
Late Glacial Marine Transgression and Ecosystem Response in the Landlocked Elefsis Bay (Northern Saronikos Gulf, Greece)

 ,
,  ,
, 

 ,
, 

Abstract
1. Introduction
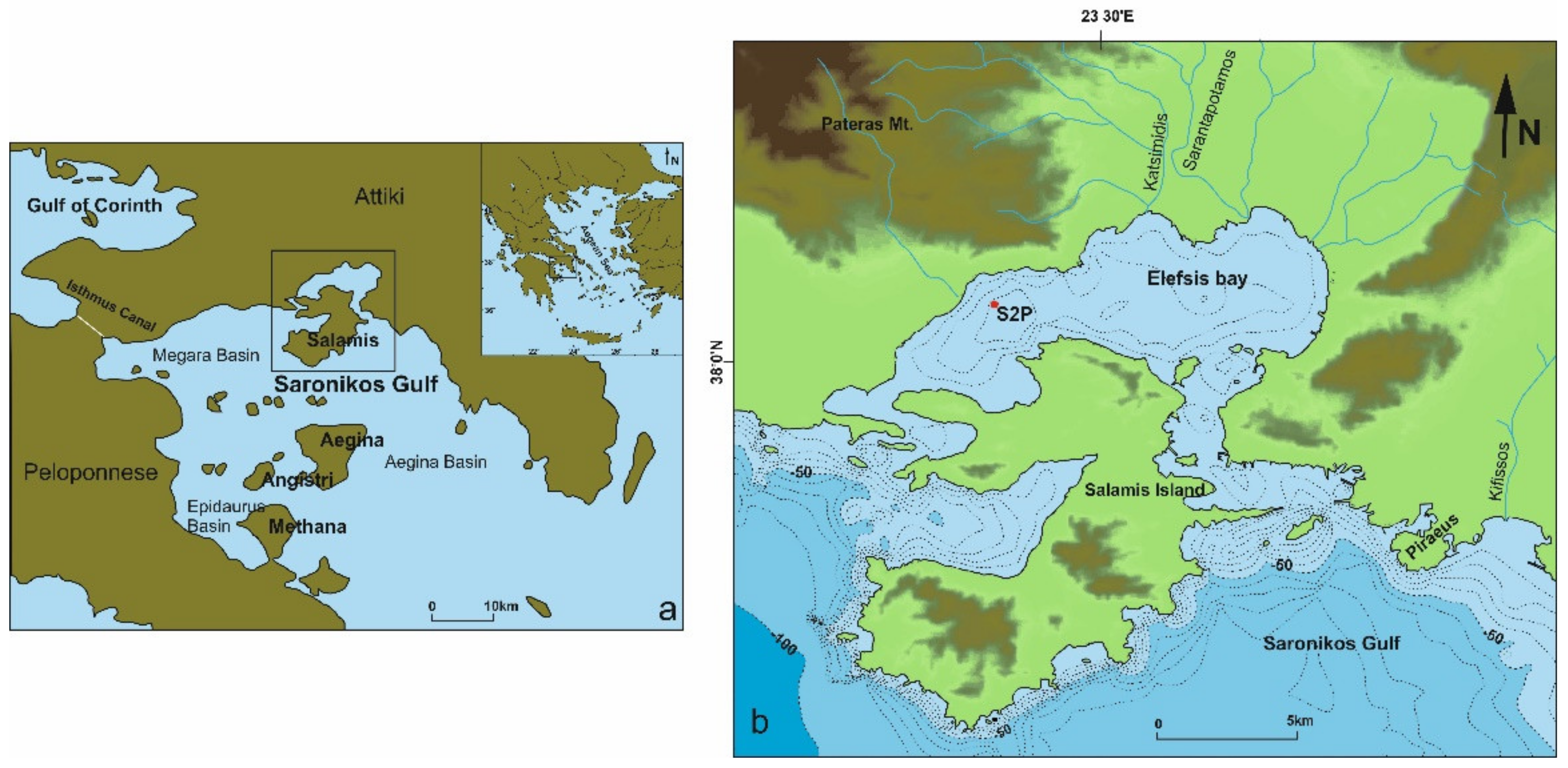
2. Regional Setting
3. Materials and Methods
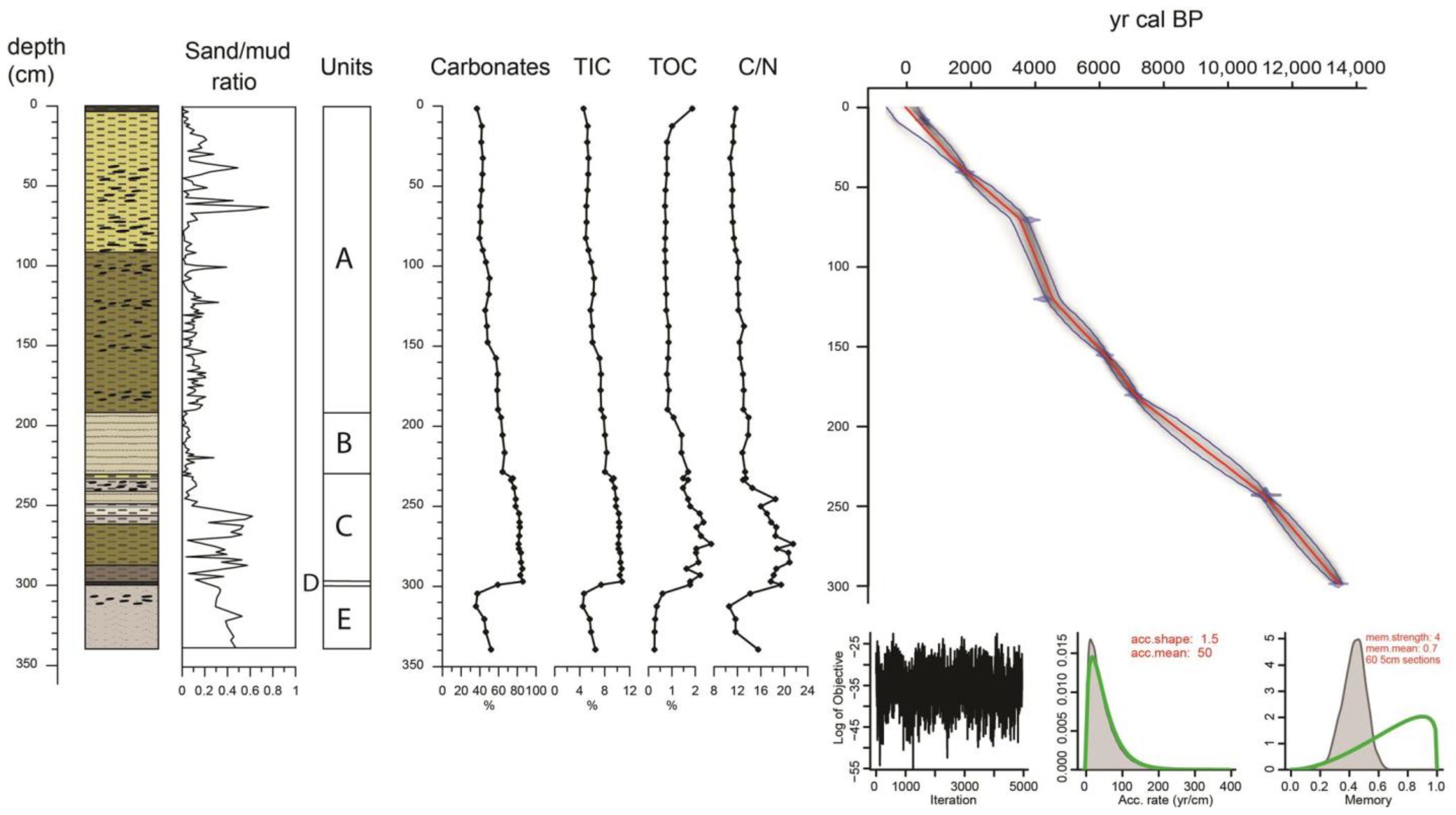
3.1. Core Chronology
3.2. Foraminifera and Ostracod Analysis
3.3. Calcareous Nannoplankton Analysis
3.4. Palynological Analysis
3.5. Mollusc Analysis
3.6. Organic Geochemistry
4. Results
4.1. Sedimentology/Age Depth Model
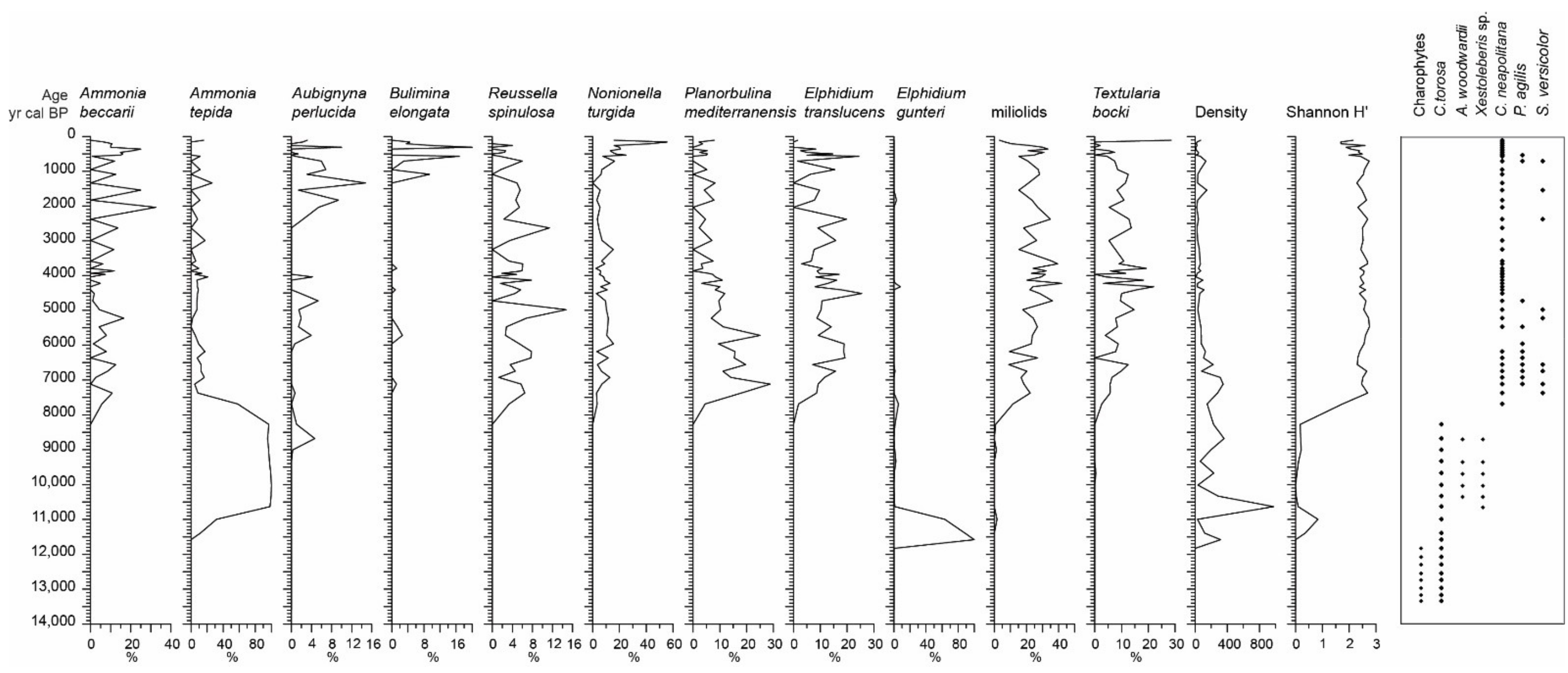
4.2. Foraminiferal and Ostracod Analysis
4.3. Calcareous Nannoplankton Analysis
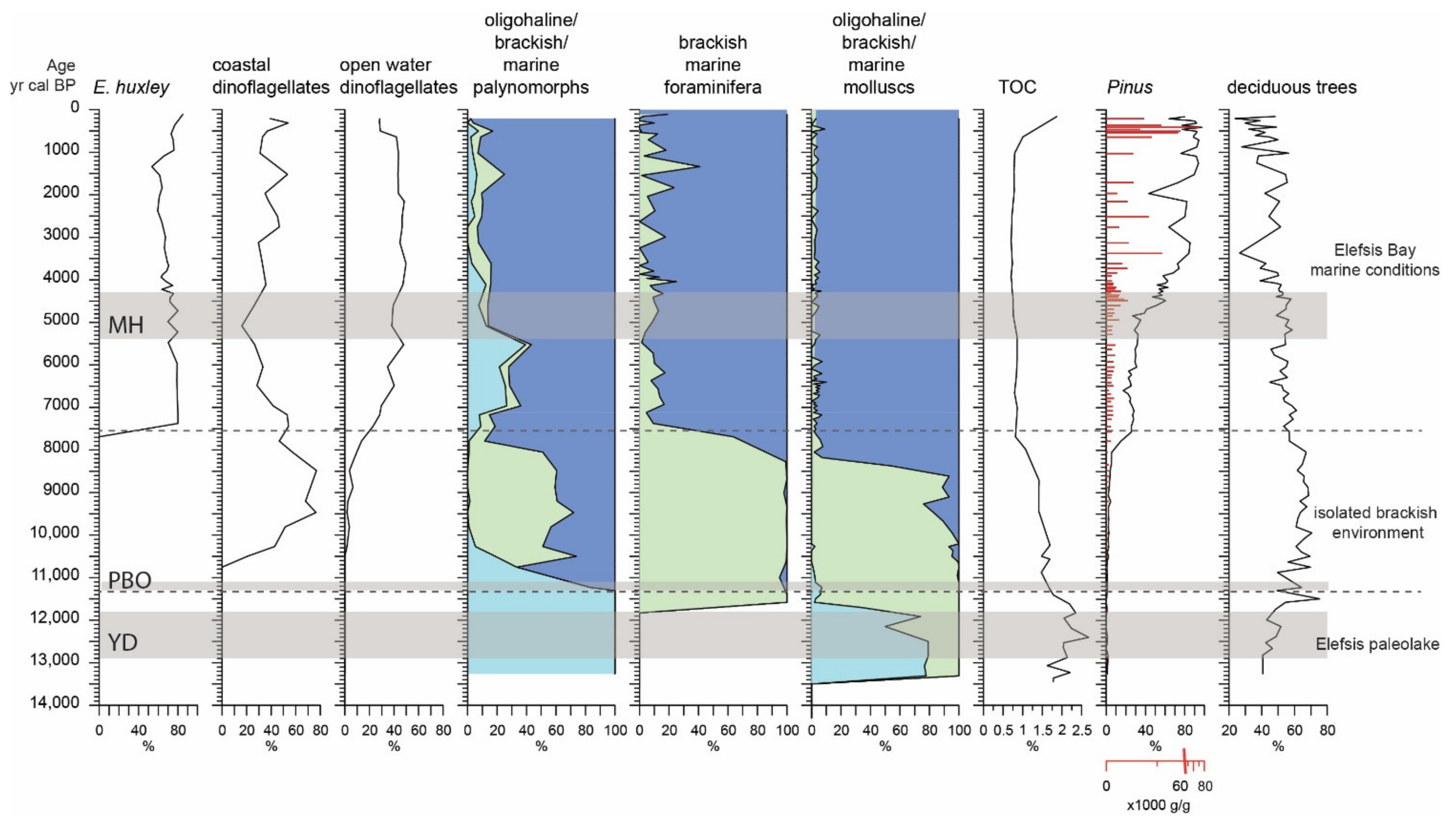
4.4. Palynological Analysis
- (a)
- PZ-I (297–248 cm; 13,500–11,350 cal BP): Botryoccocus coenobia are the dominant aquatic palynomorphs within this zone, displaying a mean concentration of 150,000 c/g (coenobia/g) and a distinct maximum concentration of 330,000 c/g. The mean concentration of aquatic vascular plants is 870 g/g, while dinoflagellate cysts are absent.
- (b)
- PZ-II (248–187 cm; 11,350–7500 cal BP): this zone is marked by a high dinoflagellate cyst concentration (15,300 c/g) and Botryoccocus decrease to 320 c/g. The morphotype of L. machaerophorum sh.p. dominates the dinoflagellate assemblage (60%), which is complemented by Spiniferites cruciformis (mean of 3%, max 5.7%) and Pyxidiniopsis psilata (max 2%). The aquatic palynomorph assemblages are complemented by the long processes morphotype of L. machaerophorum (4%) and S. bentorii (3%). Foraminifera test linings are recorded for the first time in the record with a mean concentration of 780 p/g. A decrease in the aquatic vascular plant mean concentration (310 g/g) is also observed.
- (c)
- PZ-III (187–1.5 cm; 7500–158 cal BP): the dinoflagellate cyst and Botryococcus concentrations decrease to 3000 c/g and 180 c/g, respectively, while foraminifera test linings increase to 1550 p/g. The zone is characterized by a diverse dinoflagellate assemblage, including: L. machaerophorum (mean of 14%, max 35%, min 3%), S. membranaceus (16%), S. delicatus (13%), Polysphaeridium zoharyi (9%), and Operculodinium centrocarpum (9%). The L. machaerophorum sh.p. sharply decreases to 7% at the bottom of the zone and disappears after 4600 cal BP. The abundance of Pinus, both relative and absolute, gradually increases from the bottom of the zone, reaching maximum values since 3500 cal BP. Aquatic vascular plants are present until 4000 cal BP and reoccur after 1700 cal BP, while Pseudoschizaeae spores follow a similar pattern. Finally, after 2500 cal BP, Glomus reaches maximum concentrations of 1700 p/g.
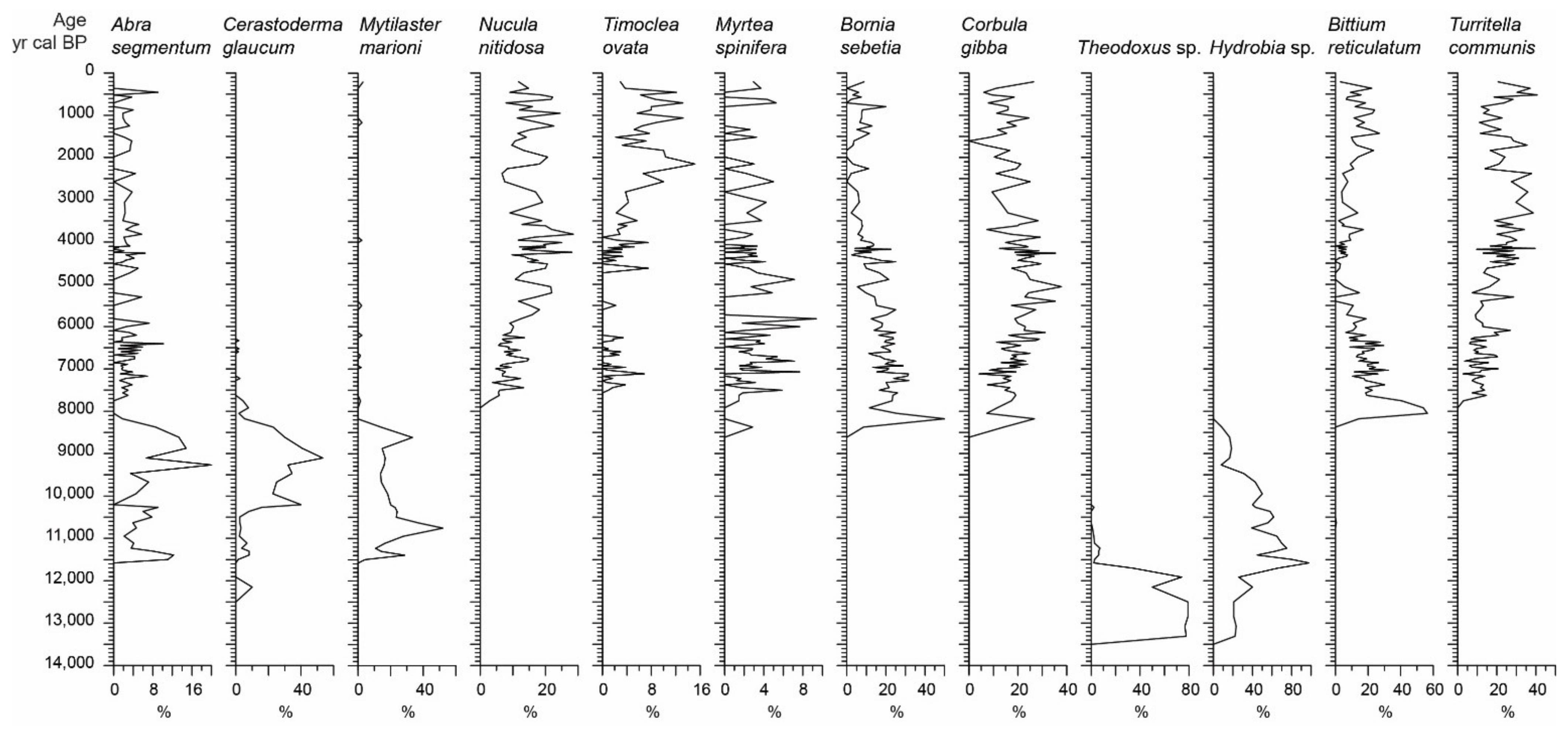
4.5. Mollusc Analysis
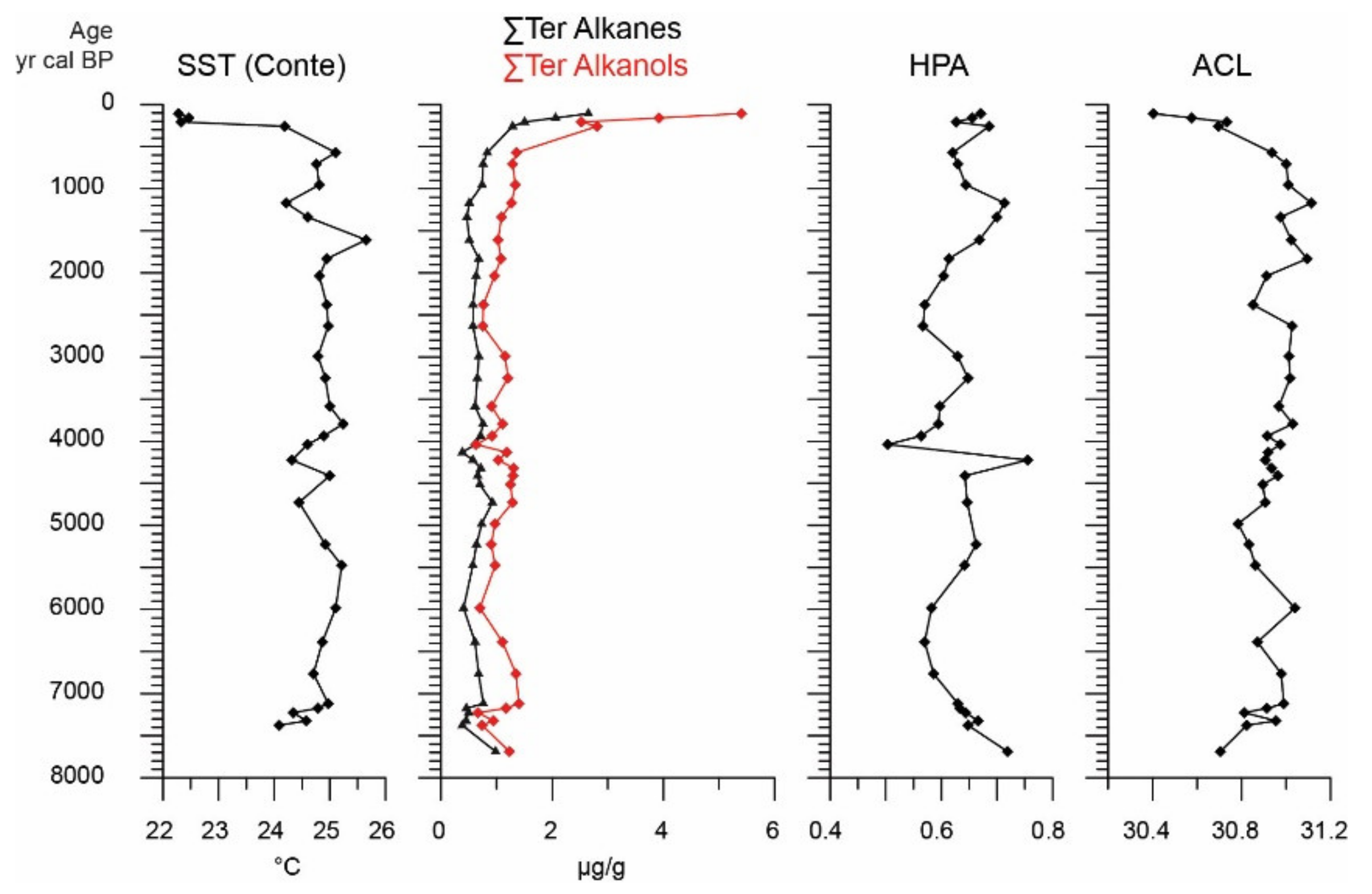
4.6. Alkenone-Based SSTs and Paleoenvironmental Indices
5. Paleoenvironmental Interpretation
5.1. Stratigraphic Interval of 300–248 cm: The Elefsis Paleolake (13,500–11,350 cal BP)
5.2. Stratigraphic Interval of 248–187 cm: The Isolated Brackish Environment (11,350–7500 cal BP)
5.3. Sratigraphic Interval of 187 cm to the Top: The Elefsis Bay (7500 cal BP to Present)
6. Conclusions
- The bottom part of the S2P sequence, most probably representing the Pliocene/Pleistocene substrate of the area, was rather deposited in a tidal system as implied by the presence of calcite layering and heavily reworked and rounded mollusc fragments. A well-defined disconformity with the rest of the sequence is recorded.
- Between 13,500 and 11,350 cal BP, a freshwater to oligohaline lacustrine environment developed, i.e., the Elefsis paleolake, characterized by green algae blooms and monospecific ostracod and gastropod oligohaline faunas.
- The occurrence of an isolated brackish environment is evidenced after 11,350 cal BP by a marked shift in the aquatic biota. Brackish dinoflagellate assemblages, mainly composed of low salinity morphotypes of L. machaerophorum, replace the green algae in the assemblages. In parallel, benthic communities are featured by high foraminiferal density values of the euryhaline species A. tepida, A. perlucida, and E. gunteri, while the prevailing ostracods C. torosa and Xestoleberis sp. together with the molluscs C. glaucum, M. marioni, and Hydrobia sp. imply an inner lagoon depositional system. As the Early Holocene relative sea level stand was not high enough to allow a direct communication of the Elefsis waterbody with the sea, the established brackish conditions were most probably the result of salinization caused by lateral encroachment from coastal waters.
- The marine conditions in Elefsis Bay are established after 7500 cal BP, evidenced by the development of a diverse dinoflagellate assemblage, gradually enriched with pelagic species, and the contemporaneous occurrence of calcareous nannoplankton assemblages, dominated by E. huxleyi. In the benthic communities the marine transgression is reflected by a sudden increase in the foraminiferal diversity and the appearance of several marine species.
- After 1500 cal BP, both benthic foraminiferal and bivalve communities are dominated by taxa tolerant in dysoxic conditions, suggesting organic matter preservation. In accordance, the contemporaneous increase of ΣTerNA and ΣTerN-OH content, high HPA values as well as the minima in the ACL profile are linked to an enhanced freshwater supply and subsequent stratification/oxygen depletion in the marine water column, implying the earliest establishment of the present-day seasonal anoxia in the Elefsis Bay.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Siddall, M.; Rohling, E.J.; Almogi-Labin, A.; Hemleben, C.; Meischner, D.; Schmelzer, I.; Smeed, D.A. Sea-level fluctuations during the last glacial cycle. Nature 2003, 423, 853–858. [Google Scholar] [CrossRef]
- Lambeck, K.; Purcell, A. Sea-level change in the Mediterranean Sea since the LGM: Model predictions for tectonically stable areas. Quat. Sci. Rev. 2005, 24, 1969–1988. [Google Scholar] [CrossRef]
- Rossi, V.; Amorosi, A.; Marchesini, M.; Marvelli, S.; Cocchianella, A.; Lorenzini, L.; Trigona, S.L.; Valle, G.; Bini, M. Late Quaternary Landscape Dynamics at the La Spezia Gulf (NW Italy): A Multi-Proxy Approach Reveals Environmental Variability within a Rocky Embayment. Water 2021, 13, 427. [Google Scholar] [CrossRef]
- Morellón, M.; Anselmetti, F.S.; Ariztegui, D.; Brushulli, B.; Sinopoli, G.; Wagner, B.; Sadori, L.; Gilli, A.; Pambuku, A. Human-climate interactions in the central Mediterranean region during the last millennia: The laminated record of Lake Butrint (Albania). Quat. Sci. Rev. 2016, 136, 134–152. [Google Scholar] [CrossRef]
- Di Rita, F.; Simone, O.; Caldara, M.; Gehrels, W.R.; Magri, D. Holocene environmental changes in the coastal Tavoliere Plain (Apulia, southern Italy): A multiproxy approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 139–151. [Google Scholar] [CrossRef]
- Miola, A.; Favaretto, S.; Sostizzo, I.; Valentini, G.; Asioli, A. Holocene salt marsh plant communities in the North Adriatic coastal plain (Italy) as reflected by pollen, non-pollen palynomorphs and plant macrofossil analyses. Veg. Hist. Archaeobot. 2010, 19, 513–529. [Google Scholar] [CrossRef]
- Alday, M.; Cearreta, A.; Cachão, M.; Freitas, M.C.; Andrade, C.; Gama, C. Micropalaeontological record of Holocene estuarine and marine stages in the Corgo do Porto rivulet (Mira River, SW Portugal). Estuar. Coast. Shelf Sci. 2006, 66, 532–543. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Pavlopoulos, K.; Tsourou, T.; Dermitzakis, M.D. Brackish marsh benthic microfauna and paleoenvironmental changes during the last 6000 years on the coastal plain of marathon (SE Greece). Riv. Ital. Paleontol. Stratigr. 2003, 109, 539–547. [Google Scholar]
- Hernández, A.; Cachão, M.; Sousa, P.; Trigo, R.M.; Luterbacher, J.; Vaquero, J.M.; Freitas, M.C. External forcing mechanisms controlling the North Atlantic coastal upwelling regime during the mid-Holocene. Geology 2020, 49, 433–437. [Google Scholar] [CrossRef]
- Orefice, M.D.; Bellotti, P.; Bertini, A.; Calderoni, G.; Neri, P.C.; Di Bella, L.; Fiorenza, D.; Foresi, L.M.; Louvari, M.A.; Rainone, L.; et al. Holocene Evolution of the Burano Paleo-Lagoon (Southern Tuscany, Italy). Water 2020, 12, 1007. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Kapsimalis, V.; Theodorakopoulou, K. Relative sea-level changes in Aegean coastal areas during Holocene: A geoarchaeological view. J. Earth Sci. 2010, 21, 244–246. [Google Scholar]
- Avramidis, P.; Iliopoulos, G.; Nikolaou, K.; Kontopoulos, N.; Koutsodendris, A.; van Wijngaarden, G.J. Holocene sedimentology and coastal geomorphology of Zakynthos Island, Ionian Sea: A history of a divided Mediterranean island. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 340–354. [Google Scholar] [CrossRef]
- Vouvalidis, K.; Syrides, G.; Pavlopoulos, K.; Pechlivanidou, S.; Tsourlos, P.; Papakonstantinou, M.-F. Palaeogeographical reconstruction of the battle terrain in Ancient Thermopylae, Greece. Geodin. Acta 2010, 23, 241–253. [Google Scholar] [CrossRef]
- Goiran, J.-P.; Pavlopoulos, K.P.; Fouache, E.; Triantaphyllou, M.V.; Etienne, R. Piraeus, the ancient island of Athens: Evidence from Holocene sediments and historical archives. Geology 2011, 39, 531–534. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Theodorakopoulou, K.; Bassiakos, Y.; Hayden, B.; Tsourou, T.; Triantaphyllou, M.; Kouli, K.; Vandarakis, D. Paleoenvironmental evolution of Istron (N.E Crete), during the last 6000 years: Depositional environment, climate and sea level changes. Geodin. Acta 2007, 20, 219–229. [Google Scholar] [CrossRef][Green Version]
- Pavlopoulos, K.; Kapsimalis, V.; Theodorakopoulou, K.; Panagiotopoulos, I.P. Vertical displacement trends in the Aegean coastal zone (NE Mediterranean) during the Holocene assessed by geo-archaeological data. Holocene 2012, 22, 717–728. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Karkanas, P.; Triantaphyllou, M.; Karymbalis, E.; Tsourou, T.; Palyvos, N. Paleoenvironmental evolution of the coastal plain of Marathon, Greece, during the Late Holocene: Depositional environment, climate, and sea level changes. J. Coast. Res. 2006, 22, 424–438. [Google Scholar] [CrossRef]
- Kontopoulos, N.; Avramidis, P. A late Holocene record of environmental changes from the Aliki lagoon, Egion, North Peloponnesus, Greece. Quat. Int. 2003, 111, 75–90. [Google Scholar] [CrossRef]
- Vött, A.; Schriever, A.; Handl, M.; Brückner, H. Holocene Palaeogeographies of the Eastern Acheloos River Delta and the Lagoon of Etoliko (NW Greece). J. Coast. Res. 2007, 234, 1042–1066. [Google Scholar] [CrossRef]
- Evelpidou, N.; Pavlopoulos, K.; Vassilopoulos, A.; Triantaphyllou, M.V.; Vouvalidis, K.; Syrides, G. Yria (western Naxos island, Greece): Sea level changes in upper holocene and palaeogeographical reconstruction. Geodin. Acta 2010, 23, 233–240. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Triantaphyllou, M.V.; Karkanas, P.; Kouli, K.; Syrides, G.; Vouvalidis, K.; Palyvos, N.; Tsourou, T. Paleoenvironmental evolution and prehistoric human environment, in the embayment of Palamari (Skyros Island, Greece) during Middle-Late Holocene. Quat. Int. 2010, 216, 41–53. [Google Scholar] [CrossRef]
- Koukousioura, O.; Triantaphyllou, M.V.; Dimiza, M.D.; Pavlopoulos, K.; Syrides, G.; Vouvalidis, K. Benthic foraminiferal evidence and paleoenvironmental evolution of Holocene coastal plains in the Aegean Sea (Greece). Quat. Int. 2012, 261, 105–117. [Google Scholar] [CrossRef]
- Syrides, G. Marine mollusk fauna and Holocene stratigraphy of the marsh of Agia Paraskevi, (Lamia, Fthiotida) Greece. Bull. Geol. Soc. Greece 2008, 42, 1–14. [Google Scholar]
- Triantaphyllou, M.V.; Ziveri, P.; Gogou, A.; Marino, G.; Lykousis, V.; Bouloubassi, I.; Emeis, K.-C.C.; Kouli, K.; Dimiza, M.; Rosell-Melé, A.; et al. Late Glacial-Holocene climate variability at the south-eastern margin of the Aegean Sea. Mar. Geol. 2009, 266, 182–197. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V. Coccolithophore assemblages during the Holocene Climatic Optimum in the NE Mediterranean (Aegean and northern Levantine Seas, Greece): Paleoceanographic and paleoclimatic implications. Quat. Int. 2014, 345, 56–67. [Google Scholar] [CrossRef]
- Marret, F.; Bradley, L.R.; Tarasov, P.E.; Ivanova, E.V.; Zenina, M.A.; Murdmaa, I.O. The Holocene history of the NE Black Sea and surrounding areas: An integrated record of marine and terrestrial palaeoenvironmental change. Holocene 2019, 29, 648–661. [Google Scholar] [CrossRef]
- Kouli, K.; Brinkhuis, H.; Dale, B. Spiniferites cruciformis: A fresh water dinoflagellate cyst? Rev. Palaeobot. Palynol. 2001, 113, 273–286. [Google Scholar] [CrossRef]
- Aksu, A.E.; Yaşar, D.; Mudie, P.J.; Gillespie, H. Late glacial-Holocene paleoclimatic and paleoceanographic evolution of the Aegean Sea: Micropaleontological and stable isotopic evidence. Mar. Micropaleontol. 1995, 25, 1–28. [Google Scholar] [CrossRef]
- Mudie, P.J.; Aksu, A.E.; Yasar, D. Late Quaternary dinofagellate cysts from the Black, Marmara and Aegean seas: Variations in assemblages, morphology and paleosalinity. Mar. Micropaleontol. 2001, 43, 155–178. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Fouache, E.; Sidiropoulou, M.; Triantaphyllou, M.; Vouvalidis, K.; Syrides, G.; Gonnet, A.; Greco, E. Palaeoenvironmental evolution and sea-level changes in the coastal area of NE Lemnos Island (Greece) during the Holocene. Quat. Int. 2013, 308–309, 80–88. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Kouli, K.; Tsourou, T.; Koukousioura, O.; Pavlopoulos, K.; Dermitzakis, M.D.D. Paleoenvironmental changes since 3000 BC in the coastal marsh of Vravron (Attica, SE Greece). Quat. Int. 2010, 216, 14–22. [Google Scholar] [CrossRef]
- Harff, J.; Flemming, N.C.; Groh, A.; Hünicke, B.; Lericolais, G.; Meschede, M.; Rosentau, A.; Sakellarioü, D.; Uscinowicz, S.; Zhang, W.; et al. Sea Level and Climate. In Submerged Landscapes of the European Continental Shelf: Quaternary Paleoenvironments; Flemming, N.C., Harff, J., Moura, D., Burgess, A., Bailey, G.N., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 11–18. ISBN 9781118927823. [Google Scholar]
- Galanidou, N.; Dellaporta, K.; Sakellariou, D. Greece: Unstable Landscapes and Underwater Archaeology. In The Archaeology of Europe’s Drowned Landscapes; Springer: Cham, Switzerland, 2020; pp. 371–392. ISBN 9783030373672. [Google Scholar]
- Lambeck, K.; Rouby, H.; Purcell, A.; Sun, Y.; Sambridge, M. Sea level and global ice volumes from the Last Glacial Maximum to the Holocene. Proc. Natl. Acad. Sci. USA 2014, 111, 15296–15303. [Google Scholar] [CrossRef]
- Van Andel, T.H.; Zangger, E.; Perissoratis, C. Transgressive/Regressive Cycles in the Gulf of Argos, Greece. Quat. Res. 1990, 34, 317–329. [Google Scholar] [CrossRef]
- Perissoratis, C.; Conispoliatis, N. The impacts of sea-level changes during latest Pleistocene and Holocene times on the morphology of the Ionian and Aegean seas (SE Alpine Europe). Mar. Geol. 2003, 196, 145–156. [Google Scholar] [CrossRef]
- Karageorgis, A.P.; Kanellopoulos, T.D.; Mavromatis, V.; Anagnostou, C.L.; Koutsopoulou, E.; Schmidt, M.; Pavlopoulos, K.; Tripsanas, E.K.; Hallberg, R.O. Authigenic carbonate mineral formation in the Pagassitikos palaeolake during the latest Pleistocene, central Greece. Geo-Mar. Lett. 2013, 33, 13–29. [Google Scholar] [CrossRef]
- McNeill, L.C.; Shillington, D.J.; Carter, G.D.O.; Everest, J.D.; Gawthorpe, R.L.; Miller, C.; Phillips, M.P.; Collier, R.E.L.; Cvetkoska, A.; De Gelder, G.; et al. High-resolution record reveals climate-driven environmental and sedimentary changes in an active rift. Sci. Rep. 2019, 9, 3116. [Google Scholar] [CrossRef]
- Kapsimalis, V.; Pavlakis, P.; Poulos, S.E.; Alexandri, S.; Tziavos, C.; Sioulas, A.; Filippas, D.; Lykousis, V. Internal structure and evolution of the Late Quaternary sequence in a shallow embayment: The Amvrakikos Gulf, NW Greece. Mar. Geol. 2005, 222–223, 399–418. [Google Scholar] [CrossRef]
- Poulos, S.E.; Collins, M.B.; Lykousis, V. Late Quaternary Evolution of Amvrakikos Gulf, Western Greece. Geo-Mar. Lett. 1995, 15, 9–16. [Google Scholar] [CrossRef]
- Perissoratis, C.; Piper, D.J.W.; Lykousis, V. Alternating marine and lacustrine sedimentation during late Quaternary in the Gulf of Corinth rift basin, central Greece. Mar. Geol. 2000, 167, 391–411. [Google Scholar] [CrossRef]
- Petropoulos, A.; Androni, A.; Ntamkarelou, T.; Anagnostou, C. Carbonate and organic carbon content in the recent sediments of Elefsis bay as indicators for the paleoenviromental evolution of the system. Bull. Geol. Soc. Greece 2013, 47, 1562–1571. [Google Scholar] [CrossRef]
- Foutrakis, P.M.; Anastasakis, G.; Piper, D.J.W. Chronology of Quaternary shoreline progradational sequences related to eustatic sea-level changes: Sedimentation and subsidence in Saronikos Gulf, Greece. Mar. Geol. 2020, 428, 106278. [Google Scholar] [CrossRef]
- EC Directive of the European Parliament and of the Council 2000/60/EC establishing a framework for community action in the field of Water Policy. J. Eur. Communities Bruss. 2000, 327, 1–72.
- Kersten, M.; Garbe-Schönberg, C.D.; Thomsen, S.; Anagnostou, C.; Sioulas, A. Source apportionment of Pb pollution in the coastal waters of Elefsis Bay, Greece. Environ. Sci. Technol. 1997, 31, 1295–1301. [Google Scholar] [CrossRef]
- Karageorgis, A.P.; Botsou, F.; Kaberi, H.; Iliakis, S. Geochemistry of major and trace elements in surface sediments of the Saronikos Gulf (Greece): Assessment of contamination between 1999 and 2018. Sci. Total Environ. 2020, 717, 137046. [Google Scholar] [CrossRef]
- Pavlidou, A.; Simboura, N.; Pagou, Κ.; Assimakopoulou, G.; Gerakaris, V.; Hatzianestis, I.; Panayotidis, P.; Pantazi, M.; Papadopoulou, N.; Reizopoulou, S.; et al. Using a holistic ecosystem-integrated approach to assess the environmental status of Saronikos Gulf, Eastern Mediterranean. Ecol. Indic. 2019, 96, 336–350. [Google Scholar] [CrossRef]
- Kapetanaki, N.; Krasakopoulou, E.; Stathopoulou, E.; Dassenakis, M.; Scoullos, M. Severe coastal hypoxia interchange with ocean acidification: An experimental perturbation study on carbon and nutrient biogeochemistry. J. Mar. Sci. Eng. 2020, 8, 462. [Google Scholar] [CrossRef]
- Primpas, I.; Tsirtsis, G.; Karydis, M.; Kokkoris, G.D. Principal component analysis: Development of a multivariate index for assessing eutrophication according to the European water framework directive. Ecol. Indic. 2010, 10, 178–183. [Google Scholar] [CrossRef]
- Pavlidou, A.; Kontoyiannis, H.; Anagnostou, C.; Siokou–Frangou, I.; Pagou, K.; Krasakopoulou, E.; Assimakopoulou, G.; Zervoudaki, S.; Zeri, C.; Chatzianestis, J.; et al. Biogeochemical Characteristics in the Elefsis Bay (Aegean Sea, Eastern Mediterranean) in Relation to Anoxia and Climate Changes BT—Chemical Structure of Pelagic Redox Interfaces: Observation and Modeling; Yakushev, E.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 161–201. ISBN 978-3-642-32125-2. [Google Scholar]
- Dimiza, M.D.; Triantaphyllou, M.V.; Koukousioura, O.; Hallock, P.; Simboura, N.; Karageorgis, A.P.; Papathanasiou, E. The Foram Stress Index: A new tool for environmental assessment of soft-bottom environments using benthic foraminifera. A case study from the Saronikos Gulf, Greece, Eastern Mediterranean. Ecol. Indic. 2016, 60, 611–621. [Google Scholar] [CrossRef]
- Simboura, N.; Zenetos, A.; Pancucci-Papadopoulou, M.A. Benthic community indicators over a long period of monitoring (2000–2012) of the Saronikos Gulf, Greece, Eastern Mediterranean. Environ. Monit. Assess. 2014, 186, 3809–3821. [Google Scholar] [CrossRef] [PubMed]
- SoHelME State of the Marine Environment Report; Papathanassiou, E., Zenetos, A., Eds.; HCMR Publisher: Athens, Greece, 2005; ISBN 9608665183. [Google Scholar]
- Foutrakis, P.M.; Anastasakis, G. Quaternary continental shelf basins of Saronikos Gulf, Aegean Sea. Geo-Mar. Lett. 2020, 40, 629–647. [Google Scholar] [CrossRef]
- Makris, J.; Papoulia, J.; Drakatos, G. Tectonic deformation and microseismicity of the Saronikos Gulf, Greece. Bull. Seismol. Soc. Am. 2004, 94, 920–929. [Google Scholar] [CrossRef]
- Pantazidou, M.; Kapniaris, S.; Katsiri, A.; Christidis, A. Pollutant trends and hazard ranking in Elefsis Bay, Greece. Desalination 2007, 210, 69–82. [Google Scholar] [CrossRef]
- Alexouli-Livaditi, A.; Livaditis, G.; Sachpazis, C. Geomorphological investigation of the drainage network and calculation of the peak storm runoff (Qp) and sediment yield of Sarantapotamos and Katsimidi streams, Attica, Greece. In Proceedings of the Engineering Geology and the Environment; Marinos, P.G., Koukis, G.C., Tsiambaos, G.C., Stournaras, G.C., Eds.; Balkema: Rotterdam, The Netherlands, 1997; pp. 31–40. [Google Scholar]
- Lekkas, E. The Athens earthquake (7 September 1999): Intensity distribution and controlling factors. Eng. Geol. 2001, 59, 297–311. [Google Scholar] [CrossRef]
- Friligos, N. Nutrient and oxygen redistribution during destratification in the Elefsis Bay, an anoxic basin. Hydrobiologia 1983, 101, 223–230. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Papathanassiou, E.; Lepretre, A.; Frontier, S. Zooplankton assemblages and influence of environmental parameters on them in a Mediterranean coastal area. J. Plankton Res. 1998, 20, 847–870. [Google Scholar] [CrossRef]
- Scoullos, M.J.; Rilley, J. Water circulation in the Gulf of Elefsis, Greece. Thalass. Jugosl. 1978, 14, 357–370. [Google Scholar]
- Pavlidou, A.; Pagou, K.; Assimakopoulou, G.; Rousselaki, E. Evolution over the last 30 years of the trophic conditions in the Gulf of Elefsis. In Sustainable Mediterranean; Issue No 71; Mediterranean Information Office for Environment, Culture and Sustainable Development, in collaboration with the European Environmental Bureau and the Arab NGO Network for Environment and Development: Athens, Greece, 2015; pp. 20–22. [Google Scholar]
- Jiang, C.; Chen, Z.; Lavoie, D.; Percival, J.B.; Kabanov, P. Mineral carbon minC (%) from Rock-Eval analysis as a reliable and cost-effective measurement of carbonate contents in shale source and reservoir rocks. Mar. Pet. Geol. 2017, 83, 184–194. [Google Scholar] [CrossRef]
- Facorellis, Y. Sea Surface radiocarbon reservoir age changes in the Aegean Sea from about 11,200 BP to present. Radiocarbon 2015, 57, 493–505. [Google Scholar] [CrossRef]
- Reimer, P.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Ramsey, C.B.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and Marine13 Radiocarbon Age Calibration Curves 0–50,000 Years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J.A.; Christen, A.J. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- Loeblich, A.R.; Tappan, H. Foraminiferal Genera and Their Classification; Van Nostrand Reinhold: New York, NY, USA, 1987. [Google Scholar]
- Loeblich, A.R.; Tappan, H. Foraminifera of the Sahul Shelf and Timor Sea; Cushman Foundation for Foraminiferal Research: Washington, DC, USA, 1994; Volume 31. [Google Scholar]
- Cimerman, F.; Langer, M. Mediterranean Foraminifera; Academia Scientiarum et Artium Slovenica: Ljubljana, Slovenia, 1991; Volume 30, Classis IV: Historia naturalis. [Google Scholar]
- Hottinger, L.; Halicz, E.; Reiss, Z. Recent Foraminiferida from the Gulf of Aqaba, Red Sea; Academia Scientiarum et Artium Slovenica, Classis IV: Ljubljana, Slovenia, 1993; Volume 33. [Google Scholar]
- Sgarrella, F.; Moncharmont-Zei, M. Benthic foraminifera of the Gulf of Naples (Italy): Systematics and autoecology. Boll. Soc. Paleontol. Ital. 1993, 32, 145–264. [Google Scholar]
- Dimiza, M.D.; Koukousioura, O.; Triantaphyllou, M.V.; Dermitzakis, M.D. Live and dead benthic foraminiferal assemblages from coastal environments of the Aegean Sea (Greece): Distribution and diversity. Rev. Micropaleontol. 2016, 59, 19–32. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T. Paleontological Data Analysis; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2006; ISBN 9781405115445. [Google Scholar]
- Bonaduce, G.; Ciampo, G.; Masoli, M. Distribution of Ostracoda in the Adriatic Sea. Pubblicazioni della Stazione zoologica di Napoli,: Napoli, Italy,, 1975; Volume 40. [Google Scholar]
- Stambolidis, E. Subrezente Ostracoden aus dem Evros-Delta (Griechenland) Einschliesslich der Entwicklung des Schlosses Gewisser Arten. Acta Universitatis Upsaliensis: Uppsala, Sweden, 1984. [Google Scholar]
- Tsourou, T. Composition and Distribution of Recent Marine Ostracod Assemblages in the Bottom Sediments of Central Aegean Sea (SE Andros Island, Greece). Int. Rev. Hydrobiol. 2012, 97, 276–300. [Google Scholar] [CrossRef]
- Tsourou, T.; Drinia, H.; Anastasakis, G. Ostracod assemblages from holocene middle shelf deposits of southern Evoikos gulf (Central Aegean Sea, Greece) and their palaeoenvironmental implications. Micropaleontology 2015, 61, 85–99. [Google Scholar]
- Triantaphyllou, M.V.; Antonarakou, A.; Kouli, K.; Dimiza, M.; Kontakiotis, G.; Papanikolaou, M.D.D.; Ziveri, P.; Mortyn, P.G.G.; Lianou, V.; Lykousis, V.; et al. Late Glacial-Holocene ecostratigraphy of the south-eastern Aegean Sea, based on plankton and pollen assemblages. Geo-Mar. Lett. 2009, 29, 249–267. [Google Scholar] [CrossRef]
- Flores, J.A.; Sierro, F.J.; Francés, G.; Vázquez, A.; Zamarreno, I. The last 100,000 years in the western Mediterranean: Sea surface water and frontal dynamics as revealed by coccolithophores. Mar. Micropaleontol. 1997, 29, 351–366. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Antonarakou, A.; Dimiza, M.; Anagnostou, C. Calcareous nannofossil and planktonic foraminiferal distributional patterns during deposition of sapropels S6, S5 and S1 in the Libyan Sea (Eastern Mediterranean). Geo-Mar. Lett. 2010, 30, 1–13. [Google Scholar] [CrossRef]
- Colmenero-Hidalgo, E.; Flores, J.-A.; Sierro, F.J.; Bárcena, M.Á.; Löwemark, L.; Schönfeld, J.; Grimalt, J.O. Ocean surface water response to short-term climate changes revealed by coccolithophores from the Gulf of Cadiz (NE Atlantic) and Alboran Sea (W Mediterranean). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 205, 317–336. [Google Scholar] [CrossRef]
- Cros, L. Planktonic Coccolithophores of the NW Mediterranean. Ph.D. Thesis, Univ. de Barcelona, Barcelona, Spain, 2001. [Google Scholar]
- Giraudeau, J. Distribution of recent nannofossils beneath the Benguela system: Southwest African continental margin. Mar. Geol. 1992, 108, 219–237. [Google Scholar] [CrossRef]
- Dimiza, M.D.; Triantaphyllou, M.V.; Malinverno, E. New evidence for the ecology of Helicosphaera carteri in polluted coastal environments (Elefsis Bay, Saronikos, Greece). J. Nannoplankt. Res. 2014, 34, 37–43. [Google Scholar]
- Dimiza, M.D.; Koukousioura, O.; Michailidis, I.; Dimou, V.G.; Navrozidou, V.; Aligizaki, K.; Seferlis, M. Seasonal living coccolithophore distribution in the enclosed coastal environments of the Thessaloniki Bay (Thermaikos Gulf, NW Aegean Sea). Rev. Micropaleontol. 2020, 69, 100449. [Google Scholar] [CrossRef]
- Kyrikou, S.; Kouli, K.; Triantaphyllou, M.V.; Dimiza, M.D.; Gogou, A.; Panagiotopoulos, I.P.; Anagnostou, C.; Karageorgis, A.P. Late Glacial and Holocene vegetation patterns of Attica: A high-resolution record from Elefsis Bay, southern Greece. Quat. Int. 2020, 545, 28–37. [Google Scholar] [CrossRef]
- Mudie, P.J.P.J.; Marret, F.; Mertens, K.N.; Shumilovskikh, L.; Leroy, S.A.G.G. Atlas of modern dinoflagellate cyst distributions in the Black Sea Corridor: From Aegean to Aral Seas, including Marmara, Black, Azov and Caspian Seas. Mar. Micropaleontol. 2017, 134, 1–152. [Google Scholar] [CrossRef]
- Van Nieuwenhove, N.; Head, M.J.; Limoges, A.; Pospelova, V.; Mertens, K.N.; Matthiessen, J.; De Schepper, S.; de Vernal, A.; Eynaud, F.; Londeix, L.; et al. An overview and brief description of common marine organic-walled dinoflagellate cyst taxa occurring in surface sediments of the Northern Hemisphere. Mar. Micropaleontol. 2020, 159, 101814. [Google Scholar] [CrossRef]
- Mudie, P.; Rochon, A.; Richards, K.; Ferguson, S.; Warny, S. Spiniferites cruciformis, Pterocysta cruciformis and Galeacysta etrusca: Morphology and palaeoecology. Palynology 2018, 42, 135–161. [Google Scholar] [CrossRef]
- Mertens, K.N.; Ribeiro, S.; Bouimetarhan, I.; Caner, H.; Combourieu-Nebout, N.; Dale, B.; De Vernal, A.; Ellegaard, M.; Filipova, M.; Godhe, A.; et al. Process length variation in cysts of a dinoflagellate, Lingulodinium machaerophorum, in surface sediments: Investigating its potential as salinity proxy. Mar. Micropaleontol. 2009, 70, 54–69. [Google Scholar] [CrossRef]
- Mertens, K.N.; Bradley, L.R.; Takano, Y.; Mudie, P.J.; Marret, F.; Aksu, A.E.; Hiscott, R.N.; Verleye, T.J.; Mousing, E.A.; Smyrnova, L.L.; et al. Quantitative estimation of Holocene surface salinity variation in the Black Sea using dinoflagellate cyst process length. Quat. Sci. Rev. 2012, 39, 45–59. [Google Scholar] [CrossRef]
- WoRMs Database. Available online: www.marinespecies.org (accessed on 1 April 2021).
- MSIP Database. Available online: www.species-identification.org (accessed on 1 April 2021).
- Sakellariou, E. The Extant Molluscs of the Gulf of Thessaloniki and Their Contribution to Stratigraphy. Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 1957. [Google Scholar]
- Gogou, A.; Bouloubassi, I.; Lykousis, V.; Arnaboldi, M.; Gaitani, P.; Meyers, P.A. Organic geochemical evidence of Late Glacial-Holocene climate instability in the North Aegean Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 256, 1–20. [Google Scholar] [CrossRef]
- Brassell, S.C.; Eglinton, G.; Marlowe, I.T.; Pflaumann, U.; Sarnthein, M. Molecular stratigraphy: A new tool for climatic assessment. Nature 1986, 320, 129–133. [Google Scholar] [CrossRef]
- Conte, M.H.; Sicre, M.-A.; Rühlemann, C.; Weber, J.C.; Schulte, S.; Schulz-Bull, D.; Blanz, T. Global temperature calibration of the alkenone unsaturation index (UK′37) in surface waters and comparison with surface sediments. Geochem. Geophys. Geosyst. 2006, 7, Q02005. [Google Scholar] [CrossRef]
- Eglinton, G.; Hamilton, R.J. Leaf epicutilar waxes. Science 1967, 256, 1322–1335. [Google Scholar] [CrossRef]
- Ohkouchi, N.; Kawamura, K.; Kawahata, H.; Taira, A. Latitudinal distributions of terrestrial biomarkers in the sediments from the Central Pacific. Geochim. Cosmochim. Acta 1997, 61, 1911–1918. [Google Scholar] [CrossRef]
- Poynter, J.G.; Farrimond, P.; Robinson, N.; Eglinton, G. Aeolian-Derived Higher Plant Lipids in the Marine Sedimentary Record: Links with Palaeoclimate BT—Paleoclimatology and Paleometeorology: Modern and Past Patterns of Global Atmospheric Transport; Leinen, M., Sarnthein, M., Eds.; Springer: Dordrecht, The Netherlands, 1989; pp. 435–462. ISBN 978-94-009-0995-3. [Google Scholar]
- Apostolopoulos, G.; Pavlopoulos, K.; Goiran, J.-P.; Fouache, E. Was the Piraeus peninsula (Greece) a rocky island? Detection of pre-Holocene rocky relief with borehole data and resistivity tomography analysis. J. Archaeol. Sci. 2014, 42, 412–421. [Google Scholar] [CrossRef]
- Tang, Z.; Cao, C.; Tang, K.; Qi, H.; Sun, Y.; Yang, J. Distribution of Carbon and Nitrogen as Indictors of Environmental Significance in Coastal Sediments of Weizhou Island, Beibu Gulf. Water 2020, 12, 3285. [Google Scholar] [CrossRef]
- Nelson, S.A. Carbonates & Other Rocks. Available online: http://www.tulane.edu/~sanelson/eens212/index.html (accessed on 1 April 2021).
- Guy-Holson, D. Botryococcus as an aid in the interpretation of palaeoenviroment and depositional processes. Rev. Palaeobot. Palynol. 1992, 71, 1–15. [Google Scholar] [CrossRef]
- Mudie, P.J.; Leroy, S.A.G.; Marret, F.; Gerasimenko, N.P.; Kholeif, S.E.A.; Sapelko, T.; Filipova-Marinova, M. Nonpollen palynomorphs: Indicators of salinity and environmental change in the Caspian-Black Sea-Mediterranean corridor. Spec. Pap. Geol. Soc. Am. 2011, 473, 89–115. [Google Scholar]
- Mariolakos, E.; Theochris, D. Shorelines displacement in the Saronic gulf area during the last 18,000 yrs and the Kihrea paleolake. Bull. Geol. Soc. Greece 2018, 34, 405–413. [Google Scholar] [CrossRef][Green Version]
- Roeser, P.; Franz, S.O.; Litt, T. Aragonite and calcite preservation in sediments from Lake Iznik related to bottom lake oxygenation and water column depth. Sedimentology 2016, 63, 2253–2277. [Google Scholar] [CrossRef]
- Antczak, M. Palaeoecological significance of Late Glacial and Holocene molluscs. Acta Biol. 2014, 21, 5–21. [Google Scholar]
- Soulié-Märsche, I. Charophytes, indicators for low salinity phases in North African sebkhet. J. Afr. Earth Sci. 2008, 51, 69–76. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Boltovskoy, E.; Giussani, G.; Watanabe, S.; Wright, R. Atlas of Benthic Shelf Foraminifera to the Southwest Atlantic; Dr. W. Junk bv Publishers: The Hague, The Netherlands, 1980. [Google Scholar]
- Melis, R.; Covelli, S. Distribution and morphological abnormalities of recent foraminifera in the Marano and Grado Lagoon (North Adriatic Sea, Italy). Mediterr. Mar. Sci. 2013, 14, 432–450. [Google Scholar] [CrossRef]
- Boudreau, R.E.A.; Patterson, R.T.; Dalby, A.P.; McKillop, W.B. Non-marine occurrence of the foraminifer Cribroelphidium gunteri in northern Lake Winnipegosis, Manitoba, Canada. J. Foraminifer. Res. 2001, 31, 108–119. [Google Scholar] [CrossRef]
- Almogi-Labin, A.; Perelis-Grossovicz, L.; Raab, M. Living Ammonia from a hypersaline inland pool, Dead Sea area, Israel. J. Foraminifer. Res. 1992, 22, 257–266. [Google Scholar] [CrossRef]
- Fatela, F.; Moreno, J.; Cabral, M.C. Salinity and water temperature assessment of the tidal marshes from the W Portuguese coast, as an ecological tool to palaeoenvironmental reconstructions based on foraminifera and ostracoda assemblages. Estud. Quat. 2016, 14, 73–81. [Google Scholar] [CrossRef]
- Hohenegger, J.; Piller, W.; Baal, C. Reasons for Spatial Microdistributions of Foraminifers in an Intertidal Pool (Northern Adriatic Sea). Mar. Ecol. 1989, 10, 43–78. [Google Scholar] [CrossRef]
- Poag, W.C. A Pilot Study of Paleoecophenotypic Variation Among Quaternary Paralic Foraminifera in the USGS-Belle Fontaine No. 1 Core, Mississippi Gulf Coast. In Stratigraphic and Paleontologic Studies of the Neogene and Quaternary Sediments in Southern Jackson County, Mississippi Chapter G.; U.S. Geological Survey: Reson, USA, 2001. [Google Scholar]
- Neale, J.W. Ostracods and palaeosalinity reconstruction. In Ostracoda Earth Sciences; Elsevier: Amsterdam, The Netherlands, 1988; pp. 125–155. [Google Scholar]
- Van Morkhoven, F.P.C.M.; van Morkhoven, F.P.C.M. Post-Paleozoic Ostracoda: Their Morphology, Taxonomy and Economic Use; Elsevier: Amsterdam, The Netherlands, 1962; Volume 1. [Google Scholar]
- Keyser, D. Histological peculiarities of the noding process in Cyprideis torosa (Jones) (Crustacea, Ostracoda). Hydrobiologia 2005, 538, 95–106. [Google Scholar] [CrossRef]
- Frenzel, P.; Schulze, I.; Pint, A. Noding of Cyprideis torosa valves (Ostracoda)—A proxy for salinity? New data from field observations and a long-term microcosm experiment. Int. Rev. Hydrobiol. 2012, 97, 314–329. [Google Scholar] [CrossRef]
- Rasmussen, S.O.; Bigler, M.; Blockley, S.P.; Blunier, T.; Buchardt, S.L.; Clausen, H.B.; Cvijanovic, I.; Dahl-Jensen, D.; Johnsen, S.J.; Fischer, H.; et al. A stratigraphic framework for abrupt climatic changes during the Last Glacial period based on three synchronized Greenland ice-core records: Refining and extending the INTIMATE event stratigraphy. Quat. Sci. Rev. 2014, 106, 14–28. [Google Scholar] [CrossRef]
- Kotthoff, U.; Muller, U.C.; Pross, J.; Schmiedl, G.; Lawson, I.T.; van de Schootbrugge, B.; Schulz, H. Lateglacial and Holocene vegetation dynamics in the Aegean region: An integrated view based on pollen data from marine and terrestrial archives. Holocene 2008, 18, 1019–1032. [Google Scholar] [CrossRef]
- Müller, U.C.; Pross, J.; Tzedakis, P.C.; Gamble, C.; Kotthoff, U.; Schmiedl, G.; Wulf, S.; Christanis, K. The role of climate in the spread of modern humans into Europe. Quat. Sci. Rev. 2011, 30, 273–279. [Google Scholar] [CrossRef]
- Panagiotopoulos, K.; Aufgebauer, A.; Schäbitz, F.; Wagner, B. Vegetation and climate history of the Lake Prespa region since the Lateglacial. Quat. Int. 2013, 293, 157–169. [Google Scholar] [CrossRef]
- Lawson, I.T.; Al-Omari, S.; Tzedakis, P.C.; Bryant, C.L.; Christanis, K. Lateglacial and Holocene vegetation history at Nisi Fen and the Boras mountains, northern Greece. Holocene 2005, 15, 873–887. [Google Scholar] [CrossRef]
- Pope, R.J.; Hughes, P.D.; Skourtsos, E. Glacial history of Mt Chelmos, Peloponnesus, Greece. Geol. Soc. Spec. Publ. 2017, 433, 211–236. [Google Scholar] [CrossRef]
- Hughes, P.D.; Woodward, J.C.; Gibbard, P.L. Quaternary glacial history of the Mediterranean mountains. Prog. Phys. Geogr. 2006, 30, 334–364. [Google Scholar] [CrossRef]
- Leontaritis, A.D.D.; Kouli, K.; Pavlopoulos, K. The glacial history of Greece: A comprehensive review. Mediterr. Geosci. Rev. 2020, 2, 65–90. [Google Scholar] [CrossRef]
- Allard, J.L.; Hughes, P.D.; Woodward, J.C.; Fink, D.; Simon, K.; Wilcken, K.M. Late Pleistocene glaciers in Greece: A new 36Cl chronology. Quat. Sci. Rev. 2020, 245, 106528. [Google Scholar] [CrossRef]
- Leroy, S.A.G.; Lahijani, H.A.K.; Reyss, J.L.; Chalié, F.; Haghani, S.; Shah-Hosseini, M.; Shahkarami, S.; Tudryn, A.; Arpe, K.; Habibi, P.; et al. A two-step expansion of the dinocyst Lingulodinium machaerophorum in the Caspian Sea: The role of changing environment. Quat. Sci. Rev. 2013, 77, 31–45. [Google Scholar] [CrossRef]
- Zonneveld, K.A.F.; Marret, F.; Versteegh, G.J.M.; Bogus, K.; Bonnet, S.; Bouimetarhan, I.; Crouch, E.; de Vernal, A.; Elshanawany, R.; Edwards, L.; et al. Atlas of modern dinoflagellate cyst distribution based on 2405 data points. Rev. Palaeobot. Palynol. 2013, 191, 1–197. [Google Scholar] [CrossRef]
- Leroy, S.A.G.; Albay, M. Palynomorphs of brackish and marine species in cores from the freshwater Lake Sapanca, NW Turkey. Rev. Palaeobot. Palynol. 2010, 160, 181–188. [Google Scholar] [CrossRef]
- McNeill, L.C.; Shillington, D.J.; Carter, G.D.O.; Everest, J.D.; Le Ber, E.; Collier, R.E.; Cvetkoska, A.; De Gelder, G.; Diz, P.; Doan, M.L.; et al. Site M0078. In Proceedings of the International Ocean Discovery Program; International Ocean Discovery Program: Texas, USA, 2019; Volume 381. [Google Scholar]
- Leroy, S.; Marret, F.; Gibert, E.; Chalie, F.; Reyss, J.-L.; Arpe, K. River inflow and salinity changes in the Caspian Sea during the last 5500 years. Quat. Sci. Rev. 2007, 26, 3359–3383. [Google Scholar] [CrossRef]
- Debenay, J.P.; Millet, B.; Angelidis, M.O. Relationships between foraminiferal assemblages and hydrodynamics in the Gulf of Kalloni, Greece. J. Foraminifer. Res. 2005, 35, 327–343. [Google Scholar] [CrossRef]
- Koukousioura, O.; Dimiza, M.D.; Kyriazidou, E.; Triantaphyllou, M.V.; Syrides, G.; Aidona, E.; Vouvalidis, K.; Panagiotopoulos, I.P.; Papadopoulou, L. Environmental evolution of the Paliouras coastal lagoon in the eastern Thermaikos gulf (Greece) during Holocene. Environ. Earth Sci. 2019, 78, 313. [Google Scholar] [CrossRef]
- Pavlopoulos, K.; Triantaphyllou, M.V.; Karymbalis, E.; Karkanas, P.; Kouli, K.; Tsourou, T. Landscape evolution recorded in the embayment of Palamari (Skyros Island, Greece) from the beginning of the Bronze Age until recent times. Géomorphol. Relief Process. Environ. 2007, 1, 37–48. [Google Scholar] [CrossRef][Green Version]
- Dimou, V.-G.; Koukousioura, O.; Dimiza, M.D.; Triantaphyllou, M.V.; Less, G.; Pomoni-Papaioannou, F.; Syrides, G. A preliminary investigation of Eocene larger benthic foraminifera assemblages from Alpine and molasse-type deposits of the Hellenic peninsula (Greece). Rev. Micropaleontol. 2021, 70, 100468. [Google Scholar] [CrossRef]
- Armynot Du Chatelet, E.; Degre, D.; Sauriau, P.-G.G.; Debenay, J.-P.P.; du Chatelet, E.A.; Degre, D.; Sauriau, P.-G.G.; Debenay, J.-P.P. Distribution of living benthic foraminifera in relation with environmental variables within the Aiguillon cove (Atlantic coast, France): Improving knowledge for paleoecological interpretation. Bull. Soc. Géologique Fr. 2009, 180, 131–144. [Google Scholar] [CrossRef]
- Athersuch, J. The genus Xestoleberis with particular reference to Recent Mediterranean species. Pubbl. Staz. Zool. Napoli 1976, 40, 282–343. [Google Scholar]
- Cronin, T.M.; Holmes, C.W.; Brewster-Wingard, G.L.; Ishman, S.E.; Dowsett, H.J.; Keyser, D.; Waibel, N. Historical trends in epiphytal ostracodes from Florida Bay: Implications for seagrass and macro-benthic algal variability. Bull. Am. Paleontol. 2001, 361, 159–197. [Google Scholar]
- Viehberg, F.A.; Frenzel, P.; Hoffmann, G. Succession of late Pleistocene and Holocene ostracode assemblages in a transgressive environment: A study at a coastal locality of the southern Baltic Sea (Germany). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 264, 318–329. [Google Scholar] [CrossRef]
- Athersuch, J. The ecology and distribution of the littoral ostracods of Cyprus. J. Nat. Hist. 1979, 13, 135–160. [Google Scholar] [CrossRef]
- Cabral, M.C.; Freitas, M.C.; Andrade, C.; Cruces, A. Coastal evolution and Holocene ostracods in Melides lagoon (SW Portugal). Mar. Micropaleontol. 2006, 60, 181–204. [Google Scholar] [CrossRef]
- Ruiz, F.; Abad, M.; Olías, M.; Galán, E.; González, I.; Aguilá, E.; Hamoumi, N.; Pulido, I.; Cantano, M. The present environmental scenario of the Nador Lagoon (Morocco). Environ. Res. 2006, 102, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, M.E.; Pugliese, N.; Bonaduce, G. Shelf ostracods distribution in the Italian seas. In What about Ostracoda! Proceedings of the 3rd European Ostracodologists Meeting, Bierville, France, 8–12 July 1996; Crasquin-Soleau, S, S., Braccini, E., Lethiers, F., Eds.; Bullentin du Centre de Recherches Elf Exploration-Production Mémoire, 20: Pau, France, 1998; pp. 91–101. [Google Scholar]
- Carboni, M.G.; Bergamin, L.; Di Bella, L.; Iamundo, F.; Pugliese, N. Palaeoecological evidences from foraminifers and ostracods on Late Quaternary sea-level changes in the Ombrone river plain (central Tyrrhenian coast, Italy). Mem. Spec. 2002, 24, 40–50. [Google Scholar]
- Eisma, D. The influence of salinity on Mollusk shell mineralogy: A discussion. J. Geol. 1966, 74, 89–94. [Google Scholar] [CrossRef]
- Tarnowska, K.; Wołowicz, M.; Chenuil, A.; Féral, J.P. Comparative studies on the morphometry and physiology of European populations of the lagoon specialist Cerastoderma glaucum (Bivalvia). Oceanologia 2009, 51, 437–458. [Google Scholar] [CrossRef]
- Öztürk, B.; Poutiers, J.-M.; Musa Sari, H.; Özbek, M. On the occurrence of Mytilaster marioni (Locard, 1889) (Mollusca; Bivalvia; Mytilidae) in Bafa Lake (Turkey), with a redescription of the species. Hydrobiologia 2002, 485, 123–131. [Google Scholar] [CrossRef]
- Fabbrocini, A.; Di Matteo, O.; D’Adamo, R. Abra segmentum (Mollusca: Pelecypoda) of the Lesina Lagoon (Southern Adriatic coast, Italy): Observations on variations in the population in relation to the main environmental parameters. Transit. Waters Bull. 2008, 1, 39–44. [Google Scholar]
- Gontikaki, E.; Antoniadou, C.; Chintiroglou, C.C. Population structure of Cerastoderma glaucum and Abra ovata in Vouliagmeni Lagoon (Attiki). J. Mar. Biol. Assoc. UK 2003, 83, 1095–1097. [Google Scholar] [CrossRef]
- Koukousioura, O.; Kouli, K.; Vouvalidis, K.; Aidona, E.; Karadimou, G.; Syrides, G. A multi-proxy approach for reconstructing environmental dynamics since the mid Holocene in Lake Ismarida (Thrace, N. Greece). Rev. Micropaleontol. 2020, 68, 100443. [Google Scholar] [CrossRef]
- Nossier, M.A. Ecophysiological responses of Cerastoderma edule (L.) and C. glaucum (BRUGUIÈRE) to different salinity regimes and exposure to air. J. Molluscan Stud. 1986, 52, 110–119. [Google Scholar] [CrossRef]
- Kevrekidis, T.; Gouvis, N.; Koukouras, A. Bionomy of Macrobenthic Molluscs in Evros Delta (North Aegean Sea). Int. Rev. Ges. Hydrobiol. 1996, 81, 455–468. [Google Scholar] [CrossRef]
- Koutsoubas, D.; Arvanitidis, C.; Dounas, C.; Drummond, L. Community structure and dynamics of the Molluscan Fauna in a Mediterranean lagoon (Gialova lagoon, SW Greece). Belg. J. Zool. 2000, 130, 131–138. [Google Scholar]
- Nicolaidou, A.; Bourgoutzani, F.; Zenetos, A.; Guelorget, O.; Perthuisot, J.-P.P. Distribution of molluscs and polychaetes in coastal lagoons in Greece. Estuar. Coast. Shelf Sci. 1988, 26, 337–350. [Google Scholar] [CrossRef]
- Malham, S.K.; Hutchinson, T.H.; Longshaw, M. A review of the biology of European cockles (Cerastoderma spp.). J. Mar. Biol. Assoc. UK 2012, 92, 1563–1577. [Google Scholar] [CrossRef]
- Colombani, N.; Osti, A.; Volta, G.; Mastrocicco, M. Impact of Climate Change on salinization of coastal water resources. Water Resour. Manag. 2016, 30, 2483–2496. [Google Scholar] [CrossRef]
- Kouli, K.; Gogou, A.; Bouloubassi, I.; Triantaphyllou, M.V.V.; Ioakim, C.; Katsouras, G.; Roussakis, G.; Lykousis, V. Late postglacial paleoenvironmental change in the northeastern Mediterranean region: Combined palynological and molecular biomarker evidence. Quat. Int. 2012, 261, 118–127. [Google Scholar] [CrossRef]
- Kouli, K.; Masi, A.; Mercuri, A.M.; Florenzano, A.; Sadori, L. Regional Vegetation Histories: An Overview of the Pollen Evidence from the Central Mediterranean. Late Antiq. Archaeol. 2018, 11, 69–82. [Google Scholar] [CrossRef]
- Dormoy, I.; Peyron, O.; Combourieu-Nebout, N.; Goring, S.; Kotthoff, U.; Magny, M.; Pross, J. Terrestrial climate variability and seasonality changes in the Mediterranean region between 15,000 and 4000 years BP deduced from marine pollen records. Clim. Past 2009, 5, 615–632. [Google Scholar] [CrossRef]
- Combourieu-Nebout, N.; Peyron, O.; Bout-Roumazeilles, V.; Goring, S.; Dormoy, I.; Joannin, S.; Sadori, L.; Siani, G.; Magny, M. Holocene vegetation and climate changes in the central Mediterranean inferred from a high-resolution marine pollen record (Adriatic Sea). Clim. Past 2013, 9, 2023–2042. [Google Scholar] [CrossRef]
- Aksu, A.E.; Abrajano, T.; Mudie, P.J.; Yaşar, D. Organic geochemical and palynological evidence for terrigenous origin of the organic matter in Aegean Sea sapropel S1. Mar. Geol. 1999, 153, 303–318. [Google Scholar] [CrossRef]
- Morzadec-Kerfourn, M.T. Interaction between sea-level changes and the development of littoral herbaceous vegetation and autotrophic dinoflagellates. Quat. Int. 2005, 133–134, 137–140. [Google Scholar] [CrossRef]
- Dimiza, M.D.; Triantaphyllou, M.V.; Dermitzakis, M.D. Seasonality and ecology of living coccolithophores in Eastern Mediterranean coastal environments (Andros Island, Middle Aegean Sea). Micropaleontology 2008, 54, 159–175. [Google Scholar]
- Moulfi-El-Houari, L.; Ambroise, D.; Mathieu, R. Distribution des foraminifères benthiques actuels sur la marge continentale Algéroise (Baie De Bou-Ismaïl). Rev. Micropaleontol. 1999, 42, 315–327. [Google Scholar] [CrossRef]
- Jorissen, F.J. The distribution of benthic foraminifera in the Adriatic Sea. Mar. Micropaleontol. 1987, 12, 21–48. [Google Scholar] [CrossRef]
- Langer, M.R. Epiphytic foraminifera. Mar. Micropaleontol. 1993, 20, 235–265. [Google Scholar] [CrossRef]
- Boomer, I.; Guichard, P.; Lericolais, G. Late Pleistocene to Recent ostracod assemblages from the western Black Sea. J. Micropalaeontol. 2010, 29, 119–133. [Google Scholar] [CrossRef]
- Frezza, V.; Di Bella, L. Distribution of recent ostracods near the Ombrone River mouth (Northern Tyrrhenian Sea, Italy). Micropaleontology 2015, 61, 101–114. [Google Scholar]
- Barbieri, G.; Rossi, V.; Vaiani, S.C.; Horton, B.P. Benthic ostracoda and foraminifera from the North Adriatic Sea (Italy, Mediterranean Sea): A proxy for the depositional characterisation of river-influenced shelves. Mar. Micropaleontol. 2019, 153, 101772. [Google Scholar] [CrossRef]
- Heusser, L.; Balsam, W.L. Pollen Distribution in the Northeast Pacific Ocean. Quat. Res. 1977, 7, 45–62. [Google Scholar] [CrossRef]
- Mudie, P.J. Pollen distribution in recent marine sediments, eastern Canada. Can. J. Earth Sci. 1982, 19, 729–747. [Google Scholar] [CrossRef]
- De Vernal, A. Palynology (Pollen, Spores, etc.). In Encyclopedia of Marine Geosciences; Springer Science + Business Media: Dordrecht, The Netherlands, 2015; pp. 1–9. ISBN 9789400766440. [Google Scholar]
- Naughton, F.; Sanchez Goñi, M.F.; Desprat, S.; Turon, J.L.; Duprat, J.; Malaizé, B.; Joli, C.; Cortijo, E.; Drago, T.; Freitas, M.C. Present-day and past (last 25,000 years) marine pollen signal off western Iberia. Mar. Micropaleontol. 2007, 62, 91–114. [Google Scholar] [CrossRef]
- Gogou, A.; Triantaphyllou, M.V.; Xoplaki, E.; Izdebski, A.; Parinos, C.; Dimiza, M.; Bouloubassi, I.; Luterbacher, J.; Kouli, K.; Martrat, B.; et al. Climate variability and socio-environmental changes in the northern Aegean (NE Mediterranean) during the last 1500 years. Quat. Sci. Rev. 2016, 136, 209–228. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Mazzanti, M.B.; Torri, P.; Vigliotti, L.; Bosi, G.; Florenzano, A.; Olmi, L.; N’siala, I.M. A marine/terrestrial integration for mid-late Holocene vegetation history and the development of the cultural landscape in the Po valley as a result of human impact and climate change. Veg. Hist. Archaeobot. 2012, 21, 353–372. [Google Scholar] [CrossRef]
- Sadori, L.; Koutsodendris, A.; Masi, A.; Bertini, A.; Combourieu-Nebout, N.; Francke, A.; Kouli, K.; Joannin, S.; Mercuri, A.M.; Panagiotopoulos, K.; et al. Pollen-based paleoenvironmental and paleoclimatic change at Lake Ohrid (SE Europe) during the past 500 ka. Biogeosciences 2016, 13, 1423–1437. [Google Scholar] [CrossRef]
- Panagiotopoulos, K.; Holtvoeth, J.; Kouli, K.; Marinova, E.; Francke, A.; Cvetkoska, A.; Jovanovska, E.; Lacey, J.H.; Lyons, E.T.; Buckel, C.; et al. Insights into the evolution of the young Lake Ohrid ecosystem and vegetation succession from a southern European refugium during the Early Pleistocene. Quat. Sci. Rev. 2020, 227, 106044. [Google Scholar] [CrossRef]
- Sadori, L. The Lateglacial and Holocene vegetation and climate history of Lago di Mezzano (central Italy). Quat. Sci. Rev. 2018, 202, 30–44. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Gogou, A.; Bouloubassi, I.; Dimiza, M.; Kouli, K.; Rousakis, G.; Kotthoff, U.; Emeis, K.C.; Papanikolaou, M.; Athanasiou, M.; et al. Evidence for a warm and humid Mid-Holocene episode in the Aegean and northern Levantine Seas (Greece, NE Mediterranean). Reg. Environ. Chang. 2014, 14, 1697–1712. [Google Scholar] [CrossRef]
- Cullen, H.M.; DeMenocal, P.B.; Hemming, S.; Hemming, G.; Brown, F.H.; Guilderson, T.; Sirocko, F. Climate change and the collapse of the Akkadian empire: Evidence from the deep sea. Geology 2000, 28, 379. [Google Scholar] [CrossRef]
- Finné, M.; Holmgren, K.; Sundqvist, H.S.; Weiberg, E.; Lindblom, M. Climate in the eastern Mediterranean, and adjacent regions, during the past 6000 years—A review. J. Archaeol. Sci. 2011, 38, 3153–3173. [Google Scholar] [CrossRef]
- Marret, F.; Zonneveld, K.A.F. Atlas of modern organic-walled dinoflagellate cyst distribution. Rev. Palaeobot. Palynol. 2003, 125, 1–200. [Google Scholar] [CrossRef]
- Peyron, O.; Combourieu-Nebout, N.; Brayshaw, D.; Goring, S.; Andrieu-Ponel, V.; Desprat, S.; Fletcher, W.; Gambin, B.; Ioakim, C.; Joannin, S.; et al. Precipitation changes in the Mediterranean basin during the Holocene from terrestrial and marine pollen records: A model–data comparison. Clim. Past 2017, 13, 249–265. [Google Scholar] [CrossRef]
- Roberts, N.; Eastwood, W.J.; Kuzucuoglu, C.; Fiorentino, G.; Caracuta, V. Climatic, vegetation and cultural change in the eastern Mediterranean during the mid-Holocene environmental transition. Holocene 2011, 21, 147–162. [Google Scholar] [CrossRef]
- Sadori, L.; Jahns, S.; Peyron, O. Mid-Holocene vegetation history of the central Mediterranean. Holocene 2011, 21, 117–129. [Google Scholar] [CrossRef]
- Barmawidjaja, D.M.; Jorissen, F.J.; Puskaric, S.; van der Zwaan, G.J. Microhabitat selection by benthic foraminifera in the northern Adriatic Sea. J. Foraminifer. Res. 1992, 22, 297–317. [Google Scholar] [CrossRef]
- Jorissen, F.J.; Barmawidjaja, D.M.; Puskaric, S.; van der Zwaan, G.J. Vertical distribution of benthic foraminifera in the northern Adriatic Sea: The relation with the organic flux. Mar. Micropaleontol. 1992, 19, 131–146. [Google Scholar] [CrossRef]
- Goineau, A.; Fontanier, C.; Mojtahid, M.; Fanget, A.S.; Bassetti, M.A.; Berné, S.; Jorissen, F. Live-dead comparison of benthic foraminiferal faunas from the Rhône prodelta (Gulf of Lions, NW Mediterranean): Development of a proxy for palaeoenvironmental reconstructions. Mar. Micropaleontol. 2015, 119, 17–33. [Google Scholar] [CrossRef]
- Moodley, L.; Heip, C.H.R.; Middelburg, J.J. Benthic activity in sediments of the northwestern Adriatic Sea: Sediment oxygen consumption, macro- and meiofauna dynamics. J. Sea Res. 1998, 40, 263–280. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Sen Gupta, B.K. Foraminifera in oxygen-depleted environments. In Modern Foraminifera; Sen Gupta, B.K., Ed.; Kluwer: Dordrecht, The Netherlands, 1999; pp. 201–216. [Google Scholar]
- Diaz, R.J.; Rosenberg, R. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 245–303. [Google Scholar]
- Jensen, J.N. Increased abundance and growth of the suspension-feeding bivalve Corbula gibba in a shallow part of the eutrophic Limfjord, Denmark. Neth. J. Sea Res. 1990, 27, 101–108. [Google Scholar] [CrossRef]
- Wilding, T.A. The Benthic Impacts of the Loch Linnhe Artificial Reef. Hydrobiologia 2006, 555, 345–353. [Google Scholar] [CrossRef]
- Büyükmeriç, Y.; Wesselingh, F.P.; Alçiçek, M.C. Middle–late Pleistocene marine molluscs from Izmit Bay area (eastern Marmara Sea, Turkey) and the nature of Marmara—Black Sea corridors. Quat. Int. 2016, 401, 153–161. [Google Scholar] [CrossRef]
- Triantaphyllou, M. Coccolithophores—And related taxa—In the modern oceans. Rev. Micropaleontol. 2016, 59, 225. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab Code | Depth (cm) | Material | 14C yr BP | cal BP |
|---|---|---|---|---|
| Beta—496147 | 8–9 | Turritella communis | 970 ± 30 | 451–563 1 |
| Beta—441157 | 40–42 | Turritella communis | 2270 ± 30 | 1706–1872 1 |
| Beta—496148 | 70–72 | Turritella communis | 3930 ± 30 | 3711–3904 1 |
| Beta—441158 | 120–121 | Turritella communis | 4260 ± 30 | 4179–4378 1 |
| Beta—453192 | 155–156 | Turritella communis | 5840 ± 30 | 6113–6268 1 |
| Beta—453193 | 180–181 | Turritella communis | 6640 ± 30 | 6992–7163 1 |
| Beta—441159 | 242–244 | Cerastoderma glacum | 10,220 ± 30 | 11,083–11,218 1 |
| Beta—441160 | 297–300 | peat | 11,650 ± 40 | 13,439–13,496 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouli, K.; Triantaphyllou, M.V.; Koukousioura, O.; Dimiza, M.D.; Parinos, C.; Panagiotopoulos, I.P.; Tsourou, T.; Gogou, A.; Mavrommatis, N.; Syrides, G.; et al. Late Glacial Marine Transgression and Ecosystem Response in the Landlocked Elefsis Bay (Northern Saronikos Gulf, Greece). Water 2021, 13, 1505. https://doi.org/10.3390/w13111505
Kouli K, Triantaphyllou MV, Koukousioura O, Dimiza MD, Parinos C, Panagiotopoulos IP, Tsourou T, Gogou A, Mavrommatis N, Syrides G, et al. Late Glacial Marine Transgression and Ecosystem Response in the Landlocked Elefsis Bay (Northern Saronikos Gulf, Greece). Water. 2021; 13(11):1505. https://doi.org/10.3390/w13111505
Chicago/Turabian StyleKouli, Katerina, Maria V. Triantaphyllou, Olga Koukousioura, Margarita D. Dimiza, Constantine Parinos, Ioannis P. Panagiotopoulos, Theodora Tsourou, Alexandra Gogou, Nikolaos Mavrommatis, George Syrides, and et al. 2021. "Late Glacial Marine Transgression and Ecosystem Response in the Landlocked Elefsis Bay (Northern Saronikos Gulf, Greece)" Water 13, no. 11: 1505. https://doi.org/10.3390/w13111505
APA StyleKouli, K., Triantaphyllou, M. V., Koukousioura, O., Dimiza, M. D., Parinos, C., Panagiotopoulos, I. P., Tsourou, T., Gogou, A., Mavrommatis, N., Syrides, G., Kyrikou, S., Skampa, E., Skylaki, E., Anagnostou, C., & Karageorgis, A. P. (2021). Late Glacial Marine Transgression and Ecosystem Response in the Landlocked Elefsis Bay (Northern Saronikos Gulf, Greece). Water, 13(11), 1505. https://doi.org/10.3390/w13111505