The Influence of Aquaculture Effluents on the Prevalence and Biocides Resistance of Opportunistic Pseudomonas fluorescens Bacteria in the Drwęca River Protected under the Natura 2000 Network


Abstract
1. Introduction
2. Materials and Methods
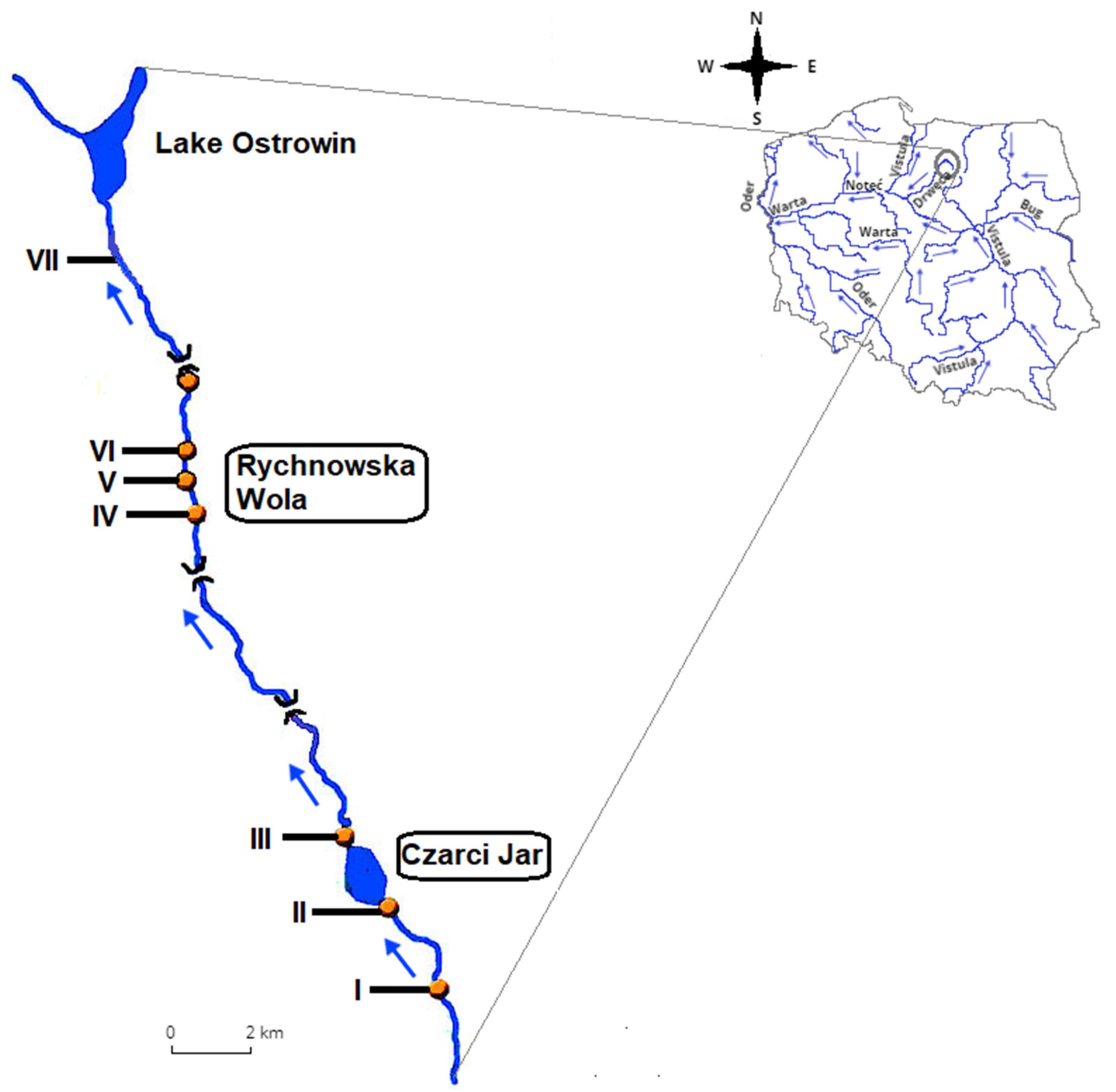
2.1. Study Area and Sampling Sites
- Sampling site I—control site located around 2 km downstream from the river source;
- Sampling site II—water intake for fish farm No. 1 in Czarci Jar with an estimated annual output 10 tons of trout broodstock;
- Sampling site III—directly downstream from the effluent discharge point of fish farm No. 1 in Czarci Jar;
- Sampling site IV—water intake for fish farms No. 2 and 3 in Rychnowska Wola, situated at a distance of 12 km from sampling site III;
- Sampling site V—directly downstream from the effluent discharge point of fish farm No. 2 in Rychnowska Wola with an estimated annual output of 50 tons market-sized trout from purchased fry;
- Sampling site VI—directly downstream from the effluent discharge point of fish farm No. 3 in Rychnowska Wola with an estimated annual output of 30 tons market-sized trout from purchased fry;
- Sampling site VII—situated at a distance of around 6 km downstream from fish farms No. 2 and 3.
2.2. Microbiological Methods
2.2.1. Total Pseudomonas fluorescens Population (TPFP) in the Drwęca River
2.2.2. Counts of Opportunistic Pathogens P. fluorescens (OPPF) in the Drwęca River
2.2.3. Antibiotic Resistance of Opportunistic Pathogens P. fluorescens (OPPF)
2.2.4. Minimum Inhibitory Concentrations (MIC) of Antibiotics
2.2.5. Resistance of Opportunistic Pathogens P. fluorescens (OPPF) to Disinfectants
2.3. Statistical Analysis
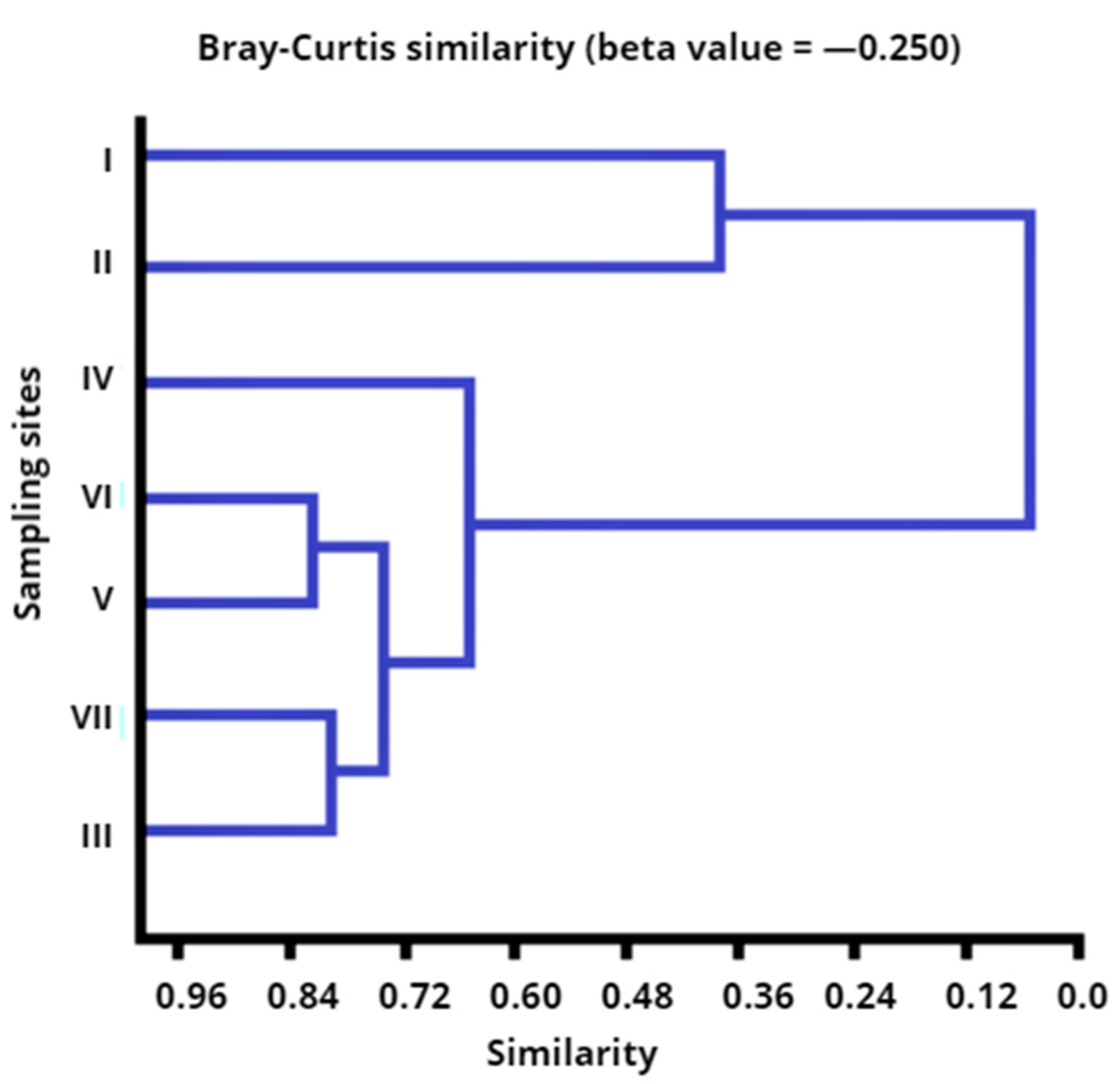
3. Results
3.1. Counts of Opportunistic Pathogens P. fluorescens (OPPF) in the Drwęca River
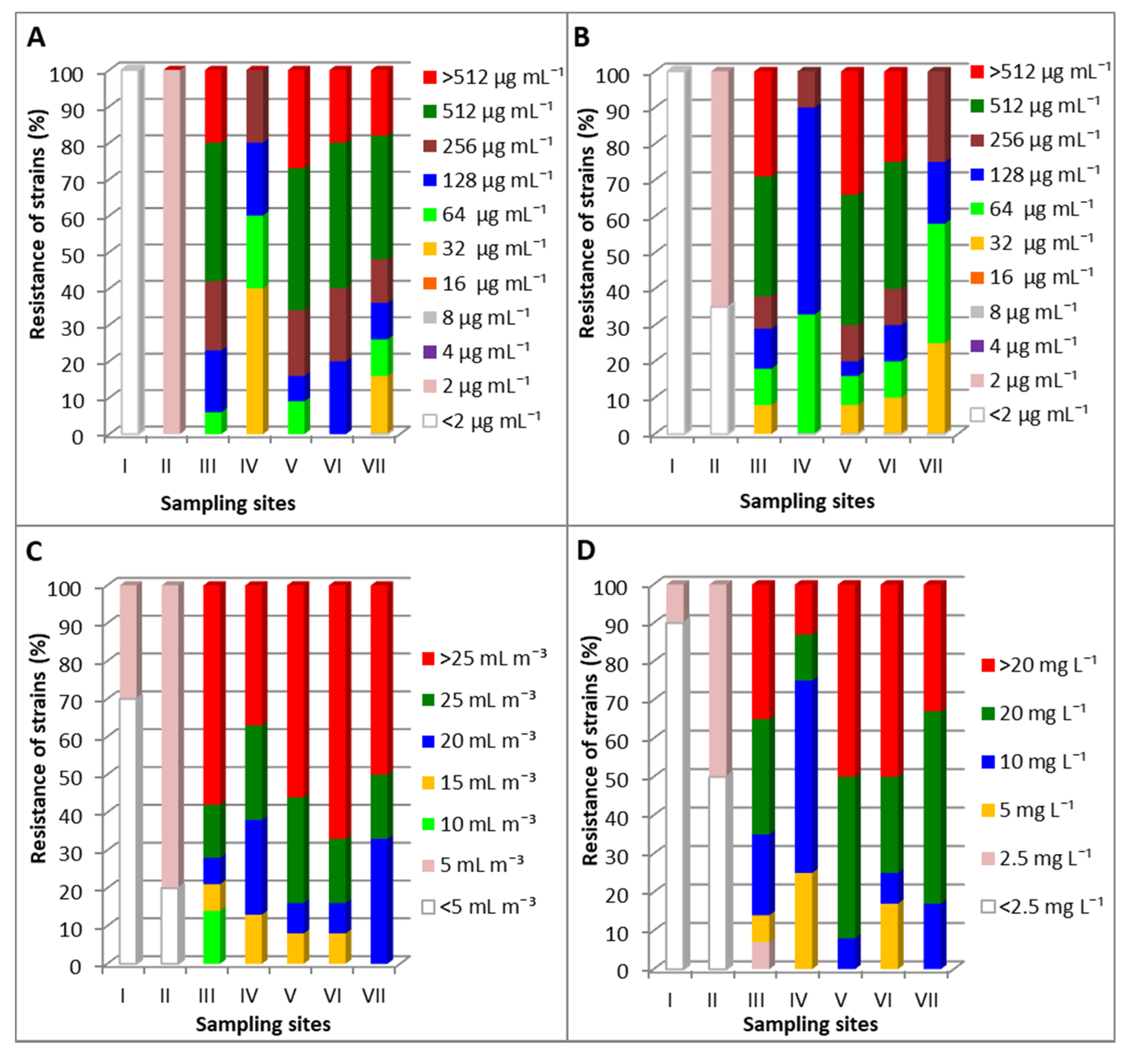
3.2. Resistance of Opportunistic Pathogens P. fluorescens (OPPF) to the Examined Antibiotics and Disinfectants
3.3. Minimum Inhibitory Concentrations (MIC) of Antibiotics and Disin fectants
4. Discussion
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Heinaru, E.; Truu, J.; Stottmeister, U.; Heinaru, A. Three types of phenol and p-cresol catabolism in phenol- and p-cresol-degrading bacteria isolated from river water continuously polluted with phenolic compounds. FEMS Microbiol. Ecol. 2000, 31, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Palleroni, N.J. Pseudomonas Bacteriology Organisms and Their Biology. In Topley & Wilson’s Microbiology and Microbial Infections; John Wiley & Sons, Ltd.: New York, NY, USA, 2010. [Google Scholar]
- Picot, L.; Abdelmoula, S.M.; Merieau, A.; Leroux, P.; Cazin, L.; Orange, N.; Feuilloley, M.G.J. Pseudomonas fluorescens as a potential pathogen: Adherence to nerve cells. Microbes Infect. 2001, 3, 985–995. [Google Scholar] [CrossRef]
- Terech-Majewska, E. Improving disease prevention and treatment in controlled fish culture. Arch. Pol. Fish. 2016, 24, 115–165. [Google Scholar] [CrossRef]
- Abd El Tawab, A.A.; Ahmed, A.; Maarouf, A.; Nesma, A. Detection of Virulence factors of Pseudomonas species isolated from fresh water fish by PCR. BVMJ 2016, 30, 199–207. [Google Scholar] [CrossRef]
- Odjadjare, E.E.; Igbinosa, E.O.; Mordi, R.; Igere, B.; Igeleke, C.L.; Okoh, A.I. Prevalence of multiple antibiotics resistant (MAR) Pseudomonas species in the final effluents of three municipal wastewater treatment facilities in south Africa. Int. J. Environ. Res. Public Health 2012, 9, 2092–2107. [Google Scholar] [CrossRef] [PubMed]
- Aich, N.; Naznin, A.; Anirban, A. Issues of antibiotic resistance in aquaculture industry and its way forward. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 26–41. [Google Scholar] [CrossRef]
- Sidoruk, M.; Cymes, I. Effect of water management technology used in trout culture on water quality in fish ponds. Water 2018, 10, 1264. [Google Scholar] [CrossRef]
- Obolewski, K.; Glińska-Lewczuk, K.; Kobus, S. An attempt at evaluating the influence of water quality on the qualitative and quantitative structure of epiphytic fauna dwelling on Stratiotes aloides L., a case study on an oxbow lake of the Łyna River. J. Elementol. 2009, 14, 119–134. [Google Scholar] [CrossRef]
- Astel, A.M.; Bigus, K.; Obolewski, K.; Glińska-Lewczuk, K. Spatiotemporal assessment of water chemistry in intermittently open/closed coastal lakes of Southern Baltic. Estuar. Coast. Shelf Sci. 2016, 182, 47–59. [Google Scholar] [CrossRef]
- Sidoruk, M.; Koc, J.; Cymes, I.; Rafałowska, M.; Rochwerger, A.; Sobczyńska-Wójcik, K.; Skibniewska, K.A.; Siemianowska, E.; Guziur, J.; Szarek, J. Risk assessment of surface waters associated with water circulation technologies on trout farms. J. Ecol. Eng. 2014, 15, 76–81. [Google Scholar] [CrossRef]
- Tkachenko, H.; Kurhaluk, N.; Grudniewska, J. Oxidative stress biomarkers in different tissues of rainbow trout (Oncorhynchus mykiss) exposed to Disinfectant-CIP formulated with per acetic acid and hydrogen peroxide. Arch. Pol. Fish. 2014, 22, 207–219. [Google Scholar] [CrossRef]
- Kumar, R.; Surendran, F.K. Phenotypic characterization and antibiotic resistance of Pseudomonas spp. from seafood and aquaculture farm environments. Fish. Technol. 2005, 42, 203–208. [Google Scholar]
- Harnisz, M.; Gołaś, I.; Pietruk, M. Tetracycline-resistant bacteria as indicators of antimicrobial resistance in protected waters-the example of the Drwęca River nature reserve (Poland). Ecol. Indic. 2011, 11, 663–668. [Google Scholar] [CrossRef]
- Pimentel, G.A.; Almeida, P.; Hantson, A.-L.; Rapaport, A.; Vande Wouwer, A. Experimental validation of a simple dynamic model of a laboratory scale recirculating aquaculture system fitted with a submerged membrane bioreactor. Biochem. Eng. J. 2017, 122, 1–12. [Google Scholar] [CrossRef]
- Fossmark, R.O.; Vadstein, O.; Rostend, T.W.; Bakke, I.; Košeto, D.; Bugten, A.V.; Helberg, G.A.; Nesje, J.; Jørgensen, N.O.G.; Raspati, G.; et al. Effects of reduced organic matter loading through membrane filtration on the microbial community dynamics in recirculating aquaculture systems (RAS) with Atlantic salmon parr (Salmo salar). Aquaculture 2020, 524, 735268. [Google Scholar] [CrossRef]
- Summerfelt, S.T.; Sharrer, M.J.; Tsukuda, S.M.; Gearheart, M. Process requirements for achieving full-flow disinfection of recirculating water using ozonation and UV irradiation. Aquac. Eng. 2009, 40, 17–27. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Cuesta, A.; Meseguer, J.; Esteban, M.A. Risks of using antifouling biocides in aquaculture. Int. J. Mol. Sci. 2012, 13, 1541–1560. [Google Scholar] [CrossRef]
- Research Strategy to Address the Knowledge Gaps on the Antimicrobial Resistance Effects of Biocides. Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR). 17 March 2010. Available online: https://ec.europa.eu/health/sites/health/files/scientific_committees/emerging/docs/scenihr_o_028.pdf (accessed on 2 June 2020).
- Grudniewska, J.; Terech-Majewska, E.; Goral, C. Evaluate the efficacy of the biocidal product in the hatchery during egg incubation and prophylactic bathing rainbow trout (Oncorhynchus mykiss) fry. In Biotechnology in Aquaculture; Zakęś, Z., Wolnicki, J., Demska-Zakęś, K., Kaminski, R., Eds.; Wyd. IRS: Olsztyn, Poland, 2008; pp. 395–401. (In Polish) [Google Scholar]
- Terech-Majewska, E.; Grudniewska, J.; Siwicki, A.K. Disinfection with the most effective biocides as a prophylactic method to supplement the treatment of fish diseases. Komun. Ryb 2010, 2, 11–16. (In Polish) [Google Scholar]
- Serrano, P.H. Responsible use of antibiotics in aquaculture. In FAO Fisheries Technical Paper 469; Food and Agriculture Organization of the United Nations: Rome, Italy, 2005. [Google Scholar]
- European Economic Commission. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora; European Economic Commission: Brussels, Belgium, 1992. [Google Scholar]
- Ordinance of the Minister of Forestry and Wood Industry of July 27, 1961 regarding recognition as a nature reserve (MP No. 71, item 302).
- Gołaś, I. Anthropogenic and Environmental Factors Affecting the Bacteriological Quality of the Upper Drwęca River; Wyd. UWM: Olsztyn, Poland, 2011; p. 122. (In Polish) [Google Scholar]
- Teodorowicz, M. Surface water quality and intensive fish culture. Arch. Pol. Fish. 2013, 21, 65–111. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 15th ed.; Water Pollution Control Federation: New York, NY, USA, 1992. [Google Scholar]
- Saha, R.S.; Donofrio, D.; Goeres, M.; Bagley, S.T. Rapid detection of rRNA group I pseudomonads in contaminated metalworking fluids and biofilm formation by fluorescent in situ hybridization. Appl. Microbiol. Biotechnol. 2012, 94, 799–808. [Google Scholar] [CrossRef]
- Gołaś, I.; Szmyt, M.; Potorski, J.; Łopata, M.; Gotkowska-Płachta, A.; Glińska-Lewczuk, K. Distribution of Pseudomonas fluorescens and Aeromonas hydrophila bacteria in a recirculating aquaculture system during farming of european grayling (Thymallus thymallus L.) Broodstock. Water 2019, 11, 376–391. [Google Scholar]
- Rogulska, A.; Antychowicz, J.; Żelazny, J. Haemolytic and proteolytic activity of Aeromonas hydrophila and A. sobria as markers of pathogenicity for carp (Cyprinus carpio L.). Med. Weter. 1994, 50, 55–58. [Google Scholar]
- Hsu, T.C.; Waltman, W.D.; Shotts, E.B. Correlation of extracellular enzymatic activity and biochemical characteristics with regard to virulence of Aeromonas hydrophila. Dev. Biol. Stand. 1981, 49, 101–111. [Google Scholar]
- Chang, C.I.; Liu, W.Y.; Shyu, C.Z. Use of prawn blood agar hemolysis to screen for bacteria opportunistic to cultured tiger prawns Penaeus monodon. Dis. Aquat. Organ. 2000, 43, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.D.; Kristiansen, K.R.; Hansen, T.K. Characterization of the proteolytic activity of starter cultures of Penicillium roqueforti for production of blue veined cheeses. Int. J. Food Microbiol. 1998, 43, 215–221. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.N.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. An. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Susceptibility Tests; Approved Standard, 11th ed.; CLSI document: M02–A11; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2012. [Google Scholar]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of food. Appl. Environ. Microb. 1983, 46, 165–170. [Google Scholar] [CrossRef]
- Paul, S.L.; Bezbaruah, R.L.; Roy, M.K.; Ghosh, A.C. Multiple antibiotic resistance index and its reversion in Pseudomonas aeruginosa. Lett. Appl. Microbiol. 1997, 24, 169–171. [Google Scholar] [CrossRef]
- Baker, C.N.; Stocker, S.A.; Culver, D.H.; Thornsberry, C. Comparison of the E test to agar dilution, broth microdilution, and agar diffusion susceptibility testing techniques by using a special challenge set of bacteria. J. Clin. Microbiol. 1991, 29, 533–538. [Google Scholar] [CrossRef]
- Stanisz, A. A straightforward course in statistics based on the STATISTICA PL program as applied in medicine. In Volume Basic Statistics; StatSoft: Cracow, Poland, 2006; ISBN 83-88724-18-5. (In Polish) [Google Scholar]
- Madi, A.; Lakhdari, O.; Blottière, H.M.; Guyard-Nicodeme, M.; Le Roux, K.; Groboillot, A.; Svinareff, P.; Doré, J.; Orange, N.; Feuilloley, M.G.J.; et al. The clinical Pseudomonas fluorescens MFN1032 strain exerts a cytotoxic effect on epithelial intestinal cells and induces Interleukin-8 via the AP-1 signaling pathway. BMC Microbiol. 2010, 10, 215–222. [Google Scholar] [CrossRef]
- Trivedi, M.K.; Patil, S.; Shettigar, H.; Gangwar, M.; Jana, S. Antimicrobial Sensitivity Pattern of Pseudomonas fluorescens after Biofield Treatment. J. Infect. Dis. Ther. 2015, 3, 222–226. [Google Scholar] [CrossRef]
- Mirand, C.D.; Zemelman, R. Antimicrobial multiresistance in bacteria isolated from freshwater Chilean salmon farms. Sci. Total Environ. 2002, 293, 207–218. [Google Scholar] [CrossRef]
- Spanggaard, B.; Jørgensen, F.; Gram, L.; Huss, H.H. Antibiotic resistance in bacteria isolated from three freshwater fish farms and an unpolluted stream in Denmark. Aquaculture 1993, 115, 195–207. [Google Scholar] [CrossRef]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Pedersen, K.; Larsen, J.L. Occurrence of antimicrobial resistance in fish-pathogenic and environmental bacteria associated with four Danish rainbow trout farms. Appl. Environ. Microbiol. 2000, 66, 4908–4915. [Google Scholar] [CrossRef]
- Pirnay, J.-P.; Matthijs, S.; Colak, H.; Chablain, P.; Bilocq, F.; van Eldere, J.; de Vos, D.; Zizi, M.; Triest, L.; Cornelis, P. Global Pseudomonas aeruginosa biodiversity as reflected in a Belgian river. Environ. Microbiol. 2005, 7, 969–980. [Google Scholar] [CrossRef]
- Kholil, M.I.; Hossain, M.M.M.; Neowajh, M.S.; Islam, M.S.; Kabir, M. Comparative efficiency of some commercial antibiotics against Pseudomonas infection in fish. Int. J. Fish. Aquat. Stud. 2015, 2, 114–117. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Number of Samples | TPFP 1 (cfu·mL−1) | OPPF 2 (cfu·mL−1) | Percentage of OPPF in TPFP (%) |
|---|---|---|---|---|
| Mean ± SD | Mean ± SD | Mean ± SD | ||
| I | 8 | 12 ± 2 ab | 1 ± 1 ab | 11.2 ± 9.4 |
| II | 8 | 24 ± 5 ab | 3 ± 2 ab | 11.5 ± 7.2 |
| III | 8 | 45 ± 11a | 38 ± 8 a | 84.8 ± 3.8 |
| IV | 8 | 36 ± 6 a | 16 ± 5 a | 42.9 ± 8.9 |
| V | 8 | 56 ± 21 a | 48 ± 18 a | 86.5 ± 8.0 |
| VI | 8 | 62 ± 19 a | 56 ± 18 a | 89.5 ± 3.6 |
| VII | 8 | 34 ± 9 a | 19 ± 5 a | 57.9 ± 14.5 |
| Sampling Sites | Number of Isolates | Antibiotic Groups | MAR Index 13 | Disinfectants | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Penicillins | Fluoroquinolones | Sulfonamides | Aminoglycosides | Tetracyclines | Ster 14 | Chlor 15 | ||||||||||
| PRL 1 | CAZ 2 | AMC 3 | MEZ 4 | CTX 5 | CIP 6 | NOR 7 | SXT 8 | CN 9 | TOB 10 | TGC 11 | TE 12 | |||||
| Number of Resistant Isolates | ||||||||||||||||
| I | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.03 | 0 | 0 |
| II | 6 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.04 | 0 | 0 |
| III | 14 | 6 | 0 | 8 | 8 | 11 | 1 | 4 | 5 | 0 | 0 | 1 | 3 | 0.27 | 9 | 5 |
| IV | 8 | 4 | 0 | 4 | 1 | 5 | 2 | 1 | 3 | 0 | 0 | 0 | 1 | 0.20 | 3 | 1 |
| V | 14 | 4 | 3 | 7 | 10 | 12 | 5 | 12 | 5 | 3 | 5 | 2 | 11 | 0.47 | 9 | 7 |
| VI | 12 | 4 | 6 | 6 | 4 | 10 | 2 | 10 | 6 | 5 | 5 | 5 | 11 | 0.51 | 8 | 7 |
| VII | 6 | 3 | 0 | 2 | 6 | 6 | 0 | 6 | 4 | 1 | 1 | 0 | 3 | 0.45 | 3 | 2 |
| Total | 63 | 22 | 9 | 29 | 29 | 45 | 10 | 33 | 23 | 8 | 11 | 8 | 29 | - | 32 | 22 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gołaś, I. The Influence of Aquaculture Effluents on the Prevalence and Biocides Resistance of Opportunistic Pseudomonas fluorescens Bacteria in the Drwęca River Protected under the Natura 2000 Network. Water 2020, 12, 1947. https://doi.org/10.3390/w12071947
Gołaś I. The Influence of Aquaculture Effluents on the Prevalence and Biocides Resistance of Opportunistic Pseudomonas fluorescens Bacteria in the Drwęca River Protected under the Natura 2000 Network. Water. 2020; 12(7):1947. https://doi.org/10.3390/w12071947
Chicago/Turabian StyleGołaś, Iwona. 2020. "The Influence of Aquaculture Effluents on the Prevalence and Biocides Resistance of Opportunistic Pseudomonas fluorescens Bacteria in the Drwęca River Protected under the Natura 2000 Network" Water 12, no. 7: 1947. https://doi.org/10.3390/w12071947
APA StyleGołaś, I. (2020). The Influence of Aquaculture Effluents on the Prevalence and Biocides Resistance of Opportunistic Pseudomonas fluorescens Bacteria in the Drwęca River Protected under the Natura 2000 Network. Water, 12(7), 1947. https://doi.org/10.3390/w12071947