Assessing the Water-Stress Baselines by Thermal Imaging for Irrigation Management in Almond Plantations under Water Scarcity Conditions

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Irrigation Treatments
2.3. Plant Measurements
2.4. Experimental Design and Statistical Analysis
3. Results
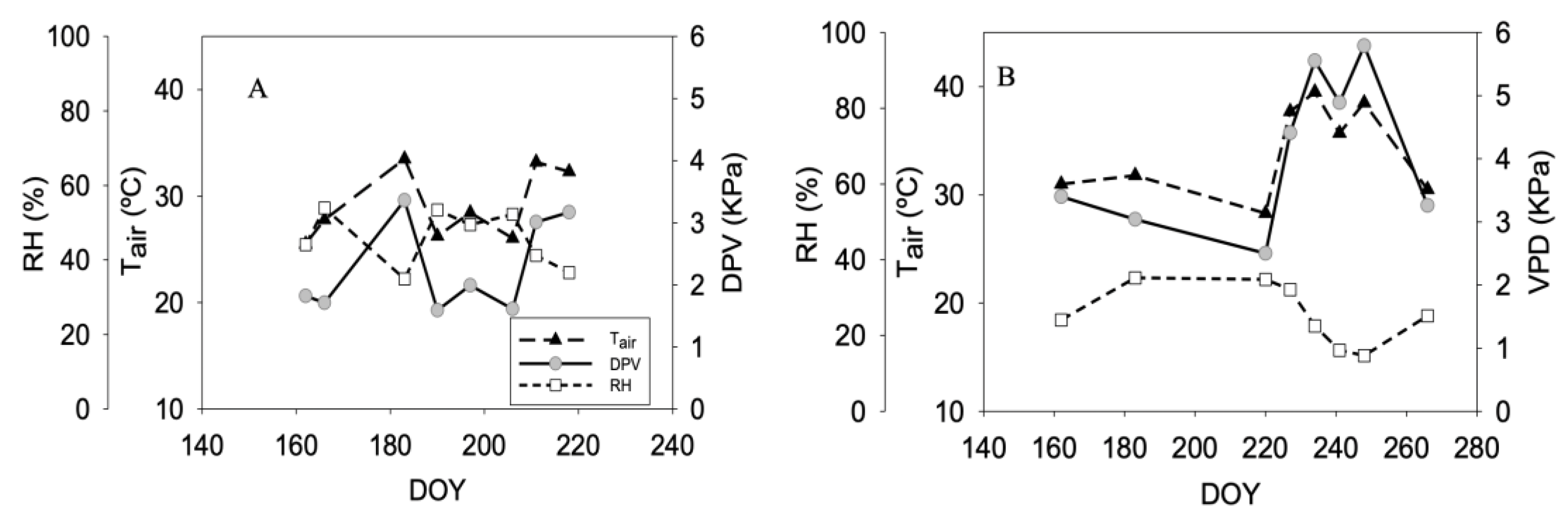
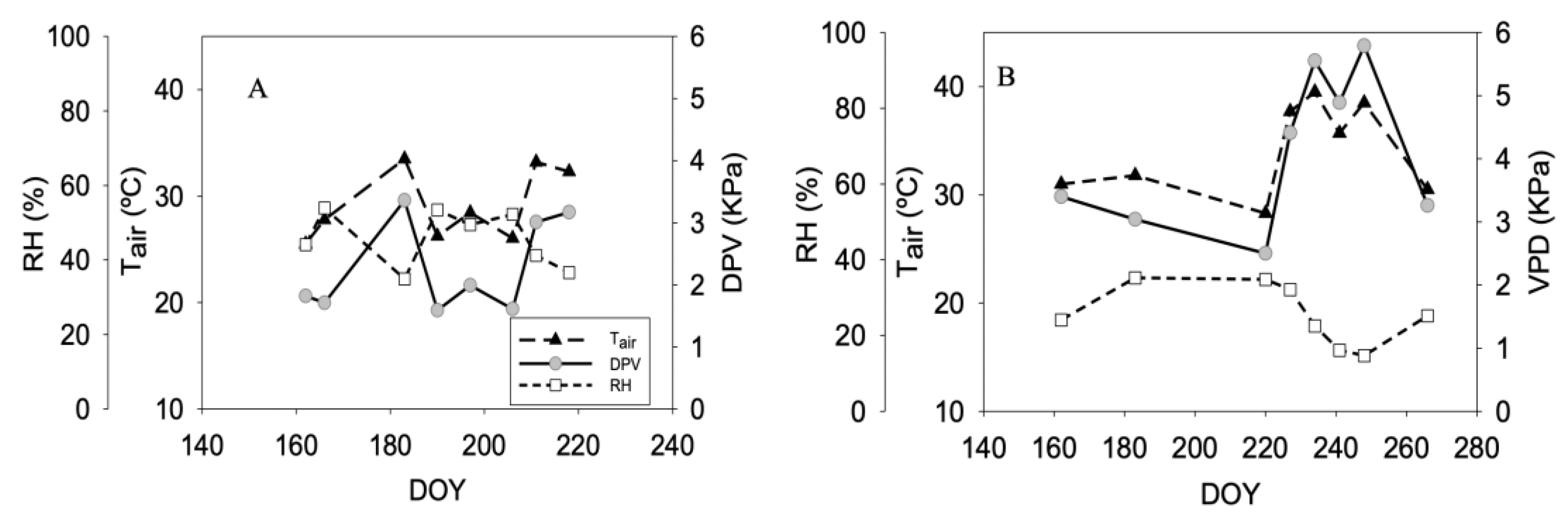
3.1. Climate Condition and Irrigation Water Amount Applied
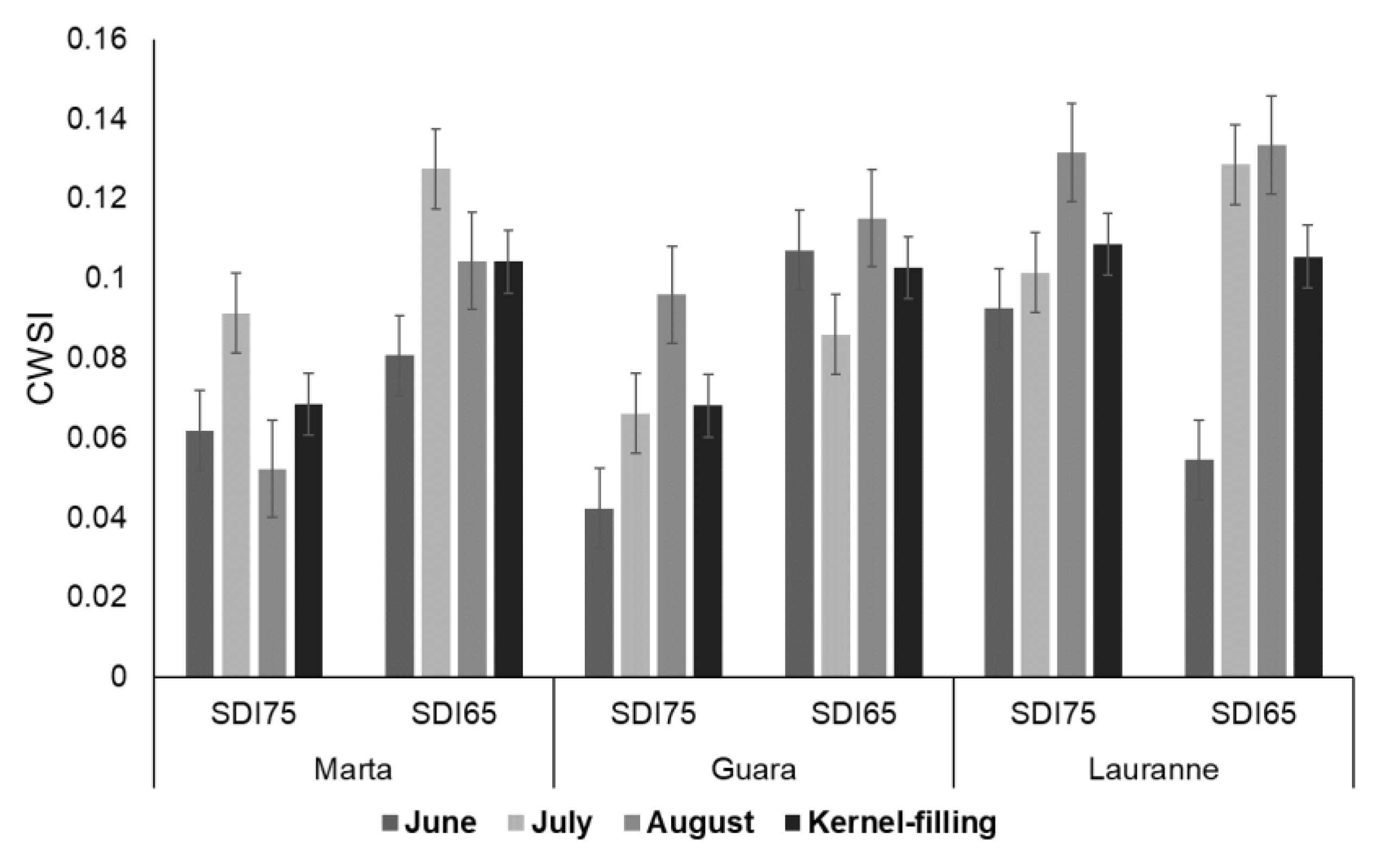
3.2. Physiological Response to Irrigation Treatments
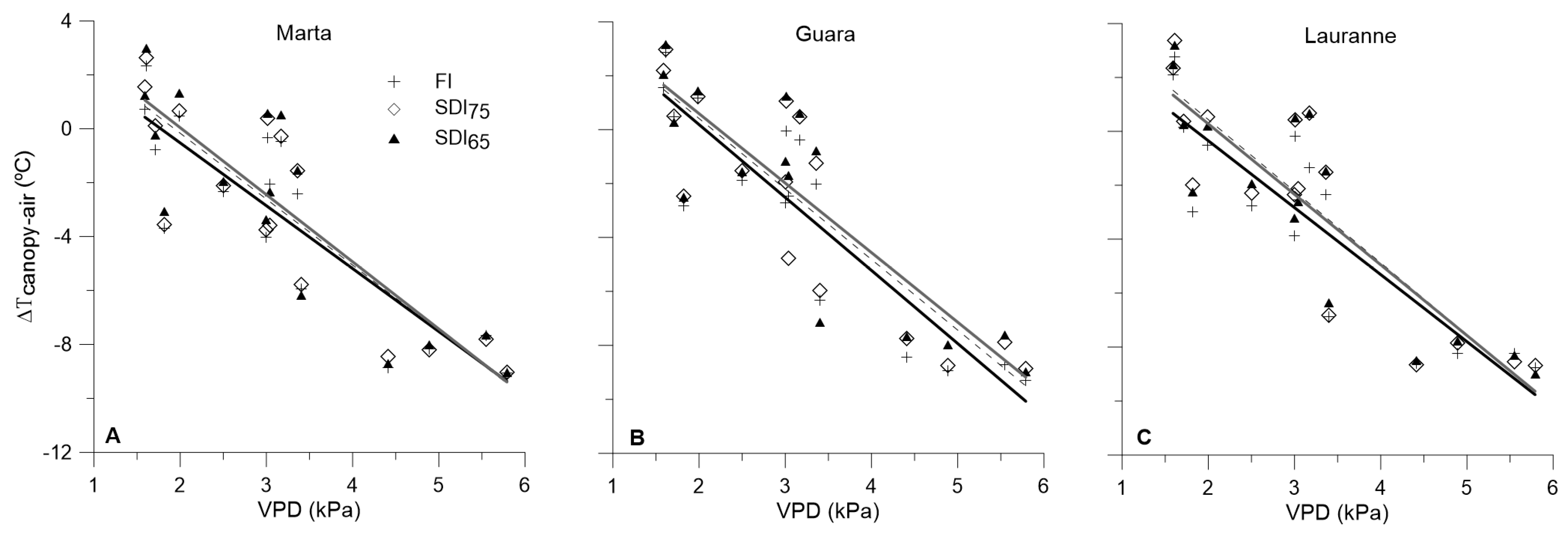
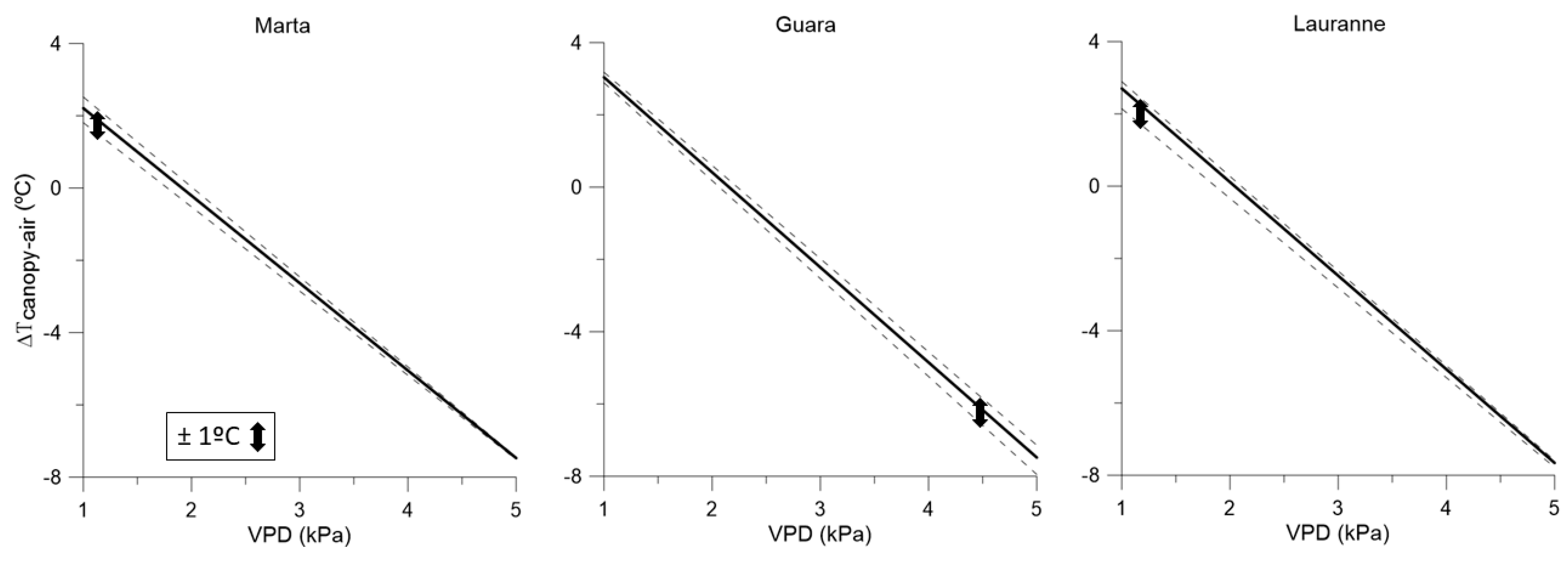
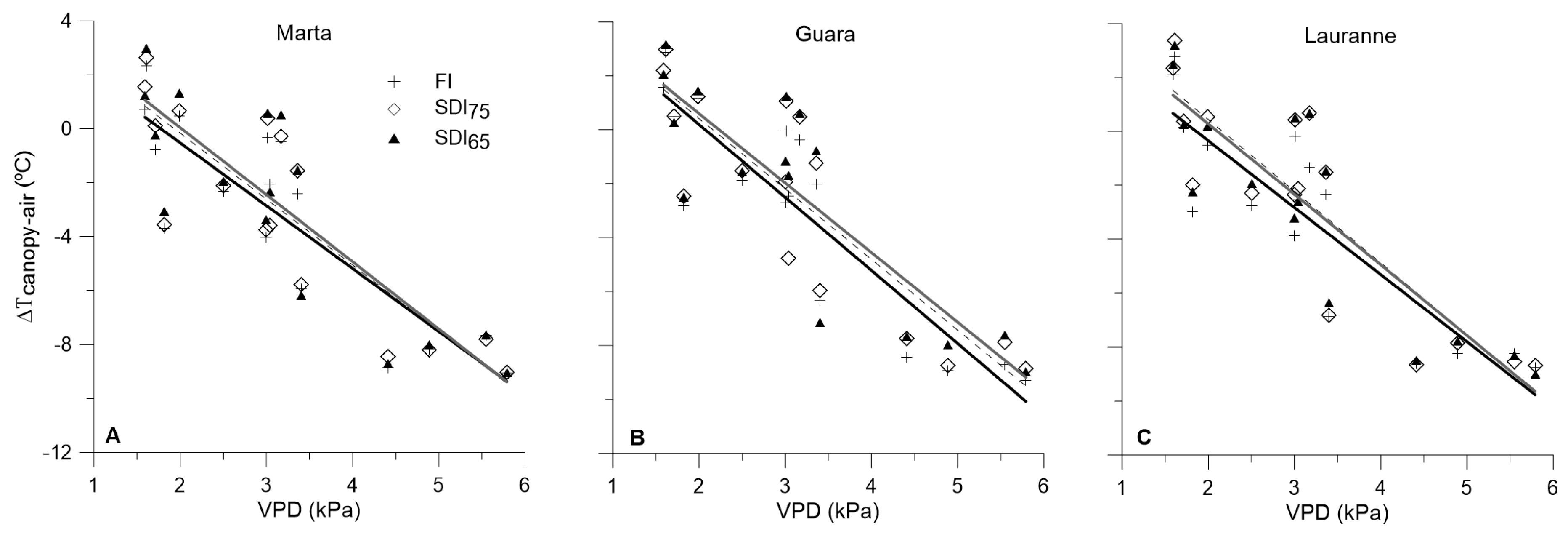
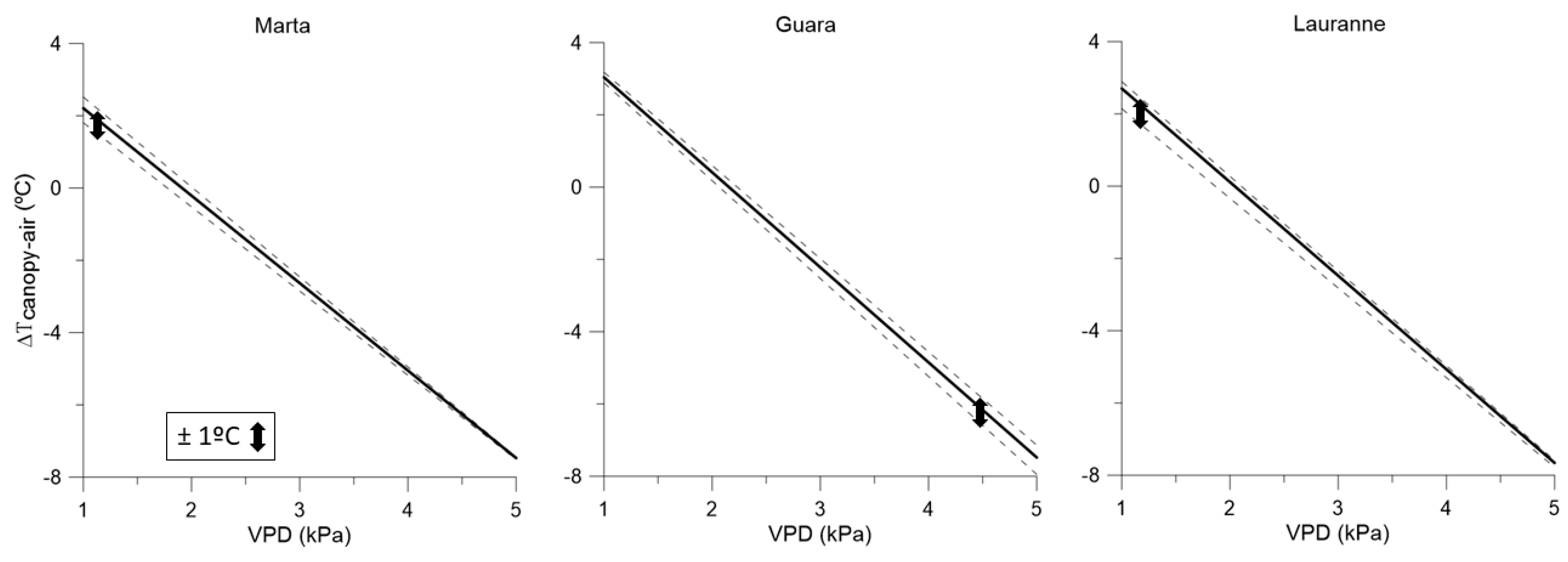
3.3. Water-Stress Baselines for Each Cultivar and Irrigation Treatment, and Their Interactions
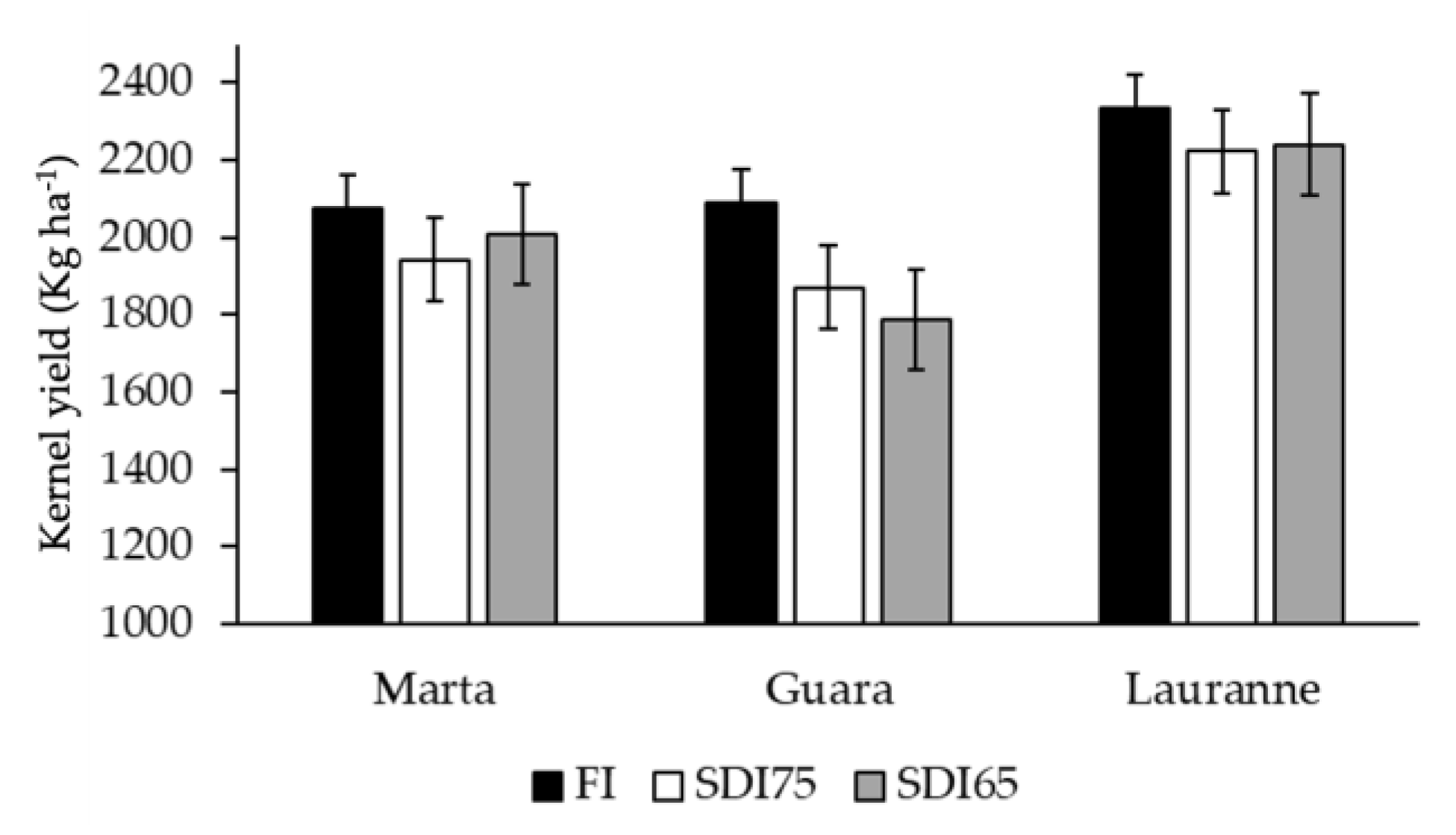
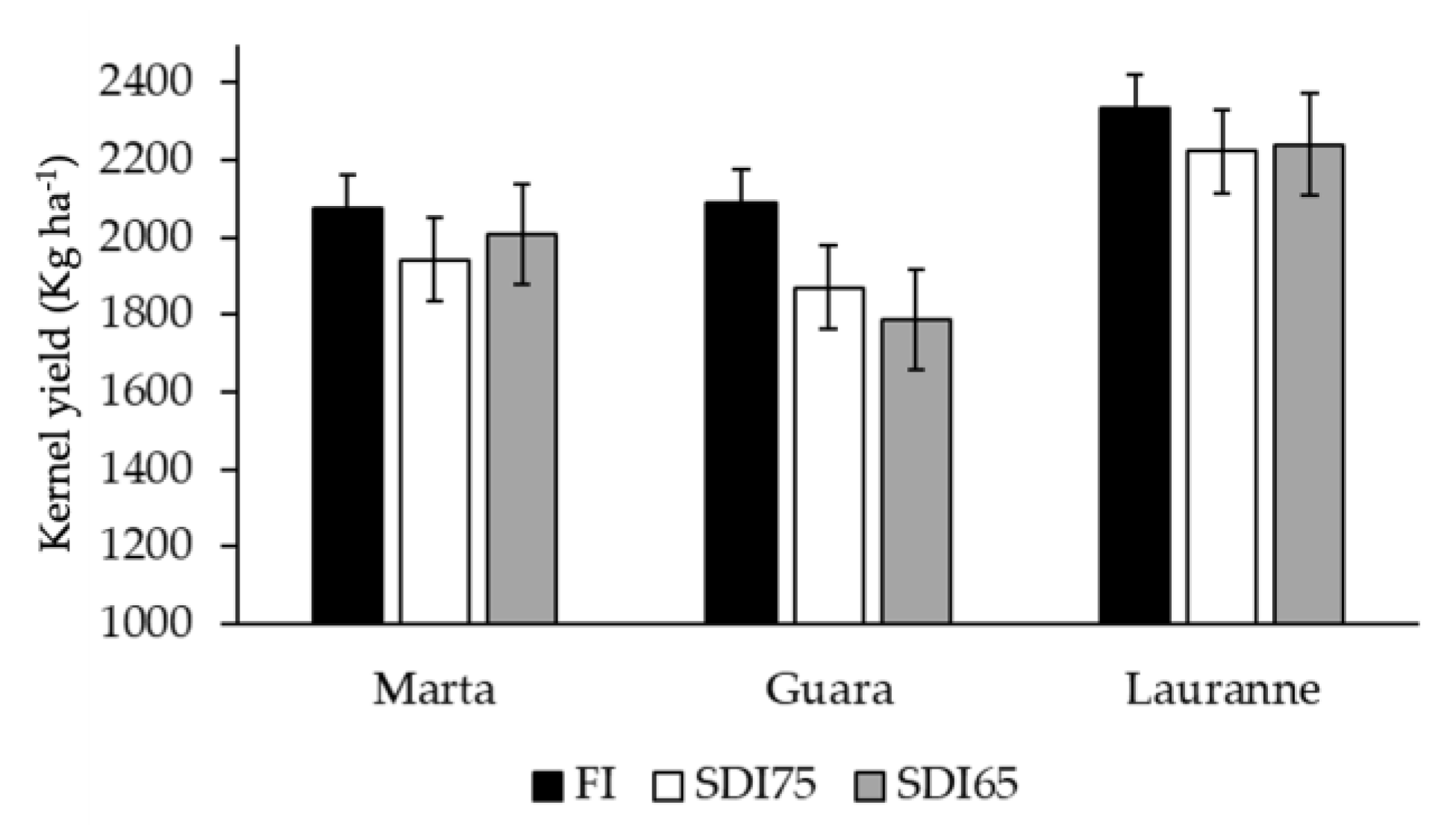
3.4. Linking the Yield with Water-Stress Baselines Defined for Each Cultivar and Irrigation Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Iglesias, A.; Mougou, R.; Moneo, M.; Quiroga, S. Towards adaptation of agriculture to climate change in the Mediterranean. Reg. Environ. Chang. 2011, 11, 159–166. [Google Scholar] [CrossRef]
- Fereres, E.; Soriano, M.A. Deficit irrigation for reducing agricultural water use. J. Exp. Bot. 2007, 58, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, J.E. Understanding olive adaptation to abiotic stresses as a tool to increase crop performance. Environ. Exp. Bot. 2014, 103, 158–179. [Google Scholar] [CrossRef] [Green Version]
- Durán Zuazo, V.H.; Rodríguez Pleguezuelo, C.R.; Franco Tarifa, D. Impact of sustained-deficit irrigation on tree growth, mineral nutrition, fruit yield and quality of mango in Spain. Fruits 2011, 66, 257–268. [Google Scholar] [CrossRef]
- Buchner, R.P.; Fulton, A.E.; Gilles, C.K.; Prichard, T.L.; Lampinen, B.D.; Shackel, K.A.; Metcalf, S.G.; Little, C.C.; Schwankl, L.J. Effects of regulated deficit irrigation on walnut grafted on “Northern California Black” or “Paradox” rootstock. Acta Hortic. 2008, 792, 141–146. [Google Scholar] [CrossRef]
- García-Tejero, I.; Romero-Vicente, R.; Jiménez-Bocanegra, J.A.; Martínez-García, G.; Durán Zuazo, V.H.; Muriel-Fernández, J.L. Response of citrus trees to deficit irrigation during different phenological periods in relation to yield, fruit quality, and water productivity. Agric. Water Manag. 2010, 97, 689–699. [Google Scholar] [CrossRef]
- Memmi, H.; Gijón, M.C.; Couceiro, J.F.; Pérez-López, D. Water stress thresholds for regulated deficit irrigation in pistachio trees: Rootstock influence and effects on yield quality. Agric. Water Manag. 2016, 164, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Caracterización del sector de la almendra en Andalucía. Secretaría General de Agricultura y Alimentación. Junta de Andalucía. 2016, 34. Available online: https://www.juntadeandalucia.es/export/drupaljda/estudios_informes/16/12/Caracterizaci%C3%B3n%20del%20sector%20de%20la%20almendra_0.pdf (accessed on 5 April 2020).
- Egea, G.; González-Real, M.M.; Baille, A.; Nortes, P.A.; Sánchez-Bel, P.; Domingo, R. The effects of contrasted deficit irrigation strategies on the fruit growth and kernel quality of mature almond trees. Irrig. Sci. 2009, 36, 1–8. [Google Scholar] [CrossRef]
- Romero, P.; Botia, P.; Garcia, F. Effects of regulated deficit irrigation under subsurface drip irrigation conditions on vegetative development and yield of mature almond trees. Plant Soil 2004, 260, 169–181. [Google Scholar] [CrossRef]
- Espadafor, M.; Orgaz, F.; Testi, L.; Lorite, I.J.; González-Dugo, V.; Fereres, E. Responses of transpiration and transpiration efficiency of almond trees to moderate water deficits. Sci. Hortic. (Amsterdam) 2017, 225, 6–14. [Google Scholar] [CrossRef]
- Nortes, P.A.; Gonzalez-Real, M.M.; Egea, G.; Baille, A. Seasonal effects of deficit irrigation on leaf photosynthetic traits of fruiting and non-fruiting shoots in almond trees. Tree Physiol. 2009, 29, 375–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girona, J.; Mata, M.; Marsal, J. Regulated deficit irrigation during the kernel-filling period and optimal irrigation rates in almond. Agric. Water Manag. 2005, 75, 152–167. [Google Scholar] [CrossRef]
- Rojo, F.; Kizer, E.; Upadhyaya, S.; Ozmen, S.; Ko-Madden, C.; Zhang, Q. A Leaf Monitoring System for Continuous Measurement of Plant Water Status to Assist in Precision Irrigation in Grape and Almond crops. IFAC-PapersOnLine 2016, 49, 209–215. [Google Scholar] [CrossRef]
- Dhillon, R.; Rojo, F.; Roach, J.; Upadhyaya, S.; Delwiche, M. A Continuous Leaf Monitoring System for Precision Irrigation Management in Orchard Crops. Tarım Makinaları Bilim. Derg. 2014, 10, 267–272. [Google Scholar]
- Fernández, J.E. Plant-based methods for irrigation scheduling of woody crops. Horticulturae 2017, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Idso, S.B. Non-water-stressed baselines: A key to measuring and interpreting plant water stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Jones, H.G. Irrigation scheduling: Advantages and pitfalls of plant-based methods. J. Exp. Bot. 2004, 55, 2427–2436. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Bello, M.A.; Ballester, C.; Castel, J.R.; Intrigliolo, D.S. Development and validation of an automatic thermal imaging process for assessing plant water status. Agric. Water Manag. 2011, 98, 1497–1504. [Google Scholar] [CrossRef] [Green Version]
- Egea, G.; Padilla-Díaz, C.M.; Martinez-Guanter, J.; Fernández, J.E.; Pérez-Ruiz, M. Assessing a crop water stress index derived from aerial thermal imaging and infrared thermometry in super-high density olive orchards. Agric. Water Manag. 2017, 187, 210–221. [Google Scholar] [CrossRef] [Green Version]
- García-Tejero, I.F.; Gutiérrez Gordillo, S.; Souza, L.; Cuadros-Tavira, S.; Durán Zuazo, V.H. Fostering sustainable water use in almond (Prunus dulcis Mill.) orchards in a semiarid Mediterranean environment. Arch. Agron. Soil Sci. 2019, 65, 164–181. [Google Scholar] [CrossRef]
- Pou, A.; Diago, M.P.; Medrano, H.; Baluja, J.; Tardaguila, J. Validation of thermal indices for water status identification in grapevine. Agric. Water Manag. 2014, 134, 60–72. [Google Scholar] [CrossRef]
- Poirier-Pocovi, M.; Volder, A.; Bailey, B.N. Modeling of reference temperatures for calculating crop water stress indices from infrared thermography. Agric. Water Manag. 2020, 233, 106070. [Google Scholar] [CrossRef]
- Jones, H.G.; Serraj, R.; Loveys, B.R.; Xiong, L.; Wheaton, A.; Price, A.H. Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of plant responses to water stress in the field. Funct. Plant Biol. 2009, 36, 978–989. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G.; Vaughan, R. Remote Sensing of Vegetation: Principles, Techniques and Applications. Soil Use Manag. 2012, 28, 134. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Hernández, A.; Padilla-Díaz, C.M.; Diaz-Espejo, A.; Fernández, J.E. Assessing plant water status in a hedgerow olive orchard from thermography at plant level. Agric. Water Manag. 2017, 188, 50–60. [Google Scholar] [CrossRef] [Green Version]
- García-Tejero, I.F.; Gutiérrez-Gordillo, S.; Ortega-Arévalo, C.; Iglesias-Contreras, M.; Moreno, J.M.; Souza-Ferreira, L.; Durán Zuazo, V.H. Thermal imaging to monitor the crop-water status in almonds by using the non-water stress baselines. Sci. Hortic. 2018, 238, 91–97. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements-Irrigation and Drainage Paper 56; Food and Agriculture Organization (FAO): Rome, Italy, 1998; Available online: http://www.fao.org/docrep/x0490e/x0490e00.htm (accessed on 12 February 2020).
- García-Tejero, I.F.; Hernandez, A.; Rodriguez, V.M.; Ponce, J.R.; Ramos, V.; Muriel, J.L.; Durán Zuazo, V.H. Estimating Almond Crop Coefficients and Physiological Response to Water Stress in Semiarid Environments (SW Spain). J. Agric. Sci. Technol. 2015, 17, 1255–1266. [Google Scholar]
- García-Tejero, I.F.; Rubio, A.E.; Viñuela, I.; Hernández, A.; Gutiérrez-Gordillo, S.; Rodríguez-Pleguezuelo, C.R.; Durán Zuazo, V.H. Thermal imaging at plant level to assess the crop-water status in almond trees (cv. Guara) under deficit irrigation strategies. Agric. Water Manag. 2018, 208. [Google Scholar] [CrossRef]
- Fu, X.; Meinzer, F.C. Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): A global data set reveals coordination and trade-offs among water transport traits. Tree Physiol. 2018, 39, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Santana, V.; Rodriguez-Dominguez, C.M.; Fernández, J.E.; Diaz-Espejo, A. Role of leaf hydraulic conductance in the regulation of stomatal conductance in almond and olive in response to water stress. Tree Physiol. 2016, 36, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Dominguez, C.M.; Buckley, T.N.; Egea, G.; de Cires, A.; Hernandez-Santana, V.; Martorell, S.; Diaz-Espejo, A. Most stomatal closure in woody species under moderate drought can be explained by stomatal responses to leaf turgor. Plant Cell Environ. 2016, 39, 2014–2026. [Google Scholar] [CrossRef]
- González-Dugo, V.; Zarco-Tejada, P.J.; Fereres, E. Applicability and limitations of using the crop water stress index as an indicator of water deficits in citrus orchards. Agric. For. Meteorol. 2014, 198–199, 94–104. [Google Scholar] [CrossRef]
- Bellvert, J.; Zarco-Tejada, P.J.; Girona, J.; Fereres, E. Mapping crop water stress index in a ‘Pinot-noir’ vineyard: Comparing ground measurements with thermal remote sensing imagery from an unmanned aerial vehicle. Precis. Agric. 2014, 15, 361–376. [Google Scholar] [CrossRef]
- Romero-Trigueros, C.; Bayona Gambín, J.M.; Nortes Tortosa, P.A.; Alarcón Cabañero, J.J.; Nicolás, E.N. Determination of Crop Water Stress index by infrared thermometry in grapefruit trees irrigated with saline reclaimed water combined with deficit irrigation. Remote Sens. 2019, 11, 757. [Google Scholar] [CrossRef] [Green Version]
- Egea, G.; Nortes, P.A.; Domingo, R.; Baille, A.; Pérez-Pastor, A.; González-Real, M.M. Almond agronomic response to long-term deficit irrigation applied since orchard establishment. Irrig. Sci. 2013, 31, 445–454. [Google Scholar] [CrossRef]
- López-López, M.; Espadador, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Water use of irrigated almond trees when subjected to water deficits. Agric. Water Manag. 2018, 195, 84–93. [Google Scholar] [CrossRef]
- Monks, D.P.; Taylor, C.; Sommer, K.; Treeby, M.T. Deficit irrigation of almond trees did not decrease yield. Acta Hortic. 2017, 1150, 251–260. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficients | March | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|---|
| KC | 0.4 | 0.6 | 0.9 | 1.1 | 1.2 | 1.1 | 0.8 | 0.7 |
| Kr | 0.4 | 0.7 | 0.8 | 0.9 | 0.9 | 0.8 | 0.8 | 0.7 |
| Parameters | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|
| 2018 | |||||||
| Tmax | 22.0 | 25.6 | 30.5 | 33.7 | 37.7 | 33.6 | 26.3 |
| Tmin | 9.5 | 12.1 | 15.2 | 15.9 | 18.7 | 18.3 | 13.1 |
| Tav | 15.2 | 18.2 | 22.5 | 24.4 | 27.6 | 24.9 | 19.0 |
| RHmax | 967.3 | 936.5 | 93.6 | 96.1 | 87.9 | 89.7 | 95.3 |
| RHmin | 44.1 | 37.3 | 33.3 | 27.5 | 20.7 | 30.7 | 41.0 |
| RHav | 75.6 | 71.3 | 62.4 | 60.9 | 51.9 | 61.9 | 71.8 |
| Rad | 17.3 | 21.9 | 24.9 | 27.0 | 23.5 | 19.6 | 13.9 |
| R | 97.2 | 103.0 | 5.4 | 0.0 | 0.6 | 21.4 | 98.4 |
| ET0 | 96.6 | 125.5 | 150.8 | 172.1 | 168.8 | 125.4 | 198.1 |
| ETc | 57.9 | 113.0 | 165.9 | 206.5 | 185.6 | 100.3 | 49.9 |
| 2019 | |||||||
| Tmax | 22.2 | 30.4 | 31.3 | 34.5 | 36.5 | 32.4 | 27.6 |
| Tmin | 7.2 | 12.2 | 17.5 | 17.9 | 17.9 | 16.3 | 11.7 |
| Tav | 19.8 | 21.5 | 22.7 | 25.8 | 26.9 | 23.8 | 18.9 |
| RHmax | 97.8 | 85.2 | 83.2 | 84.0 | 77.2 | 81.9 | 90.7 |
| RHmin | 39.8 | 23.3 | 23.4 | 25.3 | 18.7 | 27.6 | 32.9 |
| RHav | 72.2 | 52.3 | 51.4 | 55.4 | 45.9 | 54.4 | 63.2 |
| Rad | 1.9 | 2.1 | 2.1 | 2.9 | 0.8 | 0.9 | 0.7 |
| R | 71.2 | 0.0 | 0.0 | 0.0 | 0.0 | 3.4 | 10.4 |
| ET0 | 111.0 | 198.0 | 202.9 | 238.7 | 170.1 | 121.0 | 76.4 |
| ETc | 61.6 | 126.2 | 151.3 | 209.8 | 140.1 | 92.8 | 54.8 |
| DOY | Treat | Marta | Guara | Lauranne | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ψleaf (MPa) | gs (mmol m−2 s−1) | Tc (°C) | Ψleaf (MPa) | gs (mmol m−2 s−1) | Tc (°C) | Ψleaf (MPa) | gs (mmol m−2 s−1) | Tc (°C) | ||
| 162 | FI | −0.92a | 154.50a | 21.80a | −0.92a | 143.48a | 22.68a | −0.97a | 129.08a | 22.50a |
| SDI75 | −0.82a | 153.17a | 21.94a | −0.95a | 117.80a | 23.04a | −0.98a | 125.30a | 23.51a | |
| SDI65 | −0.88a | 127.90b | 22.39a | −1.04a | 174.48a | 22.95a | −0.93a | 130.25a | 23.21a | |
| 166 | FI | −1.06a | 144.27a | 27.02a | −0.93a | 180.65a | 28.29a | −1.05a | 187.85a | 27.94a |
| SDI75 | −1.13a | 160.50a | 27.90a | −1.21b | 175.07a | 28.30a | −1.08a | 179.68a | 28.18a | |
| SDI65 | −1.11a | 173.93a | 27.53a | −1.22b | 191.10a | 28.03a | −1.05a | 180.88a | 28.00a | |
| 190 | FI | −1.35a | 76.28a | 31.09a | −1.58a | 99.21a | 31.49a | −1.54a | 112.00a | 31.14a |
| SDI75 | −1.44b | 84.09a | 31.94a | −1.50a | 99.83a | 32.26a | −1.70b | 102.88a | 31.97a | |
| SDI65 | −1.40ab | 79.24a | 31.91a | −1.67a | 109.66a | 32.67a | −1.72b | 110.64a | 31.99a | |
| 197 | FI | −1.16a | 77.14a | 26.96a | −1.44a | 79.79a | 27.81a | −1.40a | 93.25a | 28.34a |
| SDI75 | −1.27b | 72.86a | 27.80a | −1.45a | 84.64a | 28.45a | −1.50b | 89.73a | 28.58a | |
| SDI65 | −1.31b | 78.03a | 27.44a | −1.42a | 84.13a | 28.25a | −1.55b | 89.35a | 28.68a | |
| 206 | FI | −1.01a | 84.41a | 28.93a | −1.38a | 107.59a | 29.63a | −1.38a | 126.57a | 27.92b |
| SDI75 | −1.09a | 90.90a | 29.12a | −1.36a | 107.90a | 29.68a | −1.21a | 128.36a | 29.00a | |
| SDI65 | −1.06a | 89.11a | 29.73a | −1.40a | 107.34a | 29.85a | −1.38a | 129.50a | 29.12a | |
| 211 | FI | −1.15a | 110.80a | 28.39a | −1.49a | 120.86a | 28.91a | −1.36a | 126.43a | 28.80a |
| SDI75 | −1.31b | 99.64a | 28.69a | −1.57a | 110.65b | 29.04a | −1.42b | 133.20a | 29.43a | |
| SDI65 | −1.38b | 96.58a | 28.99a | −1.53a | 98.80b | 29.16a | −1.44b | 129.07a | 29.21a | |
| 218 | FI | −1.87a | 138.02a | 32.85a | −2.17a | 154.36a | 33.15b | −1.84a | 189.04a | 33.01a |
| SDI75 | −1.82a | 134.86a | 33.58a | −2.12a | 166.15a | 34.28a | −2.05a | 181.77a | 33.62a | |
| SDI65 | −1.68a | 140.90a | 33.7a | −2.18a | 150.73a | 34.40a | −1.71a | 172.70a | 33.66a | |
| 225 | FI | −1.74a | 129.10a | 31.83a | −1.76a | 138.99a | 31.93a | −1.88a | 166.89a | 30.95b |
| SDI75 | −1.79a | 116.47a | 32.02a | −2.29b | 139.86a | 32.78a | −2.10a | 165.27a | 32.97a | |
| SDI65 | −2.03b | 122.47a | 32.77a | −2.01b | 129.44a | 32.87a | −1.92a | 163.96a | 32.92a | |
| DOY | Treat | Marta | Guara | Lauranne | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ψleaf (MPa) | gs (mmol m−2 s−1) | Tc (°C) | Ψleaf (MPa) | gs (mmol m−2 s−1) | Tc (°C) | Ψleaf (MPa) | gs (mmol m−2 s−1) | Tc (°C) | ||
| 162 | FI | −1.47a | 279.70a | 29.65a | −1.46a | 314.73a | 30.24a | −1.41a | 271.48a | 30.91a |
| SDI75 | −1.52a | 298.97a | 29.94a | −1.75b | 280.65a | 31.91a | −1.58a | 266.93a | 30.73a | |
| SDI65 | −1.60a | 308.45a | 30.54a | −2.11b | 274.88a | 30.47a | −1.49a | 275.62a | 30.10a | |
| 175 | FI | −1.42a | 274.55a | 29.74a | −1.75a | 222.67a | 29.34b | −1.43a | 205.53a | 29.25a |
| SDI75 | −1.54b | 298.87a | 28.23a | −2.04b | 215.02a | 27.02a | −1.87b | 206.88a | 29.67a | |
| SDI65 | −1.52b | 278.13a | 29.41a | −1.77a | 213.72a | 30.06c | −1.83b | 192.12a | 29.15a | |
| 183 | FI | −1.36a | 197.80a | 25.95a | −1.61a | 199.80a | 26.41a | −1.50a | 217.97a | 25.53a |
| SDI75 | −1.52b | 190.88a | 26.18a | −1.80b | 195.37a | 26.79a | −1.80b | 203.43a | 26.01a | |
| SDI65 | −1.54b | 192.73a | 26.29a | −2.03b | 209.57a | 26.69a | −1.64b | 206.38a | 26.31a | |
| 189 | FI | −1.36a | 197.80a | 25.95b | −1.61a | 199.80a | 26.41a | −1.50a | 217.97a | 25.53a |
| SDI75 | −1.52b | 190.88a | 26.18a | −1.80b | 195.37a | 26.79a | −1.80b | 203.43a | 26.01a | |
| SDI65 | −1.54b | 192.73a | 26.29a | −2.03b | 209.57a | 26.69a | −1.64b | 206.38a | 26.31ab | |
| 196 | FI | −1.08a | 175.82a | 28.83a | −1.4a | 165.17a | 29.26a | −1.31a | 163.80a | 29.10a |
| SDI75 | −1.29b | 173.17a | 29.25a | −1.74b | 174.03a | 29.96a | −1.57b | 164.20a | 29.06a | |
| SDI65 | −1.28b | 172.30a | 28.95a | −1.71b | 175.37a | 30.00a | −1.53b | 158.13a | 29.15a | |
| 203 | FI | −1.67a | 152.37a | 31.83a | −1.88a | 163.55a | 30.79b | −1.69a | 161.07a | 31.26a |
| SDI75 | −1.69a | 144.55a | 31.69a | −2.01b | 145.10a | 31.63a | −1.90b | 157.03a | 30.95a | |
| SDI65 | −1.87b | 155.22a | 31.79a | −2.11b | 155.72a | 31.83a | −1.89b | 170.28a | 31.16a | |
| 210 | FI | −1.19a | 165.10a | 27.53a | −1.38a | 177.73a | 26.78a | −1.44a | 174.07a | 27.45a |
| SDI75 | −1.26a | 168.58a | 27.51a | −1.55b | 187.20a | 26.97a | −1.57a | 185.95a | 27.85a | |
| SDI65 | −1.35b | 177.20a | 27.65a | −1.50b | 182.55a | 27.67a | −1.49a | 188.20a | 27.89a | |
| 217 | FI | −1.55a | 180.50a | 29.44a | −1.73a | 179.65a | 29.20a | −1.72a | 191.52a | 29.73a |
| SDI75 | −1.80b | 167.98a | 29.47a | −1.95b | 194.50a | 29.65a | −1.97b | 184.35a | 29.81a | |
| SDI65 | −1.72b | 176.20a | 29.41a | −1.90b | 195.53a | 29.49a | −1.95b | 185.43a | 29.45a | |
| Baseline | cv. Guara | cv. Marta | cv. Lauranne | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Slope | Intercept | R2 | Slope | Intercept | R2 | Slope | Intercept | R2 | |
| NWSB | −2.71a | 5.60a | 0.82 | −2.33a | 4.14a | 0.75 | −2.48a | 4.62a | 0.78 |
| WSB75 | −2.62a | 5.65a | 0.76 | −2.45a | 4.73a | 0.75 | −2.66a | 5.75a | 0.77 |
| WSB65 | −2.58a | 5.76a | 0.74 | −2.49a | 5.01a | 0.74 | −2.62a | 5.51a | 0.77 |
| Cultivars | Slope | Intercept | R2 |
|---|---|---|---|
| Marta | −2.42 | 4.63 | 0.74 |
| Guara | −2.63 | 5.67 | 0.77 |
| Lauranne | −2.59 | 5.29 | 0.74 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Gordillo, S.; García-Tejero, I.F.; Durán Zuazo, V.H.; García Escalera, A.; Ferrera Gil, F.; Amores-Agüera, J.J.; Cárceles Rodríguez, B.; Hernández-Santana, V. Assessing the Water-Stress Baselines by Thermal Imaging for Irrigation Management in Almond Plantations under Water Scarcity Conditions. Water 2020, 12, 1298. https://doi.org/10.3390/w12051298
Gutiérrez-Gordillo S, García-Tejero IF, Durán Zuazo VH, García Escalera A, Ferrera Gil F, Amores-Agüera JJ, Cárceles Rodríguez B, Hernández-Santana V. Assessing the Water-Stress Baselines by Thermal Imaging for Irrigation Management in Almond Plantations under Water Scarcity Conditions. Water. 2020; 12(5):1298. https://doi.org/10.3390/w12051298
Chicago/Turabian StyleGutiérrez-Gordillo, Saray, Iván Francisco García-Tejero, Víctor Hugo Durán Zuazo, Amelia García Escalera, Fernando Ferrera Gil, José Juan Amores-Agüera, Belén Cárceles Rodríguez, and Virginia Hernández-Santana. 2020. "Assessing the Water-Stress Baselines by Thermal Imaging for Irrigation Management in Almond Plantations under Water Scarcity Conditions" Water 12, no. 5: 1298. https://doi.org/10.3390/w12051298
APA StyleGutiérrez-Gordillo, S., García-Tejero, I. F., Durán Zuazo, V. H., García Escalera, A., Ferrera Gil, F., Amores-Agüera, J. J., Cárceles Rodríguez, B., & Hernández-Santana, V. (2020). Assessing the Water-Stress Baselines by Thermal Imaging for Irrigation Management in Almond Plantations under Water Scarcity Conditions. Water, 12(5), 1298. https://doi.org/10.3390/w12051298