Spatiotemporal Changes of Antibiotic Resistance and Bacterial Communities in Drinking Water Distribution System in Wrocław, Poland



Abstract
1. Introduction
2. Materials and Methods
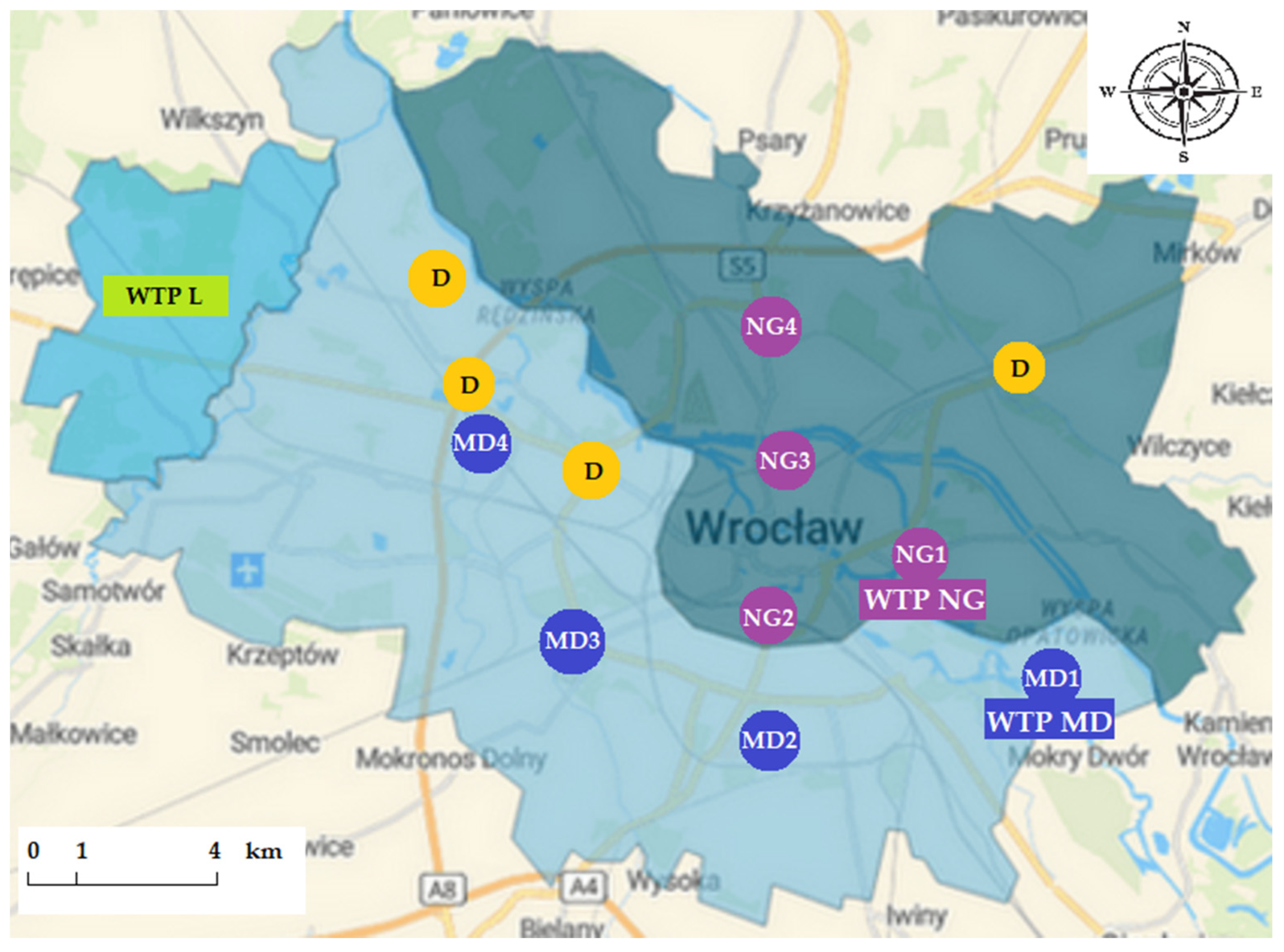
2.1. DWDS in Wrocław
2.2. Sample Collection
2.3. Relative Abundances of ARB and Total Number of Bacteria
2.4. DNA Extraction
2.5. Detection of ARGs and Other Genes in Environmental DNA
2.6. DGGE Analysis of Community Structures
2.7. Statistical Analyses
3. Results
3.1. Tap Water Parameters
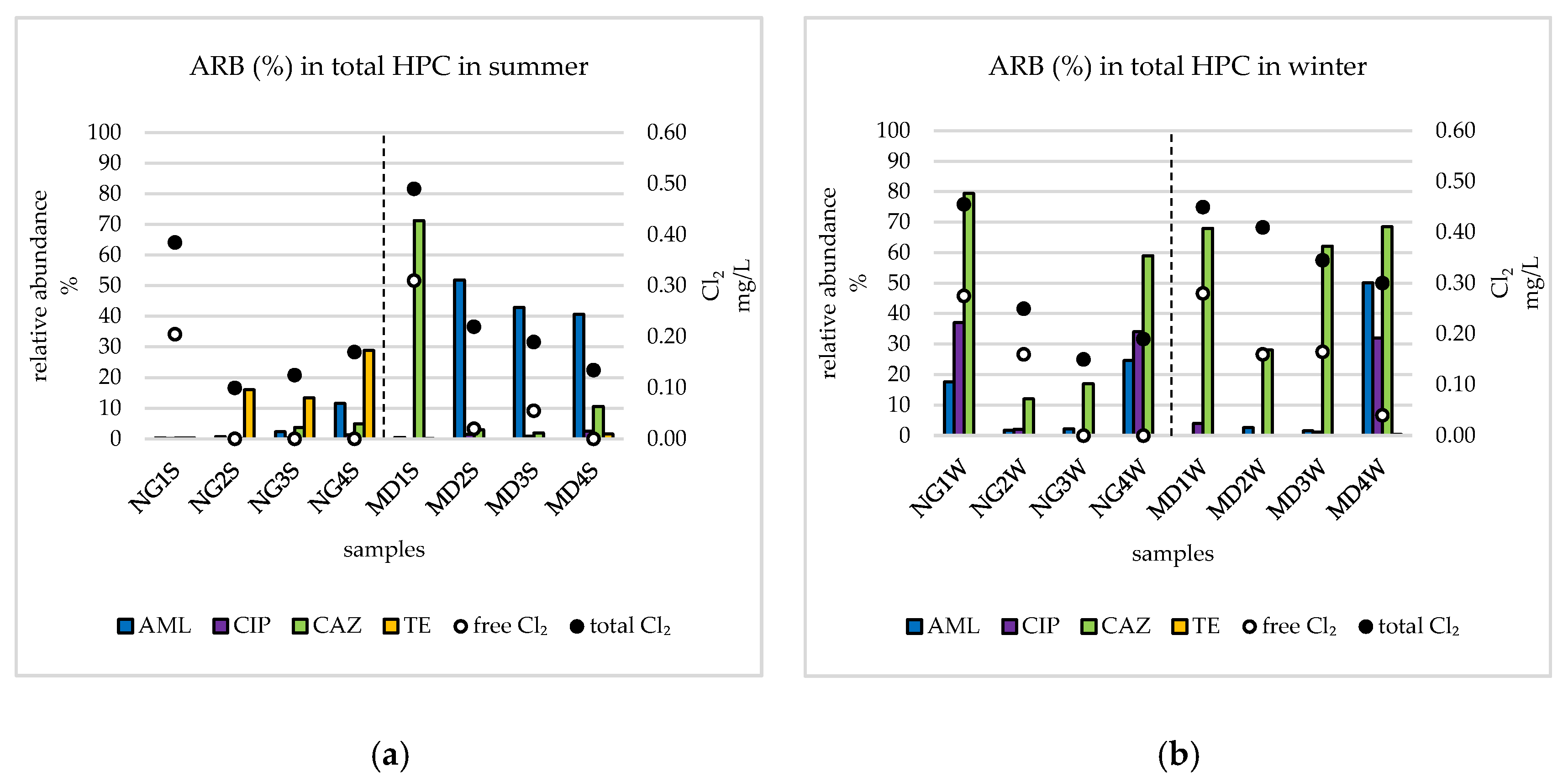
3.2. Relative Abundances of ARB and Total Number of Bacteria
3.3. Detection of ARGs and Other Genes in Environmental DNA
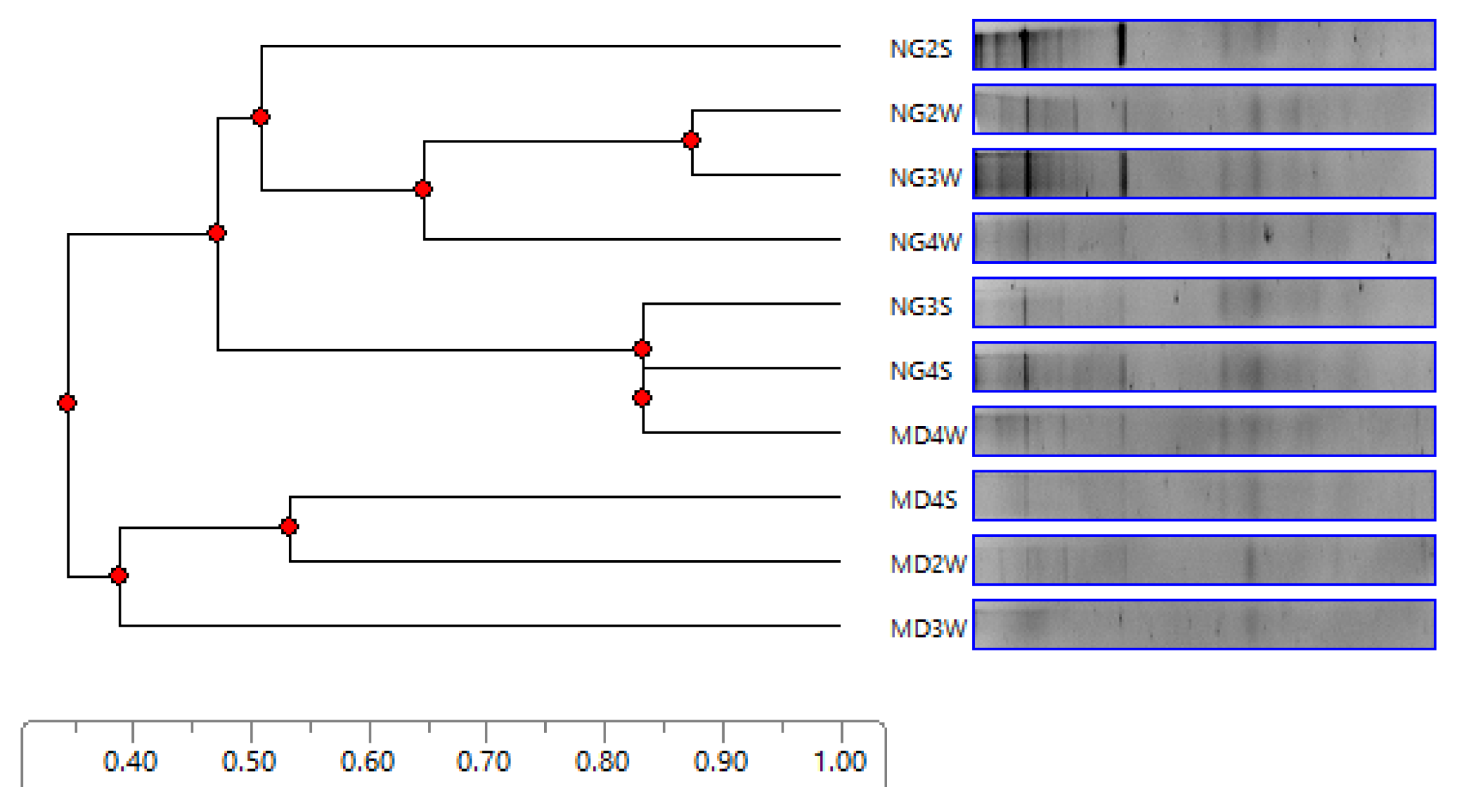
3.4. DGGE Analysis of the Community Structures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diwan, V.; Hanna, N.; Purohit, M.; Chandran, S.; Riggi, E.; Parashar, V.; Tamhankar, A.J.; Lundborg, C.S. Seasonal variations in water-quality, antibiotic residues, resistant bacteria and antibiotic resistance genes of Escherichia coli isolates from water and sediments of the Kshipra River in Central India. Int. J. Environ. Res. Public Health 2018, 15, 1281. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.L. The role of natural environments in the evolution of resistance traits in pathogenic bacteria. Proc. R. Soc. B Biol. Sci. 2009, 276, 2521–2530. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.W.; Giesen, M.J.; Bunce, J.T. Strategic approach for prioritising local and regional sanitation interventions for reducing global antibiotic resistance. Water 2018, 11, 27. [Google Scholar] [CrossRef]
- Ng, C.; Gin, K.Y.H. Monitoring antimicrobial resistance dissemination in aquatic systems. Water 2019, 11, 71. [Google Scholar] [CrossRef]
- Hong, P.Y.; Julian, T.R.; Pype, M.L.; Jiang, S.C.; Nelson, K.L.; Graham, D.; Pruden, A.; Manaia, C.M. Reusing treated wastewater: Consideration of the safety aspects associated with antibiotic-resistant bacteria and antibiotic resistance genes. Water 2018, 10, 244. [Google Scholar] [CrossRef]
- WHO. Antibiotic Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance (accessed on 1 July 2020).
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Bacterial diversity and antibiotic resistance in water habitats: Searching the links with the human microbiome. FEMS Microbiol. Rev. 2014, 38, 761–778. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.-L.; Cantón, R. Antibiotics and antibiotic resistance in water environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef]
- Ma, L.; Li, B.; Jiang, X.T.; Wang, Y.L.; Xia, Y.; Li, A.D.; Zhang, T. Catalogue of antibiotic resistome and host-tracking in drinking water deciphered by a large scale survey. Microbiome 2017, 5, 154. [Google Scholar] [CrossRef]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, H.; Zhang, L.; Jiang, Y.; Gin, K.Y.H.; He, Y. Occurrence, distribution, and risk assessment of antibiotics in a subtropical river-reservoir system. Water 2018, 10, 104. [Google Scholar] [CrossRef]
- Pruden, A.; Larsson, D.G.J.; Amézquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management of Options for Reducing the Release of Antibiotics. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Berglund, B. Environmental dissemination of antibiotic resistance genes and correlation to anthropogenic contamination with antibiotics. Infect. Ecol. Epidemiol. 2015, 5, 28564. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, U.; Wiergowski, M.; Sołtyszewski, I.; Kuzemko, J.; Wiergowska, G.; Woźniak, M.K. Presence of antibiotics in the aquatic environment in Europe and their analytical monitoring: Recent trends and perspectives. Microchem. J. 2019, 147, 729–740. [Google Scholar] [CrossRef]
- Casanova, L.M.; Sobsey, M.D. Antibiotic-resistant enteric bacteria in environmental waters. Water 2016, 8, 561. [Google Scholar] [CrossRef]
- Kubera, Ł.; Malecka-Adamowicz, M.; Jankowiak, E.; Dembowska, E.; Perliński, P.; Hejze, K. Influence of environmental and anthropogenic factors on microbial ecology and sanitary threat in the final stretch of the Brda River. Water 2019, 11, 922. [Google Scholar] [CrossRef]
- Thakali, O.; Tandukar, S.; Brooks, J.P.; Sherchan, S.P.; Sherchand, J.B.; Haramoto, E. The occurrence of antibiotic resistance genes in an Urban River in Nepal. Water 2020, 12, 450. [Google Scholar] [CrossRef]
- Xu, L.; Ouyang, W.; Qian, Y.; Su, C.; Su, J.; Chen, H. High-throughput profiling of antibiotic resistance genes in drinking water treatment plants and distribution systems. Environ. Pollut. 2016, 213, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Ma, X.; Xu, F.; Li, J.; Zhang, H.; Xiao, X. The drinking water treatment process as a potential source of affecting the bacterial antibiotic resistance. Sci. Total Environ. 2015, 533, 24–31. [Google Scholar] [CrossRef]
- Chao, Y.; Ma, L.; Yang, Y.; Ju, F.; Zhang, X.X.; Wu, W.M.; Zhang, T. Metagenomic analysis reveals significant changes of microbial compositions and protective functions during drinking water treatment. Sci. Rep. 2013, 3, 3550. [Google Scholar] [CrossRef]
- Guo, X.; Li, J.; Yang, F.; Yang, J.; Yin, D. Prevalence of sulfonamide and tetracycline resistance genes in drinking water treatment plants in the Yangtze River Delta, China. Sci. Total Environ. 2014, 493, 626–631. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, T.; Chen, H. Antibiotic resistome promotion in drinking water during biological activated carbon treatment: Is it influenced by quorum sensing? Sci. Total Environ. 2018, 612, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ferro, G.; Guarino, F.; Cicatelli, A.; Rizzo, L. β-lactams resistance gene quantification in an antibiotic resistant Escherichia coli water suspension treated by advanced oxidation with UV/H2O2. J. Hazard. Mater. 2017, 323, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Jikumaru, A.; Ueno, T.; Sei, K. Inactivation effect of antibiotic-resistant gene using chlorine disinfection. Water 2017, 9, 547. [Google Scholar] [CrossRef]
- Hao, H.; Shi, D.-Y.; Yang, D.; Yang, Z.-W.; Qiu, Z.-G.; Liu, W.-L.; Shen, Z.-Q.; Yin, J.; Wang, H.-R.; Li, J.-W.; et al. Profiling of intracellular and extracellular antibiotic resistance genes in tap water. J. Hazard. Mater. 2019, 365, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Shi, P.; Hu, Q.; Li, B.; Zhang, T.; Zhang, X.X. Bacterial Community Shift Drives Antibiotic Resistance Promotion during Drinking Water Chlorination. Environ. Sci. Technol. 2015, 49, 12271–12279. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, M.; Zhang, S.; Yu, X. Can chlorination co-select antibiotic-resistance genes? Chemosphere 2016, 156, 412–419. [Google Scholar] [CrossRef]
- Liu, S.S.; Qu, H.M.; Yang, D.; Hu, H.; Liu, W.L.; Qiu, Z.G.; Hou, A.M.; Guo, J.; Li, J.W.; Shen, Z.Q.; et al. Chlorine disinfection increases both intracellular and extracellular antibiotic resistance genes in a full-scale wastewater treatment plant. Water Res. 2018, 136, 131–136. [Google Scholar] [CrossRef]
- Lu, J.; Tian, Z.; Yu, J.; Yang, M.; Zhang, Y. Distribution and abundance of antibiotic resistance genes in sand settling reservoirs and drinking water treatment plants across the Yellow River, China. Water 2018, 10, 246. [Google Scholar] [CrossRef]
- Shi, P.; Jia, S.; Zhang, X.-X.; Zhang, T.; Cheng, S.; Li, A. Metagenomic insights into chlorination effects on microbial antibiotic resistance in drinking water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef]
- Su, H.C.; Liu, Y.S.; Pan, C.G.; Chen, J.; He, L.Y.; Ying, G.G. Persistence of antibiotic resistance genes and bacterial community changes in drinking water treatment system: From drinking water source to tap water. Sci. Total Environ. 2018, 616–617, 453–461. [Google Scholar] [CrossRef]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl. Environ. Microbiol. 2009, 75, 5714–5718. [Google Scholar] [CrossRef] [PubMed]
- McCoy, S.T.; Vanbriesen, J.M. Temporal variability of bacterial diversity in a chlorinated drinking water distribution system. J. Environ. Eng. 2012, 138, 786–795. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Egas, C.; Nunes, O.C.; Manaia, C.M. Bacterial diversity from the source to the tap: A comparative study based on 16S rRNA gene-DGGE and culture-dependent methods. FEMS Microbiol. Ecol. 2013, 83, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Edberg, S.C.; Allen, M.J. Virulence and risk from drinking water of heterotrophic plate count bacteria in human population groups. Int. J. Food Microbiol. 2004, 92, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Seidman, J.C.; Anitha, K.P.; Kanungo, R.; Bourgeois, A.L.; Coles, C.L. Risk factors for antibiotic-resistant E. coli in children in a rural area. Epidemiol. Infect. 2009, 137, 879–888. [Google Scholar] [CrossRef]
- Coleman, B.L.; Salvadori, M.I.; McGeer, A.J.; Sibley, K.A.; Neumann, N.F.; Bondy, S.J.; Gutmanis, I.A.; McEwn, S.A.; Lavoie, M.; Strong, D.; et al. The role of drinking water in the transmission of antimicrobial-resistant E. coli. Epidemiol. Infect. 2012, 140, 633–642. [Google Scholar] [CrossRef]
- Salyers, A.A.; Gupta, A.; Wang, Y. Human intestinal bacteria as reservoirs for antibiotic resistance genes. Trends Microbiol. 2004, 12, 412–416. [Google Scholar] [CrossRef]
- Armstrong, J.L.; Calomiris, J.J.; Seidler, R.J. Selection of antibiotic-resistant standard plate count bacteria during water treatment. Appl. Environ. Microbiol. 1982, 44, 308–316. [Google Scholar] [CrossRef]
- Armstrong, J.L.; Shigeno, D.S.; Calomiris, J.J.; Seidler, R.J. Antibiotic-resistant bacteria in drinking water. Appl. Environ. Microbiol. 1981, 42, 277–283. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC244002/ (accessed on 1 July 2020).
- Bergeron, S.; Boopathy, R.; Nathaniel, R.; Corbin, A.; LaFleur, G. Presence of antibiotic resistant bacteria and antibiotic resistance genes in raw source water and treated drinking water. Int. Biodeterior. Biodegrad. 2015, 102, 370–374. [Google Scholar] [CrossRef]
- Faria, C.; Vaz-Moreira, I.; Serapicos, E.; Nunes, O.C.; Manaia, C.M. Antibiotic resistance in coagulase negative staphylococci isolated from wastewater and drinking water. Sci. Total Environ. 2009, 407, 3876–3882. [Google Scholar] [CrossRef]
- Figueira, V.; Serra, E.A.; Vaz-Moreira, I.; Brandão, T.R.S.; Manaia, C.M. Comparison of ubiquitous antibiotic-resistant Enterobacteriaceae populations isolated from wastewaters, surface waters and drinking waters. J. Water Health 2012, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Figueira, V.; Vaz-Moreira, I.; Silva, M.; Manaia, C.M. Diversity and antibiotic resistance of Aeromonas spp. in drinking and waste water treatment plants. Water Res. 2011, 45, 5599–5611. [Google Scholar] [CrossRef] [PubMed]
- Flores Ribeiro, A.; Bodilis, J.; Alonso, L.; Buquet, S.; Feuilloley, M.; Dupont, J.P.; Pawlak, B. Occurrence of multi-antibiotic resistant Pseudomonas spp. in drinking water produced from karstic hydrosystems. Sci. Total Environ. 2014, 490, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Beattie, T.K.; Knapp, C.W. Relationship between antibiotic- and disinfectant-resistance profiles in bacteria harvested from tap water. Chemosphere 2016, 152, 132–141. [Google Scholar] [CrossRef]
- Khan, S.; Knapp, C.W.; Beattie, T.K. Antibiotic resistant bacteria found in municipal drinking water. Environ. Process. 2016, 3, 541–552. [Google Scholar] [CrossRef]
- Leginowicz, M.; Siedlecka, A.; Piekarska, K. Biodiversity and antibiotic resistance of bacteria isolated from tap water in Wrocław, Poland. Environ. Prot. Eng. 2018, 44, 85–98. [Google Scholar] [CrossRef]
- Narciso-da-Rocha, C.; Vaz-Moreira, I.; Manaia, C.M. Genotypic diversity and antibiotic resistance in Sphingomonadaceae isolated from hospital tap water. Sci. Total Environ. 2014, 466–467, 127–135. [Google Scholar] [CrossRef]
- Narciso-Da-Rocha, C.; Vaz-Moreira, I.; Svensson-Stadler, L.; Moore, E.R.B.; Manaia, C.M. Diversity and antibiotic resistance of Acinetobacter spp. in water from the source to the tap. Appl. Microbiol. Biotechnol. 2013, 97, 329–340. [Google Scholar] [CrossRef]
- Rocha, J.; Fernandes, T.; Riquelme, M.V.; Zhu, N.; Pruden, A.; Manaia, C.M. Comparison of culture-and quantitative PCR-based indicators of antibiotic resistance in wastewater, recycled water, and tap water. Int. J. Environ. Res. Public Health 2019, 16, 4217. [Google Scholar] [CrossRef]
- Schwartz, T.; Kohnen, W.; Jansen, B.; Obst, U. Detection of antibiotic-resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms. FEMS Microbiol. Ecol. 2003, 43, 325–335. [Google Scholar] [CrossRef]
- Siedlecka, A.; Piekarska, K. Application of NGS for studying the antibiotic resistant microbiome in Wrocław tap water. In Proceedings of the 10th Conference on Interdisciplinary Problems in Environmental Protection and Engineering EKO-DOK 2018, E3S Web of Conferences, 44(00161), Polanica-Zdrój, Poland, 16–18 April 2018. [Google Scholar] [CrossRef]
- Siedlecka, A.; Piekarska, K. Antibiotic resistance in tap water during the summer season—Preliminary research. In Proceedings of the Nternational Conference on Advances in Energy Systems and Environmental Engineering (β19), E3S Web of Conferences, 116(00077), Wrocław, Poland, 9–12 June 2019. [Google Scholar] [CrossRef]
- Siedlecka, A.; Wolf-Baca, M.; Piekarska, K. Seasonal variability of antibiotic resistance and biodiversity of tap water bacteria in Wrocław, Poland. Environ. Prot. Eng. 2020, 46, 93–109. [Google Scholar] [CrossRef]
- Subba, P.; Joshi, D.R.; Bhatta, D.R. Antibiotic resistance pattern and plasmid profiling of thermotolerant Escherichia coli isolates in drinking water. J. Nepal Health Res. Counc. 2013, 11, 44–48. [Google Scholar] [PubMed]
- Talukdar, P.K.; Rahman, M.; Rahman, M.; Nabi, A.; Islam, Z.; Hoque, M.M.; Endtz, H.P.; Islam, M.A. Antimicrobial Resistance, Virulence Factors and Genetic Diversity of Escherichia coli Isolates from Household Water Supply in Dhaka, Bangladesh. PLoS ONE 2013, 8, e61090. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance patterns of Sphingomonadaceae isolates from drinking water. Appl. Environ. Microbiol. 2011, 77, 5697–5706. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance in Pseudomonas spp. from drinking water. Sci. Total Environ. 2012, 426, 366–374. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Ubiquitous and persistent Proteobacteria and other Gram-negative bacteria in drinking water. Sci. Total Environ. 2017, 586, 1141–1149. [Google Scholar] [CrossRef]
- European Surveillance of Antimicrobial Consumption Network (ESAC-Net). Available online: https://www.ecdc.europa.eu/en/about-us/partnerships-and-networks/disease-and-laboratory-networks/esac-net (accessed on 1 July 2020).
- Wolf-Baca, M.; Piekarska, K. Biodiversity of organisms inhabiting the water supply network of Wroclaw. Detection of pathogenic organisms constituting a threat for drinking water recipients. Sci. Total Environ. 2020, 715, 136732. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, H.M.; Yoo, D.G.; Kim, J.H. A comparative study on a hydraulic and water-quality analysis method for determining rechlorination injection points for a water-supply network. Water 2019, 11, 697. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2002, 49, 1049. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing—EUCAST. Available online: https://eucast.org/ (accessed on 1 July 2020).
- Macrogen USA—Universal Primer List. Available online: https://www.macrogenusa.com/support/seq/primer.jsp (accessed on 1 July 2020).
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC202176/ (accessed on 1 July 2020).
- Regulation of the Minister of Health from December 7, 2017 on the Quality of Water Intended for Human Consumption, Dz. U. 2017, 2294. Available online: http://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20170002294/O/D20172294.pdf (accessed on 1 July 2020). (In Polish)
- Pruden, A.; Arabi, M.; Storteboom, H.N. Correlation between upstream human activities and riverine antibiotic resistance genes. Environ. Sci. Technol. 2012, 46, 11541–11549. [Google Scholar] [CrossRef]
- Leung, H.W.; Jin, L.; Wei, S.; Tsui, M.M.P.; Zhou, B.; Jiao, L.; Cheung, P.C.; Chun, Y.K.; Murphy, M.B.; Lam, P.K.S. Pharmaceuticals in tap water: Human health risk assessment and proposed monitoring framework in China. Environ. Health Perspect. 2013, 121, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, N.; Wang, B.; Zhao, Q.; Fang, H.; Fu, C.; Tang, C.; Jiang, F.; Zhou, Y.; Chen, Y.; et al. Antibiotics in Drinking Water in Shanghai and Their Contribution to Antibiotic Exposure of School Children. Environ. Sci. Technol. 2016, 50, 2692–2699. [Google Scholar] [CrossRef] [PubMed]
- Yiruhan; Wang, Q.J.; Mo, C.H.; Li, Y.W.; Gao, P.; Tai, Y.P.; Zhang, Y.; Ruan, Z.L.; Xu, J.W. Determination of four fluoroquinolone antibiotics in tap water in Guangzhou and Macao. Environ. Pollut. 2010, 158, 2350–2358. [Google Scholar] [CrossRef]
- Segura, P.A.; François, M.; Gagnon, C.; Sauvé, S. Review of the occurrence of anti-infectives in contaminated wastewaters and natural and drinking waters. Environ. Health Perspect. 2009, 117, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Jiang, T.; Zhang, S.; Yu, X. Exposure to mutagenic disinfection byproducts leads to increase of antibiotic resistance in Pseudomonas aeruginosa. Environ. Sci. Technol. 2014, 48, 8188–8195. [Google Scholar] [CrossRef]
- Farkas, A.; Butiuc-Keul, A.; Ciatarâş, D.; Neamţu, C.; Crǎciunaş, C.; Podar, D.; Drǎgan-Bularda, M. Microbiological contamination and resistance genes in biofilms occurring during the drinking water treatment process. Sci. Total Environ. 2013, 443, 932–938. [Google Scholar] [CrossRef]
- Szekeres, E.; Chiriac, C.M.; Baricz, A.; Szőke-Nagy, T.; Lung, I.; Soran, M.L.; Rudi, K.; Dragos, N.; Coman, C. Investigating antibiotics, antibiotic resistance genes, and microbial contaminants in groundwater in relation to the proximity of urban areas. Environ. Pollut. 2018, 236, 734–744. [Google Scholar] [CrossRef]
- Okitsu, N.; Kaieda, S.; Yano, H.; Nakano, R.; Hosaka, Y.; Okamoto, R.; Kobayashi, T.; Inoue, M. Characterization of ermB gene transposition by Tn1545 and Tn917 in macrolide-resistant Streptococcus pneumoniae isolates. J. Clin. Microbiol. 2005, 43, 168–173. [Google Scholar] [CrossRef]
- Ibáñez de Aldecoa, A.L.; Zafra, O.; González-Pastor, J.E. Mechanisms and regulation of extracellular DNA release and its biological roles in microbial communities. Front. Microbiol. 2017, 8, 1–19. [Google Scholar] [CrossRef]
- Mao, D.; Luo, Y.; Mathieu, J.; Wang, Q.; Feng, L.; Mu, Q.; Feng, C.; Alvarez, P.J.J. Persistence of extracellular DNA in river sediment facilitates antibiotic resistance gene propagation. Environ. Sci. Technol. 2014, 48, 71–78. [Google Scholar] [CrossRef]
- Lorenz, M.G.; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiol. Rev. 1994, 58, 563–602. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC372978/ (accessed on 1 July 2020).
- Davis, M.A.; Besser, T.E.; Orfe, L.H.; Baker, K.N.K.; Lanier, A.S.; Broschat, S.L.; New, D.; Call, D.R. Genotypic-phenotypic discrepancies between antibiotic resistance characteristics of Escherichia coli isolates from calves in management settings with high and low antibiotic use. Appl. Environ. Microbiol. 2011, 77, 3293–3299. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Zhou, Q.; Wu, Q.; Gu, Q.; Sun, M.; Zhang, J. Spatiotemporal changes in bacterial community and microbial activity in a full-scale drinking water treatment plant. Sci. Total Environ. 2018, 625, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Nescerecka, A.; Juhna, T.; Hammes, F. Identifying the underlying causes of biological instability in a full-scale drinking water supply system. Water Res. 2018, 135, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Perrin, Y.; Bouchon, D.; Delafont, V.; Moulin, L.; Héchard, Y. Microbiome of drinking water: A full-scale spatio-temporal study to monitor water quality in the Paris distribution system. Water Res. 2019, 149, 375–385. [Google Scholar] [CrossRef]
- Shrivastava, R.; Upreti, R.K.; Jain, S.R.; Prasad, K.N.; Seth, P.K.; Chaturvedi, U.C. Suboptimal chlorine treatment of drinking water leads to selection of multidrug-resistant Pseudomonas aeruginosa. Ecotoxicol. Environ. Saf. 2004, 58, 277–283. [Google Scholar] [CrossRef]
- Asghari, F.B.; Pakdel, M.; Mohammadi, A.A.; Yousefi, M. Spatial and temporal variation of physicochemical and microbial quality of drinking water for the distribution network in Maku, Iran. Desalin. Water Treat. 2019, 142, 82–89. [Google Scholar] [CrossRef]
- Henne, K.; Kahlisch, L.; Höfle, M.G.; Brettar, I. Seasonal dynamics of bacterial community structure and composition in cold and hot drinking water derived from surface water reservoirs. Water Res. 2013, 47, 5614–5630. [Google Scholar] [CrossRef]
- Girolamini, L.; Lizzadro, J.; Mazzotta, M.; Iervolino, M.; Dormi, A.; Cristino, S. Different trends in microbial contamination between two types of microfiltered water dispensers: From risk analysis to consumer health preservation. Int. J. Environ. Res. Public Health 2019, 16, 272. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Chen, J.; Wang, F.; Qi, W.; Li, Y. Impact of disinfectant on bacterial antibiotic resistance transfer between biofilm and tap water in a simulated distribution network. Environ. Pollut. 2019, 246, 131–140. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Agar Medium | Antibiotic and Concentration | Purpose |
|---|---|---|
| R2A | - | psychrophilic HPC |
| R2A + AML | amoxicillin 8 mg/L | % of bacteria resistant to amoxicillin |
| R2A + CIP | ciprofloxacin 2 mg/L | % of bacteria resistant to ciprofloxacin |
| R2A + CAZ | ceftazidime 8 mg/L | % of bacteria resistant to ceftazidime |
| R2A + TE | tetracycline 16 mg/L | % of bacteria resistant to tetracycline |
| Physical–Chemical Parameter | Range | Medium |
|---|---|---|
| temperature (°C) | 2.1–22.5 | 14.7 |
| turbidity (NTU) | <0.5 | - |
| color (mg/L Pt) | <2 | - |
| pH | 7.50–7.85 | 7.64 |
| conductivity (µS/cm) | 409–735 | 602 |
| free Cl2 (mg/L) | 0.00–0.31 | 0.10 |
| total Cl2 (mg/L) | 0.10–0.49 | 0.27 |
| chlorites and chlorates (mg/L) | 0.05–0.26 | 0.16 |
| nitrates (mg/L) | 1.70–27.00 | 7.45 |
| aluminum (µg/L) | 10–25 | 16.85 |
| Gene | Gene Classification, Resistance Target or Mechanisms | Summer | Winter | Total in Summer | Total in Winter | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NG1S | NG2S | NG3S | NG4S | MD1S | MD2S | MD3S | MD4S | NG1W | NG2W | NG3W | NG4W | MD1W | MD2W | MD3W | MD4W | ||||
| blaTEM | β-lactams | - | - | - | - | + | + | + | + | - | - | - | - | + | - | - | - | 4 | 1 |
| blaSHV | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 | |
| blaCTX-M | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 | |
| blaKPC | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 | |
| blaNDM | + | - | - | - | - | - | + | - | - | - | - | - | - | - | - | + | 2 | 1 | |
| blaOXA | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 | |
| blaOXA-48 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 | |
| ampC | - | - | - | - | + | + | - | + | - | - | - | - | - | - | - | - | 3 | 0 | |
| mecA | methicillin | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| qnrA | (fluoro)quinolones | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| qnrB | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 | |
| qnrS | + | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 0 | |
| oqxB | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | 0 | 1 | |
| tetA | tetracyclines | - | - | + | - | - | - | - | + | - | - | + | + | - | - | - | - | 2 | 2 |
| tetK | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - | - | 0 | 1 | |
| tetL | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 | |
| tetW | - | - | - | - | - | - | - | - | - | + | - | - | - | + | + | - | 0 | 3 | |
| sulI | sulphonamides | - | + | + | + | - | - | - | + | - | + | - | + | - | - | + | + | 4 | 4 |
| sulII | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 | |
| ermA | erythromycins | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| ermB | - | - | - | + | - | + | - | - | + | + | + | + | + | + | + | + | 2 | 8 | |
| vanA | vancomycin | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| mcr-1 | colistin | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| mexA | efflux | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| floR | florfenicol | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0 | 0 |
| qacEΔ1 | quaternary ammonium compounds | - | - | + | + | - | + | - | - | - | + | + | + | + | + | - | - | 3 | 5 |
| qacH | + | - | + | - | + | + | + | - | - | - | - | + | + | + | - | + | 5 | 4 | |
| tolA | transmembrane activity | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 |
| intI1 | class 1 integron | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | 8 | 8 |
| tnpA | transposon | - | + | + | + | + | + | + | - | - | - | - | - | - | - | - | - | 6 | 0 |
| total | 4 | 4 | 8 | 5 | 6 | 7 | 5 | 5 | 2 | 6 | 4 | 6 | 5 | 6 | 4 | 5 | 44 | 38 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siedlecka, A.; Wolf-Baca, M.; Piekarska, K. Spatiotemporal Changes of Antibiotic Resistance and Bacterial Communities in Drinking Water Distribution System in Wrocław, Poland. Water 2020, 12, 2601. https://doi.org/10.3390/w12092601
Siedlecka A, Wolf-Baca M, Piekarska K. Spatiotemporal Changes of Antibiotic Resistance and Bacterial Communities in Drinking Water Distribution System in Wrocław, Poland. Water. 2020; 12(9):2601. https://doi.org/10.3390/w12092601
Chicago/Turabian StyleSiedlecka, Agata, Mirela Wolf-Baca, and Katarzyna Piekarska. 2020. "Spatiotemporal Changes of Antibiotic Resistance and Bacterial Communities in Drinking Water Distribution System in Wrocław, Poland" Water 12, no. 9: 2601. https://doi.org/10.3390/w12092601
APA StyleSiedlecka, A., Wolf-Baca, M., & Piekarska, K. (2020). Spatiotemporal Changes of Antibiotic Resistance and Bacterial Communities in Drinking Water Distribution System in Wrocław, Poland. Water, 12(9), 2601. https://doi.org/10.3390/w12092601