Differences in the Vertical Distribution of Two Cladoceran Species in the Nakdong River Estuary, South Korea


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Monitoring Strategy
2.3. Data Analysis
3. Results
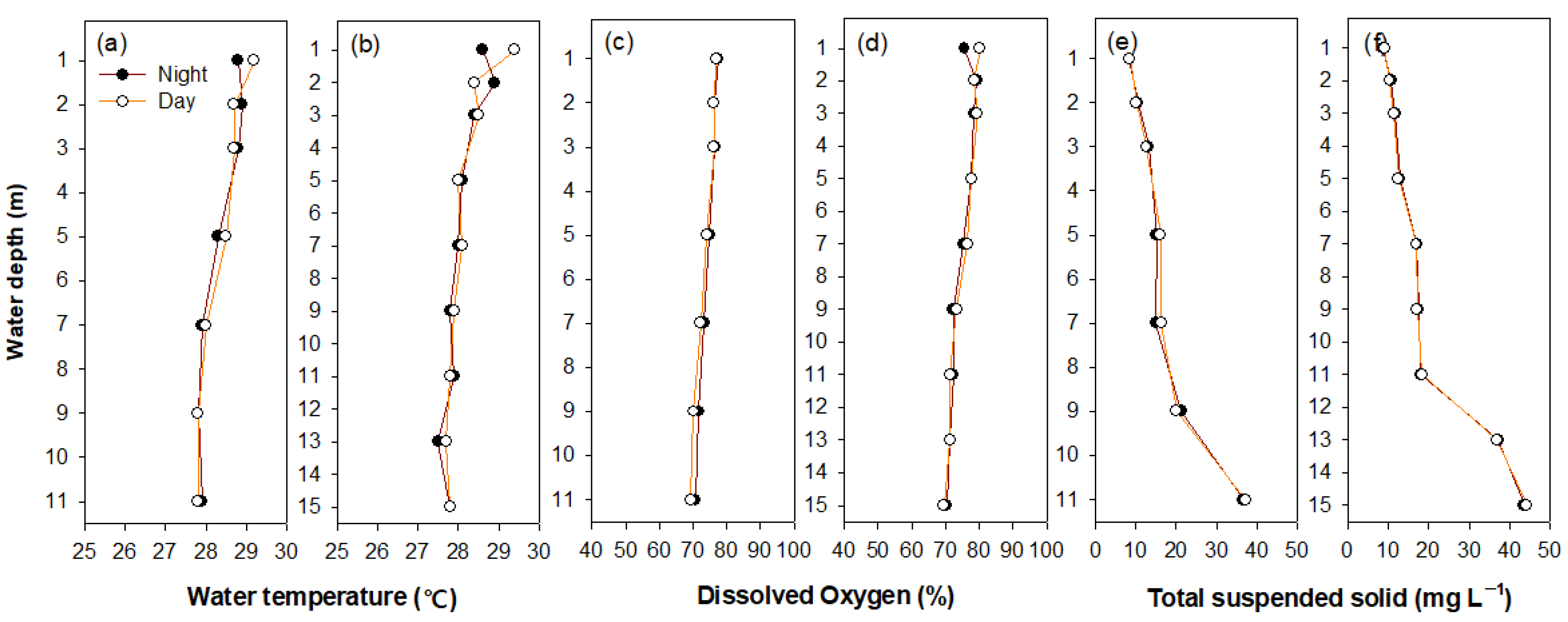
3.1. Environmental Variables and Cladocera Distributions
3.2. Vertical Distribution of Two Cladocera Species between Day and Night in Summer
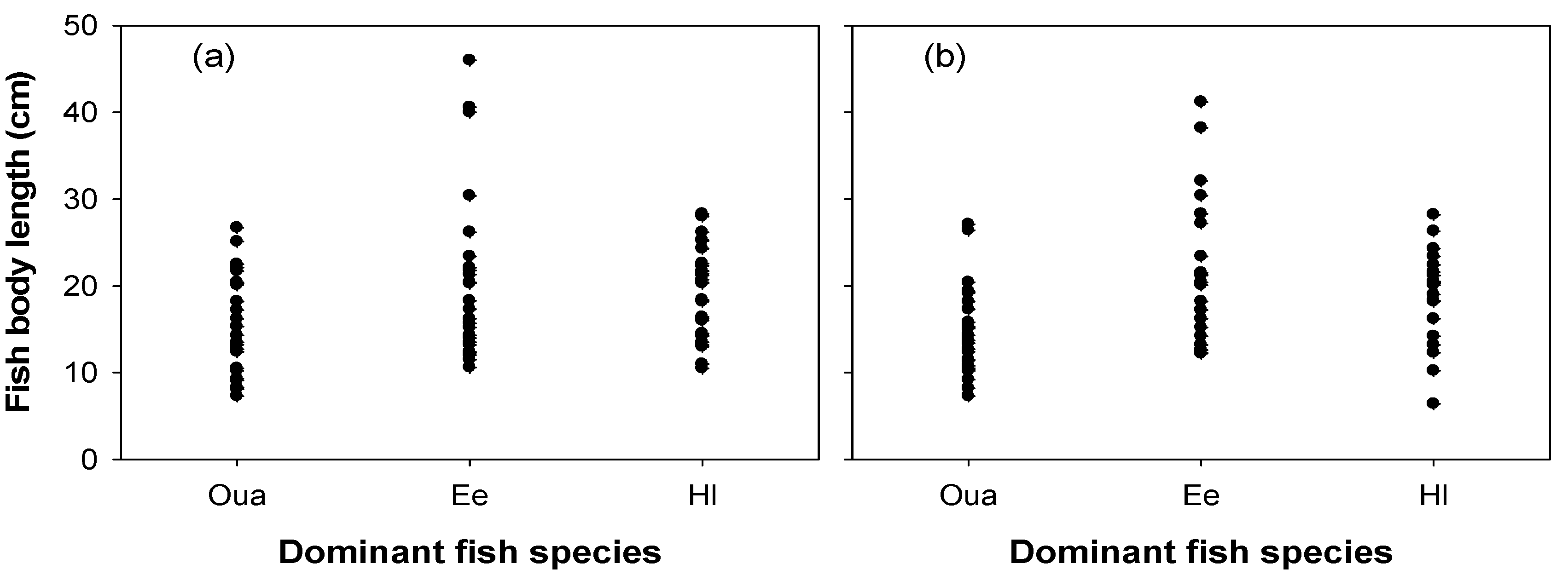
3.3. Fish Distribution in the Nakdong River Estuary
4. Discussion
4.1. Seasonal Environmental Variables between Water Layers
4.2. Vertical Distribution of Daphnia obtusa Due to Fish Predation
4.3. Behavioral Strategies by Body Size Class of the Two Cladoceran Species
Author Contributions
Funding
Conflicts of Interest
References
- Curio, E. The Ethology of Predation; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Dodson, S. Predator-induced reaction norms. Bioscience 1989, 39, 447–452. [Google Scholar] [CrossRef]
- Lima, S.L. Stress and decision making under the risk of predation: Recent developments from behavioral, reproductive, and ecological perspectives. In Advances in the Study of Behavior; Slater, P.J.B., Møller, A.P., Manfred, M., Eds.; Academic Press: London, UK, 1998; pp. 215–290. [Google Scholar]
- Pasternak, A.F.; Mikheev, V.N.; Wanzenböck, J. How plankton copepods avoid fish predation: From individual responses to variations of the life cycle. J. Ichthyol. 2006, 46, S220–S226. [Google Scholar] [CrossRef]
- Pettersson, L.B.; Nilsson, P.A.; Brönmark, C. Predator recognition and defence strategies in crucian carp, Carassius carassius. Oikos 2000, 88, 200–212. [Google Scholar] [CrossRef]
- Ángeles Esteban, M. An overview of the immunological defenses in fish skin. ISRN immunology 2012, 2012. [Google Scholar] [CrossRef]
- Luttbeg, B.; Sih, A. Predator and prey habitat selection games: The effects of how prey balance foraging and predation risk. Isr. J. Zool. 2004, 50, 233–254. [Google Scholar] [CrossRef][Green Version]
- Sagrario, M.D.L.Á.G.; Balseiro, E. Indirect enhancement of large zooplankton by consumption of predacious macroinvertebrates by littoral fish. Arch. Hydrobiol. 2003, 158, 551–574. [Google Scholar] [CrossRef]
- Tollrian, R.; Harvell, C.D. The Ecology and Evolution of Inducible Defenses; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar]
- Boeing, W.J.; Ramcharan, C.W.; Riessen, H.P. Clonal variation in depth distribution of Daphnia pulex in response to predator kairomones. Arch. Hydrobiol. 2006, 166, 241–260. [Google Scholar] [CrossRef]
- Ringelberg, J. The photobehaviour of Daphnia spp. as a model to explain diel vertical migration in zooplankton. Biol. Rev. 1999, 74, 397–423. [Google Scholar] [CrossRef]
- Walls, M.; Ketola, M. Effects of predator-induced spines on individual fitness in Daphnia pulex. Limnol. Oceanogr. 1989, 34, 390–396. [Google Scholar] [CrossRef]
- Haupt, F.; Stockenreiter, M.; Baumgartner, M.; Boersma, M.; Stibor, H. Daphnia diel vertical migration: Implications beyond zooplankton. J. Plankton Res. 2009, 31, 515–524. [Google Scholar] [CrossRef]
- Loose, C.J. Lack of endogenous rhythmicity in Daphnia diel vertical migration. Limnol. Oceanogr. 1993, 38, 1837–1841. [Google Scholar] [CrossRef]
- Ringelberg, J. A mechanism of predator-mediated induction of diel vertical migration in Daphnia hyalina. J. Plankton Res. 1991, 13, 83–89. [Google Scholar] [CrossRef]
- Battes, K.P.; Momeu, L. Diel vertical distribution of planktonic microcrustaceans (Crustacea: Cladocera, Copepoda) in a natural shallow lake from Transylvania, Romania. J. Limnol. 2014, 73, 34–44. [Google Scholar] [CrossRef]
- Jung, J.; Hojnowski, C.; Jenkins, H.; Ortiz, A.; Brinkley, C.; Cadish, L.; Evans, A.; Kissinger, P.; Ordal, L.; Osipova, S.; et al. Diel vertical migration of zooplankton in Lake Baikal and its relationship to body size. In Ecosystems and Natural Resources of Mountain Regions; Smirnov, A.I., Izmest’eva, L.R., Eds.; Nauka: Novosibirsk, Rusia, 2004; pp. 131–140. [Google Scholar]
- Semyalo, R.; Nattabi, J.K.; Larsson, P. Diel vertical migration of zooplankton in a eutrophic bay of Lake Victoria. Hydrobiologia 2009, 635, 383–394. [Google Scholar] [CrossRef]
- Brancelj, A.; Blejec, A. Diurnal vertical migration of Daphnia hyalina Leydig, 1860 (Crustacea: Cladocera) in Lake Bled (Slovenia) in relation to temperature and predation. Hydrobiologia 1994, 284, 125–136. [Google Scholar] [CrossRef]
- Degefu, F.; Schagerl, M. Zooplankton abundance, species composition and ecology of tropical high-mountain crater Lake Wonchi, Ethiopia. J. limnol. 2015, 74, 324–334. [Google Scholar] [CrossRef]
- Easton, J.; Gophen, M. Diel variation in the vertical distribution of fish and plankton in Lake Kinneret: A 24-h study of ecological overlap. Hydrobiologia 2003, 491, 91–100. [Google Scholar] [CrossRef]
- Johnsen, G.J.; Jakobsen, P.J. The effect of food limitation on vertical migration in Daphnia longispina. Limnol. Oceanogr. 1987, 32, 873–880. [Google Scholar] [CrossRef]
- Ringelberg, J.; Van Gool, E. On the combined analysis of proximate and ultimate aspects in diel vertical migration (DVM) research. Hydrobiologia 2003, 491, 85–90. [Google Scholar] [CrossRef]
- De Bernardi, R.; Giussani, G.; Manca, M. Cladocera: Predators and prey. In Cladocera; Forró, L., Frey, D.G., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 225–243. [Google Scholar]
- Ramos-Jiliberto, R.O.D.R.I.G.O.; Zúñiga, L.R. Depth-selection patterns and diel vertical migration of Daphnia ambigua (Crustacea: Cladocera) in lake El Plateado. Rev. Chil. Hist. Nat. 2001, 74, 573–585. [Google Scholar] [CrossRef]
- Horppila, J.; Malinen, T.; Nurminen, L.; Tallberg, P.; Vinni, M. A metalimnetic oxygen minimum indirectly contributing to the low biomass of cladocerans in Lake Hiidenvesi–a diurnal study on the refuge effect. Hydrobiologia 2000, 436, 81–90. [Google Scholar] [CrossRef]
- Ha, K.; Cho, E.A.; Kim, H.W.; Joo, G.J. Microcystis bloom formation in the lower Nakdong River, South Korea: Importance of hydrodynamics and nutrient loading. Mar. Freshw. Res. 1999, 50, 89–94. [Google Scholar] [CrossRef]
- Kim, H.W.; Joo, G.J.; Walz, N. Zooplankton Dynamics in the Hyper-Eutrophic Nakdong River System (Korea) Regulated by an Estuary Dam and Side Channels. Internat. Rev. Hydrobiol. 2001, 86, 127–143. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; Kim, H.W.; Chang, K.H.; Joo, G.J. Inter-annual variability of a zooplankton community: The importance of summer concentrated rainfall in a regulated river ecosystem. J. Ecol. Environ. 2011, 34, 49–58. [Google Scholar] [CrossRef][Green Version]
- Jeong, K.S.; Kim, D.K.; Joo, G.J. Delayed influence of dam storage and discharge on the determination of seasonal proliferations of Microcystis aeruginosa and Stephanodiscus hantzschii in a regulated river system of the lower Nakdong River (South Korea). Water Res. 2007, 41, 1269–1279. [Google Scholar] [CrossRef]
- An, J.H.; Lee, K.H. Correlation and hysteresis analysis of air-water temperature in four rivers: Preliminary study for water temperature prediction. J. Environ. Policy 2013, 12, 17–32. [Google Scholar]
- Wetzel, R.G.; Likens, G.E. Composition and biomass of phytoplankton. In Limnological Analyses; Springer: New York, NY, USA, 2000; pp. 147–174. [Google Scholar]
- Haney, J.F.; Hall, D.J. Sugar-coated Daphnia: A preservation technique for Cladocera 1. Limnol. Oceanogr. 1973, 18, 331–333. [Google Scholar] [CrossRef]
- Mizuno, T.; Takahashi, E. An Illustration Guide to Freshwater Zooplankton in Japan; Tokai University: Tokyo, Japan, 1999. [Google Scholar]
- Kim, I.S.; Choi, Y.; Lee, C.L.; Lee, Y.J.; Kim, B.J.; Kim, J.H. Illustrated Book of Korean Fishes; Kyohak Publishing: Seoul, Korea, 2005. [Google Scholar]
- Javahery, S.; Nekoubin, H.; Moradlu, A.H. Effect of anaesthesia with clove oil in fish. Fish Physiol. Biochem. 2012, 38, 1545–1552. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Yoon, J.D.; Jang, M.H.; Jo, H.B.; Jeong, K.S.; Kim, G.Y.; Joo, G.J. Changes of fish assemblages after construction of an estuary barrage in the lower Nakdong River, South Korea. Limnology 2016, 17, 183–197. [Google Scholar] [CrossRef]
- Read, J.S.; Hamilton, D.P.; Jones, I.D.; Muraoka, K.; Winslow, L.A.; Kroiss, R.; Wu, C.H.; Gaiser, E. Derivation of lake mixing and stratification indices from high-resolution lake buoy data. Environ. Model. Softw. 2011, 26, 1325–1336. [Google Scholar] [CrossRef]
- Kraemer, B.M.; Anneville, O.; Chandra, S.; Dix, M.; Kuusisto, E.; Livingstone, D.M.; Rimmer, A.; Schladow, S.G.; Silow, E.; Sitoki, L.M.; et al. Morphometry and average temperature affect lake stratification responses to climate change. Geophys. Res. Lett. 2015, 42, 4981–4988. [Google Scholar] [CrossRef]
- Berger, S.A.; Diehl, S.; Stibor, H.; Trommer, G.; Ruhenstroth, M. Water temperature and stratification depth independently shift cardinal events during plankton spring succession. Glob. Chang. Biol. 2010, 16, 1954–1965. [Google Scholar] [CrossRef]
- Hassan, H.; Aramaki, T.; Hanaki, K.; Matsuo, T.; Wilby, R. Lake stratification and temperature profiles simulated using downscaled GCM output. Water Sci. Technol. 1998, 38, 217–226. [Google Scholar] [CrossRef]
- Lampert, W. Ultimate causes of diel vertical migration of zooplankton: New evidence for the predator-avoidance hypothesis. Arch. Hydrobiol Beih. Ergebn. Limnol. 1993, 39, 79–88. [Google Scholar]
- Von Elert, E.; Loose, C.J. Predator-induced diel vertical migration in Daphnia: Enrichment and preliminary chemical characterization of a kairomone exuded by fish. J. Chem. Ecol. 1996, 22, 885–895. [Google Scholar] [CrossRef]
- Decker, M.B.; Breitburg, D.L.; Purcell, J.E. Effects of low dissolved oxygen on zooplankton predation by the ctenophore Mnemiopsis leidyi. Mar. Ecol. Prog. Ser. 2004, 280, 163–172. [Google Scholar] [CrossRef]
- Horppila, J.; Eloranta, P.; Liljendahl-Nurminen, A.; Niemistö, J.; Pekcan-Hekim, Z. Refuge availability and sequence of predators determine the seasonal succession of crustacean zooplankton in a clay-turbid lake. Aquat. Ecol. 2009, 43, 91–103. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; La, G.H.; Joo, G.J.; Kim, J.; Kim, S.J.; Kim, S.H.; Chung, Y.H.; Kim, D.K.; Do, Y.; et al. Spatio-temporal distribution of Diaphanosoma brachyurum (Cladocera: Sididae) in freshwater reservoir ecosystems: Importance of maximum water depth and macrophyte beds for avoidance of fish predation. J. Limnol. 2015, 74, 403–413. [Google Scholar] [CrossRef]
- Regular, P.M.; Hedd, A.; Montevecchi, W.A. Fishing in the dark: A pursuit-diving seabird modifies foraging behaviour in response to nocturnal light levels. PLoS ONE 2011, 6, e26763. [Google Scholar] [CrossRef]
- Seitz, F.; Bundschuh, M.; Rosenfeldt, R.R.; Schulz, R. Nanoparticle toxicity in Daphnia magna reproduction studies: The importance of test design. Aquat. Toxicol. 2013, 126, 163–168. [Google Scholar] [CrossRef]
- Gaevsky, N.A.; Zotina, T.A.; Gorbaneva, T.B. Vertical structure and photosynthetic activity of Lake Shira phytoplankton. Aquat. Ecol. 2002, 36, 165–178. [Google Scholar] [CrossRef]
- Lampert, W. The adaptive significance of diel vertical migration of zooplankton. Funct. Ecol. 1989, 3, 21–27. [Google Scholar] [CrossRef]
- Pearre, S. Eat and run? The hunger/satiation hypothesis in vertical migration: History, evidence and consequences. Biol. Rev. 2003, 78, 1–79. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Mills, E.L.; Idrisi, N.; Michener, R. Stable isotopes of nitrogen and carbon in an aquatic food web recently invaded by (Dreissena polymorpha)(Pallas). Can. J. Fish. Aquat. Sci. 1996, 53, 1445–1450. [Google Scholar] [CrossRef][Green Version]
- Fry, B.; Mumford, P.L.; Tam, F.; Fox, D.D.; Warren, G.L.; Havens, K.E.; Steinman, A.D. Trophic position and individual feeding histories of fish from Lake Okeechobee, Florida. Can. J. Fish. Aquat. Sci. 1999, 56, 590–600. [Google Scholar] [CrossRef]
- Ibrahim, A.A.; Huntingford, F.A. The role of visual cues in prey selection in three-spined sticklebacks (Gasterosteus aculeatus). Ethology 1989, 81, 265–272. [Google Scholar] [CrossRef]
- Thys, I.; Hoffmann, L. Diverse responses of planktonic crustaceans to fish predation by shifts in depth selection and size at maturity. Hydrobiologia 2005, 551, 87–98. [Google Scholar] [CrossRef]
- Wojtal-Frankiewicz, A.; Frankiewicz, P.; Jurczak, T.; Grennan, J.; McCarthy, T.K. Comparison of fish and phantom midge influence on cladocerans diel vertical migration in a dual basin lake. Aquat. Ecol. 2010, 44, 243–254. [Google Scholar] [CrossRef]
- Mehner, T.; Hölker, F.; Kasprzak, P. Spatial and temporal heterogeneity of trophic variables in a deep lake as reflected by repeated singular samplings. Oikos 2005, 108, 401–409. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Tollrian, R. Prey swarming: Which predators become confused and why? Anim. Behav. 2007, 74, 387–393. [Google Scholar] [CrossRef]
- Uusitalo, L.; Horppila, J.; Eloranta, P.; Liljendahl-Nurminen, A.; Malinen, T.; Salonen, M.; Vinni, M. Leptodora kindti and flexible foraging behaviour of fish–factors behind the delayed biomass peak of cladocerans in Lake Hiidenvesi. Internat. Rev. Hydrobiol. 2003, 88, 34–48. [Google Scholar] [CrossRef]
- Chang, K.H.; Hanazato, T. Seasonal and spatial distribution of two Bosmina species (B. longirostris and B. fatalis) in Lake Suwa, Japan: Its relation to the predator Leptodora. Limnology 2003, 4, 0047–0052. [Google Scholar] [CrossRef]
- Labaj, A.L.; Korosi, J.B.; Kurek, J.; Jeziorski, A.; Keller, W.; Smol, J.P. Response of Bosmina size structure to the acidification and recovery of lakes near Sudbury. Can. J. Limnol. 2016, 75, 22–29. [Google Scholar] [CrossRef][Green Version]
- Amsinck, S.L.; Strzelczak, A.; Bjerring, R.; Landkildehus, F.; Lauridsen, T.L.; Christoffersen, K.; Jeppesen, E. Lake depth rather than fish planktivory determines cladoceran community structure in Faroese lakes–evidence from contemporary data and sediments. Freshw. Biol. 2006, 51, 2124–2142. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Variables | Site 1 | Site 2 | Site 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| U | M | B | U | M | B | U | M | B | ||
| Feb. | Depth | 1 | 3 | 8 | 1 | 1 | 3 | 1 | 5 | 12 |
| WT | 5.3 | 5.0 | 5.1 | 5.0 | 5.6 | 5.3 | 5.5 | 5.3 | 5.1 | |
| DO | 129.7 | 123.9 | 123.9 | 129.0 | 124.6 | 123.7 | 136.0 | 125.7 | 124.6 | |
| pH | 6.20 | 5.84 | 5.91 | 6.18 | 6.88 | 6.45 | 8.52 | 6.28 | 6.94 | |
| Chl. a | 22.4 | 13.3 | 14.7 | 12.6 | 10.5 | 8.4 | 8.4 | 10.5 | 9.8 | |
| TSS | 10.0 | 11.0 | 15.0 | 8.0 | 8.2 | 11.0 | 11.0 | 12.4 | 19.0 | |
| May | Depth | 1 | 5 | 9 | 1 | 2 | 3 | 1 | 7 | 13 |
| WT | 18.0 | 18.3 | 18.1 | 18.2 | 18.3 | 17.9 | 18.0 | 18.7 | 18.1 | |
| DO | 87.3 | 89.9 | 89.5 | 90.9 | 94.1 | 91.9 | 87.3 | 90.6 | 90.8 | |
| pH | 5.58 | 5.43 | 5.46 | 5.41 | 5.45 | 5.42 | 5.58 | 5.87 | 6.05 | |
| Chl. a | 12.0 | 16.0 | 16.0 | 12.0 | 16.0 | 16.0 | 12.0 | 12.0 | 16.0 | |
| TSS | 4.4 | 4.4 | 5.6 | 5.4 | 5.8 | 5.9 | 7.1 | 6.9 | 7.6 | |
| Aug. | Depth | 1 | 5 | 11 | 1 | 3 | 5 | 1 | 7 | 15 |
| WT | 29.1 | 28.0 | 27.8 | 29.1 | 28.9 | 28.0 | 29.2 | 28.3 | 28.0 | |
| DO | 74.5 | 72.9 | 70.7 | 75.0 | 73.5 | 73.2 | 77.2 | 74.8 | 71.4 | |
| pH | 7.30 | 7.14 | 7.30 | 7.22 | 7.10 | 6.55 | 7.66 | 7.68 | 7.60 | |
| Chl. a | 40.0 | 37.0 | 34.5 | 32.2 | 37.0 | 30.5 | 56.0 | 49.7 | 40.8 | |
| TSS | 9.6 | 16.0 | 37.2 | 9.6 | 18.8 | 34.4 | 8.0 | 16.8 | 43.6 | |
| Nov. | Depth | 1 | 5 | 9 | 1 | 2 | 4 | 1 | 7 | 13 |
| WT | 12.9 | 12.1 | 11.6 | 12.9 | 12.1 | 11.7 | 12.9 | 12.2 | 10.8 | |
| DO | 93.2 | 94.0 | 94.9 | 92.8 | 93.2 | 95.0 | 95.6 | 93.0 | 96.1 | |
| pH | 7.37 | 7.84 | 7.97 | 8.07 | 8.12 | 8.09 | 7.35 | 7.55 | 7.41 | |
| Chl. a | 5.5 | 4.1 | 5.3 | 2.9 | 2.9 | 3.0 | 5.4 | 4.1 | 4.8 | |
| TSS | 4.5 | 6.3 | 6.7 | 5.3 | 5.5 | 5.7 | 10.8 | 8.8 | 7.9 | |
| Species | Site | Variance | df | F | p |
|---|---|---|---|---|---|
| B. longirostris | Site 1 | Time | 1 | 1.543 | 0.098 |
| Sampling point | 4 | 0.432 | 1.753 | ||
| Site 3 | Time | 1 | 1.632 | 0.086 | |
| Sampling point | 4 | 0.642 | 1.629 | ||
| D. obtusa | Site 1 | Time | 1 | 14.24 | <0.01 |
| Sampling point | 4 | 0.141 | 0.943 | ||
| Site 3 | Time | 1 | 12.41 | <0.01 | |
| Sampling point | 4 | 0.150 | 0.911 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-K.; Choi, J.-Y. Differences in the Vertical Distribution of Two Cladoceran Species in the Nakdong River Estuary, South Korea. Water 2020, 12, 2154. https://doi.org/10.3390/w12082154
Kim S-K, Choi J-Y. Differences in the Vertical Distribution of Two Cladoceran Species in the Nakdong River Estuary, South Korea. Water. 2020; 12(8):2154. https://doi.org/10.3390/w12082154
Chicago/Turabian StyleKim, Seong-Ki, and Jong-Yun Choi. 2020. "Differences in the Vertical Distribution of Two Cladoceran Species in the Nakdong River Estuary, South Korea" Water 12, no. 8: 2154. https://doi.org/10.3390/w12082154
APA StyleKim, S.-K., & Choi, J.-Y. (2020). Differences in the Vertical Distribution of Two Cladoceran Species in the Nakdong River Estuary, South Korea. Water, 12(8), 2154. https://doi.org/10.3390/w12082154