Contrasting Effects of Bioturbation Studied in Intact and Reconstructed Estuarine Sediments

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling Activities
2.2.1. Reconstructed Sediment Cores
2.2.2. Intact Sediments
2.3. Dark Incubations for Flux and Denitrification Measurements
2.4. Statistical Analysis
3. Results and Discussion
3.1. General Features of the Study Sites
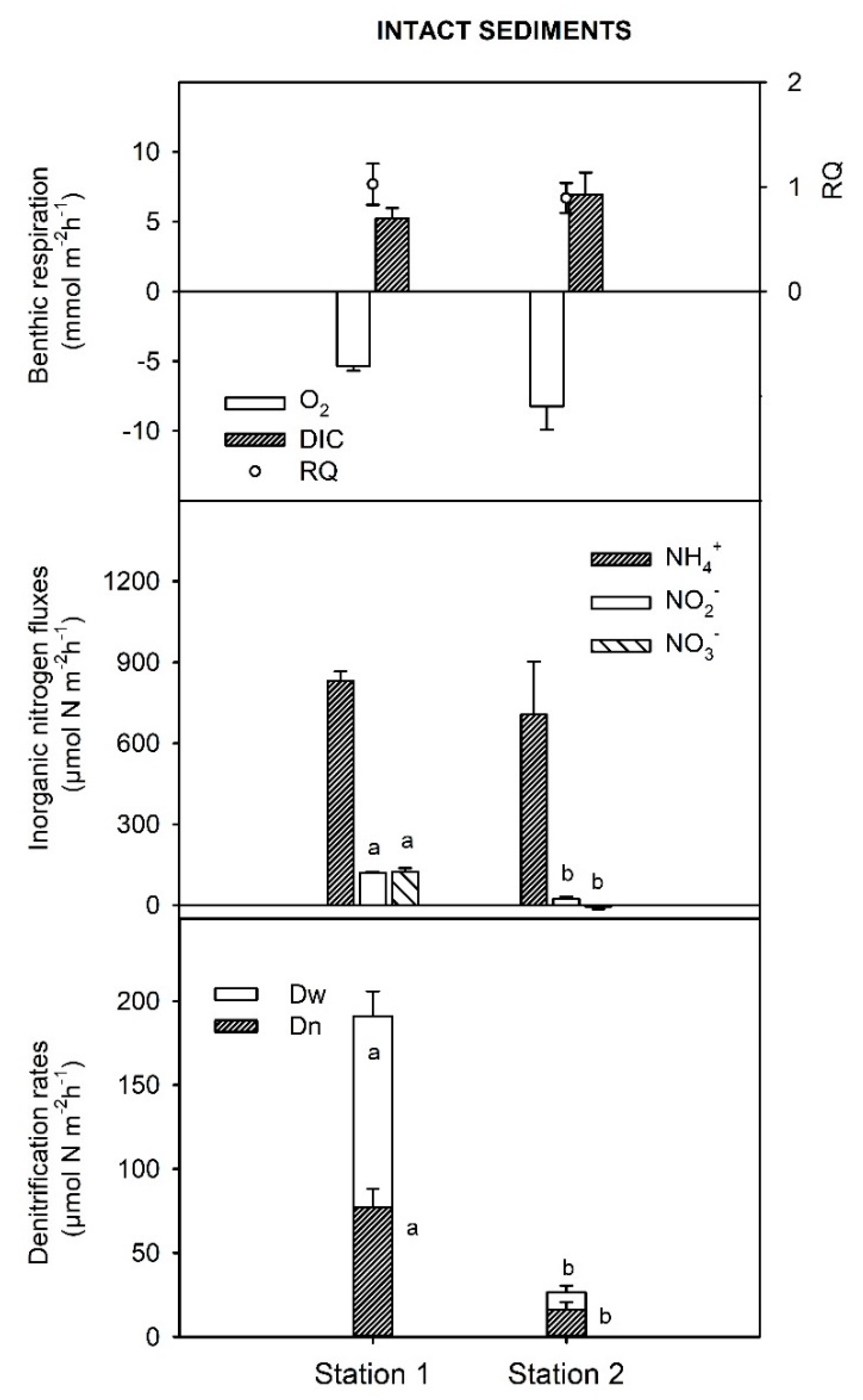
3.2. Benthic Respiration, DIN Fluxes, and Rates of Denitrification in Intact Sediment Cores
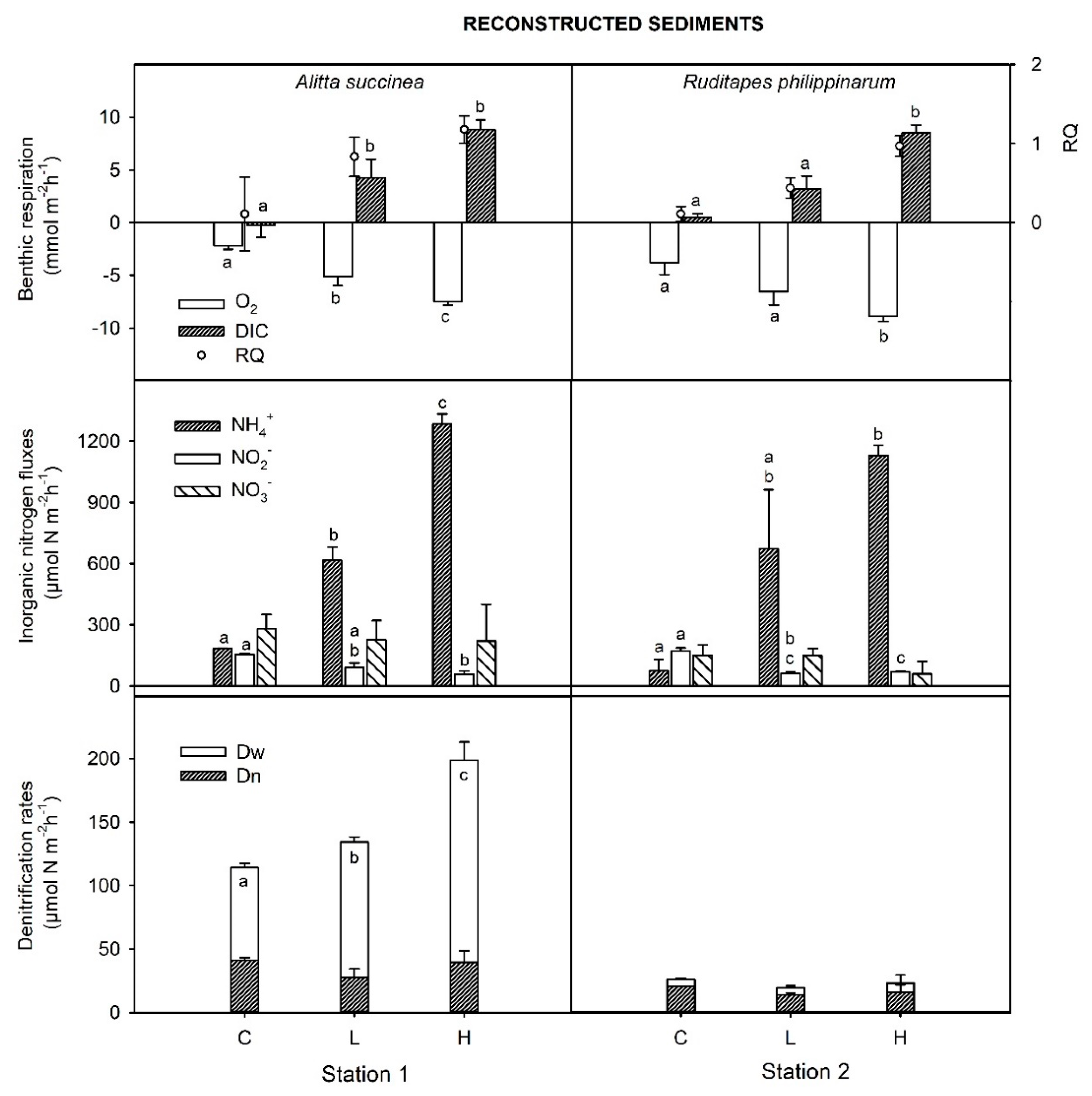
3.3. Benthic Respiration, DIN Fluxes and Rates of Denitrification in Reconstructed Cores
3.4. Macrofauna Stimulate Differentially Biogeochemical Processes in Intact and in Reconstructed Sediments
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aller, R.C. Bioturbation and remineralization of sedimentary organic matter: Effects of redox oscillation. Chem. Geol. 1994, 114, 331–345. [Google Scholar] [CrossRef]
- Welsh, D.T. It’s a dirty job but someone has to do it: The role of marine benthic macrofauna in organic matter turnover and nutrient recycling to the water column. Chem. Ecol. 2003, 19, 321–342. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Rosenberg, R.; Fluviaux, H.; Claude, U.; Lyon, B. Ecosystem engineering: The impact of bioturbation on biogeochemical processes in marine and freshwater benthic habitats. Aquat. Sci. 2006, 68, 434–442. [Google Scholar] [CrossRef]
- Laverock, B.; Gilbert, J.A.; Tait, K.; Osborn, A.M.; Widdicombe, S. Bioturbation: Impact on the marine nitrogen cycle. Biochem. Soc. Trans. 2011, 39, 315–320. [Google Scholar] [CrossRef]
- Stief, P. Stimulation of microbial nitrogen cycling in aquatic ecosystems by benthic macrofauna: Mechanisms and environmental implications. Biogeosciences 2013, 10, 7829–7846. [Google Scholar] [CrossRef]
- Zilius, M.; Bonaglia, S.; Broman, E.; Chiozzini, V.G.; Samuiloviene, A.; Nascimento, F.J.A.; Cardini, U.; Bartoli, M. N2 fixation dominates nitrogen cycling in a mangrove fiddler crab holobiont. Sci. Rep. 2020, 10, 13966. [Google Scholar] [CrossRef]
- Gergs, R.; Rinke, K.; Rothhaupt, K.O. Zebra mussels mediate benthic-pelagic coupling by biodeposition and changing detrital stoichiometry. Freshw. Biol. 2009, 54, 1379–1391. [Google Scholar] [CrossRef]
- Benelli, S.; Bartoli, M.; Ribaudo, C.; Fano, E.A. Contrasting effects of an alienworm on benthic N cycling in muddy and sandy sediments. Water 2019, 11, 465. [Google Scholar] [CrossRef]
- Swanberg, I.L. The influence of the filter-feeding bivalve Cerastoderma edule L. on microphytobenthos: A laboratory study. J. Exp. Mar. Biol. Ecol. 1991, 151, 93–111. [Google Scholar] [CrossRef]
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshw. Biol. 2001, 1431–1446. [Google Scholar] [CrossRef]
- Sandwell, D.R.; Pilditch, C.A.; Lohrer, A.M. Density dependent effects of an infaunal suspension-feeding bivalve (Austrovenus stutchburyi) on sand flat nutrient fluxes and microphytobenthic productivity. J. Exp. Mar. Biol. Ecol. 2009, 373, 16–25. [Google Scholar] [CrossRef]
- Asmus, R.M.; Asmus, H. Mussel beds: Limiting or promoting phytoplankton? J. Exp. Mar. Biol. Ecol. 1991, 148, 215–232. [Google Scholar] [CrossRef]
- Nizzoli, D.; Welsh, D.T.; Viaroli, P. Seasonal nitrogen and phosphorus dynamics during benthic clam and suspended mussel cultivation. Mar. Pollut. Bull. 2011, 62, 1276–1287. [Google Scholar] [CrossRef] [PubMed]
- Benelli, S.; Bartoli, M.; Racchetti, E.; Carpintero, P.; Mindaugas, M.; Lubiene, I.; Anna, E. Rare but large bivalves alter benthic respiration and nutrient recycling in riverine sediments. Aquat. Ecol. 2017, 51, 1–16. [Google Scholar] [CrossRef]
- Atkinson, C.L.; First, M.R.; Covich, A.P.; Opsahl, S.P.; Golladay, S.W. Suspended material availability and filtration—Biodeposition processes performed by a native and invasive bivalve species in streams. Hydrobiologia 2011, 667, 191–204. [Google Scholar] [CrossRef]
- Murphy, A.E.; Anderson, I.C.; Smyth, A.R.; Song, B.; Luckenbach, M.W. Microbial nitrogen processing in hard clam (Mercenaria mercenaria) aquaculture sediments: The relative importance of denitrification and dissimilatory nitrate reduction to ammonium (DNRA). Limnol. Oceanogr. 2016, 1589–1604. [Google Scholar] [CrossRef]
- Kristensen, E.; Kostka, J.E. Macrofaunal burrows and irrigation in marine sediment: Microbiological and biogeochemical interactions. In Interac-Tions between Macro- and Microorganisms in Marine Sediments; Kristensen, E., Haese, R.R.K.J., Eds.; American Geophysical Union: Washington, DC, USA, 2005; pp. 125–158. [Google Scholar]
- Murphy, E.A.K.; Reidenbach, M.A. Oxygen transport in periodically ventilated polychaete burrows. Mar. Biol. 2016, 163, 1–14. [Google Scholar] [CrossRef]
- Benelli, S.; Bartoli, M.; Zilius, M.; Vybernaite-Lubiene, I.; Ruginis, T.; Petkuviene, J.; Fano, E.A. Microphytobenthos and chironomid larvae attenuate nutrient recycling in shallow-water sediments. Freshw. Biol. 2018, 63, 187–201. [Google Scholar] [CrossRef]
- Volkenborn, N.; Woodin, S.A.; Wethey, D.S.; Polerecky, L. Bioirrigation in Marine Sediments. In Reference Module in Earth Systems and Environmental Sciences; Elias, S.A., Ed.; Elsevier Inc.: Amsertam, The Netherlands, 2016; pp. 1–9. ISBN 9780124095489. [Google Scholar]
- Hölker, F.; Vanni, M.J.; Kuiper, J.J.; Christof, M.; Grossart, H.-P.; Stief, P.; Adrian, R.; Lorke, A.; Dellwig, O.; Brand, A.; et al. Tube-dwelling invertebrates: Tiny ecosystem engineers have large effects in lake ecosystems. Ecol. Monogr. 2015, 85, 333–351. [Google Scholar] [CrossRef]
- Moraes, P.C.; Zilius, M.; Benelli, S.; Bartoli, M. Nitrification and denitrification in estuarine sediments with tube-dwelling benthic animals. Hydrobiologia 2018, 819, 217–230. [Google Scholar] [CrossRef]
- Mermillod-blondin, F.; Lemoine, D.G. Ecosystem engineering by tubificid worms stimulates macrophyte growth in poorly oxygenated wetland sediments. Funct. Ecol. 2010, 24, 444–453. [Google Scholar] [CrossRef]
- Herren, C.M.; Webert, K.; Drake, M.D.; Vander Zanden, M.J.; Einarsson, A.; Ives, A.R.; Gratton, C. Positive feedback between chironomids and algae creates net mutualism between benthic primary consumers and producers. Ecology 2017, 98, 447–455. [Google Scholar] [CrossRef]
- Magri, M.; Benelli, S.; Bondavalli, C.; Bartoli, M. Benthic N pathways in illuminated and bioturbated sediments studied with network analysis. Limnol. Oceanogr. 2018, 63, S68–S84. [Google Scholar] [CrossRef]
- Van der Heide, T.; Govers, L.L.; De Fouw, J.; Olff, H.; Van Der Geest, M.; Van Katwijk, M.M.; Piersma, T.; Van De Koppel, J.; Silliman, B.R.; Smolders, A.J.P.; et al. A three-stage symbiosis forms the foundation of seagrass ecosystems. Science 2012, 336, 1432–1434. [Google Scholar] [CrossRef] [PubMed]
- Svensson, J.; Enrich-Prast, A.; Leonardson, L. Nitrification and dentrification in a eutrophic lake sediment bioturbated by oligochaetes. Aquat. Microb. Ecol. 2001, 23, 177–186. [Google Scholar] [CrossRef]
- Nizzoli, D.; Bartoli, M.; Cooper, M.; Welsh, D.T.; Underwood, G.J.C.; Viaroli, P. Implications for oxygen, nutrient fluxes and denitrification rates during the early stage of sediment colonisation by the polychaete Nereis spp. in four estuaries. Estuar. Coast. Shelf Sci. 2007, 75, 125–134. [Google Scholar] [CrossRef]
- De Backer, A.; Van Coillie, F.; Provoost, P.; Van Colen, C.; Vincx, M.; Degraer, S. Bioturbation effects of Corophium volutator: Importance of density and behavioural activity. Estuar. Coast. Shelf Sci. 2011, 91, 306–313. [Google Scholar] [CrossRef]
- Bonaglia, S.; Bartoli, M.; Gunnarsson, J.S.; Rahm, L.; Raymond, C.; Svensson, O.; Yekta, S.S.; Brüchert, V. Effect of reoxygenation and Marenzelleria spp. bioturbation on Baltic Sea sediment metabolism. Mar. Ecol. Prog. Ser. 2013, 482, 43–55. [Google Scholar] [CrossRef]
- Pelegrì, S.P.; Blackburn, T.H. Bioturbation effects of the amphipod Corophium volutator on microbial nitrogen transformations in marine sediments. Mar. Biol. 1994, 121, 253–258. [Google Scholar] [CrossRef]
- Stocum, E.T.; Plante, C.J. The effect of artificial defaunation on bacterial assemblages of intertidal sediments. J. Exp. Mar. Biol. Ecol. 2006, 337, 147–158. [Google Scholar] [CrossRef]
- Kauppi, L.; Bernard, G.; Bastrop, R.; Norkko, A.; Norkko, J. Increasing densities of an invasive polychaete enhance bioturbation with variable effects on solute fluxes. Sci. Rep. 2018, 65, 1–12. [Google Scholar] [CrossRef]
- Braeckman, U.; Provoost, P.; Gribsholt, B.; Van Gansbeke, D.; Middelburg, J.J.; Soetaert, K.; Vincx, M.; Vanaverbeke, J. Role of macrofauna functional traits and density in biogeochemical fluxes and bioturbation. Mar. Ecol. Prog. Ser. 2010, 399, 173–186. [Google Scholar] [CrossRef]
- Viaroli, P.; Giordani, G.; Bartoli, M.; Naldi, M.; Azzoni, R.; Nizzoli, D.; Ferrari, I.; Comenges, J.M.Z.; Bencivelli, S.; Castaldelli, G.; et al. The Sacca di Goro lagoon and an arm of the Po River. In Estuaries; Springer: Berlin/Heidelberg, Germany, 2006; Volume 5, pp. 197–232. [Google Scholar]
- Bartoli, M.; Benelli, S.; Lauro, M.; Magri, M.; Vybernaite-Lubiene, I.; Petkuviene, J. Variable oxygen levels lead to variable stoichiometry of benthic nutrient fluxes in a hypertrophic estuary. Estuaries Coasts 2020. [Google Scholar] [CrossRef]
- Magri, M.; Benelli, S.; Bonaglia, S.; Zilius, M.; Castaldelli, G.; Bartoli, M. The effects of hydrological extremes on denitrification, dissimilatory nitrate reduction to ammonium (DNRA) and mineralization in a coastal lagoon. Sci. Total Environ. 2020, 740, 140169. [Google Scholar] [CrossRef]
- Dalsgaard, T.; Nielsen, L.P.; Brotas, V.; Viaroli, P.; Underwood, G.J.C.; Nedwell, D.B.; Sundbäck, K.; Rysgaard, S.; Miles, A.; Bartoli, M.; et al. Protocol Handbook for NICE-Nitrogen Cycling in Estuaries: A Project under the EU Research Programme: Marine Science and Technology (MAST III); Ministry of Environment and Energy National Environmental Research Institute, Denmark© Department of Lake and Estuarine Ecology: Silkeborg, Denmark, 2000; ISBN 8777725352. [Google Scholar]
- Anderson, L.G.; Hall, P.O.J.; Iverfeldt, A.; Rutgers van der Loejf, M.M.; Sundby, B.; Westerlund, S.F.G. Benthic respiration measured by total carbonate production. Limnol. Oceanogr. 1986, 31, 319–329. [Google Scholar] [CrossRef]
- Bower, C.E.; Holm-Hansen, T. A salicylate-hypochlorite method for determining ammonia in seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Golterman, H.L.; Clymo, L.S.; Ohnstand, M.A.M. Methods for physical and chemical analysis of freshwaters. In Handbook, Blackwell Science, 2nd ed.; I.B.P: Oxford, UK, 1978; Volume 8. [Google Scholar]
- APHA (American Public Health Association). Standard Methods for the Examination of Water and Wastewaters, 18th ed.; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Nielsen, L.P. Denitrification in sediment determined from nitrogen isotope pairing. FEMS Microbiol. Lett. 1992, 86, 357–362. [Google Scholar] [CrossRef]
- Kana, T.M.; Darkangelo, C.; Hunt, M.D.; Oldham, J.B.; Bennett, G.E.; Cornwell, J.C. Membrane Inlet Mass Spectometer for rapid high-precision determination of N2, O2, and Ar in environment water samples. Anal. Chem. 1994, 66, 4166–4170. [Google Scholar] [CrossRef]
- Eyre, B.D.; Ferguson, A.J.P. Comparison of carbon production and decomposition, benthic nutrient fluxes and denitrification in seagrass, phytoplankton, benthic microalgae- and macroalgae- dominated warm-temperate Australian lagoons. Mar. Ecol. Prog. Ser. 2002, 229, 43–59. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Nizzoli, D.; Bartoli, M.; Viaroli, P. Oxygen and ammonium dynamics during a farming cycle of the bivalve Tapes philippinarum. Hydrobiologia 2007, 587, 25–36. [Google Scholar] [CrossRef]
- Pelegri, S.P.; Blackburn, T.H. Effect of bioturbation by Nereis sp., Mya Arenaria and Cerastoderma sp. on nitrification and denitrification in estuarine sediments. Ophelia 1995, 42, 289–299. [Google Scholar] [CrossRef]
- Bosch, J.A.; Cornwell, J.C.; Kemp, W.M. Short-term effects of nereid polychaete size and density on sediment inorganic nitrogen cycling under varying oxygen conditions. Mar. Ecol. Prog. Ser. 2015, 524, 155–169. [Google Scholar] [CrossRef]
- Bartoli, M.; Nizzoli, D.; Viaroli, P.; Turolla, E.; Castaldelli, G.; Fano, E.A.; Rossi, R. Impact of Tapes philippinarum farming on nutrient dynamics and benthic respiration in the Sacca di Goro. Hydrobiologia 2001, 455, 203–212. [Google Scholar] [CrossRef]
- Welsh, D.T.; Nizzoli, D.; Fano, E.A.; Viaroli, P. Direct contribution of clams (Ruditapes philippinarum) to benthic fluxes, nitrification, denitrification and nitrous oxide emission in a farmed sediment. Estuar. Coast. Shelf Sci. 2015, 154, 84–93. [Google Scholar] [CrossRef]
- Murphy, A.E.; Nizzoli, D.; Bartoli, M.; Smyth, A.R.; Castaldelli, G.; Anderson, I.C. Variation in benthic metabolism and nitrogen cycling across clam aquaculture sites. Mar. Pollut. Bull. 2018, 127, 524–535. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sediment Features | |||||||
| Density | Porosity | Organic Matter | C | N | Chl a | Macrofauna | |
| (g mL−1) | (%) | (%) | (%) | (mg m−2) | (gdw m−2) | ||
| Station 1 | 1.14 ± 0.11 | 0.74 ± 0.06 | 6.03 ± 0.07 | 1.79 ± 0.04 | 0.20 ± 0.01 | 37.3 ± 9.8 | 16.0 ± 9.1 |
| Station 2 | 1.74 ± 0.02 | 0.47 ± 0.01 | 1.61 ± 0.08 | 0.35 ± 0.02 | 0.04 ± 0.00 | 4.7 ± 0.8 | 220.8 ± 292.1 |
| Granulometry | |||||||
| <0.063 mm | 0.063 < x < 0.125 mm | 0.125 < x < 0.5 mm | 0.5 < x < 2 mm | >2 mm | |||
| (%) | (%) | (%) | (%) | (%) | |||
| Station 1 | 24.4 | 22.0 | 50.4 | 3.1 | 0 | ||
| Station 2 | 10.7 | 35.2 | 51.3 | 2.7 | 0 | ||
| Water features | |||||||
| Temperature | Salinity | pH | NH4+ | NO2− | NO3− | ||
| (°C) | (µM) | (µM) | (µM) | ||||
| Station 1 | 25 | 5 | 7.77 | 1.8 | 6.6 | 31.6 | |
| Station 2 | 25 | 17 | 7.96 | 14.6 | 1.3 | 3.1 | |
| Alitta succinea | Ruditapes philippinarum | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Intact Sediments | Reconstructed Sediments | Intact Sediments | Reconstructed Sediments | |||||||||||||
| Slope | Intercept | p | r2 | Slope | Intercept | p | r2 | Slope | Intercept | p | r2 | Slope | Intercept | p | r2 | |
| O2 | −81.75 (25.67) | −4031.78 (461.74) | 0.033 | 0.72 | −158.53 (13.25) | −2283.70 (291.41) | <0.001 | 0.95 | −12.80 (2.94) | −5417.04 (1016.76) | 0.012 | 0.83 | −6.98 (2.27) | −4149.12 (969.48) | 0.018 | 0.57 |
| DIC | −137.11 (75.55) | 7417.94 (1359.12) | 0.144 | 0.45 | 264.35 (45.38) | −144.03 (997.82) | <0.001 | 0.83 | 12.57 (2.26) | 4150.56 (781.78) | 0.005 | 0.89 | 12.09 (1.75) | 112.07 (748.16) | <0.001 | 0.87 |
| RQ | −0.04 (0.02) | 1.71 (0.29) | 0.055 | 0.64 | 0.04 (0.01) | −0.09 (0.28) | 0.015 | 0.59 | −0.0001 (0.0006) | 0.918 (0.207) | 0.850 | 0.01 | 0.001 (0.0002) | 0.067 (0.082) | <0.001 | 0.88 |
| NH4+ | 5.90 (3.86) | 736.22 (69.37) | 0.201 | 0.37 | 31.43 (2.71) | 169.36 (59.60) | <0.001 | 0.95 | 0.87 (0.69) | 514.09 (239.52) | 0.276 | 0.28 | 1.47 (0.40) | 145.15 (169.82) | 0.008 | 0.66 |
| NO2− | 1.13 (0.45) | −101.37 (8.12) | 0.067 | 0.61 | −2.66 (0.71) | 145.65 (15.53) | 0.007 | 0.67 | −0.03 (0.02) | 30.42 (7.14) | 0.286 | 0.27 | −0.15 (0.05) | 149.04 (19.52) | 0.014 | 0.60 |
| NO3− | −2.46 (1.63) | 161.53 (29.26) | 0.205 | 0.36 | −2.19 (4.68) | 279.11 (102.81) | 0.654 | 0.03 | −0.05 (0.02) | 5.76(8.53) | 0.103 | 0.53 | −0.09 (0.11) | 148.70 (46.32) | 0.458 | 0.08 |
| DIN | 4.57 (4.49) | 999.12 (80.79) | 0.367 | 0.21 | 26.59 (4.77) | 594.12 (104.93) | <0.001 | 0.82 | 0.79 (0.68) | 550.26 (234.96) | 0.307 | 0.26 | 1.24 (0.40) | 442.88 (168.65) | 0.017 | 0.58 |
| Dw | 1.28 (0.93) | 93.19 (16.65) | 0.239 | 0.32 | 2.48 (0.29) | 71.43 (6.30) | <0.001 | 0.91 | 0.004 (0.005) | 9.33(1.75) | 0.432 | 0.16 | 0.001 (0.001) | 5.66 (0.59) | 0.434 | 0.09 |
| Dn | 2.61 (0.75) | 35.31 (13.51) | 0.025 | 0.75 | 0.06 (0.30) | 34.99 (6.53) | 0.852 | 0.01 | −0.007 (0.017) | 17.90 (5.91) | 0.705 | 0.04 | −0.009 (0.007) | 19.98 (2.86) | 0.213 | 0.21 |
| Dtot | 3.89 (0.61) | 128.50 (10.88) | 0.003 | 0.91 | 2.53 (0.33) | 106.42 (7.29) | <0.001 | 0.89 | −0.003 (0.016) | 27.22 (5.67) | 0.885 | 0.01 | −0.008 (0.008) | 25.64 (3.27) | 0.329 | 0.14 |
| DE | 0.21 (0.08) | 11.73 (1.52) | 0.068 | 0.61 | −0.11 (0.05) | 15.20 (1.09) | 0.054 | 0.43 | −0.005 (0.006) | 6.21(2.08) | 0.436 | 0.16 | −0.007 (0.003) | 6.29 (1.20) | 0.042 | 0.47 |
| Alitta succinea | Ruditapes philippinarum | |||
|---|---|---|---|---|
| F | p | F | p | |
| O2 | 6.559 | 0.027 | 2.486 | 0.143 |
| DIC | 16.723 | 0.002 | 0.029 | 0.869 |
| RQ | 10.439 | 0.008 | 7.809 | 0.017 |
| NH4+ | 20.424 | 0.001 | 0.662 | 0.433 |
| NO2− | 7.909 | 0.017 | 4.115 | 0.067 |
| NO3− | 0.001 | 0.976 | 0.057 | 0.816 |
| DIN | 5.550 | 0.038 | 0.371 | 0.555 |
| Dw | 2.149 | 0.171 | 0.586 | 0.459 |
| Dn | 11.740 | 0.006 | 0.121 | 0.735 |
| Dtot | 3.409 | 0.092 | 0.118 | 0.737 |
| DE | 9.041 | 0.012 | 0.089 | 0.771 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartoli, M.; Benelli, S.; Magri, M.; Ribaudo, C.; Moraes, P.C.; Castaldelli, G. Contrasting Effects of Bioturbation Studied in Intact and Reconstructed Estuarine Sediments. Water 2020, 12, 3125. https://doi.org/10.3390/w12113125
Bartoli M, Benelli S, Magri M, Ribaudo C, Moraes PC, Castaldelli G. Contrasting Effects of Bioturbation Studied in Intact and Reconstructed Estuarine Sediments. Water. 2020; 12(11):3125. https://doi.org/10.3390/w12113125
Chicago/Turabian StyleBartoli, Marco, Sara Benelli, Monia Magri, Cristina Ribaudo, Paula Carpintero Moraes, and Giuseppe Castaldelli. 2020. "Contrasting Effects of Bioturbation Studied in Intact and Reconstructed Estuarine Sediments" Water 12, no. 11: 3125. https://doi.org/10.3390/w12113125
APA StyleBartoli, M., Benelli, S., Magri, M., Ribaudo, C., Moraes, P. C., & Castaldelli, G. (2020). Contrasting Effects of Bioturbation Studied in Intact and Reconstructed Estuarine Sediments. Water, 12(11), 3125. https://doi.org/10.3390/w12113125