Influence of Local Habitat and Climatic Factors on the Distribution of Fish Species in the Tonle Sap Lake


Abstract
1. Introduction
2. Materials and Methods
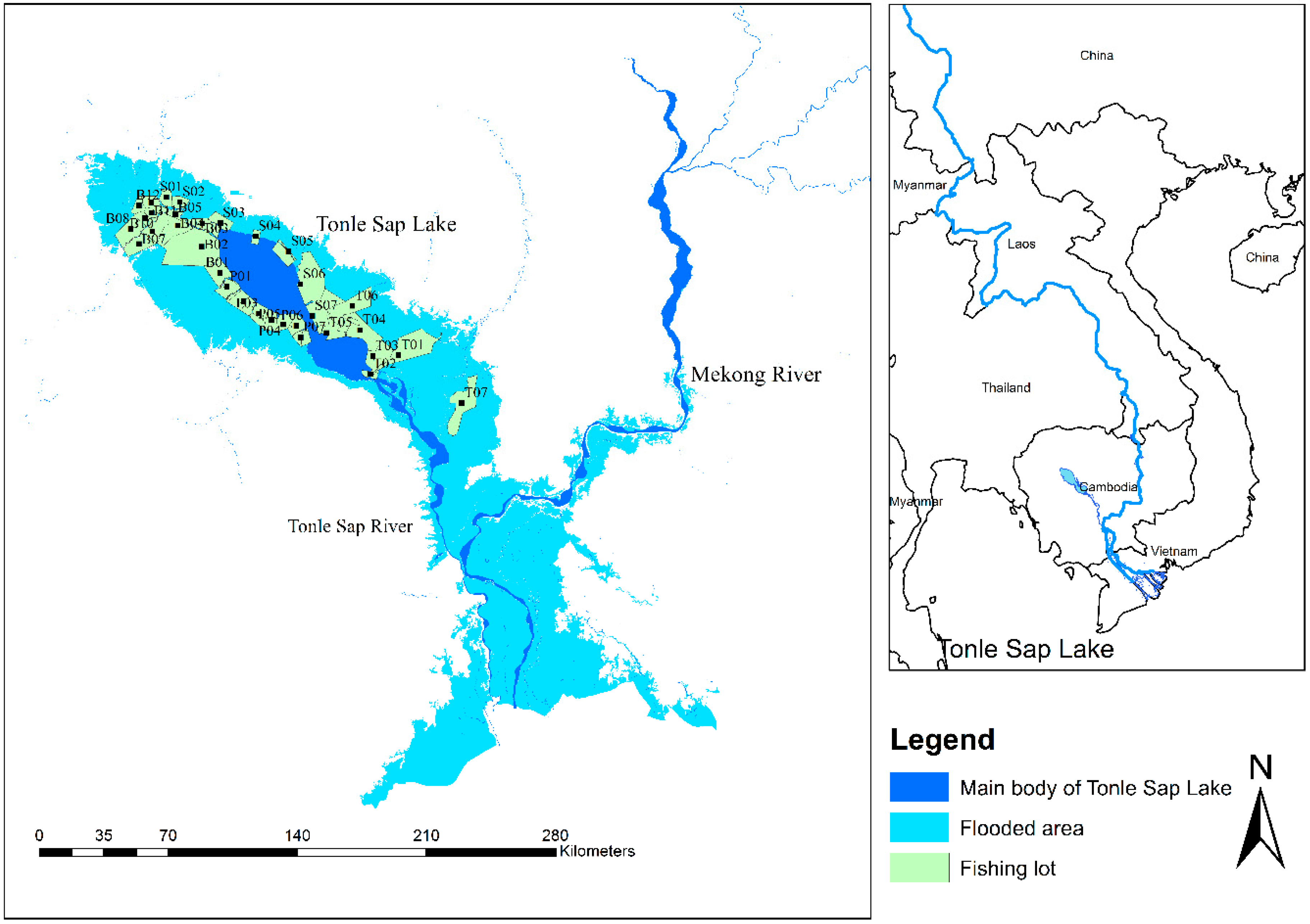
2.1. Study Area
2.2. Data Collection
2.3. Data Preparation
2.4. Statistical Analyses
3. Results
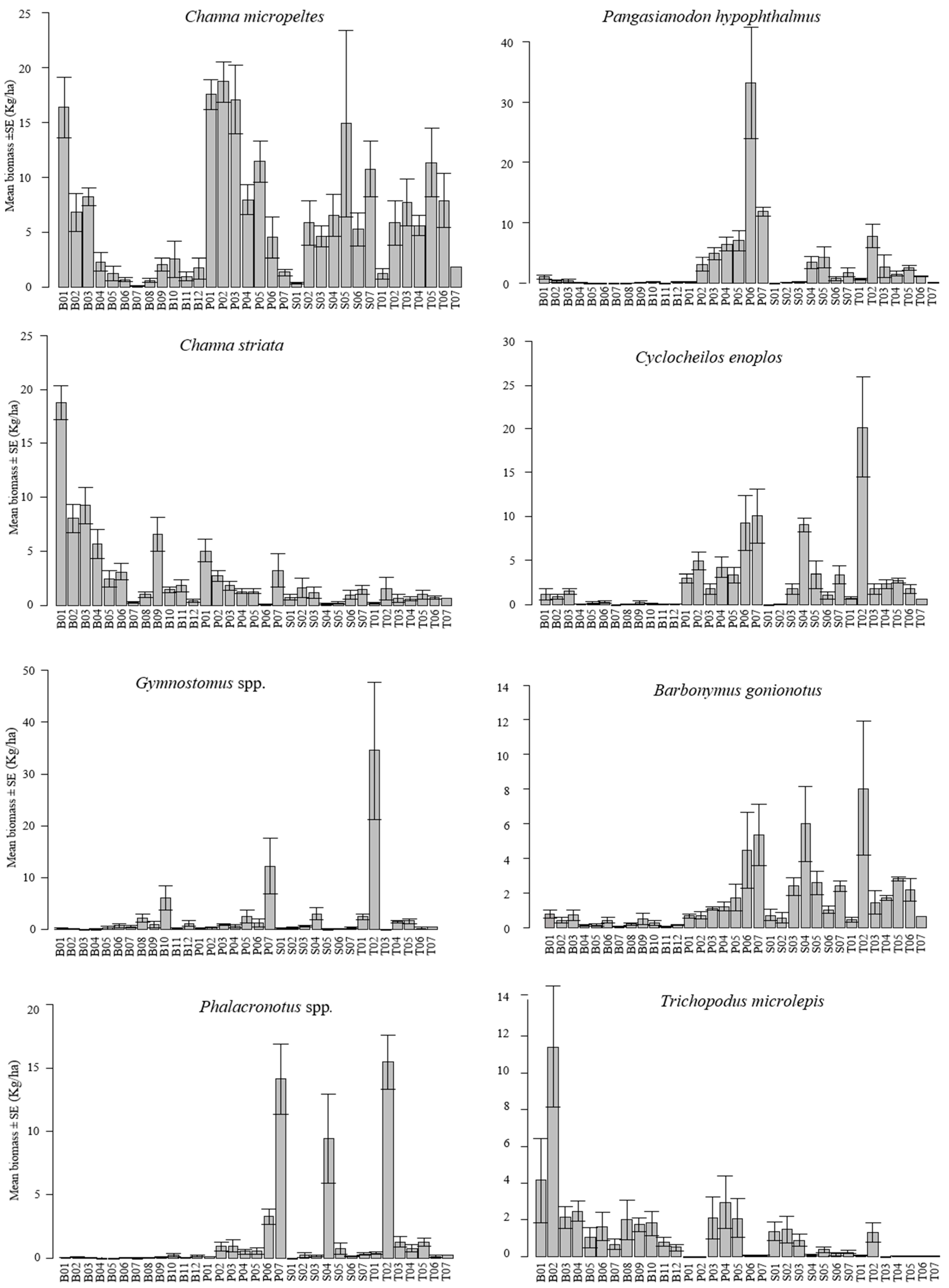
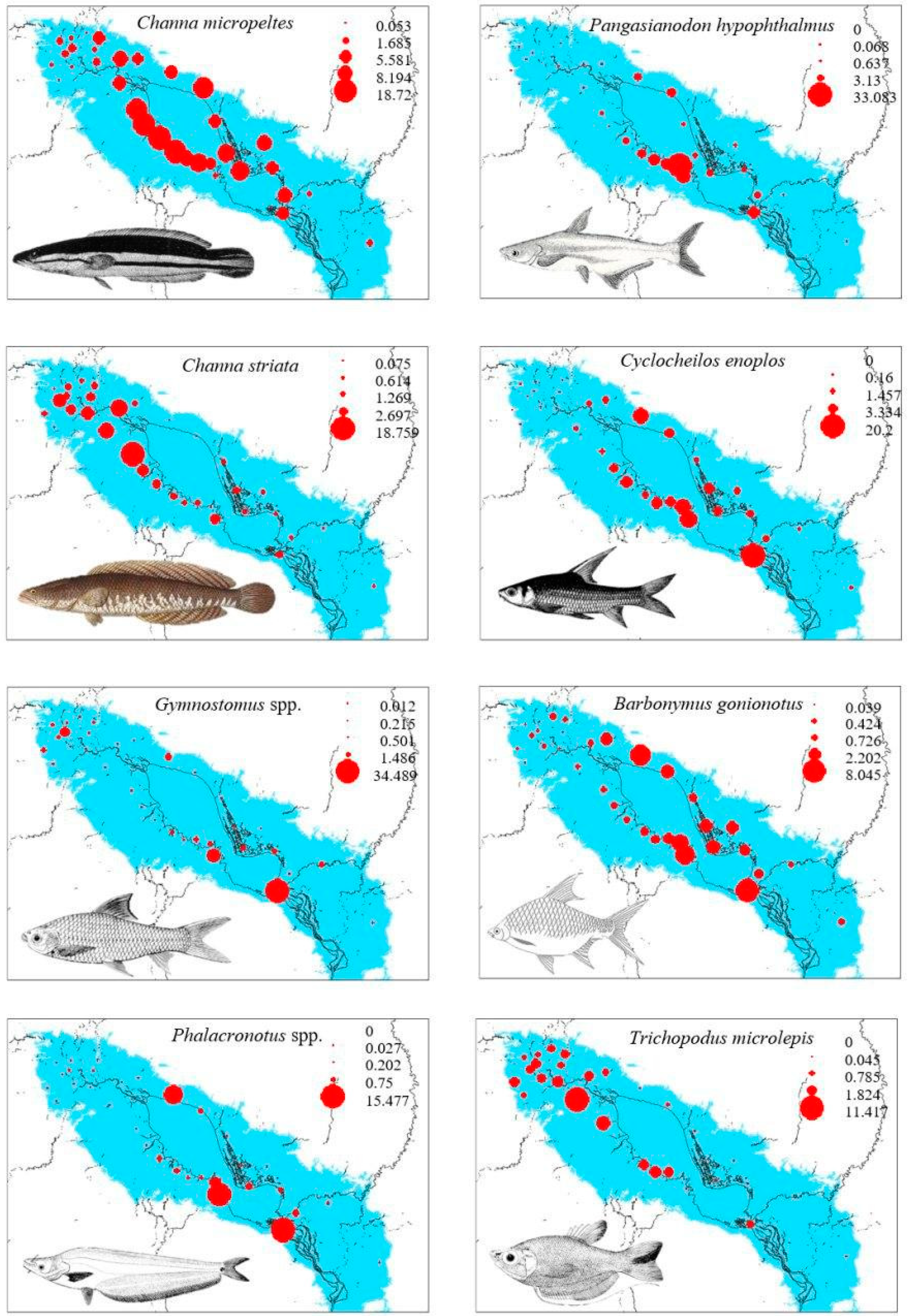
3.1. Spatial Distribution
3.2. Relationships between Fish Distribution and Local Habitat and Climatic Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mekong River Commission (MRC). Overview of the Hydrology of the Mekong Basin; Red Plough International Co. Ltd., Ed.; Mekong River Commission: Vientiane, Laos, 2005; ISSN 17283248. [Google Scholar]
- Allen, D.J.; Smith, K.G.; Darwall, W.R.T (Compilers). The Status and Distribution of Freshwater Biodiversity in Indo-Burma; IUCN: Cambridge, UK; Gland, Switzerland, 2012; ISBN 9782831714240. [Google Scholar]
- Brosse, S.; Beachard, O.; Blanchet, S.; Durr, H.H.; Grenouillet, G.; Hugueny, B.; Lauzeral, C.; Leprieur, F.; Tedesco, P.A.; Villeger, S.; et al. Fish-SPRICH: A database of freshwater fish species richness throughout the World. Hydrobiologia 2013, 700, 343–349. [Google Scholar] [CrossRef]
- Baran, E. Cambodian Inland Fisheries: Facts, Figures and Context; WorldFish Center: Penang, Malaysia; Inland Fisheries Research and Development Institute: Phnom Penh, Cambodia, 2005; ISBN 983-2346-38-X. [Google Scholar]
- Hortle, K.G. Consumption and the Yield of Fish and Other Aquatic Animals from the Lower Mekong Basin; Mekong River Commission: Vientiane, Laos, 2007; 87p. [Google Scholar]
- Mekong River Commission (MRC). State of the Basin Report 2010; Mekong River Commission: Vientiane, Laos, 2010; ISBN 978-993-208-057-1. [Google Scholar]
- Hortle, K.G.; Bamrungrach, P. Fisheries Habitat and Yield in the Lower Mekong Basin; Mekong River Commission: Phnom Penh, Cambodia, 2015; ISSN 1683-1489. [Google Scholar]
- Ahmed, M.; Hap, N.; Ly, V.; Tiongco, M. Socioeconomic Assessment of Freshwater Capture Fisheries in Cambodia: Report on a Household Survey; Mekong River Commission: Phnom Penh, Cambodia, 1998; 186p. [Google Scholar]
- Lieng, S.; Zalinge, V.N. Fish Yield Estimation in the Floodplains of the Tonle Sap Great Lake and River; Mekong River Commission: Phnom Penh, Cambodia, 2001. [Google Scholar]
- Baran, E.; So, N.; Degen, P.; Chen, X.-Y.; Starr, P. Updated information on fish and fisheries in the Mekong Basin. Catch Cult. 2013, 19, 24–25. [Google Scholar]
- UNESCO. Tonle Sap Biosphere Reserve. Available online: http://www.unesco.org/new/en/phnompenh/natural-sciences/biosphere-reserves/tonle-sap-biosphere-reserve/ (accessed on 12 January 2018).
- Ngor, P.B.; Grenouillet, G.; Phem, S.; So, N.; Lek, S. Spatial and temporal variation in fish community structure and diversity in the largest tropical flood-pulse system of South-East Asia. Ecol. Freshw. Fish 2018, 27, 1087–1100. [Google Scholar] [CrossRef]
- Campbell, I.C.; Poole, C.; Giesen, W.; Valbo-Jorgensen, J. Species diversity and ecology of Tonle Sap Great Lake, Cambodia. Aquat. Sci. 2006, 68, 355–373. [Google Scholar] [CrossRef]
- Enomoto, K.; Ishikawa, S.; Hori, M.; Hort, S.; Srun, L.S.; Nao, T.; Kurokura, H. Data mining and stock assessment of fisheries resources in Tonle Sap Lake, Cambodia. Fish. Sci. 2011, 77, 713–722. [Google Scholar] [CrossRef]
- Yen, N.T.H.; Sunada, K.; Oishi, S.; Ikejima, K.; Iwata, T. Stock assessment and fishery management of Henicorhynchus spp., Cyclocheilichthys enoplos and Channa micropeltes in Tonle Sap Great Lake, Cambodia. J. Gt. Lakes Res. 2009, 35, 169–174. [Google Scholar] [CrossRef]
- Ngor, P.B.; McCann, K.S.; Grenouillet, G.; So, N.; McMeans, B.C.; Fraser, E.; Lek, S. Evidence of indiscriminate fishing effects in one of the world’s largest inland fisheries. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- McCann, K.S.; Gellner, G.; McMeans, B.C.; Deenik, T.; Holtgrieve, G.; Rooney, N.; Hannah, L.; Cooperman, M.; Nam, S.; Giacomini, H. Food webs and the sustainability of indiscriminate fisheries. Can. J. Fish. Aquat. Sci. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- KC, K.B.; Bond, N.; Fraser, E.D.G.; Elliott, V.; Farrell, T.; McCann, K.; Rooney, N.; Bieg, C. Exploring tropical fisheries through fishers’ perceptions: Fishing down the food web in the Tonlé Sap, Cambodia. Fish. Manag. Ecol. 2017, 24, 452–459. [Google Scholar] [CrossRef]
- Ngor, P.B.; Oberdorff, T.; Phen, C.; Baehr, C.; Grenouillet, G.; Lek, S. Fish assemblage responses to flow seasonality and predictability in a tropical flood pulse system. Ecosphere 2018, 9, e02366. [Google Scholar] [CrossRef]
- Sabo, J.L.; Ruhi, A.; Holtgrieve, G.W.; Elliott, V.; Arias, M.E.; Ngor, P.B.; Räsänen, T.A.; Nam, S. Designing river flows to improve food security futures in the Lower Mekong Basin. Science 2017, 358, eaao1053. [Google Scholar] [CrossRef] [PubMed]
- Boulangeat, I.; Gravel, D.; Thuiller, W. Accounting for dispersal and biotic interactions to disentangle the drivers of species distributions and their abundances. Ecol. Lett. 2012, 15, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Peoples, B.K.; Frimpong, E.A. Biotic interactions and habitat drive positive co-occurrence between facilitating and beneficiary stream fishes. J. Biogeogr. 2016, 43, 923–931. [Google Scholar] [CrossRef]
- Olden, J.D.; Jackson, D.A. Fish—Habitat relationships in lakes: Gaining predictive and explanatory insight by using artificial neural networks. Trans. Am. Fish. Soc. 2001, 130, 878–897. [Google Scholar] [CrossRef]
- Pont, D.; Hugueny, B.; Oberdorff, T. Modelling habitat requirement of European fishes: Do species have similar responses to local and regional environmental constraints? Can. J. Fish. Aquat. Sci. 2005, 62, 163–173. [Google Scholar] [CrossRef]
- Jackson, D.A.; Peres-Neto, P.R.; Olden, J.D. What controls who is where in freshwater fish communities—The roles of biotic, abiotic, and spatial factors. Can. J. Fish. Aquat. Sci. 2001, 58, 157–170. [Google Scholar]
- Mekong River Commission (MRC). State of the Basin Report: 2003; Executive Summary; Mekong River Commission: Phnom Penh, Cambodia, 2003; ISSN 1728:3248. [Google Scholar]
- Arias, M.E.; Cochrane, T.A.; Norton, D.; Killeen, T.J.; Khon, P. The flood pulse as the underlying driver of vegetation in the largest wetland and fishery of the Mekong Basin. AMBIO 2013, 42, 864–876. [Google Scholar] [CrossRef]
- Kummu, M.; Tes, S.; Yin, S.; Adamson, P.; Józsa, J.; Koponen, J.; Richey, J.; Sarkkula, J. Water balance analysis for the Tonle Sap Lake-floodplain system. Hydrol. Process. 2014, 28, 1722–1733. [Google Scholar] [CrossRef]
- Ly, S.; VanZalinge, N.P. Catch Statistics of Cambodian Freshwater Fisheries 1994–1997; Report for Project: The Management of the Freshwater Capture Fisheries of Cambodia Phase 1; Mekong River Commission: Phnom Penh, Cambodia, 1998; p. 146. [Google Scholar]
- Fisheries Administration (FiA). Law on Fisheries; Unofficial Translated by ADB/FAO; Fisheries Administration (FiA): Phnom Penh, Cambodia, 2006. [Google Scholar]
- Degen, P.; Nao, T. Historical, cultural and legal perspectives on the fishing lot system in Cambodia. In Proceedings of the Common Property in the Mekong: Issues of Sustainability and Subsistence; Ahmed, M., Hirsch, P., Eds.; ICLARM Stud. Rev.: Phnom Penh, Cambodia, 2000; pp. 49–60. [Google Scholar]
- Deap, L.; Degen, P.; VanZalinge, N. Fishing Gears of the Cambodian Mekong; Inland Fisheries Research and Development Institude of Cambodia (IFRdDI): Phnom Penh, Cambodia, 2003; ISSN 1726-3972. [Google Scholar]
- Chan, B.; Ngor, P.B.; So, N.; Lek, S. Spatial and temporal changes in fish yields and fish communities in the largest tropical floodplain lake in Asia. Ann. Limnol. Int. J. Lim. 2017, 53, 485–493. [Google Scholar] [CrossRef]
- Chheng, P.; Touch, B.T.; Baran, E.; Leng, S.V. Biological Reviews of Important Cambodian Fish Species, Based on FishBase 2004; Inland Fisheries Research and Development Institude of Cambodia (IFRdDI): Phnom Penh, Cambodia, 2005; Volume 1, p. 127. ISBN 983-2346-48-7. [Google Scholar]
- Poulsen, A.F.; Hortle, K.G.; Chan, S.; Chhuon, C.K.; Viravong, S.; Bouakhamvongsa, K.; Suntornratana, U.; Yoorong, N.; Nguyen, T.T.; Tran, B.Q. Distribution and Ecology of Some Important Riverine Fish Species of the Mekong River Basin; Mekong River Commission: Phnom Penh, Cambodia, 2004; Volume 89, ISSN 1683-1489. [Google Scholar]
- Ngor, P.B.; Chheng, P.; So, N. Declines in catches of some large and medium-sized species in Tonle Sap River. In Proceedings of the Catch and Culture; Mekong River Commission: Phnom Penh, Cambodia, 2015; Volume 21, pp. 4–7. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 10 February 2017).
- Ngor, P.B.; Legendre, P.; Oberdorff, T.; Lek, S. Flow alterations by dams shaped fish assemblage dynamics in the complex Mekong-3S river system. Ecol. Indic. 2018, 88, 103–114. [Google Scholar] [CrossRef]
- Stamatopoulos, C. Sample-Based Fishery Surveys: A Technical Handbook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; ISBN 9251046999. [Google Scholar]
- Rainboth, W.J. FAO Species Identification Field Guide for Fishery Purposes: Fish of the Cambodian Mekong; Niem, V.H., Carpenter, K.E., Angelis, N.D., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 1996; 265p, ISBN 92-5-103743-4. [Google Scholar]
- So, N.; Utsugi, K.; Shibukawa, K.; Thach, P.; Chhuoy, S.; Kim, S.; Chin, D.; Nen, P.; Chheng, P. Fishes of the Cambodian Freshwater Bodies; Inland Fisheries Research and Development Institute, Fisheries Administration: Phnom Penh, Cambodia, 2018; 197p. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, H.H.H.; Wagner, H. Vegan: Community Ecology Package; R Package Version 2.5-6, 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 9 March 2020).
- R Core Team. R: A Language and Environment for Statistical Computing 2017; European Environment Agency: København K, Denmark, 2017. [Google Scholar]
- Lim, P.; Lek, S.; Touch, S.T.; Mao, S.; Chhouk, B. Diversity and spatial distribution of freshwater fish in Great Lake and Tonle Sap river (Cambodia, Southeast Asia). Aquat. Living Resour. 1999, 12, 379–386. [Google Scholar] [CrossRef]
- Kolosa, J. Ecological systems in hierarchical perspective: Breaks in community structure and other consequences. Ecology 1989, 70, 36–47. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Winston, M.R. Local vs. regional influences on local diversity in stream fish communities of Virginia. Ecology 1998, 79, 911–927. [Google Scholar] [CrossRef]
- Dunham, J.B.; Cade, B.S.; Terrell, J.W. Influences of spatial and temporal variation on fish—Habitat relationships defined by regression quantiles. Trans. Am. Fish. Soc. 2002, 131, 86–98. [Google Scholar] [CrossRef]
- Brosse, S.; Lek, S.; Dauba, F. Predicting fish distribution in a mesotrophic lake by hydroacoustic survey and artificial neural networks. Limnol. Oceanogr. 1999, 44, 1293–1303. [Google Scholar] [CrossRef]
- Brosse, S.; Grossman, G.D.; Lek, S. Fish assemblage patterns in the littoral zone of a European reservoir. Freshw. Biol. 2007, 52, 448–458. [Google Scholar] [CrossRef]
- Mekong River Commission (MRC). Monitoring fish abundance and diversity in the Mekong Basin. Catch Cult. 2008, 14, 24–25.
- Lammert, M.; Allan, J. Assessing biotic integrity of streams: Effects of scale in measuring the influence of land use/cover and habitat structure on fish and macroinvertebrates. Environ. Manag. 1999, 23, 257–270. [Google Scholar] [CrossRef]
- Talwar, P.K.; Jhingran, A.G. Inland Fishes of India and Adjacent Countries; A.A. Balkema: Rotterdam, The Netherlands, 1991; Volume 2. [Google Scholar]
- Kottelat, M. Fishes of the Nam Theun and Xe Bangfai basins, Laos, with diagnoses of twenty-two new species (Teleostei: Cyprinidae, Balitoridae, Cobitidae, Coiidae and Odontobutidae). Ichthyol. Explor. Freshw. 1998, 9, 1–128. [Google Scholar]
- Kottelat, M.; Widjanarti, E. The fishes of Danau Sentarum National Park and the Kapuas Lakes area, Kalimantan Barat, Indonesia. Raffles Bull. Zool. 2005, Supplement 13, 139–173. [Google Scholar]
- Singhanouvong, D.; Soulignavong, C.; Vonghachak, K.; Saadsy, B.; Warren, T.J. The Main Wet-Season Migration Through Hoo Som Yai, a Steep-Gradient Channel at the Great Fault Line on the Mekong River, Champassack Province, Southern Lao PDR; Ministry of Agriculture-Forestry: Vientiane, Laos, 1996; 115p. [Google Scholar]
- Menon, A.G.K. Checklist freshwater fishes of India. Rec. Zool. Surv. India 1999, 17, 1–366. [Google Scholar]
- Un, B.; Pech, S.; Baran, E. Aquatic Agricultural Systems in Cambodia: National Situation Analysis; Program Report: AAS-2015-13; CGIAR Research Program on Aquatic Agricultural Systems: Penang, Malaysia, 2015. [Google Scholar]
- Heino, J. Concordance of species richness patterns among multiple freshwater taxa: A regional perspective. Biodivers. Conserv. 2002, 11, 137–147. [Google Scholar] [CrossRef]
- Buisson, L.; Blanc, L.; Grenouillet, G. Modelling stream fish species distribution in a river network: The relative effects of temperature versus physical factors. Ecol. Freshw. Fish 2008, 17, 244–257. [Google Scholar] [CrossRef]
- Chea, R.; Lek, S.; Ngor, P.; Grenouillet, G. Large-Scale patterns of fish diversity and assemblage structure in the longest tropical river in Asia. Ecol. Freshw. Fish 2016, 26, 575–585. [Google Scholar] [CrossRef]
- Poulsen, A.F.; Ouch, P.; Viravong, S.; Suntornratana, U.; Nguyen, T.T. Fish Migrations of the Lower Mekong River Basin: Implications for Development, Planning and Environmental Management; Mekong River Commission: Phnom Penh, Cambodia, 2002; 62p, ISSN 1683-1489. [Google Scholar]
- Baran, E. Fish Migration Triggers in the Lower Mekong Basin and Other Tropical Freshwater Systems; Mekong River Commission: Vientiane, Cambodia, 2006; ISBN 1683-1489. [Google Scholar]
- Hughes, F. The Flooded Forest: Guidance for Policy Makers and River Managers in Europe on the Restoration of Floodplain Forests, FLOBAR2; Department of Geography, University of Cambridge: Cambridge, UK, 2003; 96p. [Google Scholar]
- World Wide Fund for Nature (WWF). Mekong Flooded Forest Ecosystem Health Synthesis Report; World Wide Fund for Nature (WWF): Phnom Penh, Cambodia, 2017; 19p. [Google Scholar]
- Baran, E.; Gallego, G. Cambodia’s fisheries: A decade of changes and evolution. Catch Cult. 2015, 21, 28–31. [Google Scholar]
- National Institute of Statistics (NIS). Cambodian Inter-Census Population Survey 2013; National Institute of Statistics, Ministry of Planning: Phnom Penh, Cambodia, 2013; 142p. [Google Scholar]
- Castello, L.; Hess, L.L.; Thapa, R.; McGrath, D.G.; Arantes, C.C.; Renó, V.F.; Isaac, V.J. Fishery yields vary with land cover on the Amazon River floodplain. Fish Fish. 2018, 19, 431–440. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fishing Lot | The Percentage Cover of Flooded Forest (%) | The Percentage Cover of Open Water (%) | The Percentage Cover Agricultural Field (%) |
|---|---|---|---|
| B01 | 74.064 | 13.687 | 12.249 |
| B02 | 92.391 | 1.426 | 6.183 |
| B03 | 90.654 | 9.213 | 0.133 |
| B04 | 97.378 | 0.832 | 1.789 |
| B05 | 99.658 | 0.000 | 0.342 |
| B06 | 97.873 | 0.000 | 2.127 |
| B07 | 10.200 | 0.000 | 89.800 |
| B08 | 70.178 | 0.000 | 29.822 |
| B09 | 99.982 | 0.000 | 0.018 |
| B10 | 100.000 | 0.000 | 0.000 |
| B11 | 89.754 | 0.000 | 10.246 |
| B12 | 80.466 | 0.000 | 19.534 |
| P01 | 82.462 | 17.538 | 0.000 |
| P02 | 63.546 | 34.188 | 2.266 |
| P03 | 68.851 | 31.149 | 0.000 |
| P04 | 62.047 | 37.953 | 0.000 |
| P05 | 33.271 | 65.598 | 1.131 |
| P06 | 48.506 | 46.976 | 4.518 |
| P07 | 44.987 | 48.856 | 6.157 |
| S01 | 96.130 | 0.000 | 3.870 |
| S02 | 96.314 | 0.000 | 3.686 |
| S03 | 92.187 | 2.330 | 5.483 |
| S04 | 34.507 | 55.821 | 9.672 |
| S05 | 11.215 | 73.477 | 15.308 |
| S06 | 83.347 | 11.307 | 5.347 |
| S07 | 77.499 | 22.501 | 0.000 |
| T01 | 74.825 | 2.359 | 22.816 |
| T02 | 70.327 | 29.673 | 0.000 |
| T03 | 72.863 | 9.697 | 17.440 |
| T04 | 54.281 | 32.006 | 13.714 |
| T05 | 75.308 | 24.692 | 0.000 |
| T06 | 66.186 | 16.696 | 17.118 |
| T07 | 32.346 | 3.633 | 64.020 |
| Variable | Unit | Variable Type | Minimum | Maximum |
|---|---|---|---|---|
| B1 | (°C) | annual mean temperature | 27.3 | 27.7 |
| B2 | (°C) | mean diurnal range (mean of monthly (max temp–min temp)) | 8.8 | 9.6 |
| B3 | % | isothermality (B2/B7) (*100) | 56 | 58 |
| B4 | (°C*100) | temperature seasonality (standard deviation *100) | 1424 | 1731 |
| B5 | (°C) | max temperature of the warmest month | 34.7 | 35.5 |
| B6 | (°C) | min temperature of the coldest month | 18.4 | 19.9 |
| B7 | (°C) | temperature annual range (B5–B6) | 15 | 16.9 |
| B8 | (°C) | mean temperature of the wettest quarter | 27.2 | 27.5 |
| B9 | (°C) | mean temperature of the driest quarter | 25.3 | 25.9 |
| B10 | (°C) | mean temperature of the warmest quarter | 29.2 | 29.4 |
| B11 | (°C) | mean temperature of coldest quarter | 24.7 | 25.7 |
| B12 | mm | annual precipitation | 1169 | 1481 |
| B13 | mm | precipitation of the wettest month | 221 | 279 |
| B14 | mm | precipitation of the driest month | 1 | 6 |
| B15 | mm | precipitation seasonality (coefficient of variation) | 70 | 79 |
| B16 | mm | precipitation of the wettest quarter | 549 | 696 |
| B17 | mm | precipitation of the driest quarter | 22 | 41 |
| B18 | mm | precipitation of the warmest quarter | 209 | 369 |
| B19 | mm | precipitation of the coldest quarter | 73 | 117 |
| Variables | Chm | Pah | Chs | Cye | Gys | Bag | Phs | Trm |
|---|---|---|---|---|---|---|---|---|
| Percentage cover of the flooded forest (FF) α | +0.70 | +4.11 *** | +1.88 * | +2.71 ** | −0.63 | +1.67 | +3.14 ** | −0.85 |
| Percentage cover of the open water (OW) α | +0.48 | +5.98 *** | +0.28 | +4.03 *** | +1.59 | +2.72 ** | +4.86 *** | −1.51 |
| Percentage cover of the agricultural field (AF) α | −0.74 | +1.38 | +1.01 | −0.49 | −2.19 * | −0.19 | −0.64 | −1.1 |
| Temperature and precipitation range (clim1) β | +1.66 | +2.68 ** | −2.57 ** | +2.58 ** | +4.25 *** | +1.20 | +0.97 | −1.34 |
| Temperature range in the warm season (clim2) β | −6.66 *** | −2.23 * | −3.14 ** | −1.68 | +4.62 *** | −0.33 | +1.50 | −2.35 * |
| Precipitation range in the wet season (clim3) β | +0.39 | +0.57 | −3.46 *** | +1.77 | −1.07 | +1.95 * | +1.97 * | −2.70 ** |
| Adj-R2 | 0.68 | 0.833 | 0.62 | 0.83 | 0.709 | 0.74 | 0.76 | 0.50 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, B.; Brosse, S.; Hogan, Z.S.; Ngor, P.B.; Lek, S. Influence of Local Habitat and Climatic Factors on the Distribution of Fish Species in the Tonle Sap Lake. Water 2020, 12, 786. https://doi.org/10.3390/w12030786
Chan B, Brosse S, Hogan ZS, Ngor PB, Lek S. Influence of Local Habitat and Climatic Factors on the Distribution of Fish Species in the Tonle Sap Lake. Water. 2020; 12(3):786. https://doi.org/10.3390/w12030786
Chicago/Turabian StyleChan, Bunyeth, Sébastien Brosse, Zeb S. Hogan, Peng Bun Ngor, and Sovan Lek. 2020. "Influence of Local Habitat and Climatic Factors on the Distribution of Fish Species in the Tonle Sap Lake" Water 12, no. 3: 786. https://doi.org/10.3390/w12030786
APA StyleChan, B., Brosse, S., Hogan, Z. S., Ngor, P. B., & Lek, S. (2020). Influence of Local Habitat and Climatic Factors on the Distribution of Fish Species in the Tonle Sap Lake. Water, 12(3), 786. https://doi.org/10.3390/w12030786