Spatiotemporal Relationships of Phytoplankton Blooms, Drought, and Rainstorms in Freshwater Reservoirs



Abstract
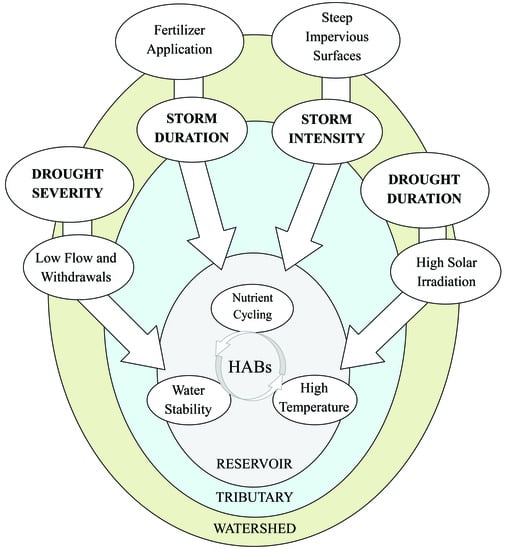
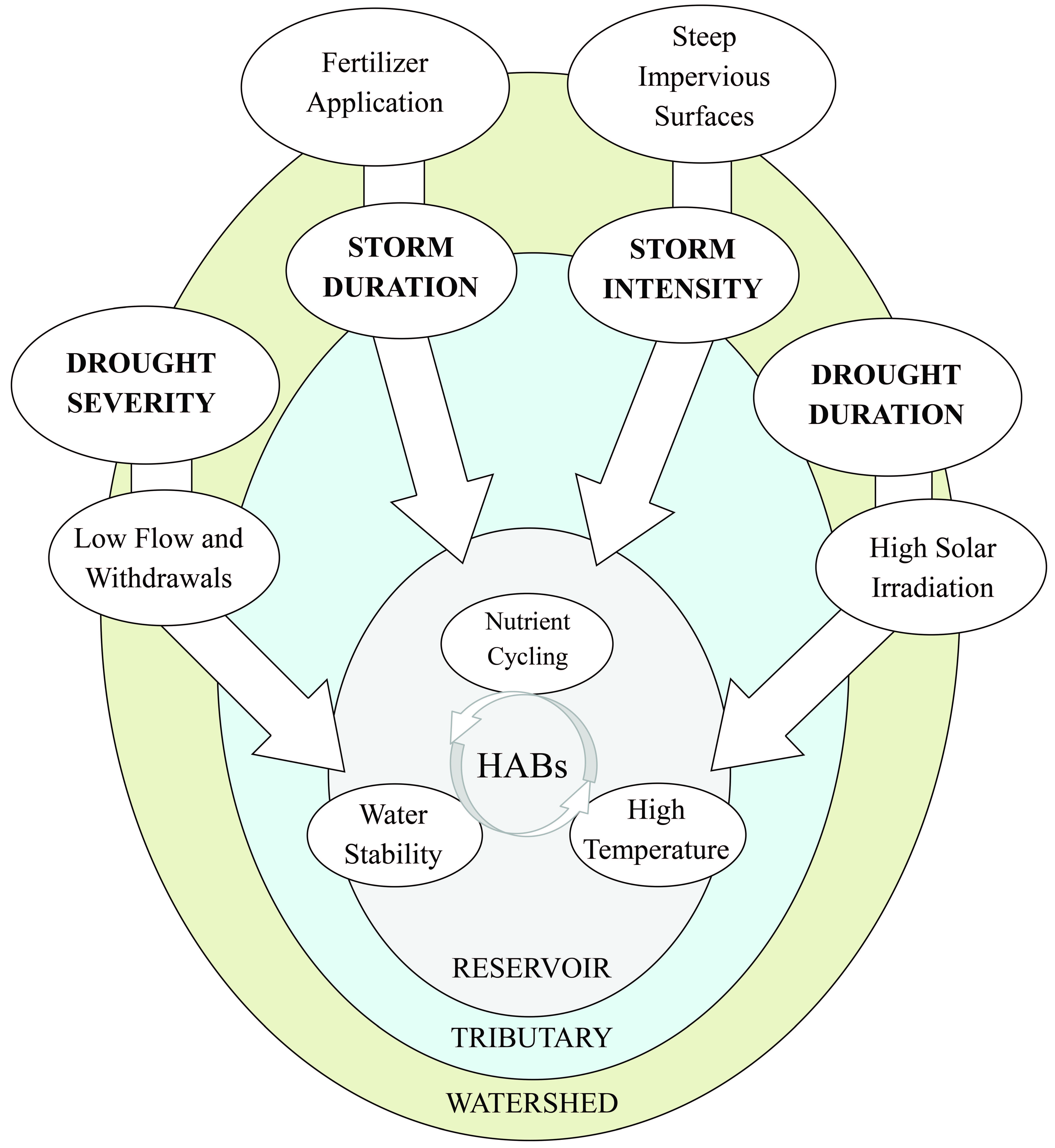
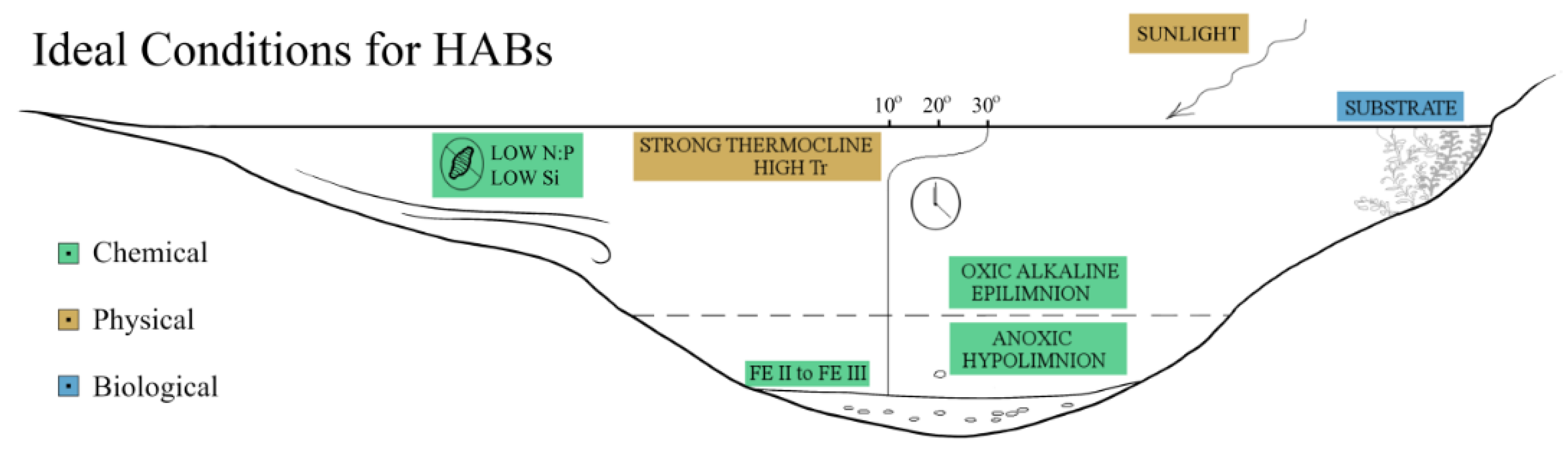
1. Introduction
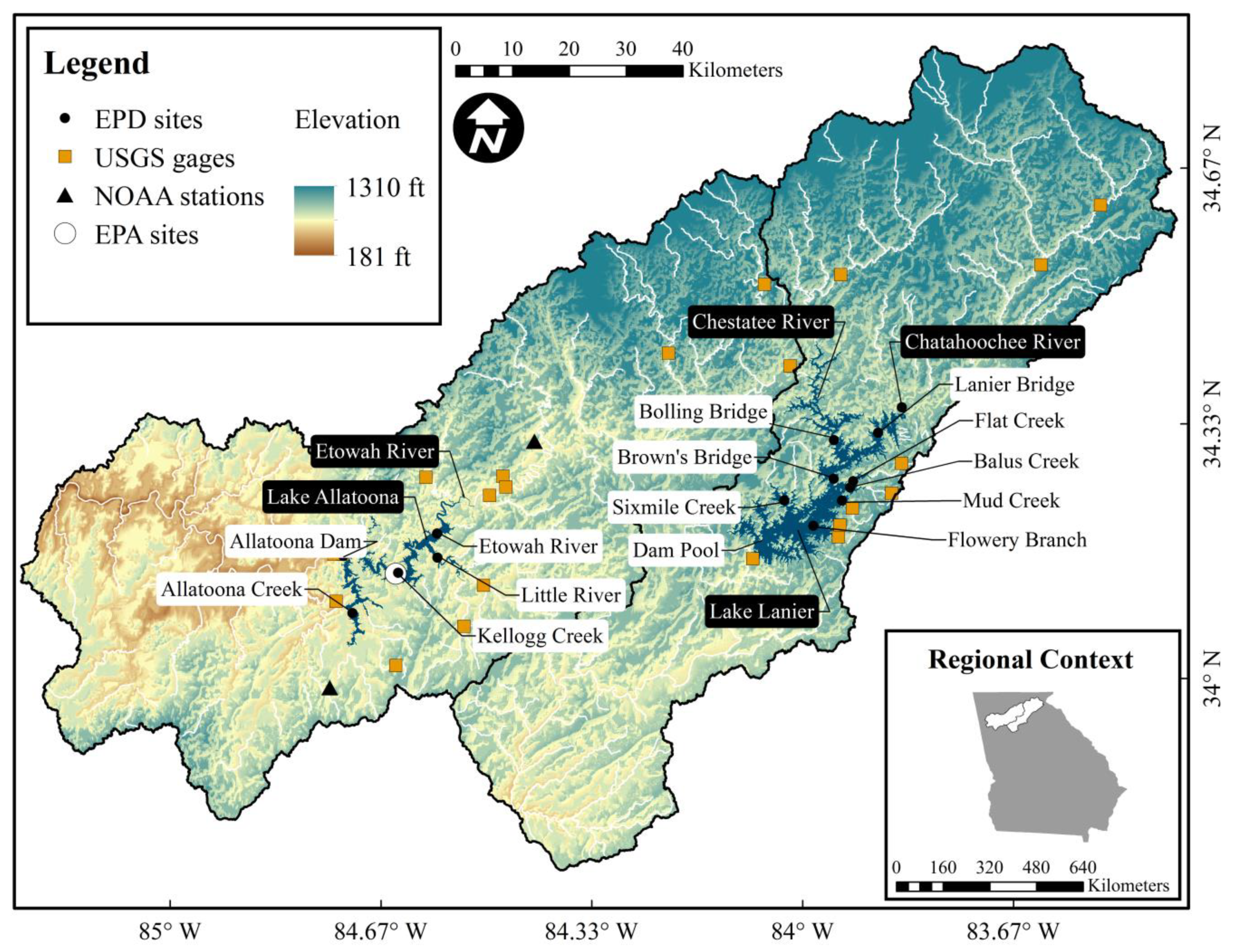
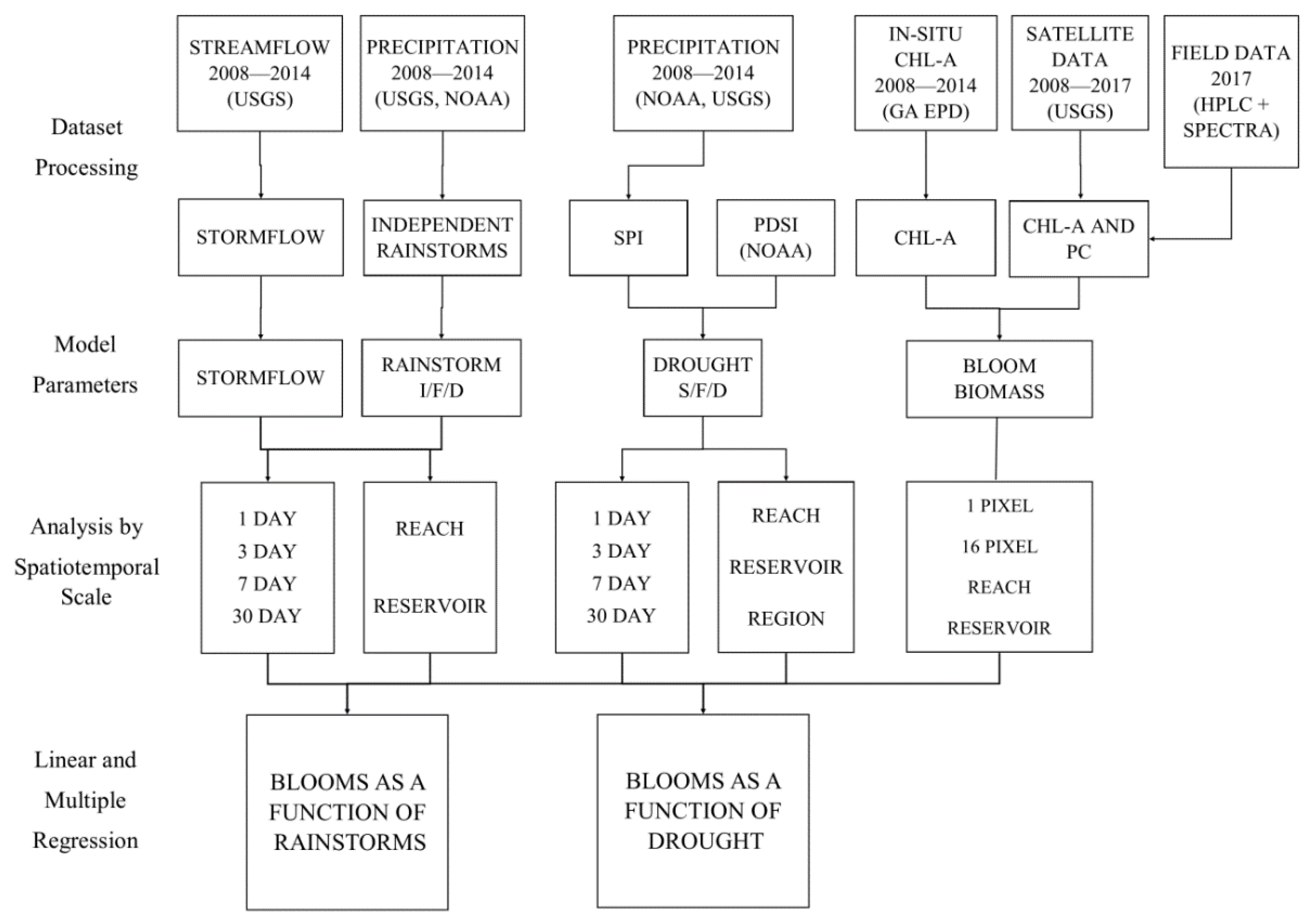
2. Materials and Methods
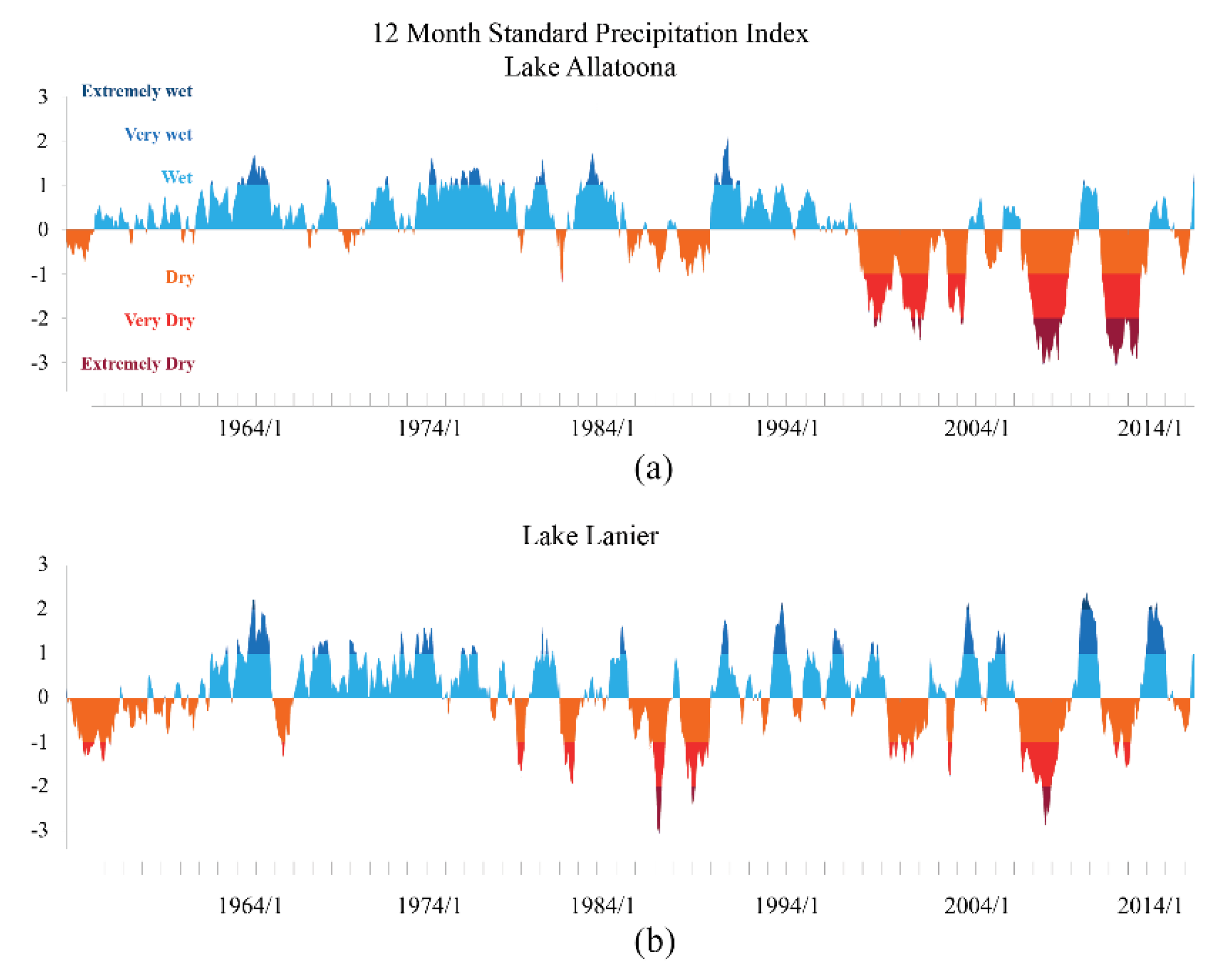
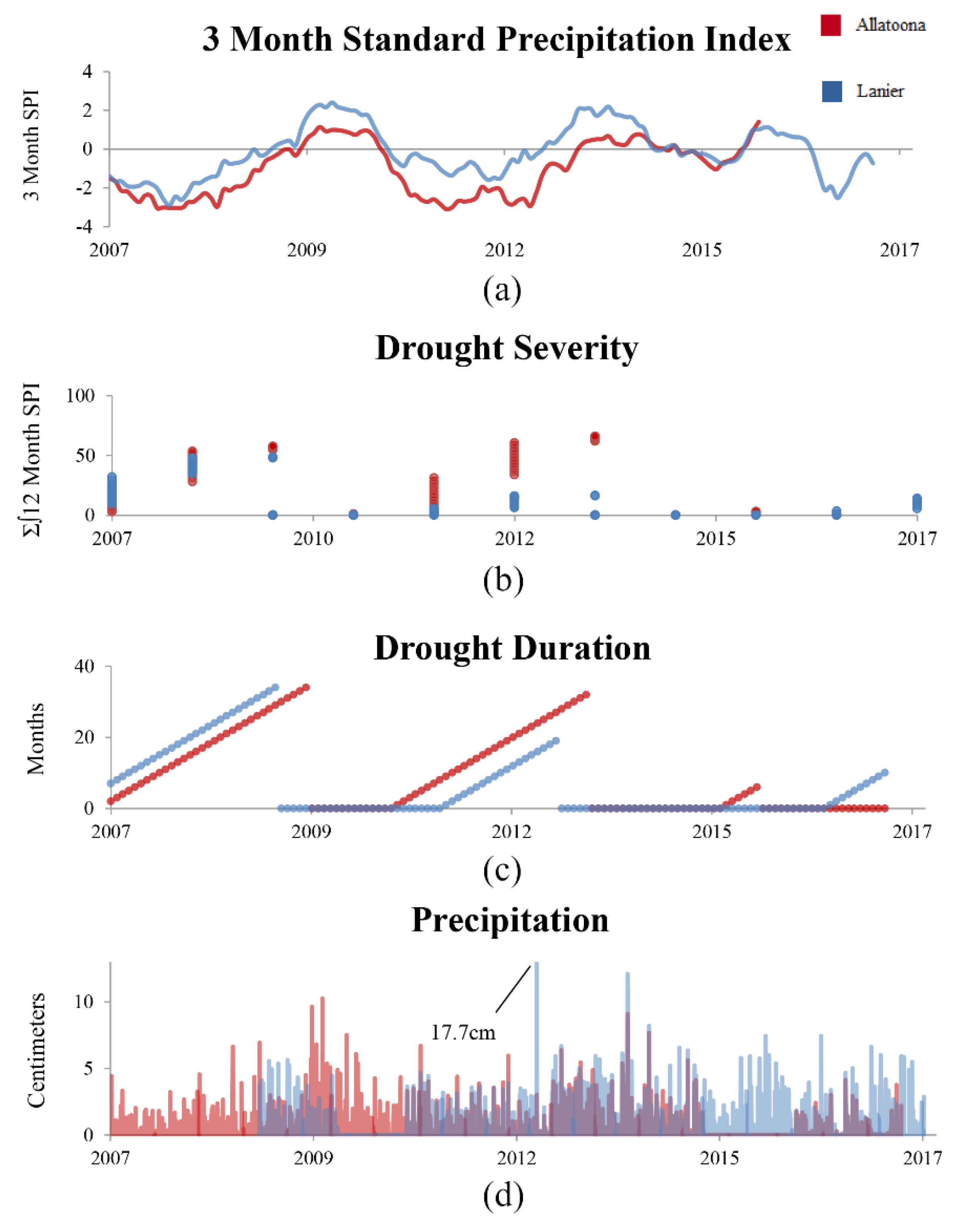
2.1. Drought
2.2. Rainstorms and Stormflow
2.3. Temperature and Water Quality
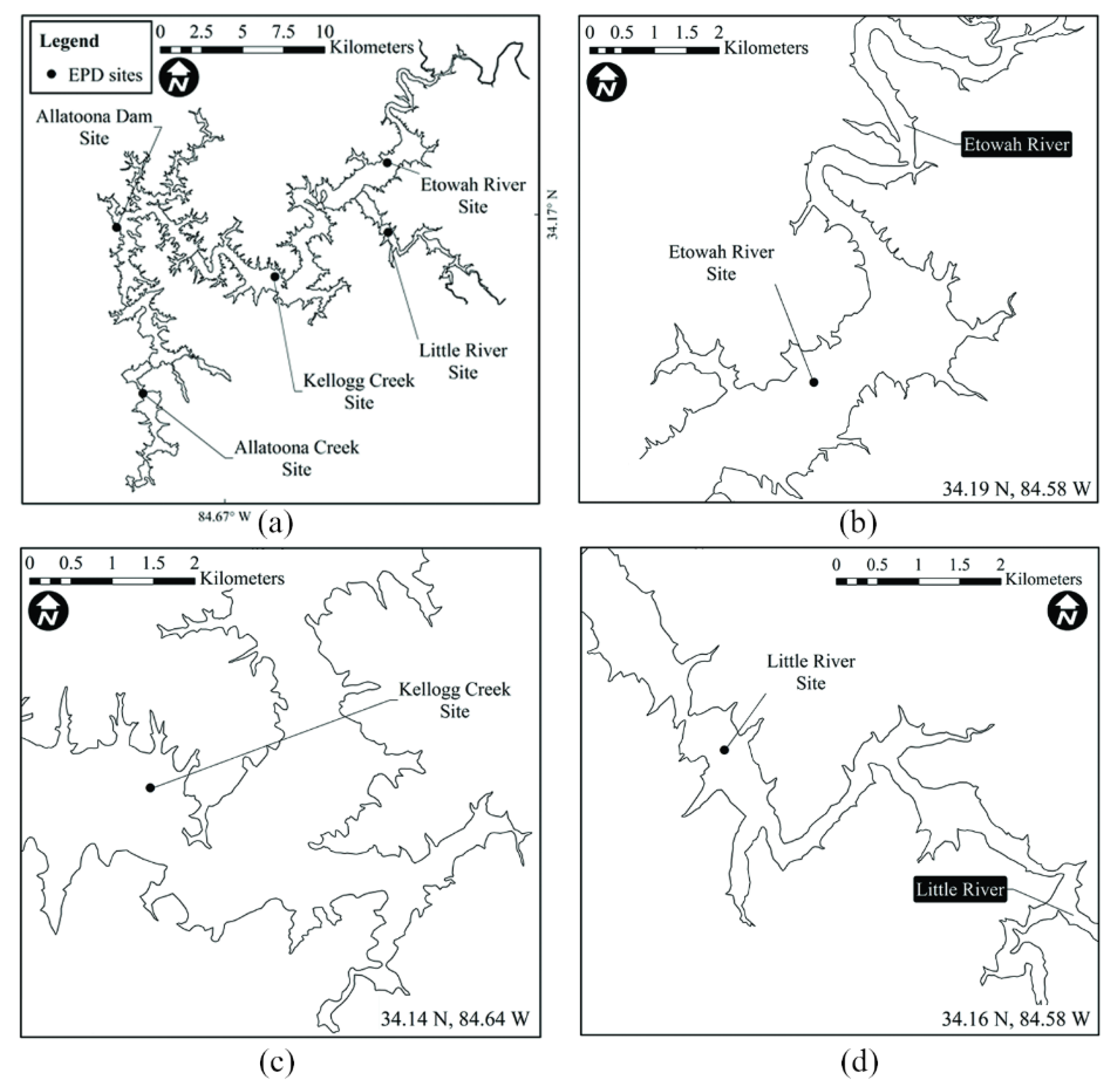
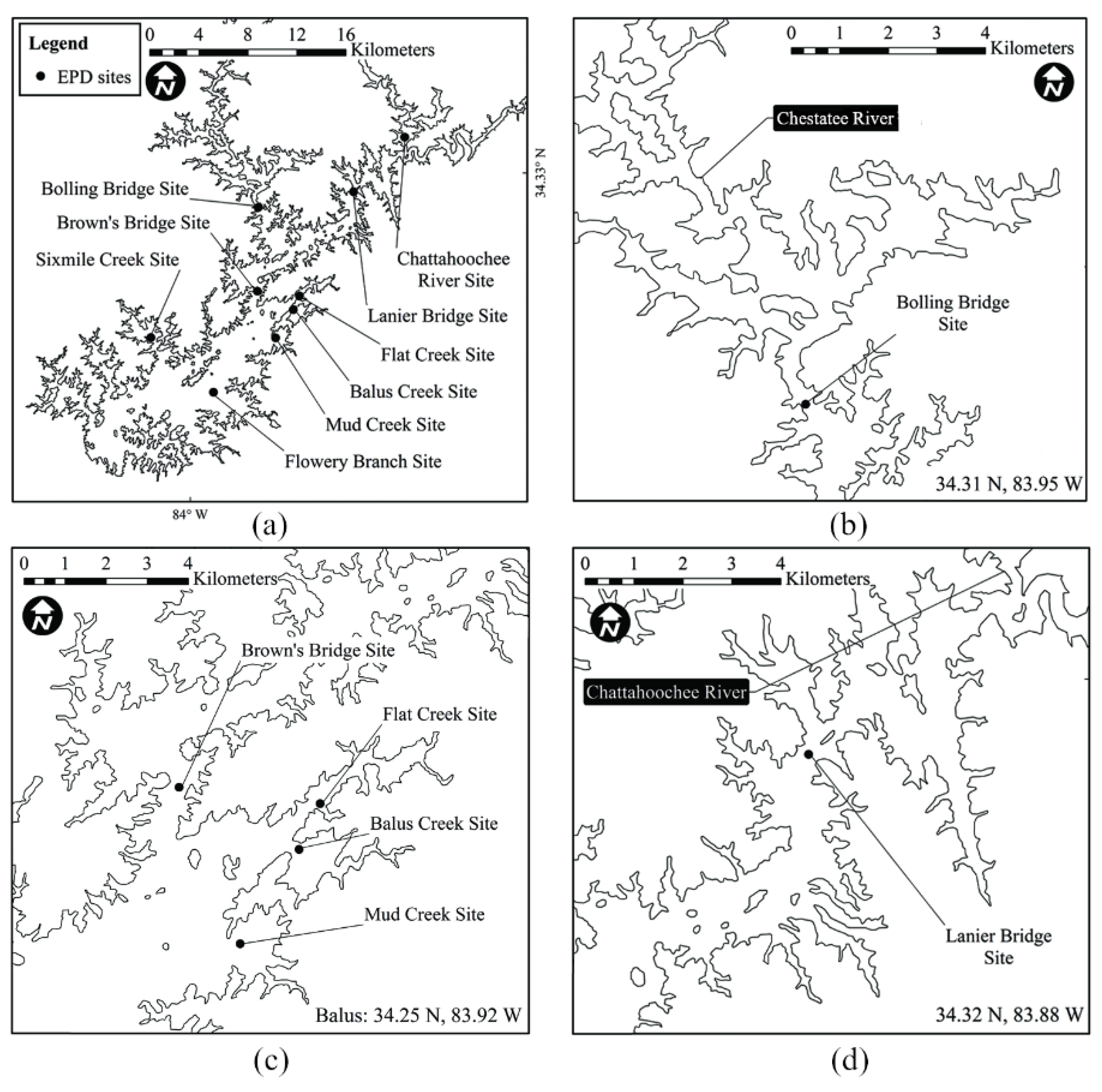
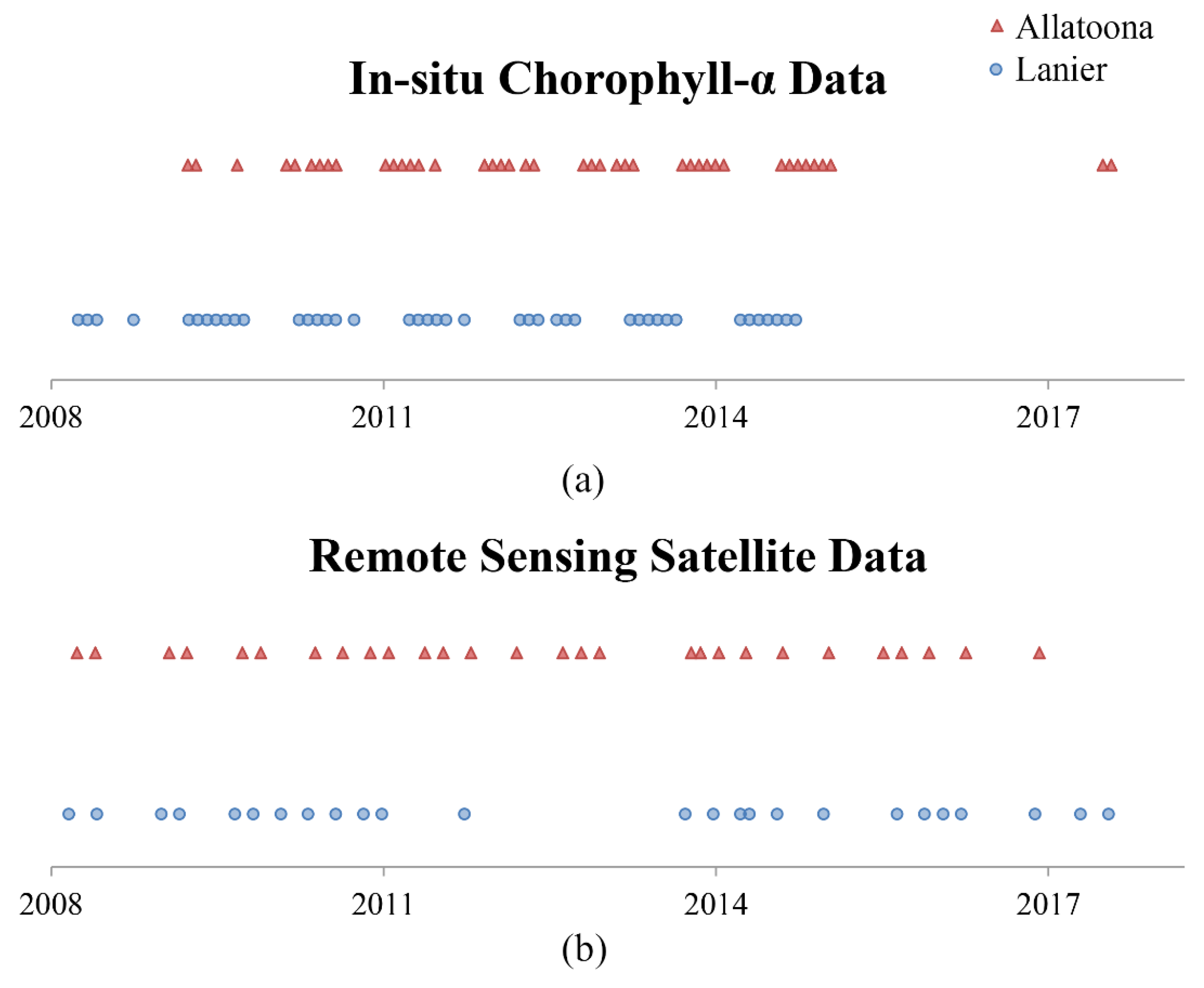
2.4. Algal Blooms
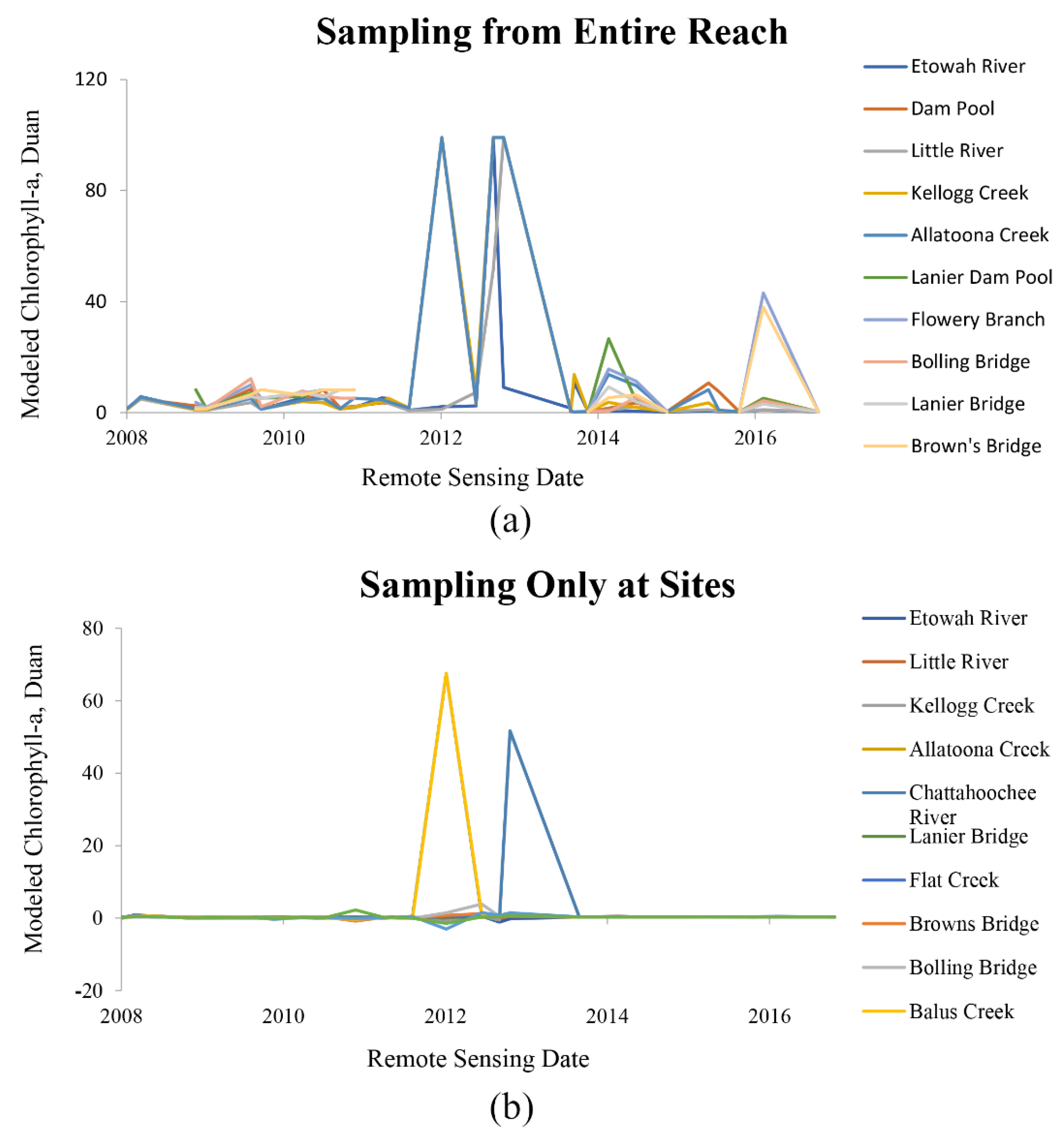
2.5. Modeling
3. Results
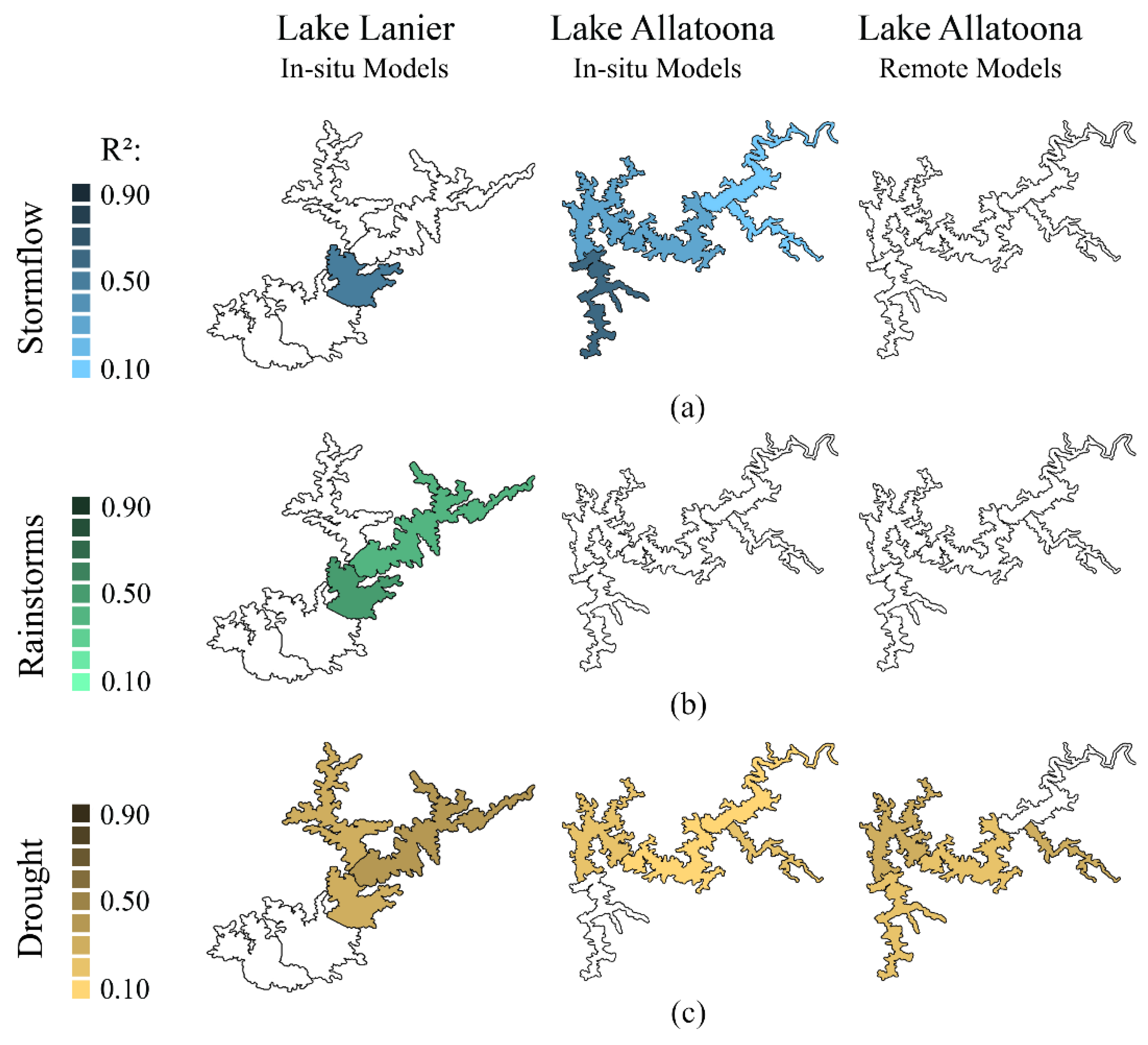
3.1. Dimensional Reduction
3.2. Linear Modeling of In Situ Data
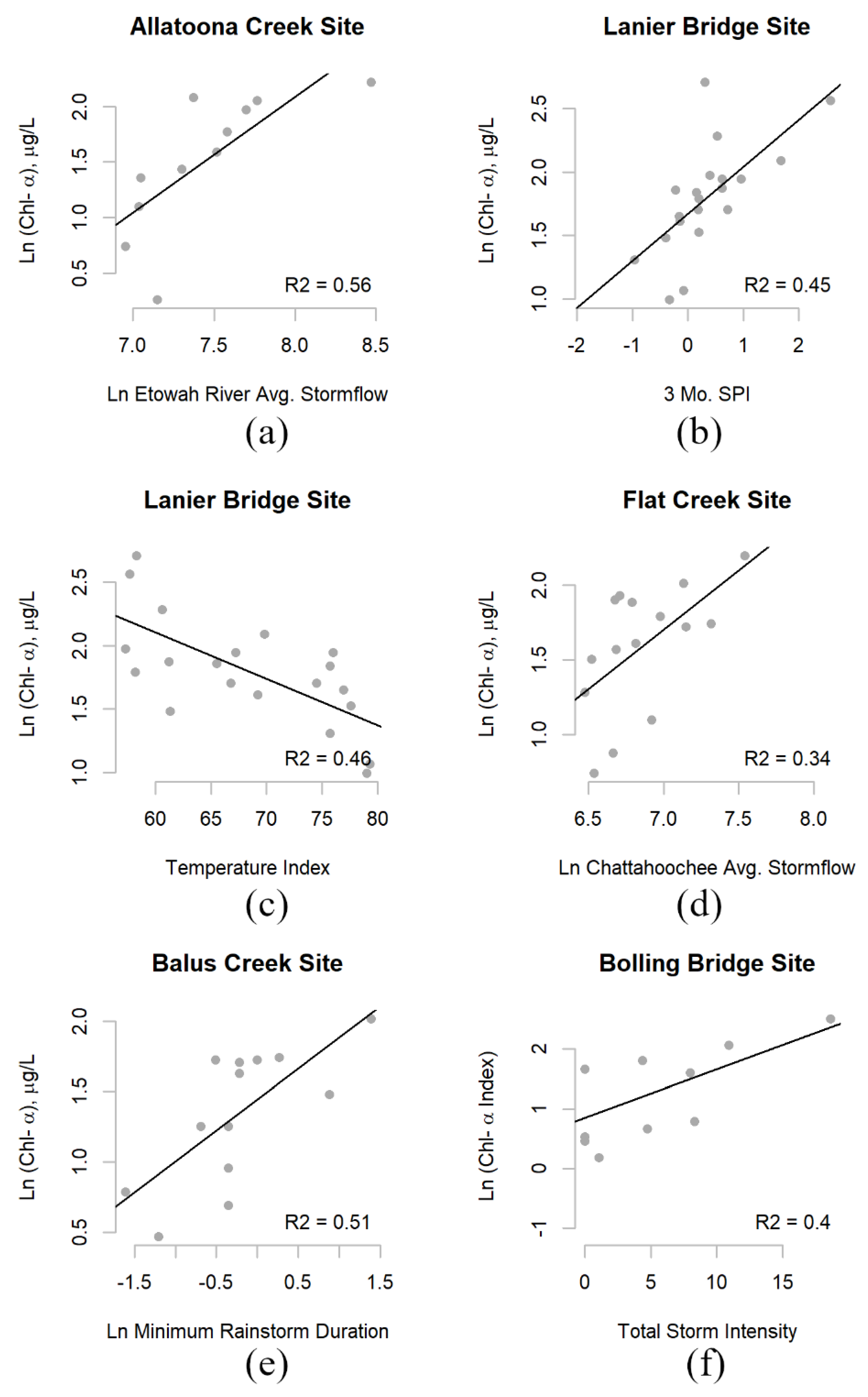
3.2.1. Stormflow
3.2.2. Rainstorms
3.2.3. Drought
3.2.4. Temperature and Chemistry
3.3. Linear Modeling of Remotely Sensed Data
4. Discussion
4.1. Lake Allatoona Models
4.2. Lake Lanier Models
4.3. Remote Sensing Models
4.4. Other Model Parameters
4.5. Model Limitations, Implications, and Future
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Unit | Lake Allatoona | Lake Sidney-Lanier |
|---|---|---|---|
| Reservoir Size 1 | km2 | 13 | 156 |
| Summer Conservation Elevation 1 | m, NGVD29 | 256 | 326 |
| Summer Conservation Storage 1 | km3 | 0.5 | 1.3 |
| Drainage Area 2 | km2 | 2900 | 2678 |
| Average Retention Time 3 | month | 4.2 | 10.2 |
| Range of Epilimnion Thickness 1 | m | 4–9 | 4–9 |
| Site | Model | R2 | Adj. R2 | F | DF | p-Value |
|---|---|---|---|---|---|---|
| Etowah River | ln (Chl-a) = 0.55 ln (Weekly Average Stormflow) − 1.10 | 0.06 | - | 0.6 | 1, 9 | 0.452 |
| Little River | ln (Chl-a) = 0.45 ln (Weekly Average Stormflow) − 0.43 | 0.12 | - | 1.27 | 1, 9 | 0.288 |
| Kellogg Creek | ln (Chl-a) = 0.90 ln (Weekly Average Stormflow) − 2.13 | 0.27 | - | 3.0 | 1, 9 | 0.101 |
| Allatoona Dam | ln (Chl-a) = 0.88 ln (Weekly Average Stormflow) − 2.24 | 0.25 | - | 3.0 | 1, 9 | 0.117 |
| Allatoona Creek | ln (Chl-a) = 1.04 ln (Weekly Average Stormflow) − 2.72 | 0.56 | - | 11.3 | 1, 9 | 0.008 |
| Site | Model | R2 | Adj. R2 | F | DF | p-Value |
|---|---|---|---|---|---|---|
| All Regression Models by Site: Lake Allatooona | ||||||
| Etowah River | ln (Chl-a) = −0.34 ln (Little River Weekly Min. Flow) + 3.36 | 0.15 | - | 6.5 | 1, 36 | 0.015 |
| ln (Chl-a) = −0.39 ln (Monthly Min. Flow) + 4.36 | 0.14 | - | 5.6 | 1, 34 | 0.024 | |
| ln (Chl-a) = −0.20 12 Mo. SPI + 1.81 | 0.13 | - | 5.6 | 1, 36 | 0.023 | |
| Kellogg Creek | ln (Chl-a) = −0.16 12 Mo. SPI + 2.26 | 0.13 | - | 5.3 | 1, 36 | 0.027 |
| Allatoona Dam | ln (Chl-a) = −0.16 12 Mo. SPI + 1.12 | 0.12 | - | 4.0 | 1, 38 | 0.032 |
| Allatoona Creek | ln (Chl-a) = 1.04 ln (Weekly Avg. Stormflow) – 6.25 | 0.56 | - | 11.3 | 1, 9 | 0.008 |
| All Regression Models by Site: Lake Lanier | ||||||
| Bolling Bridge | ln (Chl-a) = 0.37 3 Mo. SPI − 0.44 | 0.31 | - | 8.0 | 1, 18 | 0.011 |
| ln (Chl-a) = 0.26 NOAA 1 Mo. SPI + 0.19 | 0.27 | - | 6.5 | 1, 18 | 0.020 | |
| ln (Chl-a) = 0.11 ZNDX + 1.51 | 0.27 | - | 6.6 | 1, 18 | 0.019 | |
| ln (Chl-a) = 0.25 NOAA 3 Mo. SPI + 0.22 | 0.20 | - | 4.6 | 1, 18 | 0.047 | |
| Lanier Bridge | ln (Chl-a) = −0.04 Temperature Index + 4.30 | 0.46 | - | 15.5 | 1, 18 | 0.001 |
| ln (Chl-a) = 0.37 3 Mo. SPI − 0.19 | 0.45 | - | 14.5 | 1, 18 | 0.001 | |
| ln (Chl-a) = −0.10 Mo. Avg. Rainstorm Intensity + 2.32 | 0.36 | - | 6.8 | 1, 12 | 0.023 | |
| ln (Chl-a) = 0.24 NOAA 3 Mo. SPI + 0.55 | 0.26 | - | 6.4 | 1, 18 | 0.021 | |
| ln (Chl-a) = 0.18 NOAA 2 Mo. SPI + 0.86 | 0.23 | - | 5.3 | 1, 18 | 0.034 | |
| Flat Creek | ln (Chl-a) = 1.61 ln Monthly Avg. Stormflow − 9.53 | 0.49 | - | 12.4 | 1, 13 | 0.023 |
| Balus Creek | ln (Chl-a) = 0.44 ln (Mo. Min. Rainstorm Duration) + 1.44 | 0.51 | - | 11.4 | 1, 11 | 0.006 |
| ln (Chl-a) = 0.23 NOAA 1 Mo. SPI + 0.12 | 0.27 | - | 5.5 | 1, 15 | 0.033 | |
| Site | Model | R2 | Adj. R2 | F | DF | p-Value |
|---|---|---|---|---|---|---|
| All Regression Models by Site: Lake Allatoona | ||||||
| Allatoona Dam | ln (modeled chl-a) = 0.59 Drought Intensity + 0.23 | 0.27 | - | 8.9 | 1, 24 | 0.007 |
| ln (modeled chl-a) = −0.51 12 Mo. SPI + 3.25 | 0.19 | - | 5.7 | 1, 24 | 0.026 | |
| Little River | ln (modeled chl-a) = 0.40 ln (Drought Severity + 1) − 0.16 | 0.26 | - | 8.5 | 1, 24 | 0.003 |
| ln (modeled chl-a) = 0.50 ln (Drought Duration + 1) − 0.16 | 0.24 | - | 7.8 | 1, 24 | 0.007 | |
| Kellogg Creek | ln (modeled chl-a) = −0.50 12 Mo. SPI + 3.18 | 0.19 | - | 5.6 | 1, 24 | 0.026 |
| Allatoona Creek | ln (modeled chl-a) = 0.03 Drought Severity + 0.57 | 0.21 | - | 6.3 | 1, 24 | 0.019 |
| ln (modeled chl-a) = 0.51 Drought Intensity + 0.40 | 0.21 | - | 6.2 | 1, 24 | 0.020 | |
| ln (modeled chl-a) = −0.49 12 Mo. SPI + 3.23 | 0.18 | - | 5.2 | 1, 24 | 0.032 | |
| All Regression Models by Site: Lake Lanier | ||||||
| Bolling Bridge | ln (modeled chl-a) = 0.081 Monthly Total Rainstorm Intensity + 0.85 | 0.40 | - | 6.0 | 1, 9 | 0.036 |
References
- Heiskary, S.A. Developing Eutrophication Standards for Lakes and Reservoirs: A Report Prepared by the Lake Standards Subcommittee; North American Lake Management Society: Madison, WI, USA, 1992. [Google Scholar]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57. [Google Scholar] [CrossRef] [PubMed]
- Magrann, T.; Dunbar, S.G.; Boskovic, D.S.; Hayes, W.K. Impacts of Microcystis on algal biodiversity and use of new technology to remove Microcystis and dissolved nutrients. Lakes Reserv. Res. Manag. 2012, 17, 231–239. [Google Scholar] [CrossRef]
- Schindler, D.W. Evolution of phosphorus limitation in lakes. Science 1977, 195, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Meis, S.; Thackeray, S.J.; Jones, I.D. Effects of recent climate change on phytoplankton phenology in a temperate lake. Freshw. Boil. 2009, 54, 1888–1898. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef]
- Reichwaldt, E.S.; Ghadouani, A. Effects of rainfall patterns on toxic cyanobacterial blooms in a changing climate: Between simplistic scenarios and complex dynamics. Water Res. 2012, 46, 1372–1393. [Google Scholar] [CrossRef]
- Havens, K.E.; Ji, G.; Beaver, J.R.; Fulton, R.S.; Teacher, C.E. Dynamics of cyanobacteria blooms are linked to the hydrology of shallow Florida lakes and provide insight into possible impacts of climate change. Hydrobiologia 2019, 829, 43–59. [Google Scholar] [CrossRef]
- Ahn, C.-Y.; Chung, A.-S.; Oh, H.-M. Rainfall, phycocyanin, and N:P ratios related to cyanobacterial blooms in a Korean large reservoir. Hydrobiologia 2002, 474, 117–124. [Google Scholar] [CrossRef]
- Bloesch, J. Mechanisms, measurement and importance of sediment resuspension in lakes. Mar. Freshw. Res. 1995, 46, 295–304. [Google Scholar] [CrossRef]
- Robson, B.J.; Hamilton, D.P. Summer flow event induces a cyanobacterial bloom in a seasonal Western Australian estuary. Mar. Freshw. Res. 2003, 54, 139. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.A.; Jones, I.D.; Page, T. The importance of nutrient source in determining the influence of retention time on phytoplankton: An explorative modelling study of a naturally well-flushed lake. Hydrobiologia 2009, 627, 129–142. [Google Scholar] [CrossRef][Green Version]
- Bowling, L.C.; Merrick, C.; Swann, J.; Green, D.; Smith, G.; Neilan, B.A. Effects of hydrology and river management on the distribution, abundance and persistence of cyanobacterial blooms in the Murray River, Australia. Harmful Algae 2013, 30, 27–36. [Google Scholar] [CrossRef]
- Beaver, J.R.; Jensen, D.E.; Casamatta, D.A.; Tausz, C.E.; Scotese, K.C.; Buccier, K.M.; Teacher, C.E.; Rosati, T.C.; Minerovic, A.D.; Renicker, T.R. Response of phytoplankton and zooplankton communities in six reservoirs of the middle Missouri River (USA) to drought conditions and a major flood event. Hydrobiologia 2013, 705, 173–189. [Google Scholar] [CrossRef]
- Hutchinson, G.E. A Treatise on Limnology, Vol. I: Geography, Physics and Chemistry; John Wiley & Sons, Inc.: London, UK, 1957; ISBN 978-0471425687. [Google Scholar]
- Li, W.; Qin, B.; Zhu, G. Forecasting short-term cyanobacterial blooms in Lake Taihu, China, using a coupled hydrodynamic-algal biomass model. Ecohydrology 2014, 7, 794–802. [Google Scholar] [CrossRef]
- Dirnberger, J.M. The Role of Heat and Drought During Occasional Blooms of Picoplankton and Nanoplankton in a Large Drinking Water Reservoir (Allatoona Lake, Georgia). In Proceedings of the 2011 Georgia Water Resources Conference, Athens, GA, USA, 11–13 April 2011; Carroll, G.D., Ed.; Georgia Institute of Technology: Atlanta, GA, USA, 2011. [Google Scholar]
- Ní Longphuirt, S.; O’Boyle, S.; Wilkes, R.; Dabrowski, T.; Stengel, D. Influence of Hydrological Regime in Determining the Response of Macroalgal Blooms to Nutrient Loading in Two Irish Estuaries. Estuaries Coasts 2016, 39, 478–494. [Google Scholar] [CrossRef]
- Srifa, A.; Phlips, E.J.; Cichra, M.F.; Hendrickson, J.C. Phytoplankton dynamics in a subtropical lake dominated by cyanobacteria: Cyanobacteria ‘Like it Hot’ and sometimes dry. Aquat. Ecol. 2016, 50, 163–174. [Google Scholar] [CrossRef]
- Srifa, A.; Phlips, E.J.; Cichra, M.F.; Hendrickson, J.C. Effects of non-algal turbidity on cyanobacterial biomass in seven turbid Kansas reservoirs. Lake Reserv. Manag. 2011, 27, 6–14. [Google Scholar]
- Camacho, A.; Murueta, N.; Blasco, E.; Santamans, A.C.; Picazo, A. Hydrology-driven macrophyte dynamics determines the ecological functioning of a model Mediterranean temporary lake. Hydrobiologia 2016, 774, 93–107. [Google Scholar] [CrossRef]
- Royer, T.V.; David, M.B.; Gentry, L.E. Timing of Riverine Export of Nitrate and Phosphorus from Agricultural Watersheds in Illinois: Implications for Reducing Nutrient Loading to the Mississippi River. Environ. Sci. Technol. 2006, 40, 4126–4131. [Google Scholar] [CrossRef]
- Ceballos, E.L.; Rasmussen, T.C. Internal Loading in Southeastern Piedmont Impoundments. In Proceedings of the 2007 Georgia Water Resources Conference, Athens, GA, USA, 27–29 March 2007; Georgia Institute of Technology: Atlanta, GA, USA, 2007. [Google Scholar]
- Glibert, P.M.; Kelly, V.; Alexander, J.; Codispoti, L.A.; Boicourt, W.C.; Trice, T.M.; Michael, B. In situ nutrient monitoring: A tool for capturing nutrient variability and the antecedent conditions that support algal blooms. Harmful Algae 2008, 8, 175–181. [Google Scholar] [CrossRef]
- Holbach, A.; Wang, L.; Chen, H.; Hu, W.; Schleicher, N.; Zheng, B.; Norra, S. Water mass interaction in the confluence zone of the Daning River and the Yangtze River - a driving force for algal growth in the Three Gorges Reservoir. Environ. Sci. Pollut. Res. 2013, 20, 7027–7037. [Google Scholar] [CrossRef] [PubMed]
- McCullough, G.K.; Page, S.J.; Hesslein, R.H.; Stainton, M.P.; Kling, H.J.; Salki, A.G.; Barber, D.G. Hydrological forcing of a recent trophic surge in Lake Winnipeg. J. Great Lakes Res. 2012, 38, 95–105. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences, and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total. Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, V.M.; Roelke, D.L.; Grover, J.P.; Brooks, B.; Prosser, K.N.; Scott, W.C.; Laws, C.A.; Umphres, G.D. Interplay between ambient surface water mixing and manipulated hydraulic flushing: Implications for harmful algal bloom mitigation. Ecol. Eng. 2013, 60, 289–298. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, K.; Liu, J.; Deng, J.; Qin, B.; Zhu, G.; Zhou, Y. Meteorological and hydrological conditions driving the formation and disappearance of black blooms, an ecological disaster phenomena of eutrophication and algal blooms. Sci. Total. Environ. 2016, 569, 1517–1529. [Google Scholar] [CrossRef]
- Campana, P.; Knox, J.; Grundstein, A.; Dowd, J. The 2007–2009 Drought in Athens, Georgia, United States: A Climatological Analysis and an Assessment of Future Water Availability1. JAWRA J. Am. Water Resour. Assoc. 2012, 48, 379–390. [Google Scholar] [CrossRef]
- U.S. Army Corps of Engineers. Appendix A, Allatoona Dam and Lake. In Alabama-Coosa-Tallapoosa (ACT) River Basin Water Control Manual; U.S. Army Corps of Engineers: Mobile, AL, USA, 1952; revised 1962 and 2015; p. xv. Available online: https://www.sam.usace.army.mil/Portals/46/docs/planning_environmental/act/docs/New/Appendix%20A%20-%20Allatoona%20WCM%206%20May%2015.pdf?ver=2017-04-17-120210-283 (accessed on 1 February 2020).
- Dirnberger, J.M.; Weinberger, J. Influences of Lake Level Changes on Reservoir Water Clarity in Allatoona Lake, Georgia. Lake Reserv. Manag. 2005, 21, 24–29. [Google Scholar] [CrossRef][Green Version]
- Vincent, R.K.; Qin, X.; McKay, R.L.; Miner, J.; Czajkowski, K.; Savino, J.; Bridgeman, T. Phycocyanin detection from LANDSAT TM data for mapping cyanobacterial blooms in Lake Erie. Remote. Sens. Environ. 2004, 89, 381–392. [Google Scholar] [CrossRef]
- Duan, H.; Zhang, Y.; Zhang, B.; Song, K.; Wang, Z. Assessment of Chlorophyll-a Concentration and Trophic State for Lake Chagan Using Landsat TM and Field Spectral Data. Environ. Monit. Assess. 2007, 129, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Rasmussen, T.C. Short-term and long-term sediment and phosphorus inputs to Lake Lanier. In Proceedings of the 2001 Georgia Water Resources Conference, Athens, GA, USA, 26–27 April 2001; Hatcher, K.J., Ed.; Georgia Institute of Technology: Atlanta, GA, USA, 2001. [Google Scholar]
- Radcliffe, D.E.; Lin, Z.; Risse, L.M.; Romeis, J.J.; Jackson, C.R. Modeling Phosphorus in the Lake Allatoona Watershed Using SWAT: I. Developing Phosphorus Parameter Values. J. Environ. Qual. 2009, 38, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Environmental Protection Agency, U.S. EPA National Aquatic Resources Survey (NARS); Environmental Protection Agency: Washington, DC, USA, 2009.
- Environmental Protection Agency, U.S. EPA National Aquatic Resources Survey (NARS); Environmental Protection Agency: Washington, DC, USA, 2014.
- GAEPD AMU Soft Algae Taxa Data, Chattahoochee River Basin. Available online: https://gomaspublic.gaepd.org/ (accessed on 12 December 2019).
- U.S. Army Corps of Engineers. Appendix B, Buford Dam and Lake Sidney Lanier. In Apalachicola–Chattahoochee–Flint (ACF) River Basin Water Control Manual; U.S. Army Corps of Engineers: Mobile, AL, USA, 1959; revised 1991 and 2017; p. xiv.
- Griffith, G.E.; Omernik, J.M.; Comstock, J.A.; Lawrence, S.; Martin, G.; Goddard, A.; Hulcher, V.J.; Foster, T. Ecoregions of Alabama and Georgia (Color Poster with Map, Descriptive Text, Summary Tables, and Photographs); (map scale 1:1,700,000); U.S. Geological Survey: Reston, VA, USA, 2001.
- Kunkel, K.E.; Stevens, L.E.; Stevens, S.E.; Sun, L. Regional Climate Trends and Scenarios for the U.S. National Climate Assessment, Part 2 (NOAA Technical Report NESDIS 142-2); NOAA: Washington, DC, USA, 2013; p. 94.
- NCDC. The Climate of Georgia. Available online: https://www.ncdc.noaa.gov/climatenormals/clim60/states/Clim_GA_01.pdf (accessed on 30 January 2020).
- Georgia Environmental Protection Division. Total Maximum Daily Load Evaluation for Lake Allatoona in the Coosa River Basin for Chlorophyll a; Georgia Environmental Protection Division: Atlanta, GA, USA, 2013; p. 127.
- Georgia Environmental Protection Division. Final Total Maximum Daily Load Evaluation for Lake Lanier in the Chattahoochee River Basin for Chlorophyll a; Georgia Environmental Protection Division: Atlanta, GA, USA, 2017; p. 130.
- Alley, W.M. The Palmer drought severity index: Limitations and assumptions. J. Clim. Appl. Meteorol. 1984, 23, 1100–1109. [Google Scholar] [CrossRef]
- Guttman, N.B. Accepting the standardized precipitation index: A calculation algorithm. JAWRA J. Am. Water Res. Assoc. 1999, 35, 311–322. [Google Scholar] [CrossRef]
- NDMC National Drought Mitigation Center. Available online: http://drought.unl.edu (accessed on 1 June 2017).
- Edwards, D.C.; McKee, T.B. Characteristics of 20th Century Drought in the United States at Multiple Time Scales; Colorado State University: Fort Collins, CO, USA, 1997. [Google Scholar]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–23 January 1993; Volume 17, pp. 179–183. [Google Scholar]
- Svoboda, M.; Hayes, M.; Wood, D. Standardized Precipitation Index User Guide; World Meteorological Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Spinoni, J.; Naumann, G.; Carrao, H.; Barbosa, P.; Vogt, J. World drought frequency, duration, and severity for 1951–2010. Int. J. Clim. 2014, 34, 2792–2804. [Google Scholar] [CrossRef]
- Restrepo-Posada, P.J.; Eagleson, P.S. Identification of independent rainstorms. J. Hydrol. 1982, 55, 303–319. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, D.R.; Schluchter, W.M. A Novel Algorithm for Predicting Phycocyanin Concentrations in Cyanobacteria: A Proximal Hyperspectral Remote Sensing Approach. Remote. Sens. 2009, 1, 758–775. [Google Scholar] [CrossRef]
- Mishra, S.; Mishra, D.R. Normalized difference chlorophyll index: A novel model for remote estimation of chlorophyll-a concentration in turbid productive waters. Remote. Sens. Environ. 2012, 117, 394–406. [Google Scholar] [CrossRef]
- Knapp, A.; Milewski, A. Water Quality Data-Lake Allatoona, Georgia, U.S., 2017. Mendeley Data 2019, v2. Available online: http://dx.doi.org/10.17632/cr65yxbmvx.2 (accessed on 5 December 2019).
- Ortiz, J.D.; Avouris, D.; Schiller, S.; Luvall, J.C.; Lekki, J.D.; Tokars, R.P.; Anderson, R.C.; Shuchman, R.; Sayers, M.; Becker, R. Intercomparison of Approaches to the Empirical Line Method for Vicarious Hyperspectral Reflectance Calibration. Front. Mar. Sci. 2017, 4, 1–21. [Google Scholar] [CrossRef]
- Pinckney, J.; Millie, D.; Howe, K.; Paerl, H.; Hurley, J.; Robinson, C.J.; Goómez-Gutieérrez, J. Flow scintillation counting of 14 C-labeled microalgal photosynthetic pigments. J. Plankton Res. 1996, 18, 1867–1880. [Google Scholar] [CrossRef]
- Cotti-Rausch, B.E.; Lomas, M.W.; Lachenmyer, E.M.; Goldman, E.A.; Bell, D.W.; Goldberg, S.R.; Richardson, T.L. Mesoscale and sub-mesoscale variability in phytoplankton community composition in the Sargasso Sea. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2016, 110, 106–122. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 3, 18–22. Available online: http://cogns.northwestern.edu/cbmg/LiawAndWiener2002.pdf (accessed on 22 December 2019).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Delpla, I.; Baurès, E.; Jung, A.-V.; Thomas, O. Impacts of rainfall events on runoff water quality in an agricultural environment in temperate areas. Sci. Total. Environ. 2011, 409, 1683–1688. [Google Scholar] [CrossRef]
- Jones, I.D.; Elliott, J.A. Modelling the effects of changing retention time on abundance and composition of phytoplankton species in a small lake. Freshw. Boil. 2007, 52, 988–997. [Google Scholar] [CrossRef]
- An, K.-G.; Jones, J.R. Factors regulating bluegreen dominance in a reservoir directly influenced by the Asian monsoon. Hydrobiologia 2000, 432, 37–48. [Google Scholar] [CrossRef]
- Middleton, G.V. Sediment deposition from turbidity currents. Annu. Rev. Earth Planet. Sci. 1993, 21, 89–114. [Google Scholar] [CrossRef]









| Site | Driver | Model | R2/Adj. R2 | F/DF | p-Value |
|---|---|---|---|---|---|
| Etowah River | D | ln (Chl-a) = −0.34 Little River Weekly Min. Flow + 3.36 | 0.15 | 6.5/1, 36 | * |
| Kellogg Creek | D | ln (Chl-a) = −0.16 12 Mo. SPI + 2.26 | 0.13 | 5.3/1, 36 | * |
| Allatoona Dam | D | ln (Chl-a) = −0.16 12 Mo. SPI + 1.12 | 0.12 | 4.0/1, 38 | * |
| Allatoona Creek | W | ln (Chl-a) = 1.04 ln (Weekly Avg. Stormflow) − 6.25 | 0.56 | 11.3/1, 9 | * |
| Bolling Bridge | W | ln (Chl-a) = 0.37 3 Mo. SPI − 0.44 | 0.31 | 8.0/1, 18 | ** |
| Lanier Bridge | W | ln (Chl-a) = 0.37 3 Mo. SPI − 0.19 | 0.45 | 14.5/1, 18 | ** |
| Flat Creek | W | ln (Chl-a) = 1.61 ln Monthly Avg. Stormflow − 9.53 | 0.49 | 12.4/1, 13 | ** |
| Balus Creek | W | ln (Chl-a) = 0.44 ln (Monthly Min. Rainstorm Duration) + 1.44 | 0.51 | 11.4/1, 11 | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knapp, A.S.; Milewski, A.M. Spatiotemporal Relationships of Phytoplankton Blooms, Drought, and Rainstorms in Freshwater Reservoirs. Water 2020, 12, 404. https://doi.org/10.3390/w12020404
Knapp AS, Milewski AM. Spatiotemporal Relationships of Phytoplankton Blooms, Drought, and Rainstorms in Freshwater Reservoirs. Water. 2020; 12(2):404. https://doi.org/10.3390/w12020404
Chicago/Turabian StyleKnapp, Abigail S., and Adam M. Milewski. 2020. "Spatiotemporal Relationships of Phytoplankton Blooms, Drought, and Rainstorms in Freshwater Reservoirs" Water 12, no. 2: 404. https://doi.org/10.3390/w12020404
APA StyleKnapp, A. S., & Milewski, A. M. (2020). Spatiotemporal Relationships of Phytoplankton Blooms, Drought, and Rainstorms in Freshwater Reservoirs. Water, 12(2), 404. https://doi.org/10.3390/w12020404