Understanding the Role of Organic Matter Cycling for the Spatio-Temporal Structure of PCBs in the North Sea


 , ,
, ,
Abstract
1. Introduction
2. Methods—Model Description
2.1. Model Components
2.1.1. Modeling Hydrodynamics and Transport with HAMSOM
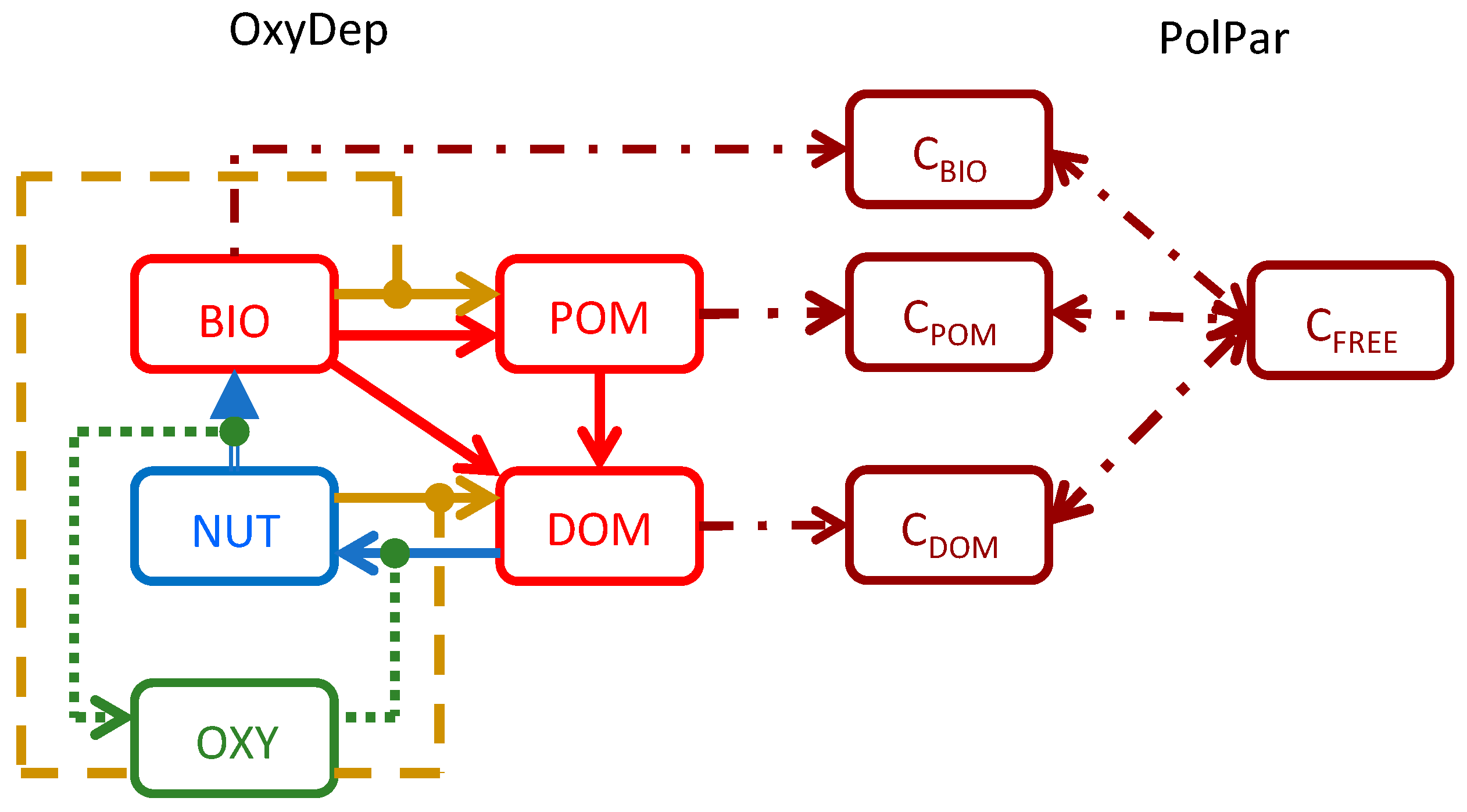
2.1.2. Modeling Biogeochemistry with OxyDep
2.1.3. Hazardous Substances Partitioning and Decay Model: PolPar
2.1.4. Degradation of the Chemicals
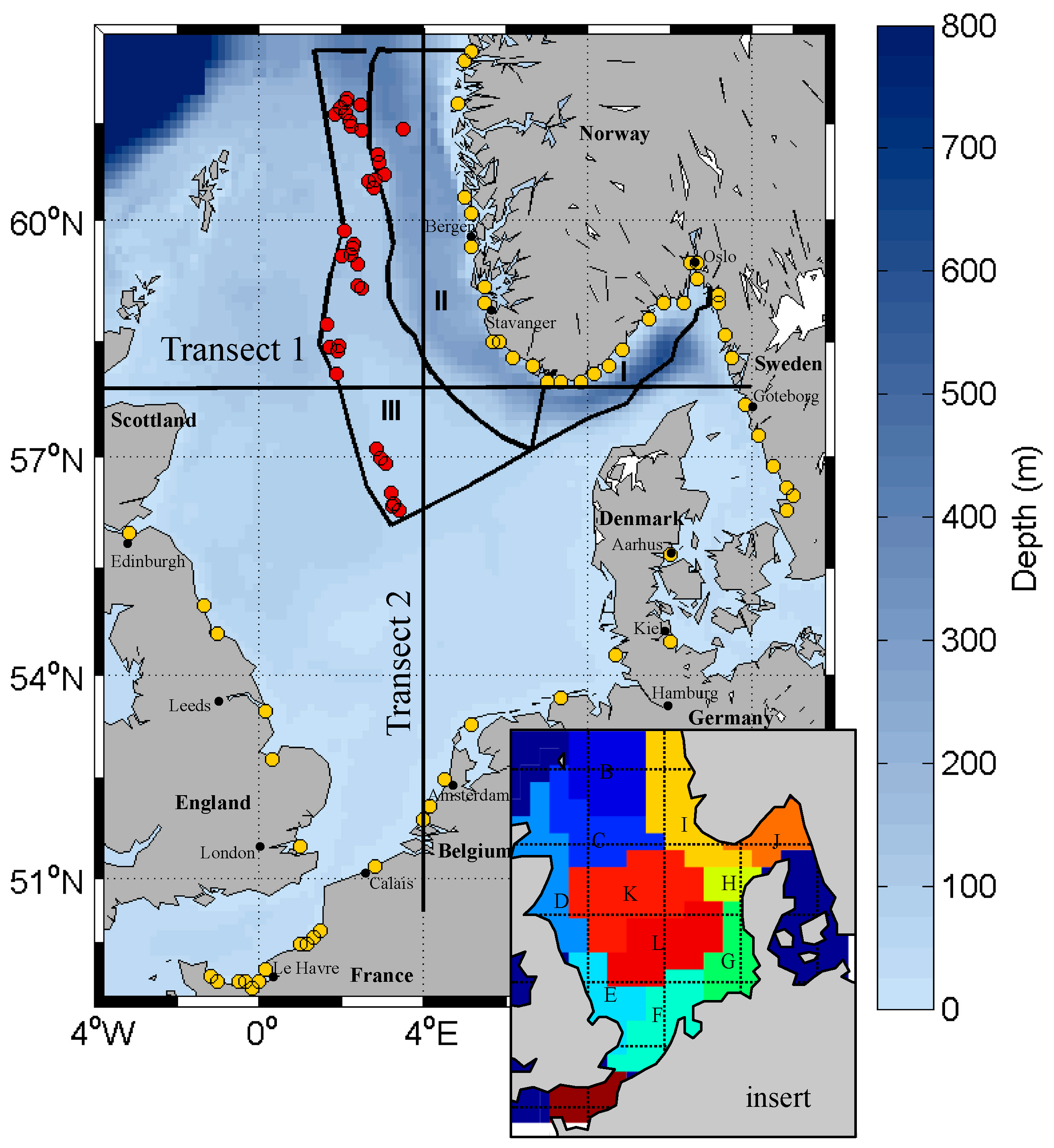
2.2. Model Setup and Boundary Conditions
2.2.1. Air–Sea Boundary
2.2.2. Sediment–Water Boundary
2.2.3. Lateral Boundaries
2.2.4. Computational Aspects
3. Results and Discussion
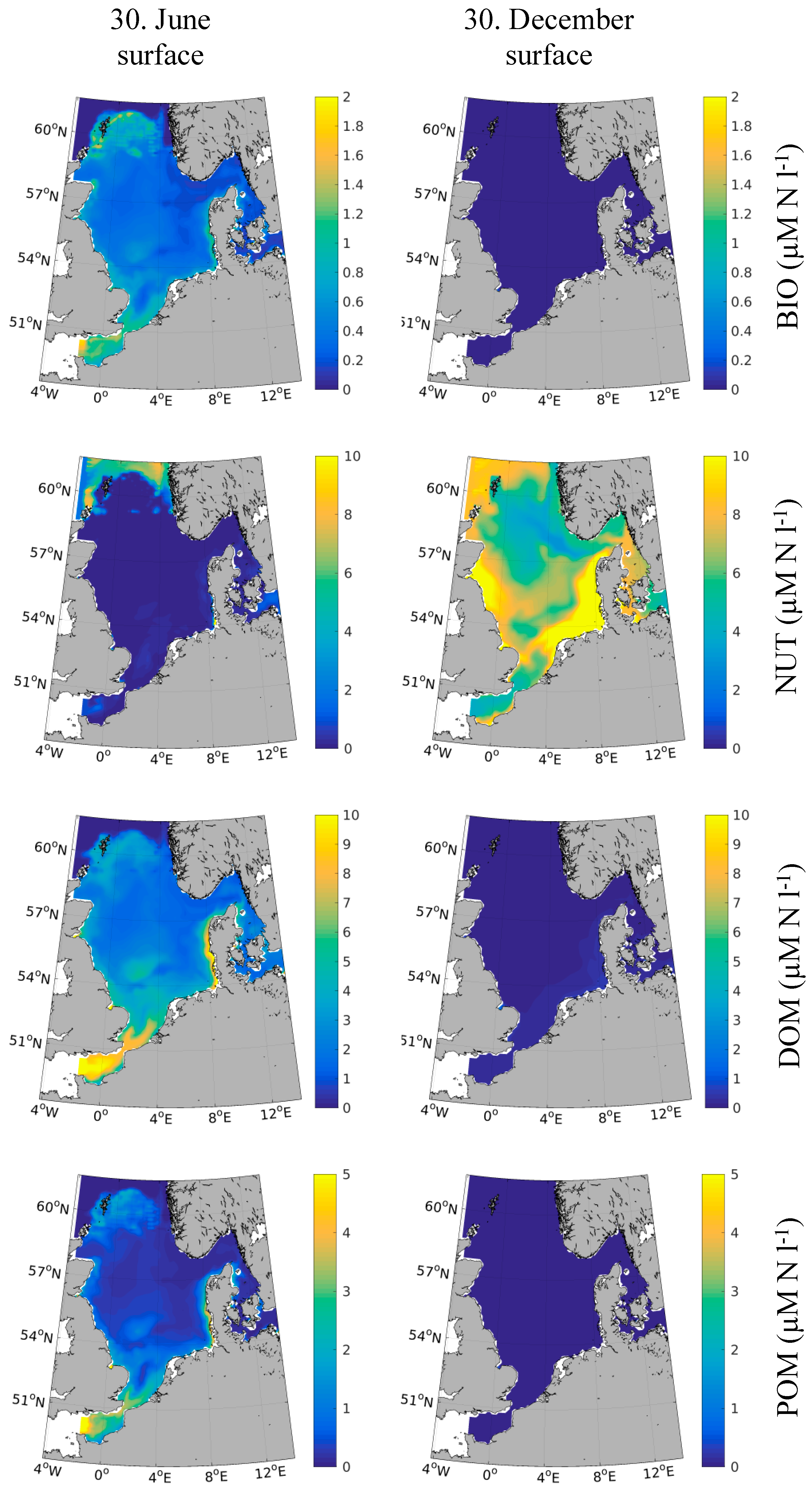
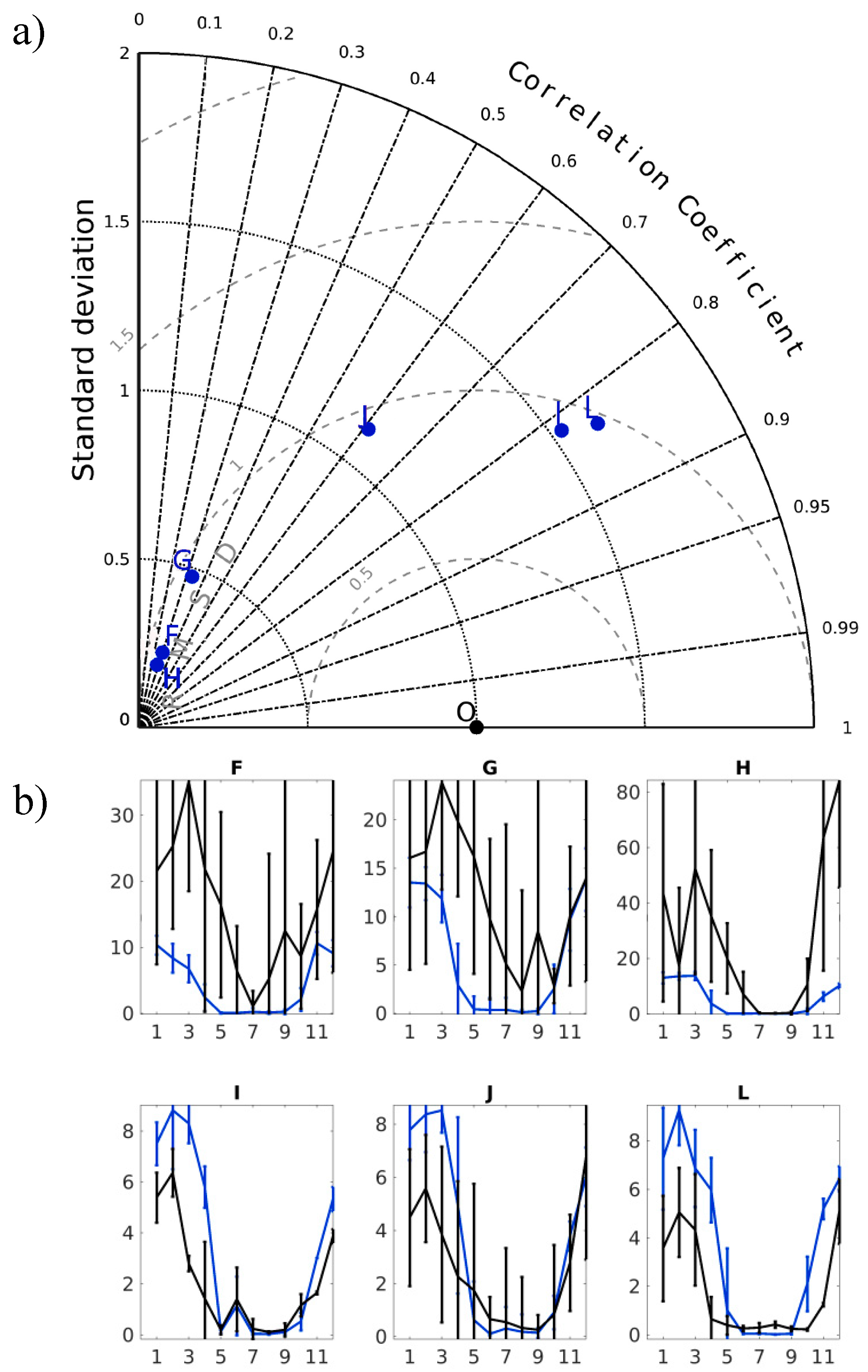
3.1. Model Performance for Biogeochemical State Variables
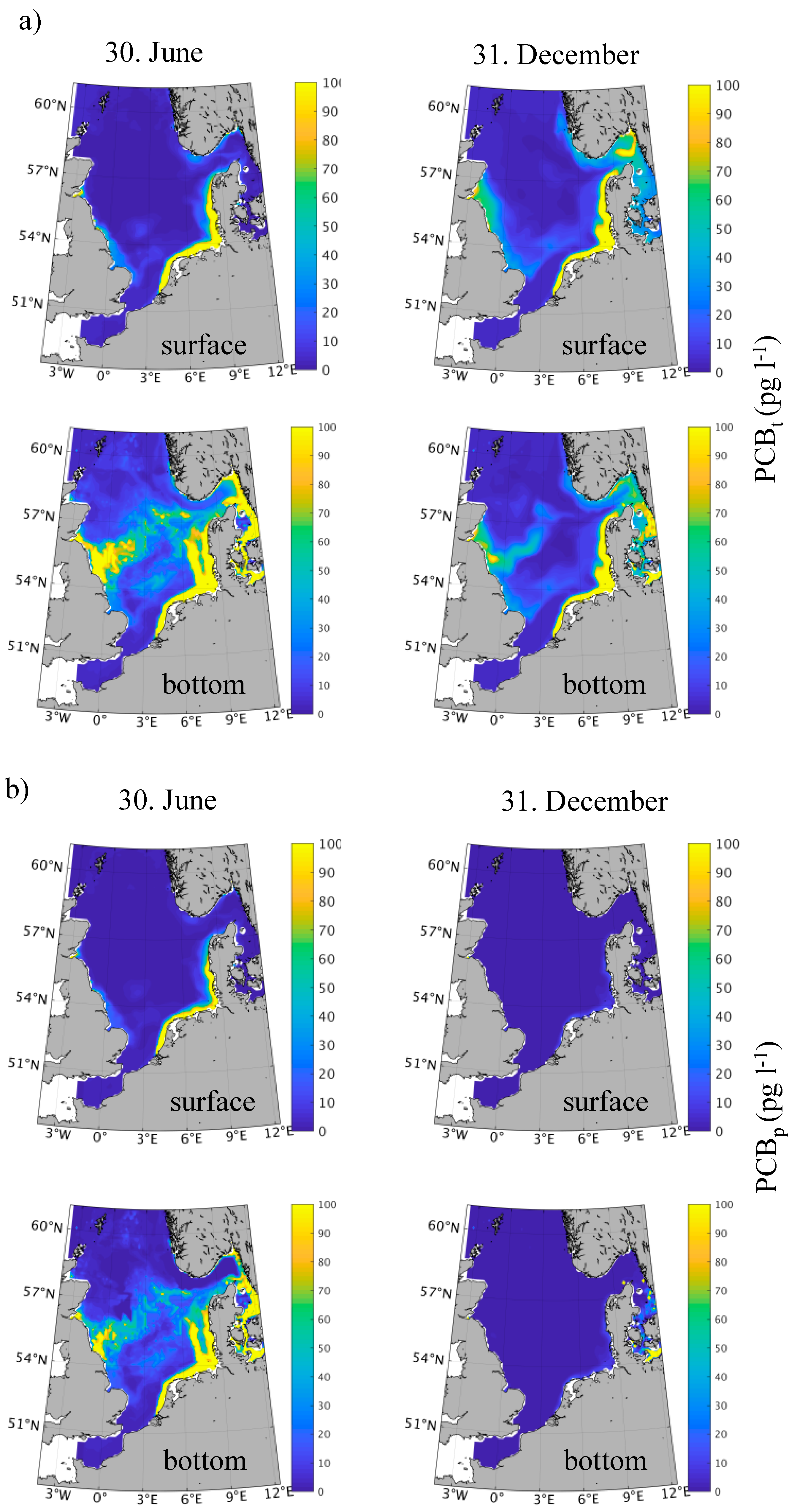
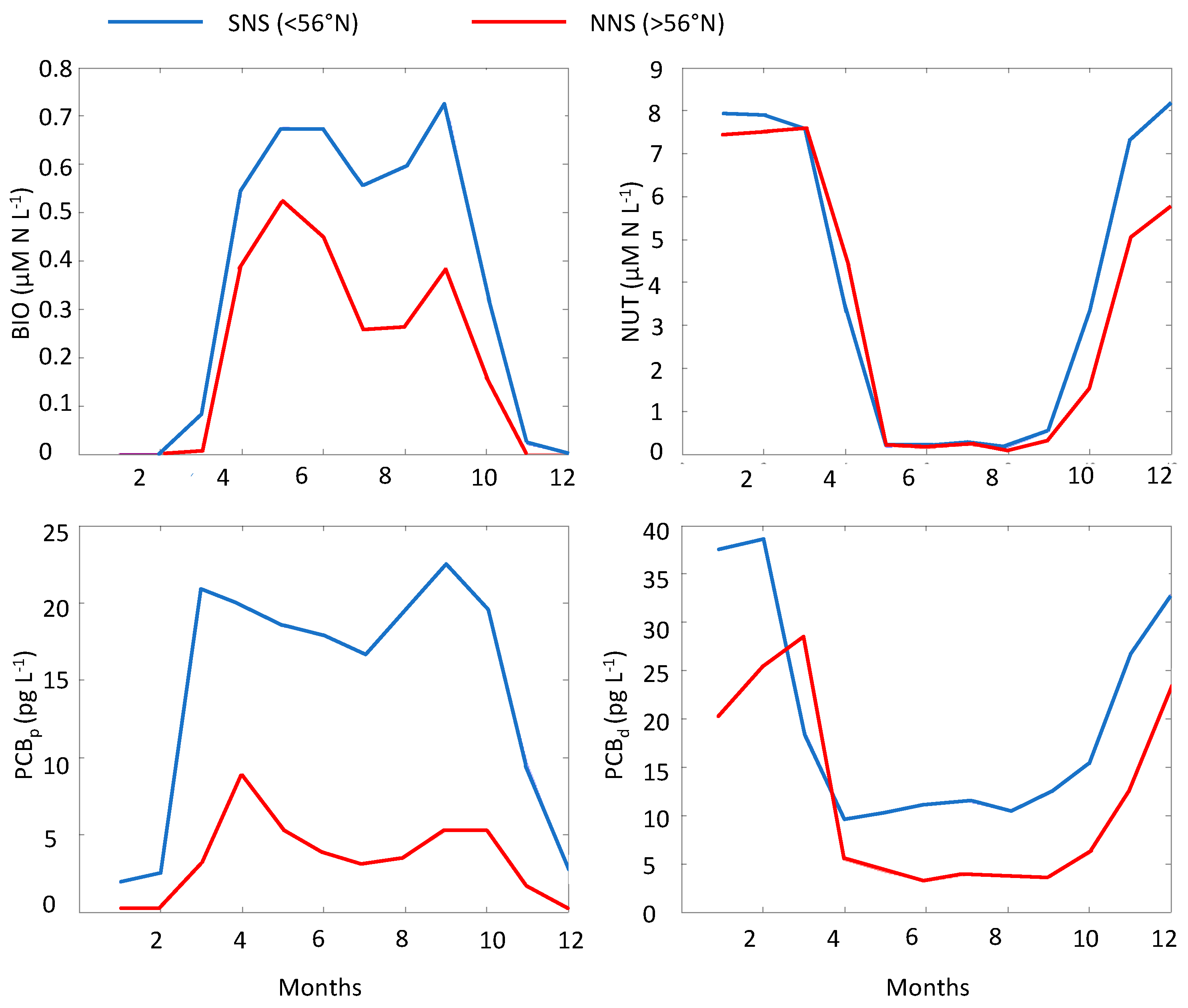
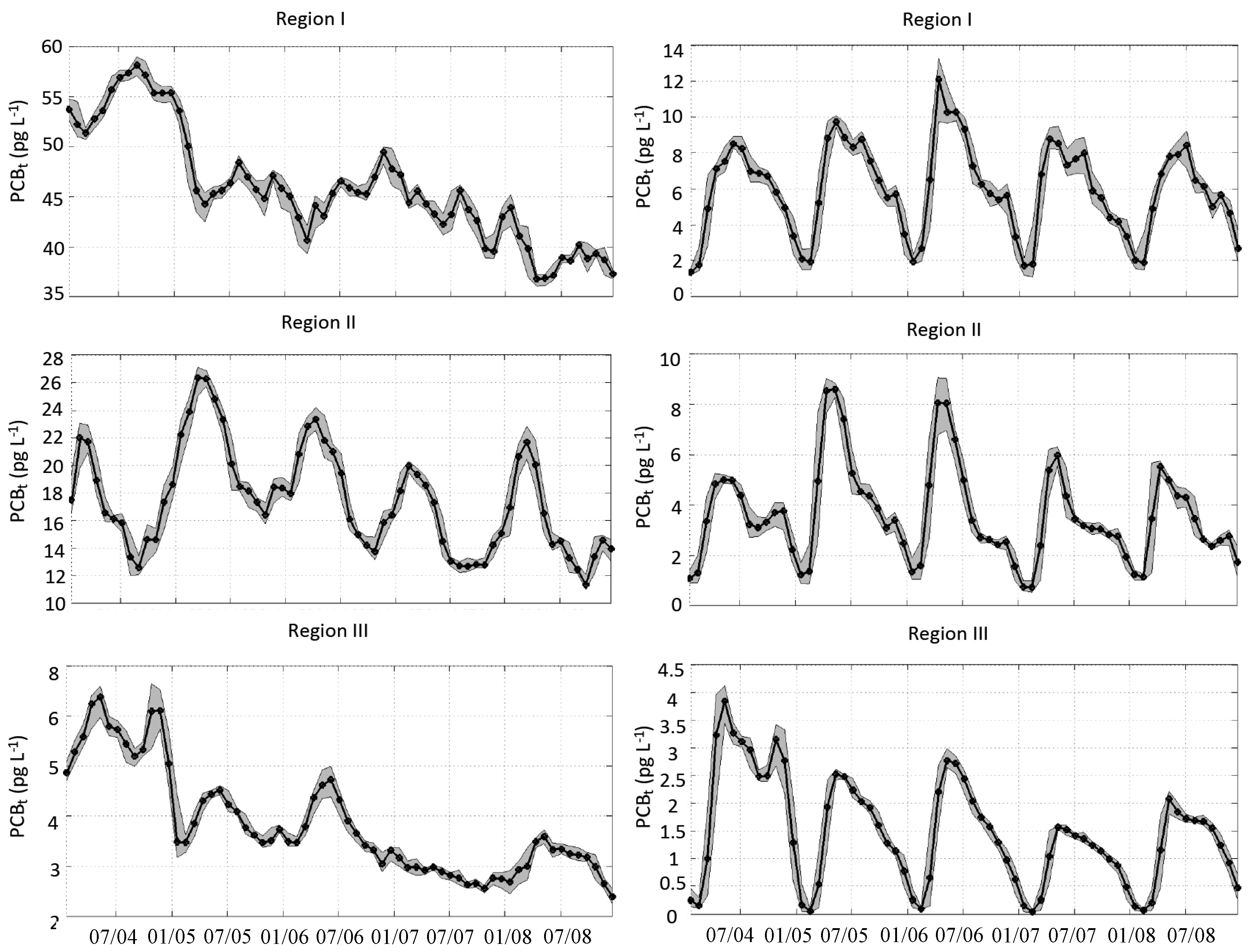
3.2. Spatial-Temporal Variability in PCB
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Notation | Value | Units | Parameter |
|---|---|---|---|
| GrowthBIO | d−1 | Specific growth rate | |
| f(I) | - | Photosynthesis dependense on irradiance | |
| f(T) | - | Photosynthesis dependense on temperature | |
| f(φ) | - | Variation of light with latitude and time | |
| f(NUT) | - | Photosynthesis dependense on nutrient | |
| KNF | 4.0 | d−1 | Maximum specific growth rate |
| I0 | 80. | W m−2 | Optimal Irradiance at the surface |
| k | 0.10 | m−1 | Extinction coefficient |
| Iopt | 25. | W m−2 | Optimal irradiance |
| φ | ° | Latitude | |
| bm | 0.12 | °C−1 | Coefficient for uptake rate dependence on t |
| cm | 1.4 | - | Coefficient for uptake rate dependence on t |
| KNUT | 0.02 | Half-saturatuib constand for uptake of NUT by BIO (squared Mikhaelis-Menten dependence) | |
| r_bio_nut | 0.05 | d−1 | Specific respiration rate |
| KBP | 0.01 | d−1 | Specific rate of mortality |
| KBD | 0.10 | d−1 | Specific rate of excretion a |
| KBPA | 0.5 | d−1 | Specific rate mortality in anoxic cond. |
| KBPC | 0.6 | d−1 | Spec.rate of addtional mortality (cannibalism) |
| BIOCan | 1. | µM N | Threshold BIO value for cannibalism |
| KPD | 0.10 | d−1 | Specific rate of POM decomposition (autolis) |
| DecayDOM | d−1 | mineralization of POM | |
| KPOM | 0.003 | d−1 | Specific rate of POM oxic decay |
| KPOMS | 0.001 | d−1 | Specific rate of POM denitrification |
| DecayDOM | d−1 | mineralization of DOM | |
| KDOM | 0.05 | d−1 | Specific rate of DOM oxic decay |
| KDOMS | 0.0005 | d−1 | Specific rate of DOM denitrification |
| Tda | 13. | - | Coefficient for dependence of decay on t |
| Bda | 20. | - | Coefficient for dependence of decay on t |
| Bu | 0.22 | d−1 m−1 | Burial coeficient for lower boundary |
| NUTDen | 1. | µM N | Threshold NUT value for denitrification |
| 20. | µM | Constant that defines the oxygen threshold | |
| COtoN | −8625 | - | O to N Redfield ratio (138/16) |
References
- Den Besten, P.J.; Valk, S.; Van Weerlee, E.; Nolting, R.F.; Postma, J.F.; Everaarts, J.M. Bioaccumulation and biomarkers in the sea star Asterias rubens (Echinodermata: Asteroidea): A North Sea field study. Mar. Environ. Res. 2001, 51, 365–387. [Google Scholar] [CrossRef]
- Jepson, P.D.; Law, R.J. Persistent pollutants, persistent threats. Science 2016, 352, 1388–1389. [Google Scholar] [CrossRef] [PubMed]
- Beyer, A.; Biziuk, M. Environmental Fate and Global Distribution of Polychlorinated Biphenyls BT—Reviews of Environmental Contamination and Toxicology Vol 201; Whitacre, D.M., Ed.; Springer US: Boston, MA, USA, 2009. [Google Scholar]
- Agudo, A.; Aronson, K.J.; Bonefeld-Jorgensen, E.C.; Cocco, P.; Cogliano, V.; Cravedi, J.-P.; Esch, H.; Fiedler, H.; Glauert, H.P.; Leon Guo, Y.-L.; et al. Polychlorinated biphenyls and polybrominated biphenyls: Vol 107. In IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; IARC Press: Lyon, France, 2016. [Google Scholar]
- Faroon, O.; Olson, J.N. Toxicological Profile for Polychlorinated Biphenyls (PCBs); Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2000.
- Mackenzie, B.R.; Almesjö, L.; Hansson, S. Fish, Fishing, and Pollutant Reduction in the Baltic Sea. Environ. Sci. Technol. 2004, 38, 1970–1976. [Google Scholar] [CrossRef] [PubMed]
- Karl, H.; Ruoff, U. Dioxins and dioxin-like PCBs in fish and fishery products on the German market. J. Verbrauch. Leb. 2008, 3, 19–27. [Google Scholar] [CrossRef]
- Kiljunen, M.; Vanhatalo, M.; Mäntyniemi, S.; Peltonen, H.; Kuikka, S.; Kiviranta, H.; Parmanne, R.; Tuomisto, J.T.; Vuorinen, P.J.; Hallikainen, A.; et al. Human dietary intake of organochlorines from Baltic herring: Implications of individual fish variability and fisheries management. Ambio 2007, 36, 257–264. [Google Scholar] [CrossRef]
- Fliedner, A.; Rüdel, H.; Knopf, B.; Lohmann, N.; Paulus, M.; Jud, M.; Pirntke, U.; Koschorreck, J. Assessment of seafood contamination under the marine strategy framework directive: Contributions of the German environmental specimen bank. Environ. Sci. Pollut. Res. 2018, 25, 26939–26956. [Google Scholar] [CrossRef]
- Wania, F.; Mackay, D. Tracking the distribution of persistent organic pollutants. Environ. Sci. Technol. 1996, 30, A390–A396. [Google Scholar] [CrossRef]
- Dachs, J.; Lohmann, R.; Ockenden, W.A.; Méjanelle, L.; Eisenreich, S.J.; Jones, K.C. Oceanic biogeochemical controls on global dynamics of persistent organic pollutants. Environ. Sci. Technol. 2002, 36, 4229–4237. [Google Scholar] [CrossRef]
- Nizzetto, L.; Lohmann, R.; Gioia, R.; Dachs, J.; Jones, K.C. Atlantic Ocean Surface Waters Buffer Declining Atmospheric Concentrations of Persistent Organic Pollutants. Environ. Sci. Technol. 2010, 44, 6978–6984. [Google Scholar] [CrossRef]
- Nizzetto, L.; Lohmann, R.; Gioia, R.; Jahnke, A.; Temme, C.; Dachs, J.; Herckes, P.; Di Guardo, A.; Jones, K.C. PAHs in air and seawater along a North-South Atlantic transect: Trends, processes and possible sources. Environ. Sci. Technol. 2008, 42, 1580–1585. [Google Scholar] [CrossRef]
- Dachs, J.; Eisenreich, S.J.; Baker, J.E.; Ko, F.-C.; Jeremiason, J.D. Coupling of phytoplankton uptake and air-water exchange of persistent organic pollutants. Environ. Sci. Technol. 1999, 33, 3653–3660. [Google Scholar] [CrossRef]
- Nizzetto, L.; Gioia, R.; Li, J.; Borgå, K.; Pomati, F.; Bettinetti, R.; Dachs, J.; Jones, K.C. Biological pump control of the fate and distribution of hydrophobic organic pollutants in water and plankton. Environ. Sci. Technol. 2012, 46, 3204–3211. [Google Scholar] [CrossRef] [PubMed]
- Larsson, P.; Okla, L.; Cronberg, G. Turnover of polychlorinated biphenyls in an oligotrophic and a eutrophic lake in relation to internal lake processes and atmospheric fallout. Can. J. Fish. Aquat. Sci. 1998, 55, 1926–1937. [Google Scholar] [CrossRef]
- Berglund, O.; Larsson, P.; Ewald, G.; Okla, L. Influence of trophic status on PCB distribution in lake sediments and biota. Environ. Pollut. 2001, 113, 199–210. [Google Scholar] [CrossRef]
- Lohmann, R.; Dachs, J. Deposition of Dissolved and Particulate-Bound Chemicals from the Surface Ocean. In Handbook of Chemical Mass Transport in the Environment; Thibodeaux, L.J., Mackay, D., Eds.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Morales, L.; Dachs, J.; Fernández-Pinos, M.C.; Berrojalbiz, N.; Mompean, C.; González-Gaya, B.; Jiménez, B.; Bode, A.; Ábalos, M.; Abad, E. Oceanic Sink and Biogeochemical Controls on the Accumulation of Polychlorinated Dibenzo-p-dioxins, Dibenzofurans, and Biphenyls in Plankton. Environ. Sci. Technol. 2015, 49, 13853–13861. [Google Scholar] [CrossRef]
- Lohmann, R.; Jurado, E.; Pilson, M.E.Q.; Dachs, J. Oceanic deep water formation as a sink of persistent organic pollutants. Geophys. Res. Lett. 2006, 33, 2–5. [Google Scholar] [CrossRef]
- Sinkkonen, S.; Paasivirta, J. Degradation half-life times of PCDDs, PCDFs and PCBs for environmental fate modeling. Chemosphere 2000, 40, 943–949. [Google Scholar] [CrossRef]
- Ma, J.; Hung, H.; Tian, C.; Kallenborn, R. Revolatilization of persistent organic pollutants in the Arctic induced by climate change. Nat. Clim. Chang. 2011, 1, 255–260. [Google Scholar] [CrossRef]
- Schrum, C.; Backhaus, J.O. Sensitivity of atmosphere-ocean heat exchange and heat content in the North Sea and the Baltic Sea. Tellus Ser. A Dyn. Meteorol. Oceanogr. 1999, 51, 526–549. [Google Scholar] [CrossRef]
- Ilyina, T.; Pohlmann, T.; Lammel, G.; Sündermann, J. A fate and transport ocean model for persistent organic pollutants and its application to the North Sea. J. Mar. Syst. 2006, 63, 1–19. [Google Scholar] [CrossRef]
- Ilyina, T. The fate of persistent organic pollutants in the North Sea: Multiple year model simulations of [gamma]-HCH, [alpha]-HCH and PCB 153. In Hamburg Studies on Maritime Affairs; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- O’Driscoll, K.; Ilyina, T.; Pohlmann, T.; Mayer, B.; Damm, P. Modelling the fate of persistent organic pollutants (POPs) in the North Sea system. Procedia Environ. Sci. 2011, 6, 169–179. [Google Scholar] [CrossRef][Green Version]
- Schrum, C.; St John, M.; Alekseeva, I. ECOSMO, a coupled ecosystem model of the North Sea and Baltic Sea: Part II. Spatial-seasonal characteristics in the North Sea as revealed by EOF analysis. J. Mar. Syst. 2006, 61, 100–113. [Google Scholar] [CrossRef]
- Daewel, U.; Schrum, C. Simulating long-term dynamics of the coupled North Sea and Baltic Sea ecosystem with ECOSMO II: Model description and validation. J. Mar. Syst. 2013, 119–120, 30–49. [Google Scholar] [CrossRef]
- Yakushev, E.V.; Debolskaya, E.I.; Kuznetsov, I.S.; Staalstrøm, A. Modelling of the Meromictic Fjord Hunnbunn (Norway) with an Oxygen Depletion Model (OxyDep). In Chemical Structure of Pelagic Redox Interfaces: Observation and Modeling, Hdb Env Chem 22; Yakushev, E.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Oguz, T.; Ducklow, H.W.; Malanotte-Rizzoli, P.; Murray, J.W. Simulations of the Black Sea pelagic ecosystem by 1-D, vertically resolved, physical–biochemical models. Fish. Oceanogr. 1998, 7, 300–304. [Google Scholar] [CrossRef]
- Savchuk, O.; Wulff, F. Biogeochemical trans-formations of nitrogen and phosphorus in the marine environment—Coupling hydrodynamic and biogeochemical processes in models for the Baltic Proper. Syst. Ecol. Contrib. Stock. Univ. 1996, 2, 79. [Google Scholar]
- Barthel, K.; Daewel, U.; Pushpadas, D.; Schrum, C.; Årthun, M.; Wehde, H. Resolving frontal structures: On the payoff using a less diffusive but computationally more expensive advection scheme. Ocean Dyn. 2012, 62, 1457–1470. [Google Scholar] [CrossRef]
- Schrum, C. Thermohaline stratification and instabilities at tidal mixing fronts: Results of an eddy resolving model for the German Bight. Cont. Shelf Res. 1997, 17, 689–716. [Google Scholar] [CrossRef]
- Backhaus, J.O.; Hainbucher, D. A Finite Difference General Circulation Model for Shelf Seas and Its Application to Low Frequency Variability on the North European Shelf. In Three-Dimensional Models of Marine and Estuarine Dynamics; Nihoul, J.C.J., Jamart, B.M., Eds.; Elsevier Oceanography Series: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Janssen, F.; Schrum, C.; Hübner, U.; Backhaus, J.O. Uncertainty analysis of a decadal simulation with a regional ocean model for the North Sea and Baltic Sea. Clim. Res. 2001, 18, 55–62. [Google Scholar] [CrossRef]
- Daewel, U.; Schrum, C. Low frequency variability in North Sea and Baltic Sea identified through simulations with the 3-d coupled physical-biogeochemical model ECOSMO. Earth Syst. Dyn. 2017, 8, 801–815. [Google Scholar] [CrossRef]
- Kalnay, E.; Kanamitsu, M.; Kistler, R.; Collins, W.; Deaven, D.; Gandin, L.; Iredell, M.; Saha, S.; White, G.; Woollen, J.; et al. The NCEP/NCAR 40-year reanalysis project. Bull. Am. Meteorol. Soc. 1996, 77, 437–471. [Google Scholar] [CrossRef]
- Thomas, G.O. Polychlorinated Biphenyls; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Schwarzenbach, R.P.; Gschwend, P.M.; Imboden, D.M. Environmental Organic Chemistry, 2nd ed.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2003. [Google Scholar]
- Mackay, D. Multimedia Environmental Models: The Fugacity Approach, 2nd ed.; Lewis Publishers: Boca Raton, FL, USA, 2001. [Google Scholar]
- Meng, Q.; Chu, S.; Xu, X. Sorption phenomena of PCBs in environment. Chin. Sci. Bull. 2001, 46, 89–97. [Google Scholar] [CrossRef]
- IPCS. Environmental Health Criteria 123; Alpha-and BetaHexachlorocyclohexanes; World Health Organization: Geneva, Switzerland, 1992. [Google Scholar]
- Yang, Z.Y.; Greenstein, D.; Zeng, E.Y.; Maruya, K.A. Determination of poly (dimethyl)siloxane-water partition coefficients for selected hydrophobic organic chemicals using 14C-labeled analogs. J. Chromatogr. A 2007, 1148, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Gupta, G.; Baker, J. Photodegradation of polychlorinated biphenyl congeners using simulated sunlight and diethylamine. Chemosphere 1995, 31, 3323–3344. [Google Scholar] [CrossRef]
- Chen, L.; Tang, X.; Shen, C.; Chen, C.; Chen, Y. Photosensitized degradation of 2,4′,5-trichlorobiphenyl (PCB 31) by dissolved organic matter. J. Hazard. Mater. 2011, 201–202, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lammel, G.; Meixner, F.X.; Vrana, B.; Efstathiou, C.I.; Kohoutek, J.; Kukučka, P.; Mulder, M.D.; Přibylová, P.; Prokeš, R.; Rusina, T.P.; et al. Bidirectional air–sea exchange and accumulation of POPs (PAHs, PCBs, OCPs and PBDEs) in the nocturnal marine boundary layer. Atmos. Chem. Phys. 2016, 16, 6381–6393. [Google Scholar] [CrossRef]
- Gusev, A.; Dutchak, S.; Rozovskaya, O.; Shatalov, V.; Sokovykh, V.; Vulykh, N. Persistent Organic Pollutants in the Environment. Available online: https://www.researchgate.net/publication/288851152_Persistent_organic_pollutants_in_the_environment (accessed on 14 March 2020).
- Schneider, B.; Nausch, G.; Kubsch, H.; Petersohn, I. Accumulation of total CO2 during stagnation in the Baltic Sea deep water and its relationship to nutrient and oxygen concentrations. Mar. Chem. 2002, 77, 277–291. [Google Scholar] [CrossRef]
- De Haas, H.; Boer, W.; Van Weering, T.C.E. Recent sedimentation and organic carbon burial in a shelf sea: The North Sea. Mar. Geol. 1997, 144, 131–146. [Google Scholar] [CrossRef]
- Savchuk, O.P.; Wulff, F. Long-term modeling of large-scale nutrient cycles in the entire Baltic Sea. Hydrobiologia 2009, 629, 209–224. [Google Scholar] [CrossRef]
- Orlanski, I. A simple boundary condition for unbounded hyperbolic flows. J. Comput. Phys. 1976, 21, 251–269. [Google Scholar] [CrossRef]
- Green, N.W.; Heldal, H.E.; Måge, A.; Aas, W.; Gafvert, T.; Schrum, C.; Boitsov, S.; Breivik, K.; Iosjpe, M.; Yakushev, E.; et al. Tilførselsprogrammet 2010 Overvåking av Tilførsler og Miljøtilstand i Nordsjøen, Statlig Program for Forurensningsovervåking Rapportnr; Klima og forurensningsdirektoratet (Klif): Oslo, Norway, 2011.
- Suratman, S.; Jickells, T.; Weston, K.; Fernand, L. Seasonal variability of inorganic and organic nitrogen in the North Sea. Hydrobiologia 2008, 610, 83–98. [Google Scholar] [CrossRef]
- Føyn, L.; Hagebø, M. Nutrients and Hydrography in the North Sea Autumn 1998; Report from a cruise with R/V “Johan Hjort” 19 October–26 November 1998; Institute of Marine Research: Bergen, Norway, 1998. [Google Scholar]
- Taylor, K.E. Summarizing multiple aspects of model performance in a single diagram. J. Geophys. Res. 2001, 106, 7183–7192. [Google Scholar] [CrossRef]
- Zhao, C.; Daewel, U.; Schrum, C. Tidal impacts on primary production in the North Sea. Earth Syst. Dyn. Discuss. 2019, 10, 287–317. [Google Scholar] [CrossRef]
- Richardson, K.; Visser, A.W.; Pedersen, F.B. Subsurface phytoplankton blooms fuel pelagic production in the North Sea. J. Plankton Res. 2000, 22, 1663–1671. [Google Scholar] [CrossRef]
- Gerofke, A.; Kömp, P.; McLachlan, M.S. Bioconcentration of persistent organic pollutants in four species of marine phytoplankton. Environ. Toxicol. Chem. 2005, 24, 2908–2917. [Google Scholar] [CrossRef]
- Fellman, J.; Spencer, R.; Raymond, P.; Pettit, N.; Skrzypek, G.; Hernes, P.; Grierson, P. Dissolved organic carbon biolability decreases along with its modernization in fluvial networks in an ancient landscape. Ecology 2014, 95, 2622–2632. [Google Scholar] [CrossRef]
- EEC Council Directive Relating to the Quality of Water Intended for Human Consumption, 80/778/EEC No. L229; Council of the European Communities: Brussels, Belgium, 1980.
- ICES. Report of the Working Group on Biological Effects of Contaminents (WGBEC); 31 March–4 April, Sète, France; ICES: Sète, France, 2008. [Google Scholar]
- Sonne, C. Health effects from long-range transported contaminants in Arctic top predators: An integrated review based on studies of polar bears and relevant model species. Environ. Int. 2010, 36, 461–491. [Google Scholar] [CrossRef]
- Hufnagl, M.; Peck, M.A.; Nash, R.D.M.; Pohlmann, T.; Rijnsdorp, A.D. Changes in potential North Sea spawning grounds of plaice (Pleuronectes platessa L.) based on early life stage connectivity to nursery habitats. J. Sea Res. 2013, 84, 26–39. [Google Scholar] [CrossRef]
- Delbeke, K.; Joiris, C. Accumulation mechanism and geographical distribution of PCBs in the North Sea. Océanis 1988, 14, 399–410. [Google Scholar]
- Van Zoest, R.; Van Eck, G.T.M. Behavior of selected PCBs, PAHs, gHCH in the Sheldt estuary. Neth. J. Aquat. Ecol. 1993, 23, 301–308. [Google Scholar] [CrossRef]
- Huhnerfuss, H.; Bester, K.; Landgraff, O.; Pohlmann, T.; Selke, T. Annual balances of hexachlorocyclohexanes, polychlorinated biphenyls and triazines in the German Bight. Mar. Pollut. Bull. 1997, 34, 419–426. [Google Scholar] [CrossRef]
- Dias-Ferreira, C.; Pato, R.L.; Varejão, J.B.; Tavares, A.O.; Ferreira, A.J.D. Heavy metal and PCB spatial distribution pattern in sediments within an urban catchment—contribution of historical pollution sources. J. Soils Sediments 2016, 16, 2594–2605. [Google Scholar] [CrossRef]
- Ferreira, C.; Walsh, R.; Ferreira, A. Degradation in Urban Areas. Curr. Opin. Environ. Sci. Heal. 2018, 5, 19–25. [Google Scholar] [CrossRef]
- Mai, C.; Theobald, N.; Hühnerfuss, H.; Lammel, G. Persistent organochlorine pesticides and polychlorinated biphenyls in air of the North Sea region and air-sea exchange. Environ. Sci. Pollut. Res. 2016, 23, 23648–23661. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Bull, D.; Petrick, G.; Bruhn, R.; Duinker, J. Chlorobiphenyls (PCB) and PAHs in water masses of the northern North Atlantic. Mar. Chem. 1998, 61, 101–114. [Google Scholar] [CrossRef]
- Nilsen, B.M.; Frantzen, S.; Nedreaas, K.; Julshamn, K. Basisundersøkelse av Fremmedstoffer i Blåkveite (Reinhardtius Hippoglossoides). Available online: https://nifes.hi.no/wp-content/uploads/sluttrapport_basisundersokelse_blakveite_16_april_2010-.pdf (accessed on 13 March 2020).
- Karl, H.; Lahrssen-Wiederholt, M. Dioxin and dioxin-like PCB levels in cod-liver and -muscle from different fishing grounds of the North- and Baltic Sea and the North Atlantic. J. Verbrauch. Leb. 2009, 4, 247–255. [Google Scholar] [CrossRef]
- Julshamn, K.; Frantzen, S.; Valdersnes, S.; Nilsen, B.; Maage, A.; Nedreaas, K. Concentrations of mercury, arsenic, cadmium and lead in Greenland halibut (Reinhardtius hippoglossoides) caught off the coast of northern Norway. Mar. Biol. Res. 2011, 7, 733–745. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daewel, U.; Yakushev, E.V.; Schrum, C.; Nizzetto, L.; Mikheeva, E. Understanding the Role of Organic Matter Cycling for the Spatio-Temporal Structure of PCBs in the North Sea. Water 2020, 12, 817. https://doi.org/10.3390/w12030817
Daewel U, Yakushev EV, Schrum C, Nizzetto L, Mikheeva E. Understanding the Role of Organic Matter Cycling for the Spatio-Temporal Structure of PCBs in the North Sea. Water. 2020; 12(3):817. https://doi.org/10.3390/w12030817
Chicago/Turabian StyleDaewel, Ute, Evgeniy V. Yakushev, Corinna Schrum, Luca Nizzetto, and Elena Mikheeva. 2020. "Understanding the Role of Organic Matter Cycling for the Spatio-Temporal Structure of PCBs in the North Sea" Water 12, no. 3: 817. https://doi.org/10.3390/w12030817
APA StyleDaewel, U., Yakushev, E. V., Schrum, C., Nizzetto, L., & Mikheeva, E. (2020). Understanding the Role of Organic Matter Cycling for the Spatio-Temporal Structure of PCBs in the North Sea. Water, 12(3), 817. https://doi.org/10.3390/w12030817