
Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
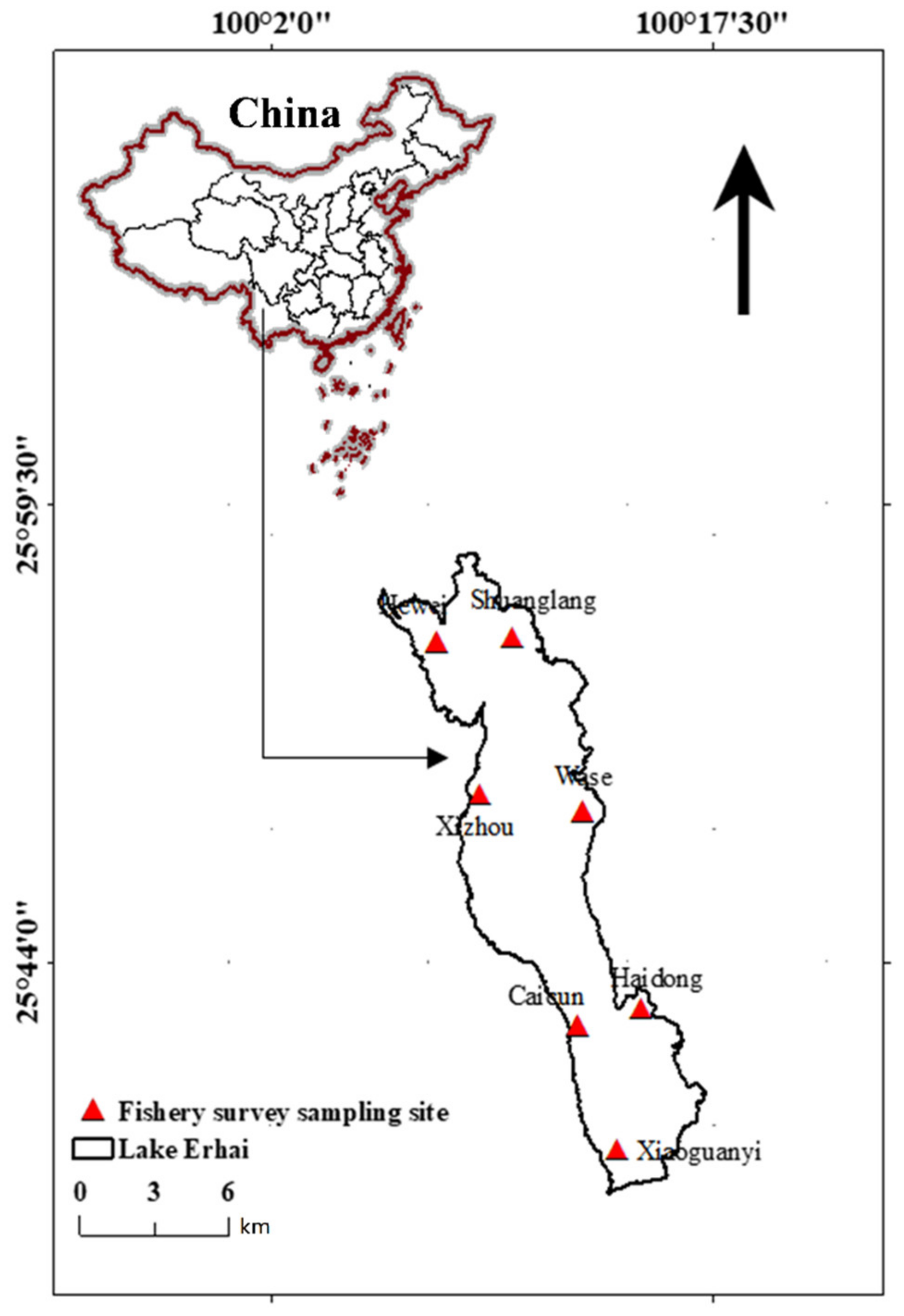
2.2. Sampling Sites
2.3. Sampling Methods
2.4. Data Analysis
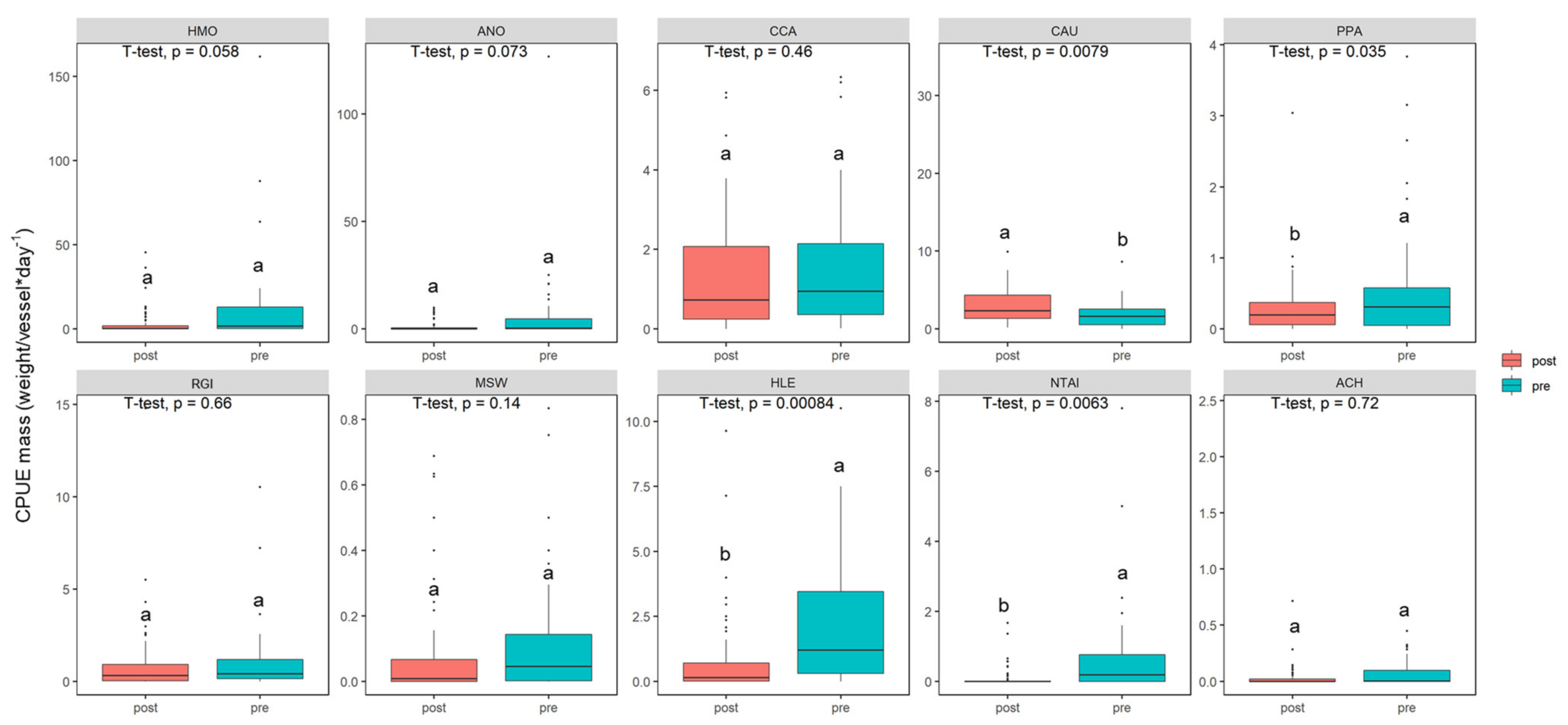
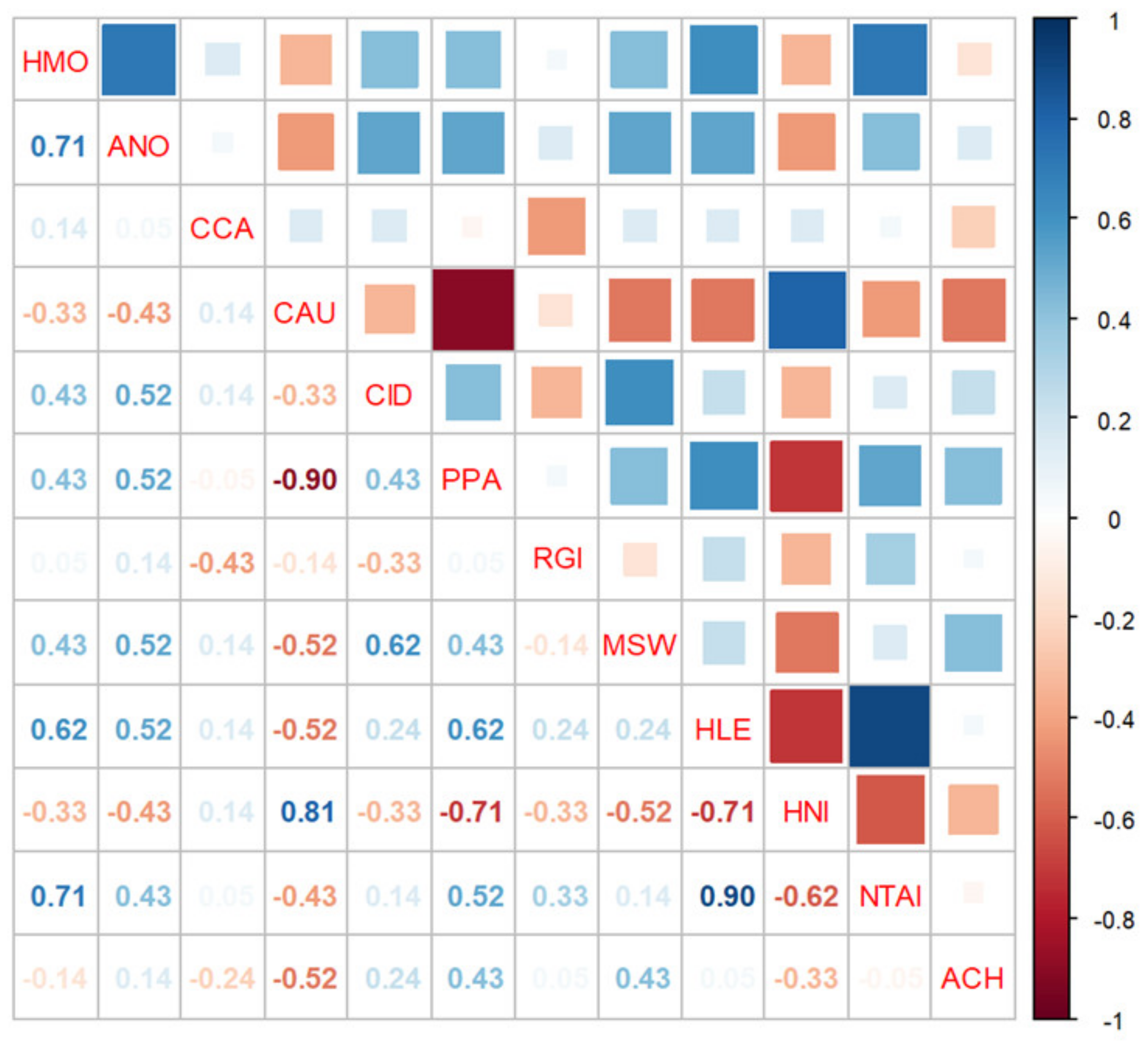
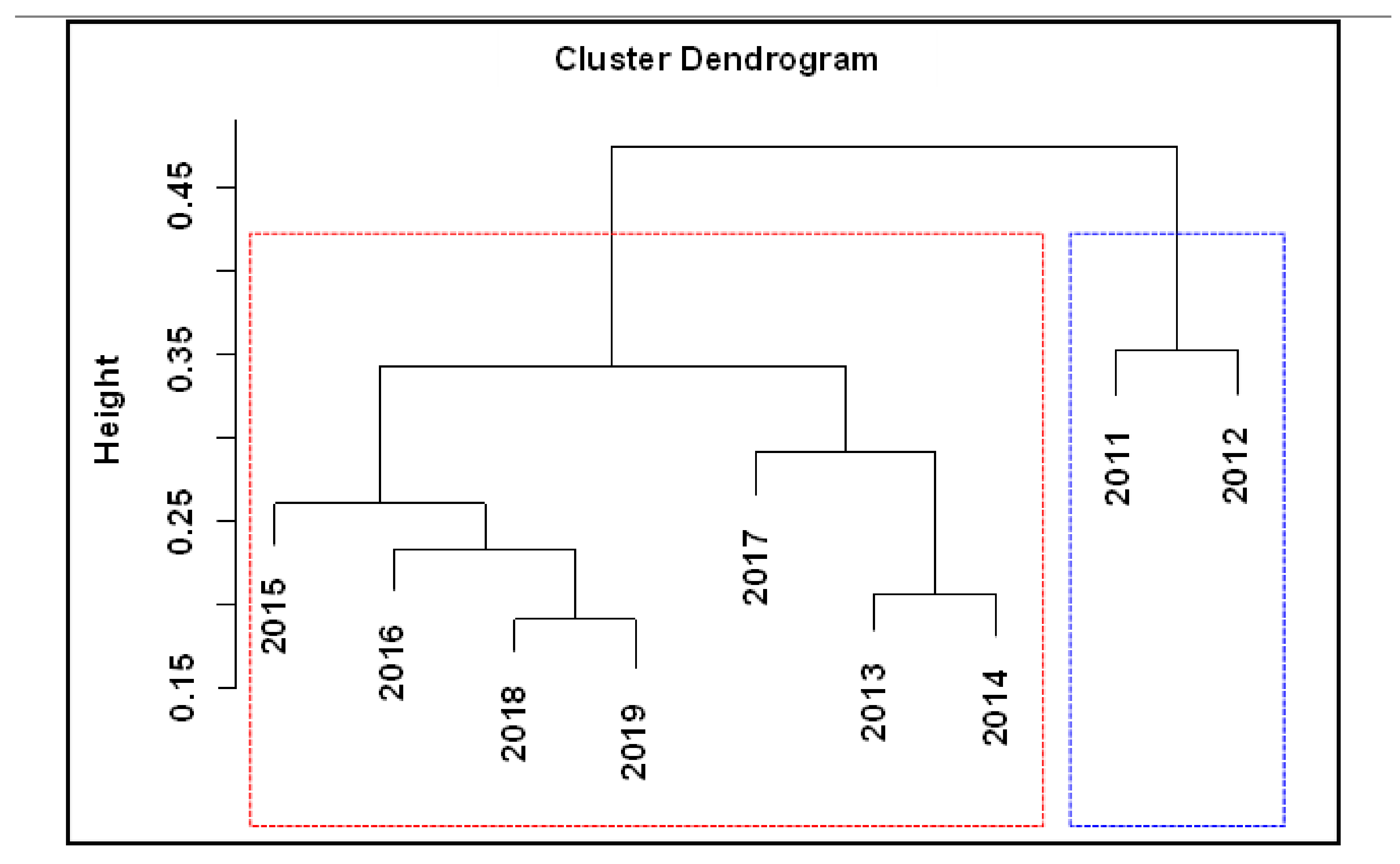
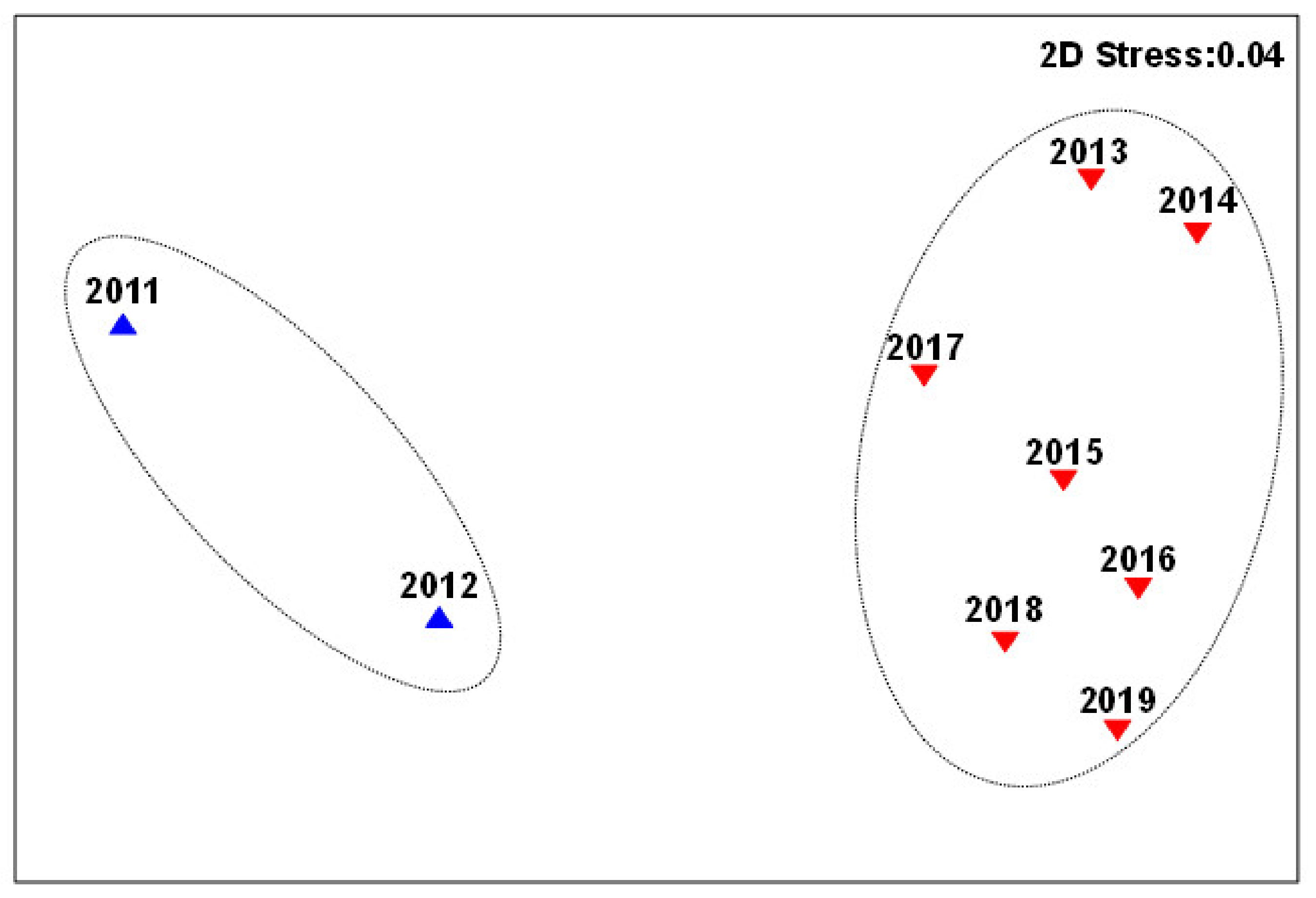
3. Results
3.1. Fish Species Composition
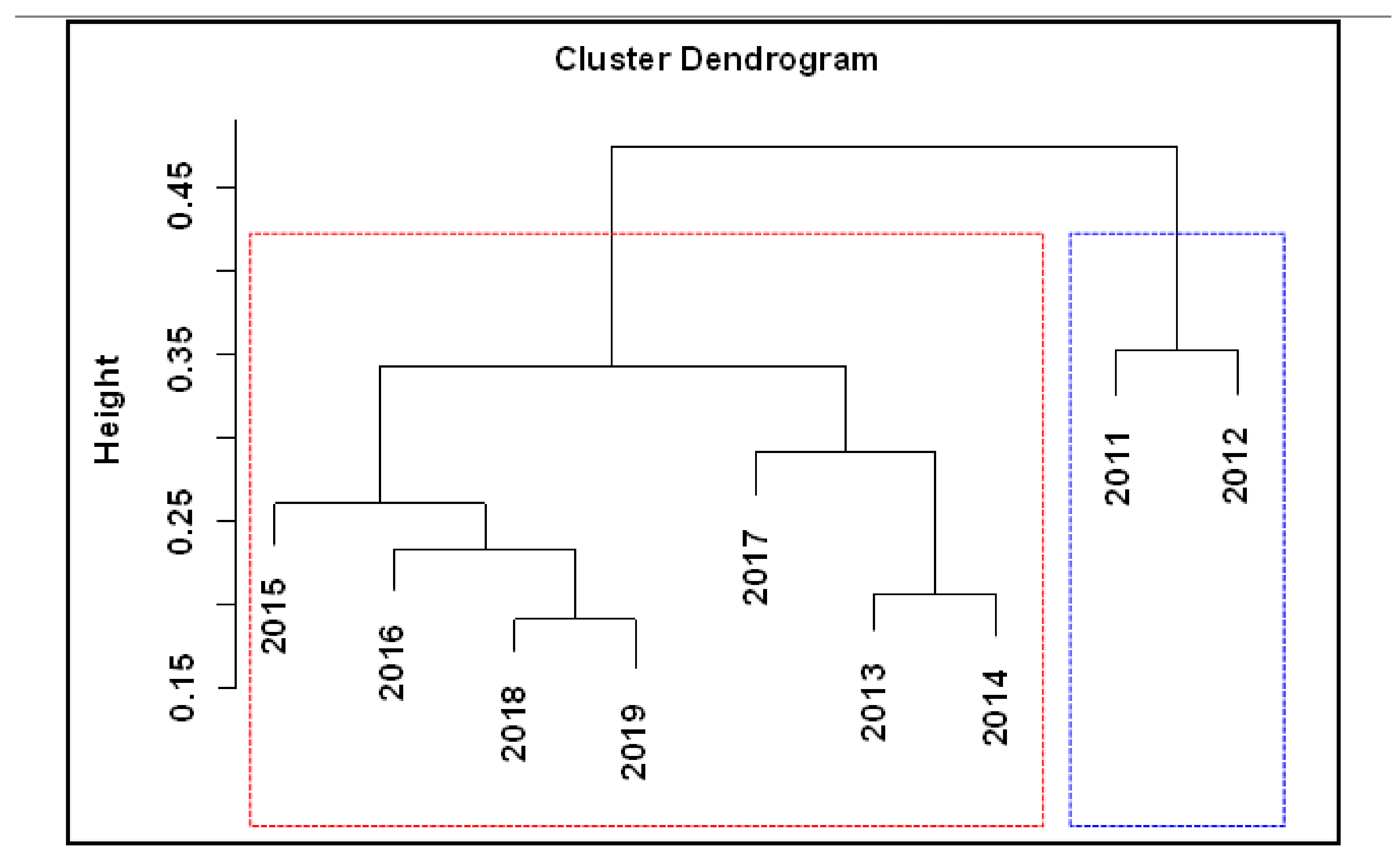
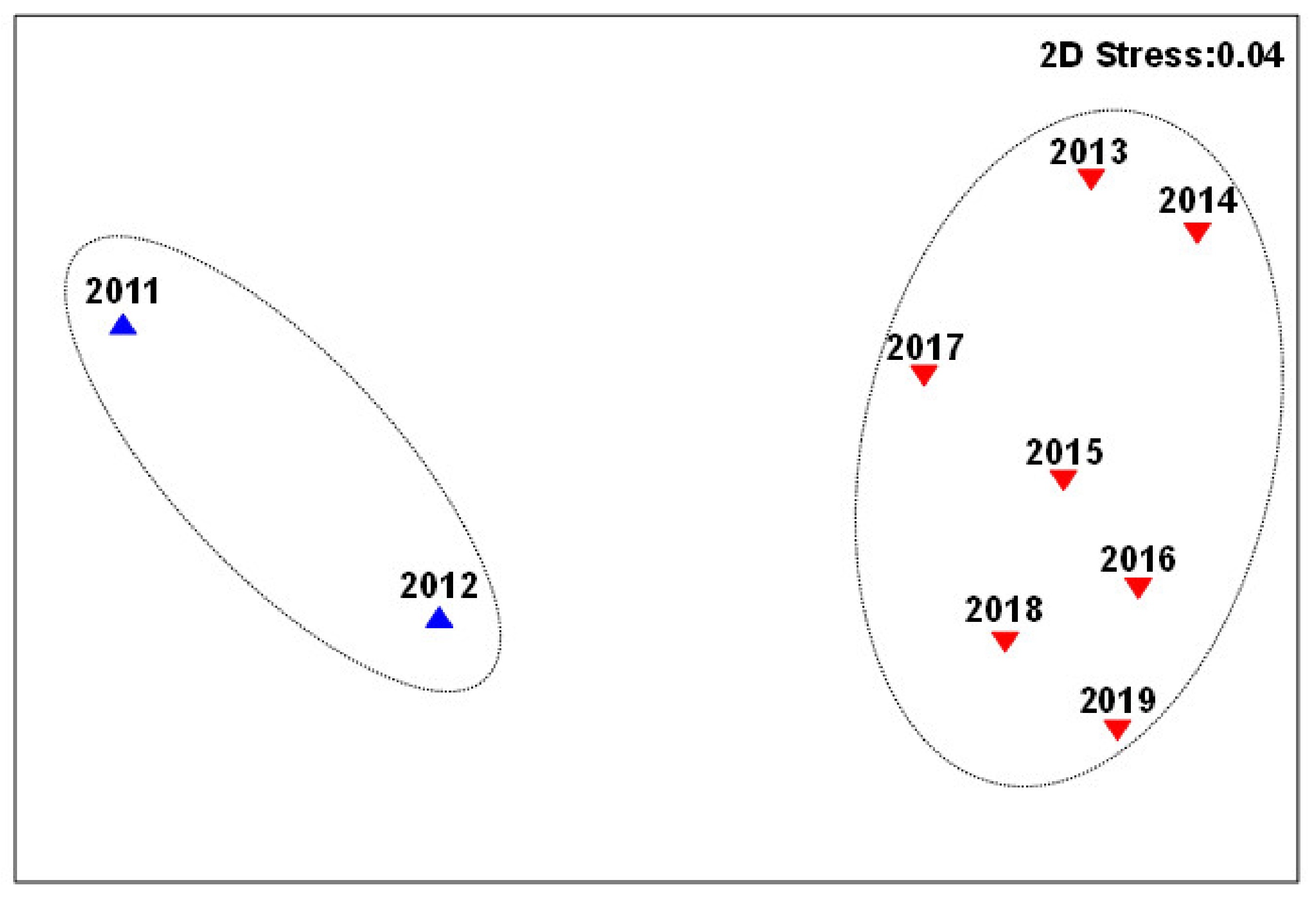
3.2. Changes in Fish Communities
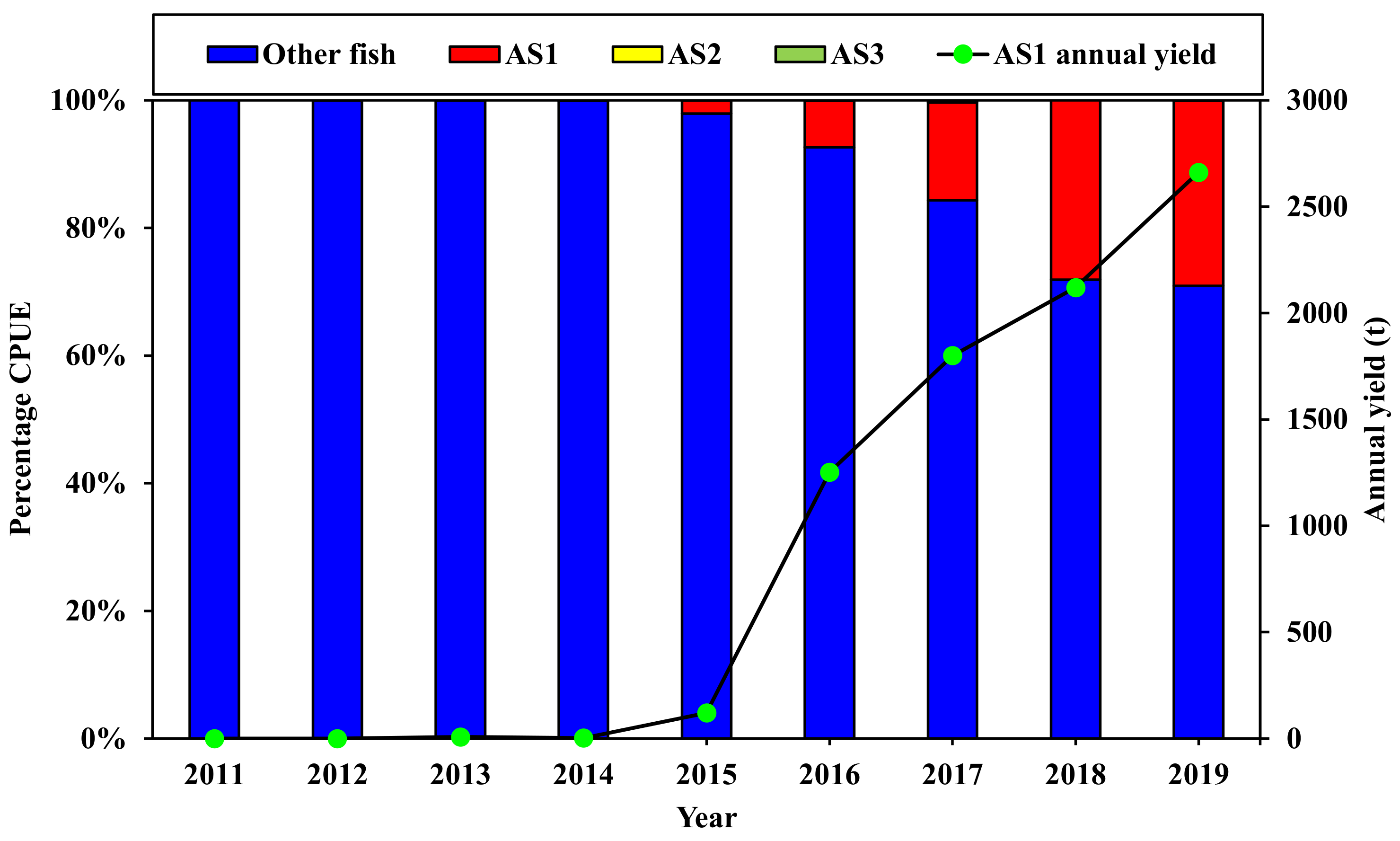
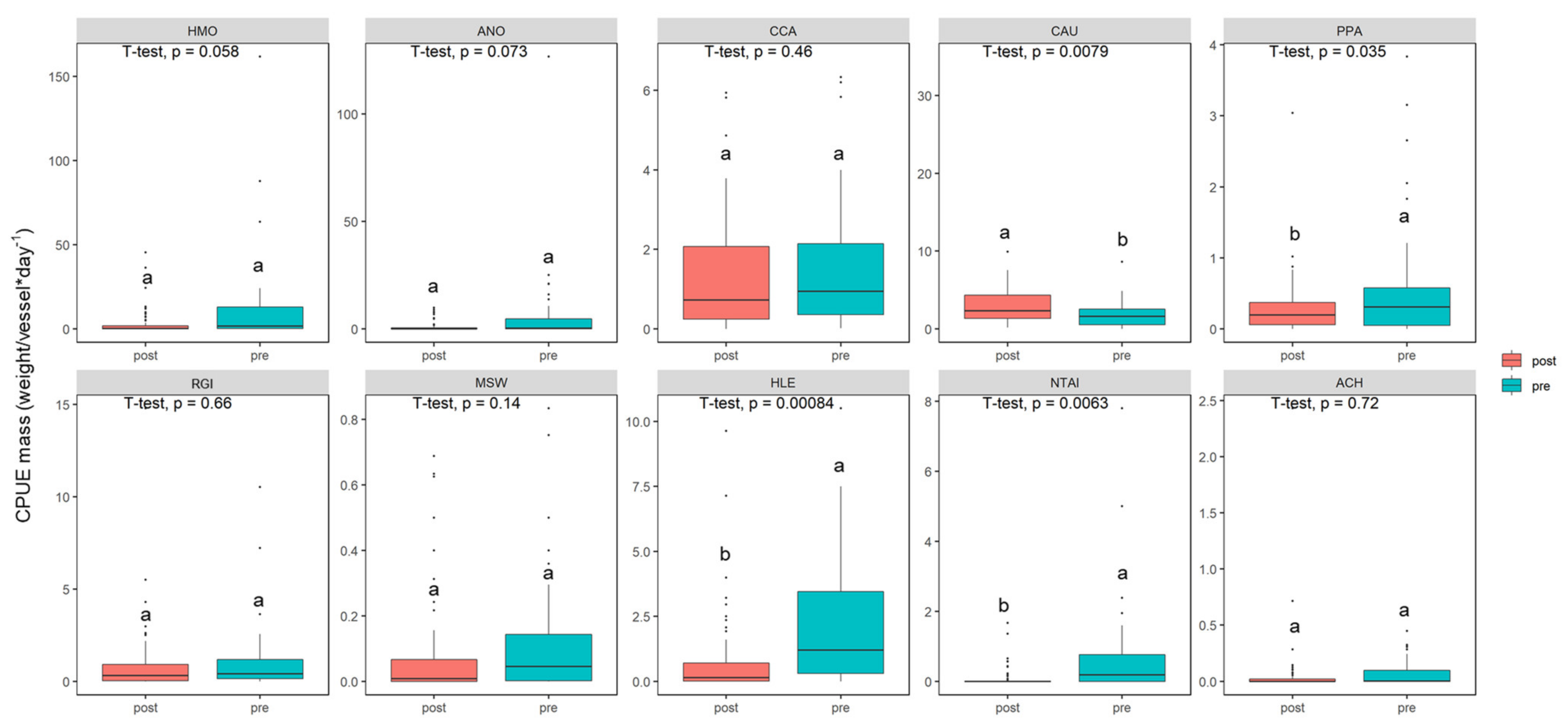
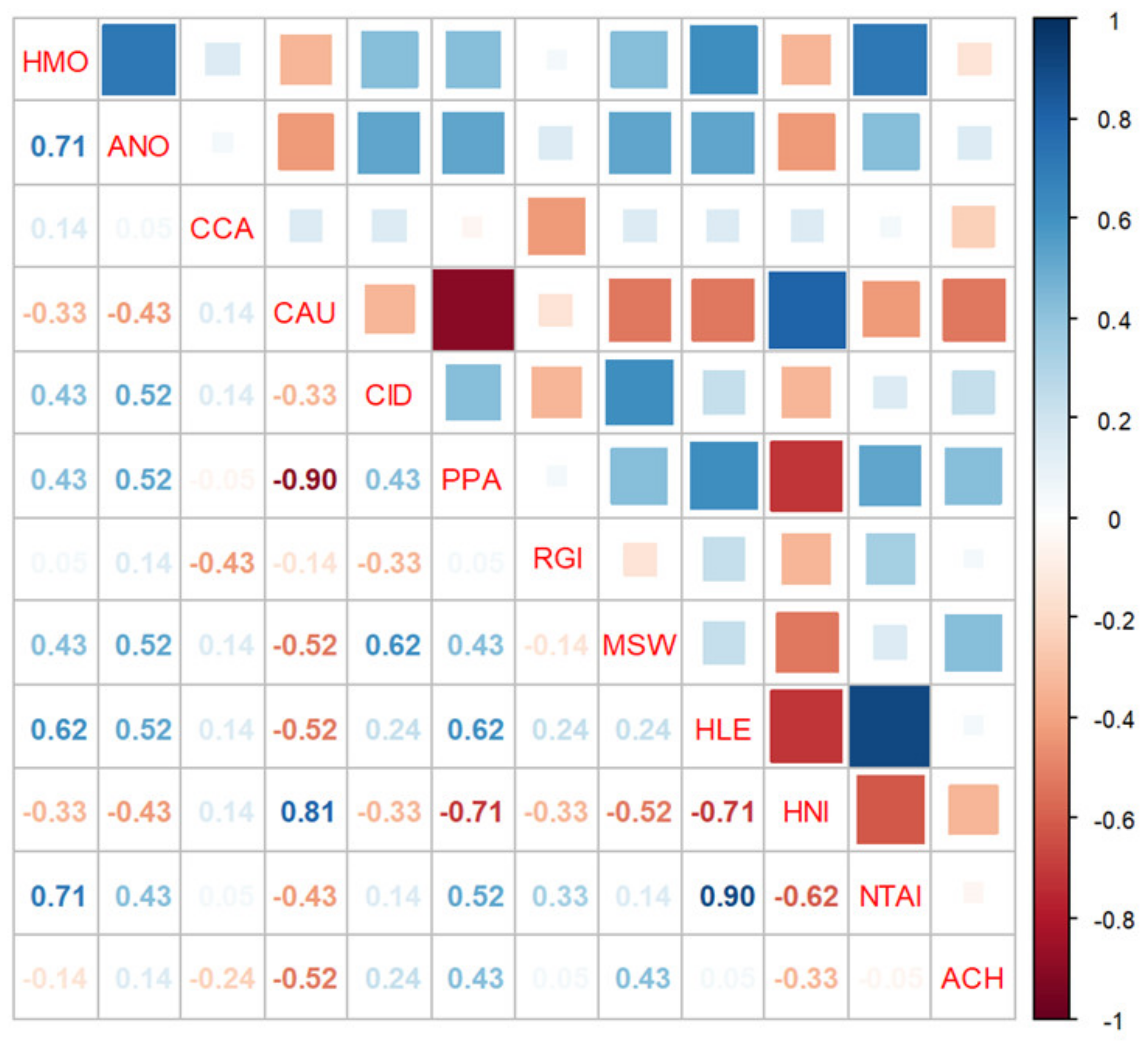
3.3. Variations in Population Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Britton, J.R.; Gozlan, R.E.; Copp, G.H. Managing non-native fish in the environment. Fish Fish. 2011, 12, 256–274. [Google Scholar] [CrossRef]
- Peeler, E.J.; Oidtmann, B.C.; Midtlyng, P.J.; Miossec, L.; Gozlan, R.E. Non-native aquatic animals introductions have driven disease emergence in Europe. Biol. Invasions 2010, 13, 1291–1303. [Google Scholar] [CrossRef] [Green Version]
- Jeschke, J.M.; Strayer, D.L. Invasion success of vertebrates in Europe and North America. Proc. Natl. Acad. Sci. USA 2005, 102, 7198–7202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricciardi, A.; Kipp, R. Predicting the number of ecologically harmful exotic species in an aquatic system. Divers. Distrib. 2007, 14, 374–380. [Google Scholar] [CrossRef]
- Cucherousset, J.; Olden, J.D. Ecological Impacts of Nonnative Freshwater Fishes. Fisheries 2011, 36, 215–230. [Google Scholar] [CrossRef]
- Gurevitch, J.; Padilla, D.K. Are invasive species a major cause of extinctions? Trends Ecol. Evol. 2004, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M. Invasive Species, Environmental Change and Management, and Health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, F.; Leunda, P.M. Non-native fish impacts on Mediterranean freshwater ecosystems: Current knowledge and research needs. Fish. Manag. Ecol. 2012, 19, 142–156. [Google Scholar] [CrossRef]
- Marr, S.M.; Ellender, B.R.; Woodford, D.J.; Alexander, M.E.; Wasserman, R.J.; Ivey, P.; Zengeya, T.; Weyl, O.L. Evaluating invasion risk for freshwater fishes in South Africa. Bothalia 2017, 47. [Google Scholar] [CrossRef]
- Ellender, B.; Weyl, O. A review of current knowledge, risk and ecological impacts associated with non-native freshwater fish introductions in South Africa. Aquat. Invasions 2014, 9, 117–132. [Google Scholar] [CrossRef]
- Matsuzaki, S.S.; Usio, N.; Takamura, N.; Washitani, I. Contrasting impacts of invasive engineers on freshwater ecosystems: An experiment and meta-analysis. Oecologia 2009, 158, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Ellis, B.K.; Stanford, J.A.; Goodman, D.; Stafford, C.P.; Gustafson, D.L.; Beauchamp, D.A.; Chess, D.W.; Craft, J.A.; Deleray, M.A.; Hansen, B.S. Long-term effects of a trophic cascade in a large lake ecosystem. Proc. Natl. Acad. Sci. USA 2011, 108, 1070–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rypel, A.L. Do invasive freshwater fish species grow better when they are invasive? Oikos 2014, 123, 279–289. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Britton, J.R.; Cowx, I.; Copp, G.H. Current knowledge on non-native freshwater fish introductions. J. Fish Biol. 2010, 76, 751–786. [Google Scholar] [CrossRef]
- Leino, J.R.; Mensinger, A.F. The benthic fish assemblage of the soft-bottom community of the Duluth-Superior Harbor before and after round goby invasion (1989–2011). J. Great Lakes Res. 2016, 42, 829–836. [Google Scholar] [CrossRef]
- Wang, S.H.; Wang, J.; Li, M.B.; Du, F.; Yang, Y.M.; Lassoie, J.P.; Hassan, M.Z. Six decades of changes in vascular hydrophyte and fish species in three plateau lakes in Yunnan, China. Biodivers. Conserv. 2013, 22, 3197–3221. [Google Scholar] [CrossRef]
- Fei, J.H. Studies on the Present Condition of Fish Stocks and Its Spatial Structure, Growth Characteristics in Erhai Lake. Master’s Thesis, Hangzhou Normal University, Hangzhou, China, 2012. [Google Scholar]
- Zhu, F.Y.; Ye, S.W.; Li, Z.J.; Zhang, T.L.; Yuan, J.; Guo, Z.Q.; Tang, J.F.; Liu, J.S. Growth and reproduction of the non-native icefish Neosalanx taihuensis Chen, 1956 (Salangidae) in a plateau lake, southwestern China. J. Appl. Ichthyol. 2014, 30, 862–869. [Google Scholar] [CrossRef]
- Du, B.H.; Li, Y.A. Danger Risk to Fish Diversity in Erhai Lake and Proposals to Dispel It. Res. Environ. Sci. 2001, 14, 42–44. [Google Scholar] [CrossRef]
- Wu, Q.L.; Wang, Y.F. On the succession of Aquatic Communities in Erhai Lake. J. Lake Sci. 1999, 11, pp. 267–273. Available online: https://www.semanticscholar.org/paper/On-the-Succession-of-Aquatic-Communities-in-Erhai-Qinglon/af6872c768d2839fc7b5355bc8144bb4ef37a153.
- Jiang, Z.G.; Brosse, S.; Jiang, X.M.; Zhang, E. Measuring ecosystem degradation through half a century of fish species introductions and extirpations in a large isolated lake. Ecol. Indic. 2015, 58, 104–112. [Google Scholar] [CrossRef]
- Lin, Q.Q.; Zhao, P.P.; Wu, Q.T.; Wei, Z.H.; Li, H.M.; Han, B.P. Predation pressure induced by seasonal fishing moratorium changes the dynamics of subtropical Cladocera populations. Hydrobiologia 2013, 710, 73–81. [Google Scholar] [CrossRef]
- Tang, J.F.; Ye, S.W.; Li, W.; Liu, J.S.; Zhang, T.L.; Guo, Z.Q.; Zhu, F.Y.; Li, Z.J. Status and historical changes in the fish community in Erhai Lake. Chin. J. Oceanol. Limnol. 2013, 31, 712–723. [Google Scholar] [CrossRef]
- Saruwatari, T.; Lopez, J.A.; Pietsch, T.W. A revision of the osmerid genus Hypomesus Gill (Teleostei: Salmoniformes), with the description of a new species from the southern Kuril Islands. Species Divers. 1997, 2, 59–82. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.H.; Hanazato, T.; Ueshima, G.; Tahara, H. Feeding habit of pond smelt (Hypomesus transpacificus nipponensis) and its impact on the zooplankton community in lake suwa, Japan. J. Freshw. Ecol. 2005, 20, 129–138. [Google Scholar] [CrossRef]
- Sakamoto, D.; Nemoto, T.; Sunoh, N.; Iwasaki, J.; Niwa, S.; Arayama, K.; Suzuki, N.; Pai, S.; Takagi, K.; Sakuramoto, K. Population size estimation of the pond smelt Hypomesus nipponensis in Lake Kasumigaura and Lake Kitaura, Japan. Fish. Sci. 2014, 80, 907–914. [Google Scholar] [CrossRef]
- Xie, Y.H.; Li, B.; Li, W.L.; Liu, C.X.; Xu, S.Q.; Zhao, X.L. The Fishes of Genus Hypomesus and Utilization of Its Resource; Liaoning Science and Technology Press: Liaoning, China, 1992; pp. 159–164. [Google Scholar]
- Katano, O.; Hakoyama, H.; Matsuzaki, S.S. Japanese inland fisheries and aquaculture: Status and trends. Freshw. Fish. Ecol. 2015, 231–240. [Google Scholar] [CrossRef]
- Swanson, C.; Reid, T.; Young, P.S.; Cech, J.J. Comparative environmental tolerances of threatened delta smelt (Hypomesus transpacificus) and introduced wakasagi (H. nipponensis) in an altered California estuary. Oecologia 2000, 123, 384–390. [Google Scholar] [CrossRef]
- Makino, W.; Mikami, H.; Katano, N.; Nakagawa, M.; Takamura, N. Biological productivity of Lake Towada, a north temperate, oligotrophic, kokanee fishery lake. Limnology 2003, 4, 79–90. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, R.M.; Cai, L.G.; Tian, Y.; Tilawalti, T.; Lai, Y. Fish Resources and Fishery in the Lake Bositenghu; The Science and Technology Press of Xinjiang: Xinjiang, China, 2005; pp. 1–160. [Google Scholar]
- Zhou, X.Y.; Hu, Z.J.; Liu, Q.G.; Yang, L.L.; Wang, Y.B. Feeding ecology of the non-indigenous fish Hypomesus nipponensis in Lake Ulungur, China: Insight into the relationship between its introduction and the collapse of the native Eurasian perch population. Mar. Freshw. Res. 2013, 64, 549–557. [Google Scholar] [CrossRef]
- Adakbek, K.; Su, D.X.; Yang, Y.; Ye, S.M.; Huderat. The current situation and the countermeasures of protection and development of fish resources in Ulungur Lake. J. Shanghai Fish. Univ. 2006, 15, 308–314. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, R.M.; Li, H. Degeneration Causes of the Aboriginal Fishes and Some Measures for Protecting Them in the Ergis River, Xinjiang, China. Arid Zone Res. 2003, 20, 152–155, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Lin, M.L. Studies on Stocking Techniques of Culter mongolicus and Ecosystem Effects of Piscivorous Fishes Releasing in Lakes along the Middle and Lower Reach of the Yangtze River. Ph.D. Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2012. (In Chinese with English Abstract). [Google Scholar]
- Ha, J.Y.; Hanazato, T.; Chang, K.H.; Jeong, K.S.; Kim, D.K. Assessment of the lake biomanipulation mediated by piscivorous rainbow trout and herbivorous daphnids using a self-organizing map: A case study in Lake Shirakaba, Japan. Ecol. Inform. 2015, 29, 182–191. [Google Scholar] [CrossRef]
- Gong, L.; Guo, L.G.; Yin, C.J.; Yang, J.J.; Yang, Y.L. Study on growth characteristics and population regulation effect of Hypomesus nipponesis and Neosalanx taihuensis in Lake Erhai. J. Hydroecology 2020, in press. [Google Scholar]
- Li, Y.; Yen, H.; Lei, Q.L.; Qiu, W.W.; Luo, J.F.; Lindsey, S.; Qin, L.H.; Zhai, L.M.; Wang, H.Y.; Wu, S.X.; et al. Impact of human activities on phosphorus flows on an early eutrophic plateau: A case study in Southwest China. Sci. Total Environ. 2020, 714, 136851. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Wang, L.; Pang, Y. Spatial-temporal variation of inflow river water quality under land use effect. J. Agro-Environ. Sci. 2020, 39, 160–170. [Google Scholar]
- Fei, J.H.; Tang, T.; Shao, X.Y. Fishery resources and developmental patterns of fishery in Erhai Lake. Wetl. Sci. 2011, 9, 277–283. [Google Scholar] [CrossRef]
- Chu, X.L.; Chen, Y.R. Fishes of Yunnan, Vol. I.; Science Press: Beijing, China, 1989. (In Chinese) [Google Scholar]
- Chu, X.L.; Chen, Y.R. Fishes of Yunnan, Vol. II.; Science Press: Beijing, China, 1990. (In Chinese) [Google Scholar]
- Knights, D.; Kuczynski, J.; Charlson, E.S.; Zaneveld, J.; Mozer, M.C.; Collman, R.G.; Bushman, F.D.; Knight, R.; Kelley, S.T. Bayesian community-wide culture-independent microbial source tracking. Nat. Methods 2011, 8, 761–763. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.; Keough, M. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Liu, G.M.; Liu, Z.W.; Li, Y.L.; Chen, F.Z.; Gu, B.H.; Smoak, J.M. Effects of fish introduction and eutrophication on the cladoceran community in Lake Fuxian, a deep oligotrophic lake in southwest China. J. Paleolimnol. 2008, 42, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Sarnelle, O.; Knapp, R.A. Nutrient recycling by fish versus zooplankton grazing as drivers of the trophic cascade in alpine lakes. Limnol. Oceanogr. 2005, 50, 2032–2042. [Google Scholar] [CrossRef]
- Gong, L. Study on Effects of Invasion of Hypomesus nipponensis on Crustacean Zooplankton in Lake Erhai. Master’s Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2020. [Google Scholar]
- Guo, N.C.; Zhang, M.; Yu, Y.; Qian, S.Q.; Li, D.M.; Kong, F.X. Crustacean zooplankton communities in 13 lakes of Yunnan-Guizhou plateau: Relationship between crustacean zooplankton biomass or size structure and trophic indicators after invasion by exotic fish. Ann. Limnol. Int. J. Limnol. 2009, 45, 279–288. [Google Scholar] [CrossRef]
- Wang, X.L.; Xiang, J.G.; Liu, J.S.; Liu, M.; Wu, L.; Murphy, B.R.; Xie, S.G. Reduced growth and reproductive investment of Hemiculter leucisculus (Cyprinidae) in a reservoir with introduced icefish Neosalanx taihuensis (Salangidae). Environ. Biol. Fishes 2013, 96, 895–903. [Google Scholar] [CrossRef]
- Hu, C.L.; Guo, L.G.; Wang, S.R. Diet and feeding ecology of invasive icefish Neosalanx taihuensis in Erhai Lake, a Chinese plateau meso-eutrophicated lake. Chin. J. Oceanol. Limnol. 2015, 33, 372–380. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Huang, D.; Zhao, Y.; Wang, H.; Hu, S.; Shen, J. Age, growth and mortality of invasive sharpbelly, Hemiculter leucisculus (Basilewski, 1855) in Erhai Lake, China. J. Appl. Ichthyol. 2013, 29, 1279–1285. [Google Scholar] [CrossRef]
- Czerniejewski, P.; Rybczyk, A.; Linowska, A.; Sobecka, E. New location, food composition, and parasitic fauna of the invasive fish Pseudorasbora parva (Temminck & Schlegel, 1846) (Cyprinidae) in Poland. Turk. J. Zool. 2019, 43, 94–105. [Google Scholar] [CrossRef]
- Yalcin-Ozdilek, S.; Kirankaya, S.G.; Ekmekci, F.G. Feeding Ecology of the Topmouth Gudgeon Pseudorasbora parva (Temminck and Schlegel, 1846) in the Gelingullu Reservoir, Turkey. Turk. J. Fish. Aquat. Sci. 2013, 13, 87–94. [Google Scholar] [CrossRef]
- Xie, S.G.; Cui, Y.; Zhang, T.L.; Li, Z. Seasonal patterns in feeding ecology of three small fishes in the Biandantang Lake, China. J. Fish Biol. 2000, 57, 867–880. [Google Scholar] [CrossRef]
- Werner, E.E.; Gilliam, J.F. The ontogenetic niche and species interactions in size structured populations. Annu. Rev. Ecol. Syst. 1984, 15, 393–425. [Google Scholar] [CrossRef]
- Yin, M.C. Feeding, growth and survival of yolk-sac larvae of silver, bighead, grass and crucian carps. Trans. Chin. Ichthyol. Soc. 1997, 6, 69–79. [Google Scholar]
- Amundsen, P.-A.; Siwertsson, A.; Primicerio, R.; Bøhn, T. Long-term responses of zooplankton to invasion by a planktivorous fish in a subarctic watercourse. Freshw. Biol. 2009, 54, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Angeler, D.G.; Álvarez-Cobelas, M.; Sánchez-Carrillo, S.; Rodrigo, M.A. Assessment of exotic fish impacts on water quality and zooplankton in a degraded semi-arid floodplain wetland. Aquat. Sci. 2002, 64, 76–86. [Google Scholar] [CrossRef]
- Bohn, T.; Amundsen, P.-A. Effects of invading vendace (Coregonus Albula L.) on species composition and body size in two zooplankton communities of the Pasvik River System, northern Norway. J. Plankton Res. 1998, 20, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.F.; Huang, X.F.; Liu, J.K.; Jiao, N.Z. Effects of fish stocking on the zooplankton community structure in a shallow lake in China. Fish. Manag. Ecol. 2005, 12, 81–89. [Google Scholar] [CrossRef]
- Tang, J.F. Studies on Fisheries Ecology in a Large Yunnan Plateau Lake, Lake Erhai, China. Ph.D. Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2013. [Google Scholar]
- Davies, G.D.; Britton, J.R. Consistency in the life history traits of four invasive Pseudorasbora parva populations in Southern England. J. Appl. Ichthyol. 2021, 37, 295–302. [Google Scholar] [CrossRef]
- Goudswaard, K.P.C.; Witte, F.; Katunzi, E.F.B. The invasion of an introduced predator, Nile perch (Lates niloticus L.) in Lake Victoria (East Africa): Chronology and causes. Environ. Biol. Fishes 2008, 81, 127–139. [Google Scholar] [CrossRef]
- Copp, G.H.; Britton, J.R.; Cucherousset, J.; García-Berthou, E.; Kirk, R.; Peeler, E.; Stakènas, S. Voracious invader or benign feline? A review of the environmental biology of European catfish (Silurus glanis) in its native and introduced ranges. Fish Fish. 2009, 10, 252–282. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation body size and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef] [PubMed]
- McCrary, J.K.; Murphy, B.R.; Stauffer, J.R.; Hendrix, S.S. Tilapia (teleostei: Cichlidae) status in Nicaraguan natural waters. Environ. Biol. Fishes 2007, 78, 107–114. [Google Scholar] [CrossRef]
- Britton, J.R.; Davies, G.D.; Brazier, M. Eradication of the invasive Pseudorasbora parva results in increased growth and production of native fishes. Ecol. Freshw. Fish 2009, 18, 8–14. [Google Scholar] [CrossRef]
- Bergman, E.; Greenberg, L.A. Competition between a planktivore, a benthivore, and a species with ontogenetic diet shifts. Ecology 1994, 75, 1233–1245. [Google Scholar] [CrossRef]
- Kornis, M.S.; Mercado-Silva, N.; Vander, Z.M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish Biol. 2014, 80, 235–285. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Water Depth Range (m) | Water Depth Gradient | Substrate Type | Aquatic Plant Richness |
|---|---|---|---|---|
| Haidong | 1–13 | Medium | Mud | Medium |
| Wase | 2–20 | Large | Silt | Medium |
| Shuanglang | 6–10 | Large | Silt | Low |
| Hewei | 2–6 | Small | Silt | Low |
| Xizhou | 2–15 | Large | Silt | High |
| Caicun | 2–5 | Small | Silt | High |
| Xiaoguanyi | 2–10 | Medium | Silt | High |
| Family | Species | Common Names | Code | Trophic Guild | Year | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2000–2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | |||||
| Cyprinidae | Carassius auratus * | Crucian carp | CAU | Omnivore | Y | 0.01 | 0.01 | 1.46 | 1.73 | 1.41 | 2.46 | 4.01 | 3.82 | 4.81 |
| Mylopharyngodon piceus ξ | Black carp | MPI | Carnivore | Y | 0.09 | 0.02 | 0.02 | 0.02 | 0.00 | 0.00 | ||||
| Ctenopharyngodon idellus ξ | Grass carp | CID | Herbivore | Y | 0.07 | 0.06 | 0.39 | 0.23 | 0.17 | 0.20 | 0.10 | 0.38 | 0.17 | |
| Hypophthalmichthys molitrix ξ | Silver carp | HMO | Phytoplanktivore | Y | 3.64 | 15.78 | 6.32 | 3.29 | 11.16 | 5.99 | 0.97 | 3.06 | 1.92 | |
| Aristichthys nobilis ξ | Bighead carp | ANO | Zooplanktivore | Y | 0.00 | 0.01 | 2.87 | 2.52 | 4.34 | 1.98 | 0.41 | 1.40 | 0.99 | |
| Cyprinus carpio ξ | Common carp | CCA | Omnivore | Y | 0.02 | 0.02 | 1.57 | 0.61 | 1.24 | 1.41 | 1.40 | 1.30 | 1.70 | |
| Cyprinus carpio chi1ia * | Chilu carp | CCHI | Omnivore | Y | 0.00 | 0.00 | 0.00 | |||||||
| Cyprinus longipectoralis * | Long-pectoral carp | CLO | Carnivore | Y | 0.00 | |||||||||
| Megalobrama amblycephala ξ | Bluntsnout bream | MAM | Herbivore | Y | 0.00 | 0.00 | 0.09 | 0.02 | 0.00 | |||||
| Pseudorasbora parva ξ | Topmouth gudgeon | PPA | Omnivore | Y | 0.16 | 0.19 | 0.37 | 0.57 | 0.32 | 0.33 | 0.40 | 0.21 | 0.17 | |
| Rhodeus ocellatus ξ | Rosy bitterling | ROC | Detritivore | Y | 0.01 | 0.01 | 0.20 | 0.21 | 0.31 | 0.00 | ||||
| Acheilognathus chankaensis ξ | Khanka spiny bitterling | ACH | Detritivore | Y | 0.01 | 0.15 | 0.21 | 0.03 | 0.00 | 0.20 | 0.07 | 0.06 | ||
| Hemiculter leucisculus ξ | Sharpbelly | HLE | Zooplanktivore | Y | 1.77 | 0.08 | 0.52 | 0.87 | 2.00 | 1.23 | 0.63 | 0.26 | 0.39 | |
| Schizothorax wangchiachii * | Duanxu schizothorax | SWA | Zooplanktivore | Y | 0.00 | 0.00 | 0.00 | 0.00 | ||||||
| Abbottina rivularis ξ | Chinese false gudgeon | ARI | Omnivore | Y | 0.00 | 0.00 | 0.01 | |||||||
| Cyprinus carpio haematopterus * | Mirror carp | CCH | Omnivore | Y | 0.07 | |||||||||
| Cobitidae | Misgurnus anguillicaudatus * | Pond loach | MAN | Omnivore | Y | 0.07 | 0.13 | 0.08 | 0.86 | 0.62 | 0.06 | 0.16 | ||
| Synbranchidae | Monopterus albus * | Ricefield eel | MAL | Carnivore | Y | 0.00 | 0.01 | 0.00 | 0.01 | |||||
| Eleotridae | Micropercops swinhonis ξ | Swinhon’s sleeper | MSW | Omnivore | Y | 0.10 | 0.15 | 0.06 | 0.10 | 0.06 | 0.10 | 0.08 | ||
| Gobiidae | Rhinogobius cliffordpopei ξ | RCL | Carnivore | Y | 1.04 | 0.10 | 0.02 | 0.19 | 0.06 | 0.13 | 0.11 | 0.03 | 0.04 | |
| Rhinogobius giurinus ξ | Barcheek goby | RGI | Carnivore | Y | 9.36 | 0.88 | 0.14 | 1.71 | 0.51 | 1.17 | 0.95 | 0.25 | 0.34 | |
| Salangidae | Neosalanx taihuensis ξ | Icefish | NTAI | Zooplanktivore | Y | 0.27 | 7.80 | 0.03 | 0.08 | 0.63 | 0.67 | 0.21 | 0.00 | 0.52 |
| Channidae | Channa argus * | Snakeheaded fish | CCR | Carnivore | Y | 0.00 | 0.02 | 0.04 | 0.02 | 0.02 | 0.08 | 0.01 | 0.01 | |
| Poeciliidae | Gambusia affinis ξ | Mosquitofish | GAF | Carnivore | Y | 0.00 | 0.00 | |||||||
| Osmeridae | Hypomesus nipponensis ξ | Japanese smelt | HNI | Zooplanktivore | 0.00 | 0.01 | 0.46 | 1.32 | 1.86 | 4.41 | 4.70 | |||
| Belontiidae | Macropodus chinensis ξ | Roundtail paradise fish | MAO | Carnivore | Y | |||||||||
| Siluridae | Clarias gariepinus ξ | Catfish | CGA | Carnivore | 0.01 | |||||||||
| Silurus meridionalis Chen ξ | Southern catfish | SMC | Carnivore | 0.01 | 0.01 | 0.04 | ||||||||
| Bagridae | Pelteobagrus fulvidraco ξ | Yellow catfish | PFU | Carnivore | Y | 0.00 | 0.00 | 0.01 | 0.00 | 0.01 | 0.00 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, C.; Chen, Y.; Guo, L.; Ni, L. Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China. Water 2021, 13, 1800. https://doi.org/10.3390/w13131800
Yin C, Chen Y, Guo L, Ni L. Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China. Water. 2021; 13(13):1800. https://doi.org/10.3390/w13131800
Chicago/Turabian StyleYin, Chengjie, Yushun Chen, Longgen Guo, and Leyi Ni. 2021. "Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China" Water 13, no. 13: 1800. https://doi.org/10.3390/w13131800
APA StyleYin, C., Chen, Y., Guo, L., & Ni, L. (2021). Fish Assemblage Shift after Japanese Smelt (Hypomesus nipponensis McAllister, 1963) Invasion in Lake Erhai, a Subtropical Plateau Lake in China. Water, 13(13), 1800. https://doi.org/10.3390/w13131800