Microbial Assisted Hexavalent Chromium Removal in Bioelectrochemical Systems

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
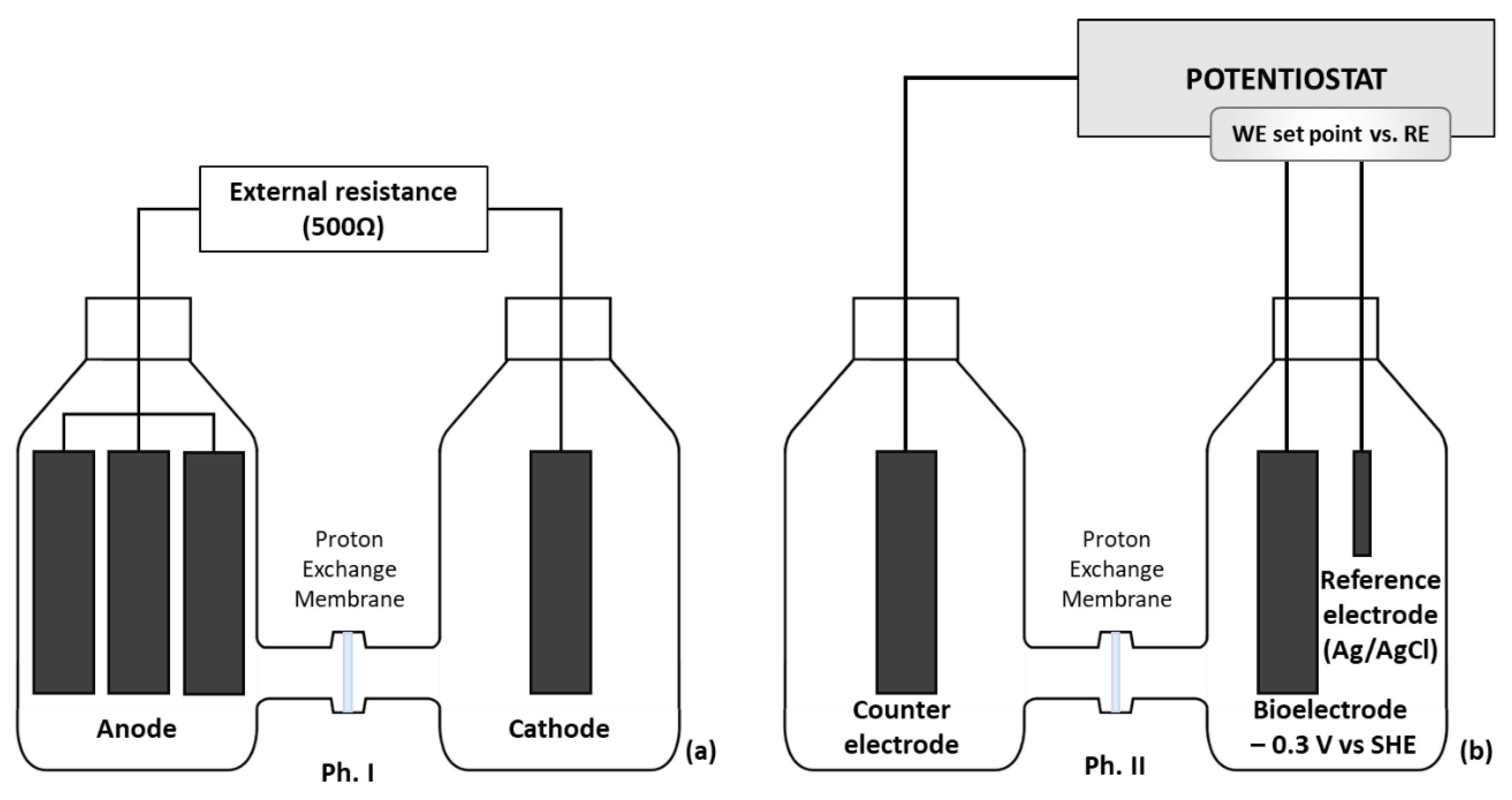
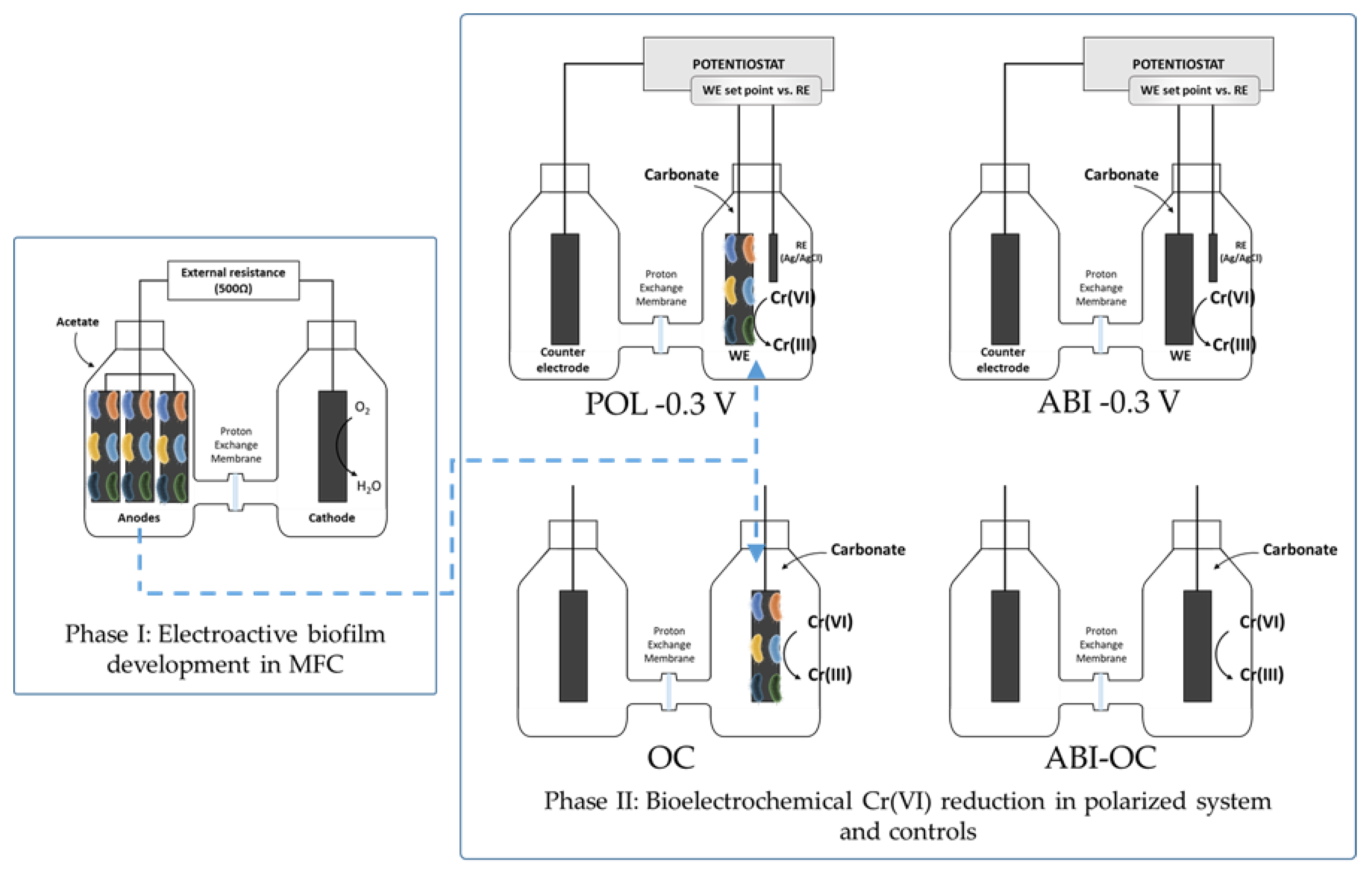
2.1. Reactors Set-Up and Operation
2.2. Electroactive Biofilm Development in MFC
2.3. Polarized Bio-Electrode for Cr(VI) Removal
2.4. Analyses and Data Processing
2.5. Calculations
3. Results
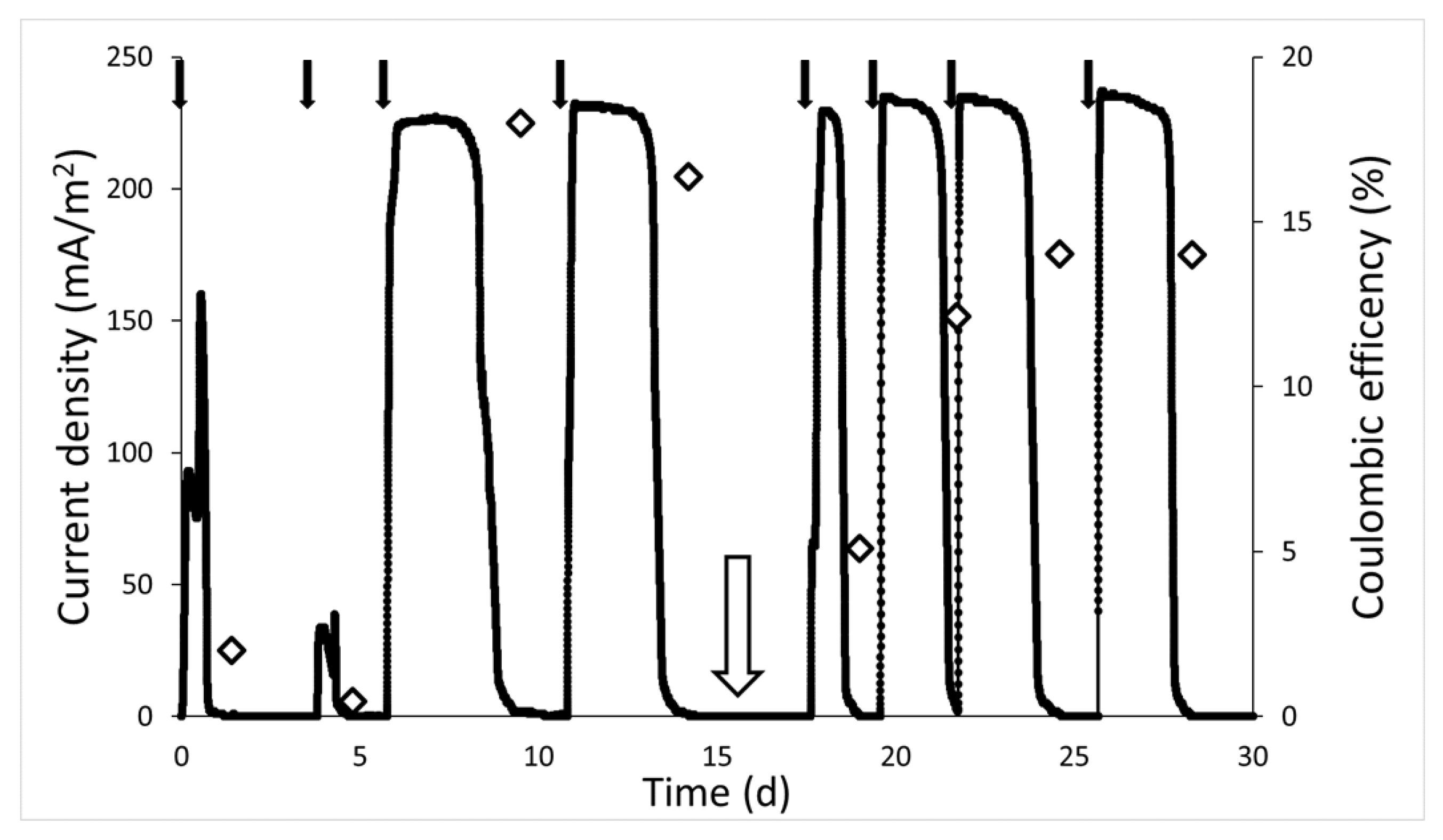
3.1. Current Density in the MFC
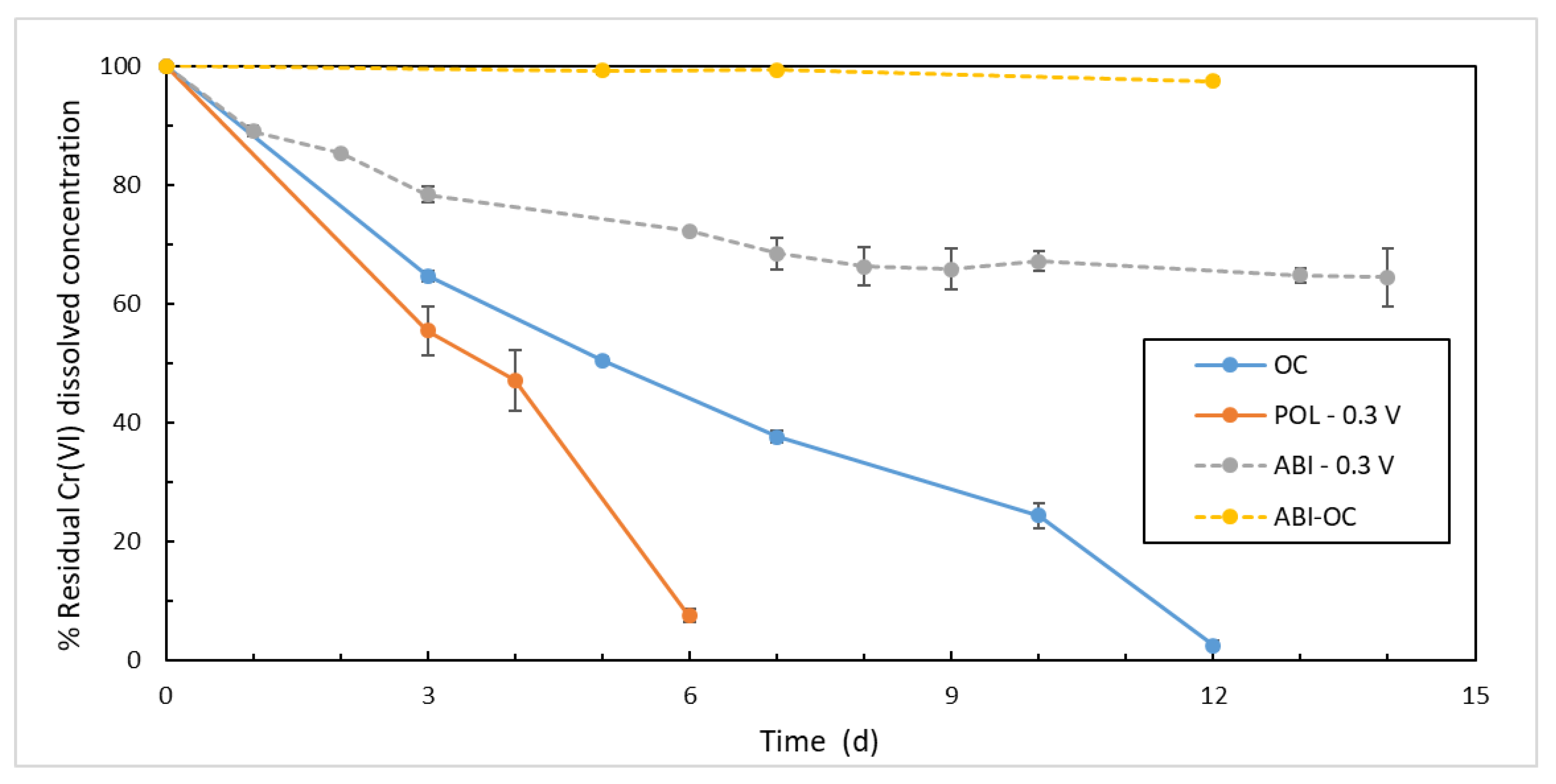
3.2. Cr(VI) trends
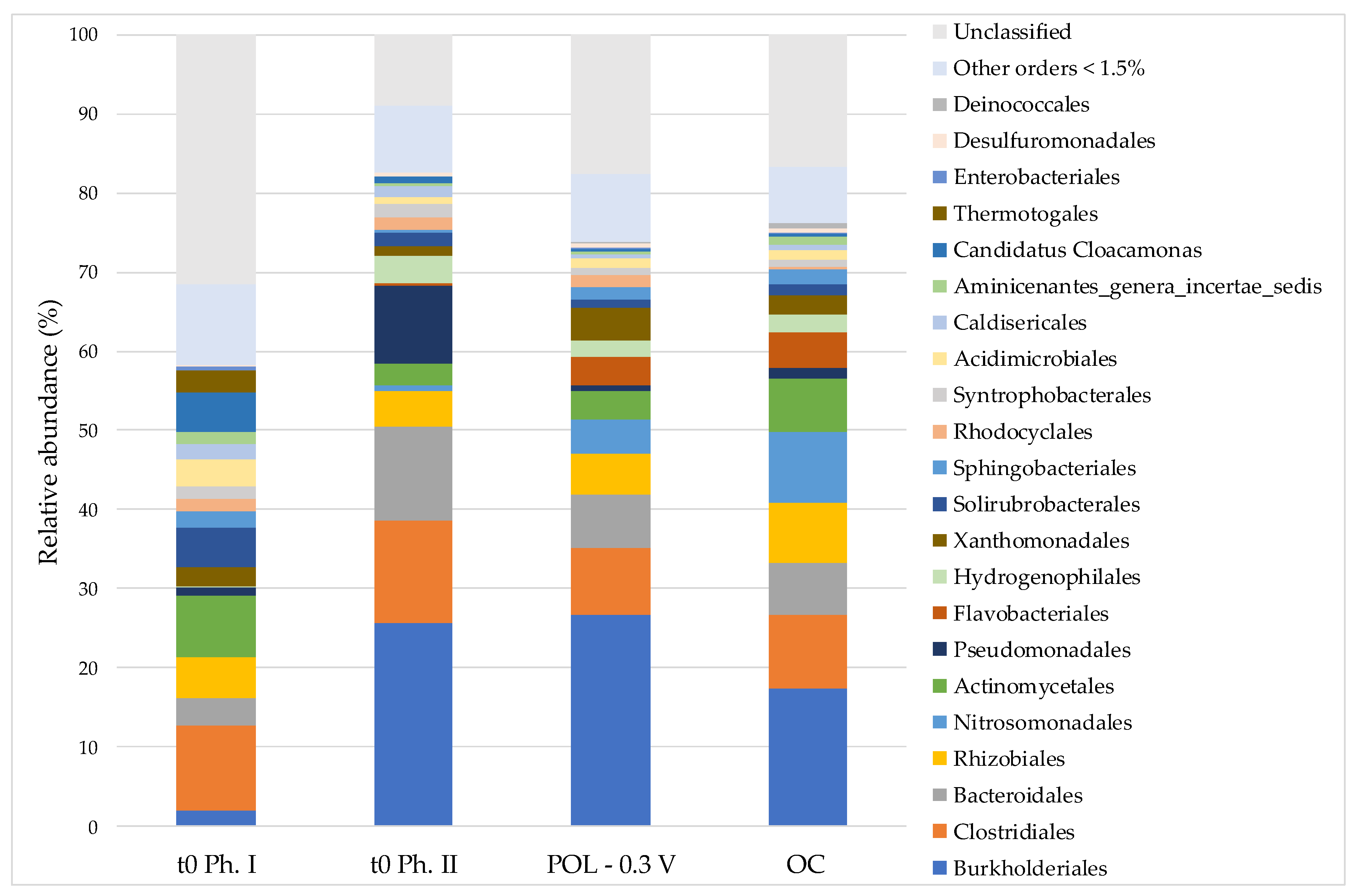
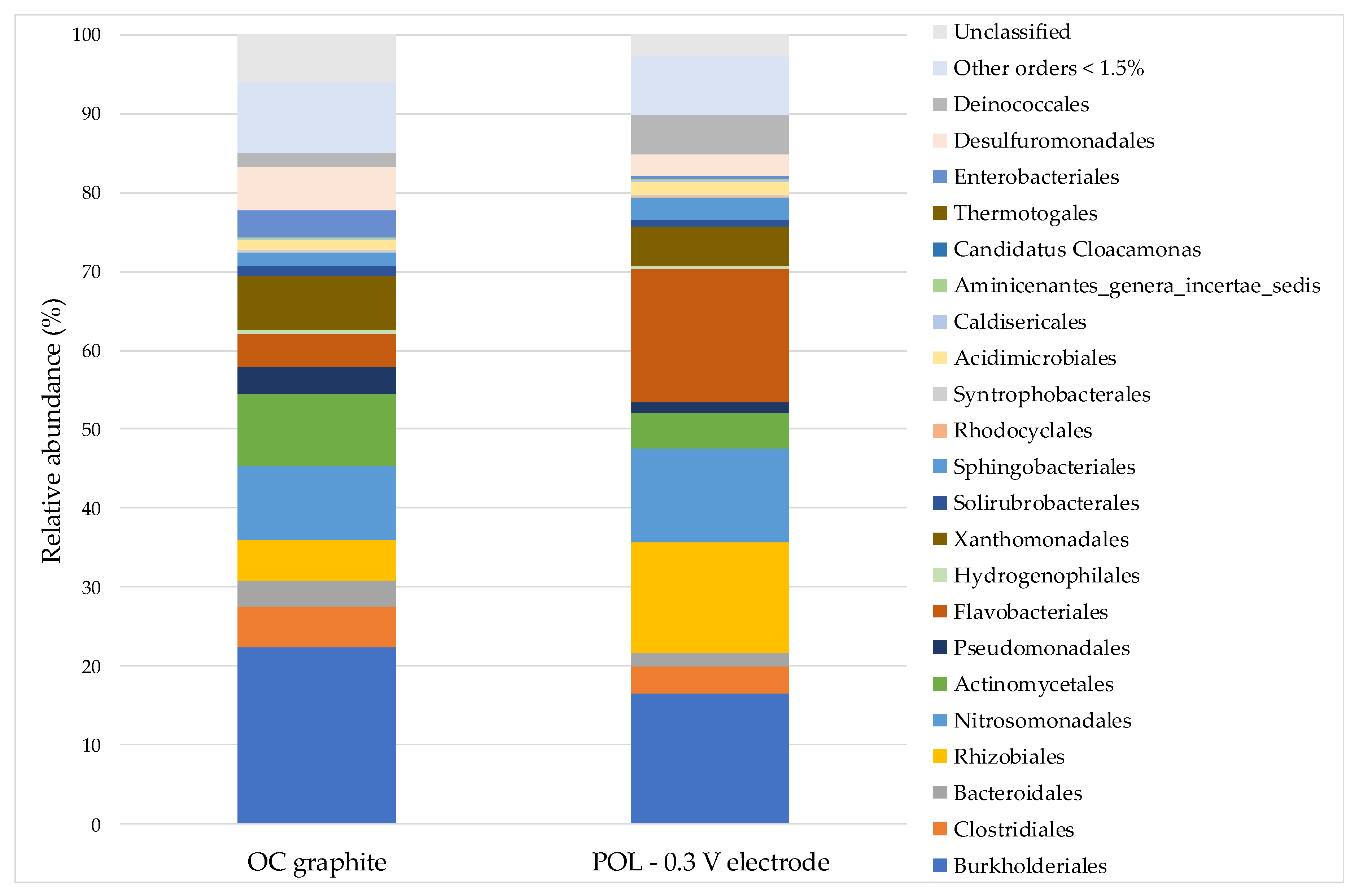
3.3. Microbial Communities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ayyappan, C.S. Microbial leaching of chromium from solidified waste forms—A kinetic study. J. Ecol. Eng. 2015, 16, 36–42. [Google Scholar] [CrossRef]
- Tschersich, C.; Barouki, R.; Uhl, M.; Klánová, J.; Horvat, M.; Alimonti, A.; Sarigiannis, D.; Santonen, T.; Lebret, E.; Schoeters, G. Scoping Documents for 2018 Deliverable Report D 4.2—Input for Cd and Cr(VI); Tenical Report for HBM4EU H2020 Programme; 2018; Available online: https://www.hbm4eu.eu/deliverables/ (accessed on 7 February 2020).
- Mohan, D.; Rajput, S.; Singh, V.K.; Steele, P.H.; Pittman, C.U. Modeling and evaluation of chromium remediation from water using low cost bio-char, a green adsorbent. J. Hazard. Mater. 2011, 188, 319–333. [Google Scholar] [CrossRef]
- Song, Z.; Williams, C.J.; Edyvean, R.G.J. Sedimentation of tannery wastewater. Water Res. 2000, 34, 2171–2176. [Google Scholar] [CrossRef]
- Madhavi, V.; Vijay, A.; Reddy, B.; Reddy, K.G.; Madhavi, G.; Nagavenkata, T.; Prasad, K.V. An Overview on Research Trends in Remediation of Chromium. Res. J. Recent Sci. Res. 2013, 2, 71–83. [Google Scholar]
- Peng, H.; Guo, J.; Li, B.; Liu, Z.; Tao, C. High-efficient recovery of chromium (VI) with lead sulfate. J. Taiwan Inst. Chem. Eng. 2018, 85, 149–154. [Google Scholar] [CrossRef]
- Peng, H.; Leng, Y.; Cheng, Q.; Shang, Q.; Shu, J.; Guo, J. Efficient Removal of Hexavalent Chromium from Wastewater with Electro-Reduction. Processes 2019, 7, 41. [Google Scholar] [CrossRef]
- Jobby, R.; Jha, P.; Yadav, A.K.; Desai, N. Biosorption and biotransformation of hexavalent chromium [Cr(VI)]: A comprehensive review. Chemosphere 2018, 207, 255–266. [Google Scholar] [CrossRef]
- Daghio, M.; Aulenta, F.; Vaiopoulou, E.; Franzetti, A.; Arends, J.B.A.; Sherry, A.; Suárez-Suárez, A.; Head, I.M.; Bestetti, G.; Rabaey, K. Electrobioremediation of oil spills. Water Res. 2017, 114, 351–370. [Google Scholar] [CrossRef]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef]
- Rabaey, K.; Angenent, L.; Schroder, U.; Keller, J. Bioelectrochemical Systems: From Extracellular Electron Transfer to Biotechnological Application; IWA Publishing: London, UK, 2009; ISBN 9781843392330. [Google Scholar]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.Q.; Fredrickson, J.K. Extracellular Electron Transfer Mechanisms between Microorganisms and Minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- An, Z.; Zhang, H.; Wen, Q.; Chen, Z.; Du, M. Desalination combined with hexavalent chromium reduction in a microbial desalination cell. Desalination 2014, 354, 181–188. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.; Lei, L. Electricity production during the treatment of real electroplating wastewater containing Cr6+ using microbial fuel cell. Process Biochem. 2008, 43, 1352–1358. [Google Scholar] [CrossRef]
- Wang, G.; Huang, L.; Zhang, Y. Cathodic reduction of hexavalent chromium [Cr(VI)] coupled with electricity generation in microbial fuel cells. Biotechnol. Lett. 2008, 30, 1959–1966. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Feng, C.; Ni, J.; Zhang, J.; Huang, W. Simultaneous reduction of vanadium (V) and chromium (VI) with enhanced energy recovery based on microbial fuel cell technology. J. Power Sources 2012, 204, 34–39. [Google Scholar] [CrossRef]
- Huang, L.; Chen, J.; Quan, X.; Yang, F. Enhancement of hexavalent chromium reduction and electricity production from a biocathode microbial fuel cell. Bioprocess Biosyst. Eng. 2010, 33, 937–945. [Google Scholar] [CrossRef]
- Tandukar, M.; Huber, S.J.; Onodera, T.; Pavlostathis, S.G. Biological chromium(VI) reduction in the cathode of a microbial fuel cell. Environ. Sci. Technol. 2009, 43, 8159–8165. [Google Scholar] [CrossRef]
- Huang, L.; Chai, X.; Quan, X.; Logan, B.E.; Chen, G. Reductive dechlorination and mineralization of pentachlorophenol in biocathode microbial fuel cells. Bioresour. Technol. 2012, 111, 167–174. [Google Scholar] [CrossRef]
- Xafenias, N.; Zhang, Y.; Banks, C.J. Enhanced performance of hexavalent chromium reducing cathodes in the presence of Shewanella oneidensis MR-1 and lactate. Environ. Sci. Technol. 2013, 47, 4512–4520. [Google Scholar] [CrossRef]
- Thatoi, H.; Das, S.; Mishra, J.; Rath, B.P.; Das, N. Bacterial chromate reductase, a potential enzyme for bioremediation of hexavalent chromium: A review. J. Environ. Manag. 2014, 146, 383–399. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E.J.P. Reduction of Chromate by Desulfovibrio-Vulgaris and Its c3 Cytochrome. Appl. Environ. Microbiol. 1994, 60, 726–728. [Google Scholar] [CrossRef]
- Chung, J.; Nerenberg, R.; Rittmann, B.E. Bio-reduction of soluble chromate using a hydrogen-based membrane biofilm reactor. Water Res. 2006, 40, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Inglett, K.S.; Bae, H.S.; Aldrich, H.C.; Hatfield, K.; Ogram, A.V. Clostridium chromiireducens sp. nov., isolated from Cr(VI)-contaminated soil. Int. J. Syst. Evol. Microbiol. 2011, 61, 2626–2631. [Google Scholar] [CrossRef] [PubMed]
- Mclean, J.; Beveridge, T.J. Chromate Reduction by a Pseudomonad Isolated from a Site Contaminated with Chromated Copper Arsenate. Appl. Environ. Microbiol. 2001, 67, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- McLean, J.S.; Beveridge, T.J.; Phipps, D. Isolation and characterization of a chromium-reducing bacterium from a chromated copper arsenate-contaminated site. Environ. Microbiol. 2000, 2, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Malaviya, P.; Singh, A. Bioremediation of chromium solutions and chromium containing wastewaters. Crit. Rev. Microbiol. 2016, 42, 607–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, S.; Wang, J.; Song, Q.; Zhang, W.; He, Q.; Song, J.; Ma, F. Comparison of performance and microbial communities in a bioelectrochemical system for simultaneous denitrification and chromium removal: Effects of pH. Process Biochem. 2018, 73, 154–161. [Google Scholar] [CrossRef]
- Xue, H.; Zhou, P.; Huang, L.; Quan, X.; Yuan, J. Cathodic Cr(VI) reduction by electrochemically active bacteria sensed by fluorescent probe. Sens. Actuators B Chem. 2017, 243, 303–310. [Google Scholar] [CrossRef]
- Pous, N.; Balaguer, M.D.; Colprim, J.; Puig, S. Opportunities for groundwater microbial electro-remediation. Microb. Biotechnol. 2018, 11, 119–135. [Google Scholar] [CrossRef]
- Williams, K.H.; Nevin, K.P.; Franks, A.; Englert, A.; Long, P.E.; Lovley, D.R. Electrode-based approach for monitoring in situ microbial activity during subsurface bioremediation. Environ. Sci. Technol. 2010, 44, 47–54. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Q.; Jiang, L.; Zhou, P.; Quan, X.; Logan, B.E. Adaptively Evolving Bacterial Communities for Complete and Selective Reduction of Cr(VI), Cu(II), and Cd(II) in Biocathode Bioelectrochemical Systems. Environ. Sci. Technol. 2015, 49, 9914–9924. [Google Scholar] [CrossRef]
- Song, T.S.; Jin, Y.; Bao, J.; Kang, D.; Xie, J. Graphene/biofilm composites for enhancement of hexavalent chromium reduction and electricity production in a biocathode microbial fuel cell. J. Hazard. Mater. 2016, 317, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhu, X.; Song, T.; Zhang, L.; Jia, H.; Wei, P. Effect of acclimatization on hexavalent chromium reduction in a biocathode microbial fuel cell. Bioresour. Technol. 2015, 180, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Kellner, K.; Posnicek, T.; Ettenauer, J.; Zuser, K.; Brandl, M. A new, low-cost potentiostat for environmental measurements with an easy-to-use PC interface. Procedia Eng. 2015, 120, 956–960. [Google Scholar] [CrossRef]
- Rowe, A.A.; Bonham, A.J.; White, R.J.; Zimmer, M.P.; Yadgar, R.J.; Hobza, T.M.; Honea, J.W.; Ben-Yaacov, I.; Plaxco, K.W. Cheapstat: An open-source, “do-it-yourself” potentiostat for analytical and educational applications. PLoS ONE 2011, 6, e23783. [Google Scholar] [CrossRef]
- Eaton, A.D.; Franson, M.A.H. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association, Ed.; American Public Health Association: Washington, DC, USA, 2005; ISBN 9780875530475. [Google Scholar]
- Huber, J.A.; Welch, D.B.M.; Morrison, H.G.; Huse, S.M.; Neal, P.R.; Butterfield, D.A.; Sogin, M.L. Microbial population structures in the deep marine biosphere. Science 2007, 318, 97–101. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, P.Y. Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef]
- Palma, E.; Daghio, M.; Espinoza Tofalos, A.; Franzetti, A.; Cruz Viggi, C.; Fazi, S.; Petrangeli Papini, M.; Aulenta, F. Anaerobic electrogenic oxidation of toluene in a continuous-flow bioelectrochemical reactor: Process performance, microbial community analysis, and biodegradation pathways. Environ. Sci. Water Res. Technol. 2018, 4, 2136–2145. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Rismani-Yazdi, H.; Carver, S.M.; Christy, A.D.; Tuovinen, O.H. Cathodic limitations in microbial fuel cells: An overview. J. Power Sources 2008, 180, 683–694. [Google Scholar] [CrossRef]
- Lu, L.; Ren, N.; Zhao, X.; Wang, H.; Wu, D.; Xing, D. Hydrogen production, methanogen inhibition and microbial community structures in psychrophilic single-chamber microbial electrolysis cells. Energy Environ. Sci. 2011, 1329–1336. [Google Scholar] [CrossRef]
- Wang, Y.-T. Microbial reduction of chromate. In Environmental Microbe-Metal Interactions; American Society of Microbiology: Washington, DC, USA, 2000; pp. 225–235. [Google Scholar]
- Viti, C.; Marchi, E.; Decorosi, F.; Giovannetti, L. Molecular mechanisms of Cr(VI) resistance in bacteria and fungi. FEMS Microbiol. Rev. 2014, 38, 633–659. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Regan, J.M.; Quan, X. Electron transfer mechanisms, new applications, and performance of biocathode microbial fuel cells. Bioresour. Technol. 2011, 102, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Focardi, S.; Pepi, M.; Focardi, S.E. Microbial Reduction of Hexavalent Chromium as a Mechanism of Detoxification and Possible Bioremediation Applications. Biodegrad. Life Sci. 2013, 12, 321–348. [Google Scholar]
- Vendruscolo, F.; da Rocha Ferreira, G.L.; Antoniosi Filho, N.R. Biosorption of hexavalent chromium by microorganisms. Int. Biodeterior. Biodegrad. 2017, 119, 87–95. [Google Scholar] [CrossRef]
- Qu, Y.; Zhang, X.; Xu, J.; Zhang, W.; Guo, Y. Removal of hexavalent chromium from wastewater using magnetotactic bacteria. Sep. Purif. Technol. 2014, 136, 10–17. [Google Scholar] [CrossRef]
- Gangadharan, P.; Nambi, I.M. Hexavalent chromium reduction and energy recovery by using dual-chambered microbial fuel cell. Water Sci. Technol. 2015, 71, 353–358. [Google Scholar] [CrossRef]
- Li, M.; Zhou, S.; Xu, Y.; Liu, Z.; Ma, F.; Zhi, L.; Zhou, X. Simultaneous Cr(VI) reduction and bioelectricity generation in a dual chamber microbial fuel cell. Chem. Eng. J. 2018, 334, 1621–1629. [Google Scholar] [CrossRef]
- Singhvi, P.; Chhabra, M.; Singhvi, P.; Chhabra, M. Simultaneous Chromium Removal and Power Generation Using Algal Biomass in a Dual Chambered Salt Bridge Microbial Fuel Cell. J. Bioremediat. Biodegrad. 2013, 4, 190. [Google Scholar]
- Chao, A. Estimating the Population Size for Capture-Recapture Data with Unequal Catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef]
- Sotres, A.; Díaz-Marcos, J.; Guivernau, M.; Illa, J.; Magrí, A.; Prenafeta-Boldú, F.X.; Bonmatí, A.; Viñas, M. Microbial community dynamics in two-chambered microbial fuel cells: Effect of different ion exchange membranes. J. Chem. Technol. Biotechnol. 2015, 90, 1497–1506. [Google Scholar] [CrossRef]
- Barbosa, S.G.; Peixoto, L.; Soares, O.S.G.P.; Pereira, M.F.R.; Heijne, A.T.; Kuntke, P.; Alves, M.M.; Pereira, M.A. Influence of carbon anode properties on performance and microbiome of Microbial Electrolysis Cells operated on urine. Electrochim. Acta 2018, 267, 122–132. [Google Scholar] [CrossRef]
- Read, S.T.; Dutta, P.; Bond, P.L.; Keller, J.; Rabaey, K. Initial development and structure of biofilms on microbial fuel cell anodes. BMC Microbiol. 2010, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Joicy, A.; Song, Y.-C.; Lee, C.-Y. Electroactive microorganisms enriched from activated sludge remove nitrogen in bioelectrochemical reactor. J. Environ. Manag. 2019, 233, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Saratale, G.D.; Saratale, R.G.; Shahid, M.K.; Zhen, G.; Kumar, G.; Shin, H.S.; Choi, Y.G.; Kim, S.H. A comprehensive overview on electro-active biofilms, role of exo-electrogens and their microbial niches in microbial fuel cells (MFCs). Chemosphere 2017, 178, 534–547. [Google Scholar] [CrossRef] [PubMed]
- Franzetti, A.; Daghio, M.; Parenti, P.; Truppi, T.; Bestetti, G.; Trasatti, S.P.; Cristiani, P. Monod Kinetics Degradation of Low concentration Residual Organics in Membraneless Microbial Fuel Cells. J. Electrochem. Soc. 2017, 164, H3091–H3096. [Google Scholar] [CrossRef]
- Yun, H.; Liang, B.; Kong, D.; Wang, A. Improving biocathode community multifunctionality by polarity inversion for simultaneous bioelectroreduction processes in domestic wastewater. Chemosphere 2018, 194, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Opitz, S.; Harzer, R.; Totsche, K.; Küsel, K. Attached and Suspended Denitrifier Communities in Pristine Limestone Aquifers Harbor High Fractions of Potential Autotrophs Oxidizing Reduced Iron and Sulfur Compounds. Microb. Ecol. 2017, 74, 264–277. [Google Scholar] [CrossRef]
- Fernandez, N.; Sierra-Alvarez, R.; Amils, R.; Field, J.A.; Sanz, J.L. Compared microbiology of granular sludge under autotrophic, mixotrophic and heterotrophic denitrification conditions. Water Sci. Technol. 2009, 59, 1227–1236. [Google Scholar] [CrossRef]
- Im, C.H.; Kim, C.; Song, Y.E.; Oh, S.E.; Jeon, B.H.; Kim, J.R. Electrochemically enhanced microbial CO conversion to volatile fatty acids using neutral red as an electron mediator. Chemosphere 2018, 191, 166–173. [Google Scholar] [CrossRef]
- Paiva, M.C.; Ávila, M.P.; Reis, M.P.; Costa, P.S.; Nardi, R.M.D.; Nascimento, A.M.A. The microbiota and abundance of the class 1 integron-integrase gene in tropical sewage treatment plant influent and activated sludge. PLoS ONE 2015, 10, e0131532. [Google Scholar] [CrossRef]
- Khanongnuch, R.; Di Capua, F.; Lakaniemi, A.-M.; Rene, E.R.; Lens, P.N.L. H2S removal and microbial community composition in an anoxic biotrickling filter under autotrophic and mixotrophic conditions. J. Hazard. Mater. 2018, 367, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Morel, M.A.; Iriarte, A.; Jara, E.; Musto, H.; Castro-Sowinski, S. Revealing the biotechnological potential of Delftia sp. JD2 by a genomic approach. AIMS Bioeng. 2016, 3, 156–175. [Google Scholar] [CrossRef]
- Chen, J.; Yang, Y.; Liu, Y.; Tang, M.; Wang, R.; Tian, Y.; Jia, C. Bacterial community shift and antibiotics resistant genes analysis in response to biodegradation of oxytetracycline in dual graphene modified bioelectrode microbial fuel cell. Bioresour. Technol. 2019, 276, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Wu, Y.; Qian, F.; Chen, C.; Shen, Y.; Zhao, Y. Methane-Oxidizing Microorganism Properties in Landfills. Polish J. Environ. Stud. 2019, 28, 3809–3818. [Google Scholar] [CrossRef]
- Schauss, T.; Busse, H.J.; Golke, J.; Kämpfer, P.; Glaeser, S.P. Moheibacter stercoris sp. Nov., isolated from an input sample of a biogas plant. Int. J. Syst. Evol. Microbiol. 2016, 66, 2585–2591. [Google Scholar] [CrossRef]
- Liao, C.; Wu, J.; Zhou, L.; Li, T.; Du, Q.; An, J.; Li, N.; Wang, X. Optimal set of electrode potential enhances the toxicity response of biocathode to formaldehyde. Sci. Total Environ. 2018, 644, 1485–1492. [Google Scholar] [CrossRef]
- Sun, J.; Xu, W.; Yang, P.; Li, N.; Yuan, Y.; Zhang, H.; Ning, X.; Zhang, Y.; Chang, K.; Peng, Y.; et al. Enhancing the performance of photo-bioelectrochemical fuel cell using graphene oxide/cobalt/polypyrrole composite modified photo-biocathode in the presence of antibiotic. Int. J. Hydrogen Energy 2018, 44, 1919–1929. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Q.; Quan, X.; Liu, Y.; Chen, G. Bioanodes/biocathodes formed at optimal potentials enhance subsequent pentachlorophenol degradation and power generation from microbial fuel cells. Bioelectrochemistry 2013, 94, 13–22. [Google Scholar] [CrossRef]
- Xiao, Y.; Zheng, Y.; Wu, S.; Yang, Z.H.; Zhao, F. Bacterial Community Structure of Autotrophic Denitrification Biocathode by 454 Pyrosequencing of the 16S rRNA Gene. Microb. Ecol. 2015, 69, 492–499. [Google Scholar] [CrossRef]
- Liao, C.; Wu, J.; Zhou, L.; Li, T.; An, J.; Huang, Z.; Li, N.; Wang, X. Repeated transfer enriches highly active electrotrophic microbial consortia on biocathodes in microbial fuel cells. Biosens. Bioelectron. 2018, 121, 118–124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Work Phase | Description | Acronym | Carbon Source | Hexavalent Chromium | Microbial Characterization |
|---|---|---|---|---|---|
| Electroactive bacteria enrichment | Inoculum in MFC | t0 Ph. I | Organic matter (anaerobic sludge) | - | Planktonic community |
| MFC until 15th day—inoculum of POL −0.3 V and OC | t0 Ph. II | Acetate | - | ||
| Cr(VI) removal tests | Potentiostatically controlled system | POL −0.3 V | KHCO3 | K2Cr2O7 (1 mg L−1 Cr(VI)) | Planktonic community and biofilm developed on polarized electrode (POL −0.3 V electrode) |
| Open circuit control | OC | Planktonic community and biofilm developed on graphite (OC graphite) | |||
| Potentiostatically controlled abiotic system | ABI −0.3 V | - | |||
| Open circuit abiotic control | ABI-OC | - |
| Order | OC Graphite (%) | POL −0.3 V Electrode (%) |
|---|---|---|
| Genus | ||
| Burkholderiales | 22.4 | 16.5 |
| Advenella | 14.7 | 8.2 |
| Cupriavidus | 2.8 | 1.6 |
| Polaromonas | 1.6 | <1.5 |
| Flavobacteriales | 4.2 | 17.0 |
| Moheibacter | 4.1 | 16.7 |
| Nitrosomonadales | 9.3 | 11.9 |
| Nitrosomonas | 9.1 | 11.9 |
| Rhizobiales | 5.2 | 14.0 |
| Nitrobacter | <1.5 | 9.9 |
| Actinomycetales | 9.2 | 4.4 |
| Rhodococcus | 5.0 | 1.5 |
| Xanthomonadales | 6.9 | 5.0 |
| Stenotrophomonas | 3.0 | 3.1 |
| Dyella | 3.3 | <1.5 |
| Clostridiales | 5.1 | 3.5 |
| Clostridium XI | 1.8 | 1.8 |
| Desulfuromonadales | 5.4 | 2.9 |
| Geobacter | 4.2 | 1.8 |
| Deinococcales | 1.8 | 4.9 |
| Truepera | 1.8 | 4.9 |
| Bacteroidales | 3.2 | 1.7 |
| Petrimonas | 1.7 | <1.5 |
| Pseudomonadales | 3.3 | 1.4 |
| Pseudomonas | 3.1 | <1.5 |
| Sphingobacteriales | 1.6 | 2.9 |
| Enterobacteriales | 3.5 | <1.5 |
| Escherichia/Shigella | 3.5 | <1.5 |
| Acidimicrobiales | <1.5 | 1.8 |
| Other orders <1.5% | 8.9 | 7.5 |
| Others genera <1.5% | 24.4 | 21.9 |
| Unclassified order | 6.1 | 2.7 |
| Unclassified genus | 16.1 | 16.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beretta, G.; Daghio, M.; Espinoza Tofalos, A.; Franzetti, A.; Mastorgio, A.F.; Saponaro, S.; Sezenna, E. Microbial Assisted Hexavalent Chromium Removal in Bioelectrochemical Systems. Water 2020, 12, 466. https://doi.org/10.3390/w12020466
Beretta G, Daghio M, Espinoza Tofalos A, Franzetti A, Mastorgio AF, Saponaro S, Sezenna E. Microbial Assisted Hexavalent Chromium Removal in Bioelectrochemical Systems. Water. 2020; 12(2):466. https://doi.org/10.3390/w12020466
Chicago/Turabian StyleBeretta, Gabriele, Matteo Daghio, Anna Espinoza Tofalos, Andrea Franzetti, Andrea Filippo Mastorgio, Sabrina Saponaro, and Elena Sezenna. 2020. "Microbial Assisted Hexavalent Chromium Removal in Bioelectrochemical Systems" Water 12, no. 2: 466. https://doi.org/10.3390/w12020466
APA StyleBeretta, G., Daghio, M., Espinoza Tofalos, A., Franzetti, A., Mastorgio, A. F., Saponaro, S., & Sezenna, E. (2020). Microbial Assisted Hexavalent Chromium Removal in Bioelectrochemical Systems. Water, 12(2), 466. https://doi.org/10.3390/w12020466