How does Urban Pollution Influence Macroinvertebrate Traits in Forested Riverine Systems?




Abstract
1. Introduction
2. Materials and Methods
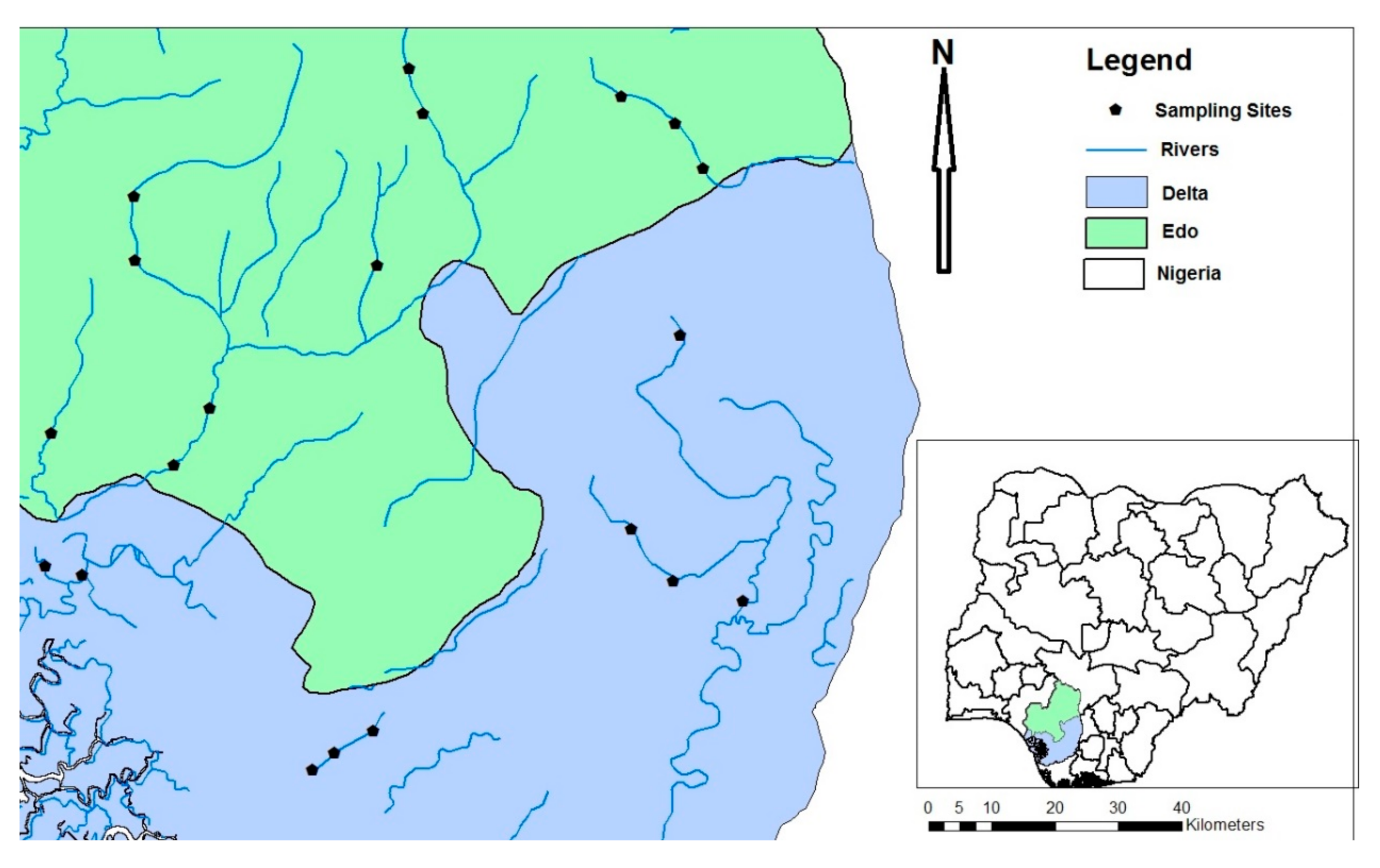
2.1. Study Sites
2.2. Sampling and Analysing of Macroinvertebrates and Physico-Chemical Variables
2.3. Data/Statistical Analyses
Physico-Chemically-Based Classification of Sites into Ecological Classes
2.4. Trait Selection for this Study
2.5. Assessing Macroinvertebrate Traits and Taxa Responses, as Well as Identifying and Classifying Potential Indicator Macroinvertebrate Traits along Urban Pollution Gradient in Forested Riverine Systems
2.6. Exploring the Distribution Patterns of Macroinvertebrate Traits along Urban Pollution Gradient in Forested Riverine Systems
3. Results
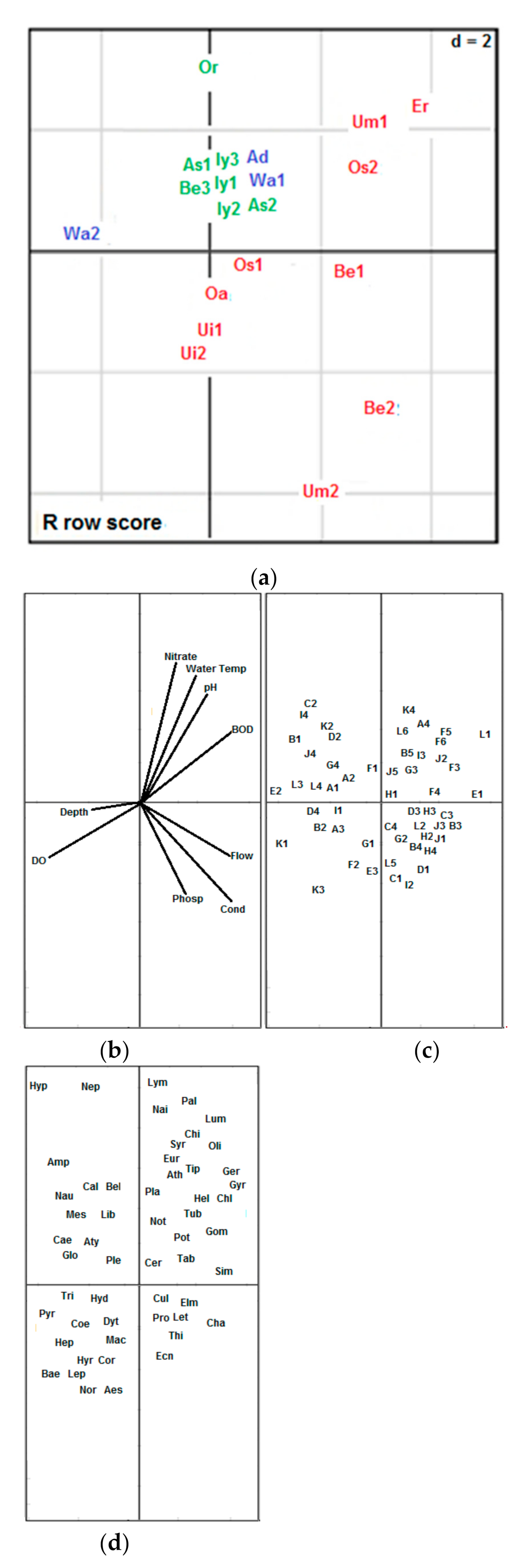
3.1. Assessing Macroinvertebrate Traits and Taxa Responses, as Well as Identifying and Classifying Potential Indicator Macroinvertebrate Traits along Urban Pollution Gradient in Forested Riverine Systems
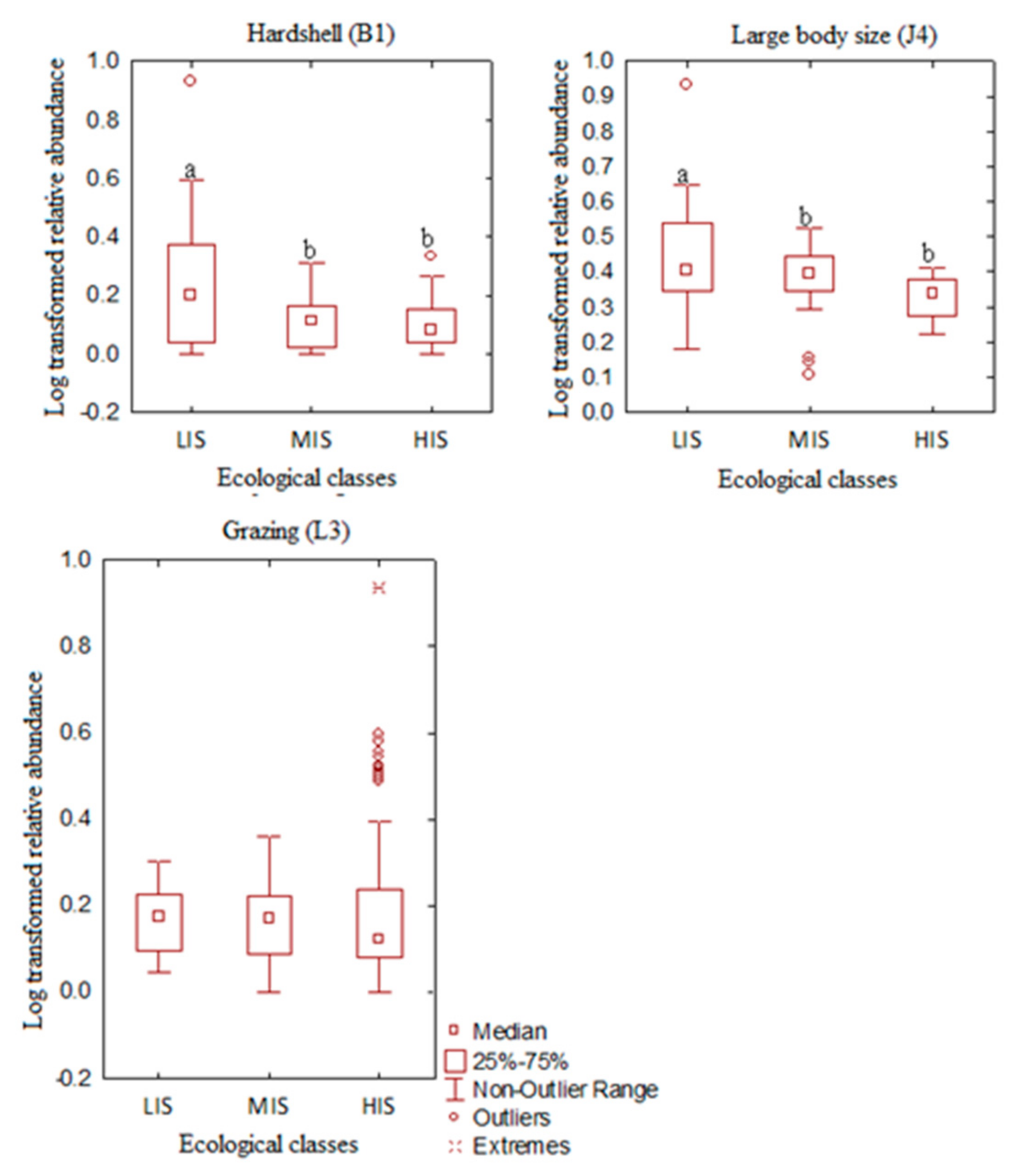
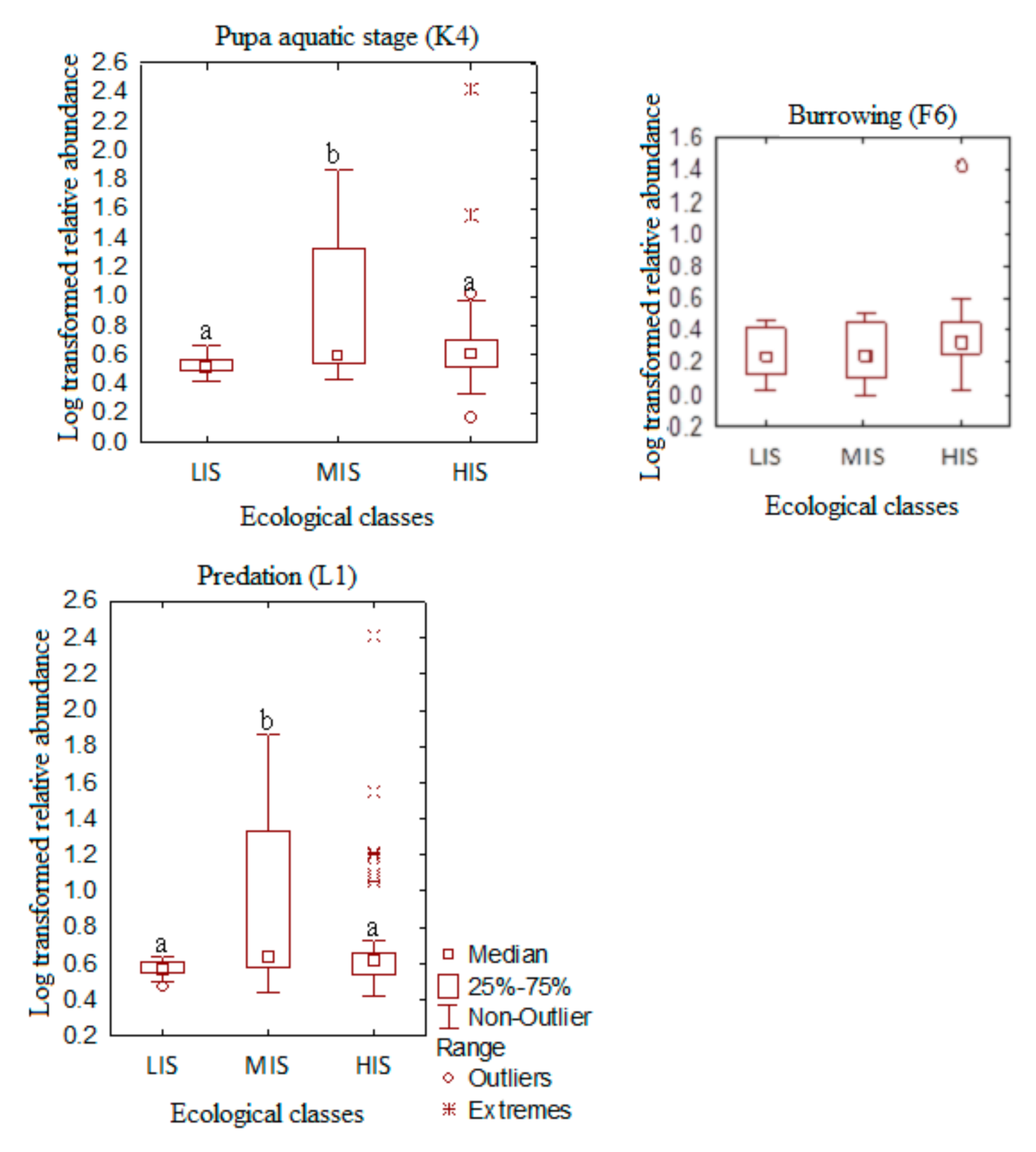
3.2. Exploring the Distribution Patterns of Macroinvertebrate Traits along Urban Pollution Gradient in Forested Riverine Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berger, E.; Haase, P.; Schafer, R.B.; Sundermann, A. Towards stressor-specific macroinvertebrate indices: Which traits and taxonomic groups are associated with vulnerable and tolerant taxa? Sci. Total Environ. 2018, 619, 144–154. [Google Scholar] [CrossRef]
- Desrosiers, M.; Usseglio-Polatera, P.; Archaimbault, V.; Larras, F.; Methot, G.; Pinel-Alloul, B. Assessing anthropogenic pressure in the St. Lawrence River using traits of benthic macroinvertebrates. Sci. Total Environ. 2019, 649, 233–246. [Google Scholar] [CrossRef]
- Olden, J.D.; Poff, N.L.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef]
- Edegbene, A.O.; Arimoro, F.O.; Odume, O.N. Developing and applying a macroinvertebrate-based multimetric index for urban rivers in the Niger Delta, Nigeria. Ecol. Evol. 2019, 9, 12869–12885. [Google Scholar] [CrossRef]
- Odume, O.N. Searching for urban pollution signature and sensitive macroinvertebrate traits and ecological preferences in a river in the Eastern Cape of South Africa. Ecol. Indic. 2020, 108. [Google Scholar] [CrossRef]
- Arimoro, F.O.; Odume, O.N.; Uhunoma, S.I.; Edegbene, A.O. Anthropogenic impact on water chemistry and benthic macroinvertebrate associated changes in a southern Nigeria stream. Environ. Monit. Assess. 2015, 187, 1–14. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Edegbene, A.O. Developing Macroinvertebrate Trait-and Taxonomically-Based Approaches for Biomonitoring Wadeable Riverine Systems in the Niger Delta, Nigeria. Ph.D. Thesis, Rhodes University, Grahamstown, South Africa, 2020. [Google Scholar] [CrossRef]
- Hulot, F.D.; Lacroix, G.; Lescher-Moutoué, F.; Loreau, M. Functional diversity governs ecosystem response to nutrient enrichment. Nature 2000, 405, 340–344. [Google Scholar] [CrossRef]
- Ishikawa, N.F.; Chikaraishi, Y.; Ohkouchi, N.; Murakami, A.R.; Tayasu, I.; Togashi, H.; Okuda, N. Integrated trophic position decreases in more diverse communities of stream food webs. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Gieswein, A.; Hering, D.; Lorens, A.W. Development and validation of a macroinvertebrate-based biomonitoring tool to assess fine sediment impact in small mountain streams. Sci. Total Environ. 2019, 652, 1290–1301. [Google Scholar] [CrossRef]
- Lakew, A.; Moog, O. A multimetric index based on benthic macroinvretebrates for assessing the ecological status of streams and rivers in central and southeast highlands of Ethiopia. Hydrobiologia 2015, 751, 229–242. [Google Scholar] [CrossRef]
- Kuzmanovic, M.; Doledec, S.; de Catro-Catala, N.; Ginebreda, A.; Sabater, S.; Munoz, I.; Barcelo, D. Environmental stressors as driver of the trait composition of benthic macroinvertebrates assemblages in polluted Iberian rivers. Environ. Res. 2017, 156, 485–493. [Google Scholar] [CrossRef]
- Walsh, C.J.; Jack, W.F.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The urban syndrome: Current knowledge and the search for a cure. J. North Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Edegbene, A.O.; Elakhame, L.A.; Arimoro, F.O.; Osimen, E.C.; Odume, O.N. Development of Macroinvertebrates Multimetric Index for Ecological Evaluation of a River in North Central Nigeria. Environ. Monit. Assess. 2019, 191, 274. [Google Scholar] [CrossRef]
- Akamagwuna, F.C.; Mensah, P.K.; Nnadozie, C.F.; Odume, O.N. Traits-based responses of Ephemeroptera, Plecoptera and Trichoptera to sediment stress in the Tsitsa River and its tributaries, Eastern Cape, South Africa. River Res. Appl. 2019, 35, 999–1012. [Google Scholar] [CrossRef]
- Edegbene, A.O.; Arimoro, F.O.; Odume, O.N. Exploring the distribution patterns of macroinvertebrate signature traits and ecological preferences and their responses to urban and agricultural pollution in selected rivers in the Niger Delta ecoregion, Nigeria. Aquat. Ecol. 2020, 54, 553–573. [Google Scholar] [CrossRef]
- Umoh, S.G. The promise of wetland farming; evidence from Nigeria. Agric. J. 2008, 3, 107–112. [Google Scholar]
- Zabbey, N.; Erondu, E.S.; Hart, A.I. Nigeria and the prospect of shrimp farming: Critical issues. Livest. Resour. Rural Dev. 2010, 22. Available online: http://www.lrrd.org/lrrd22/11/zabb22198.htm (accessed on 30 September 2020).
- Pena-Cortes, F.; Pincheira-Ulbrich, J.; Bertran, C.; Tapia, J.; Hauenstein,, E.; Fernandez, E.; Rozas, D. A study of the geographic distribution of swamp forest in the coastal zone of the Araucania Region, Chile. Appl. Geogr. 2011, 31, 545–555. [Google Scholar] [CrossRef]
- Fierro, P.; Bertran, C.; Hauenstein, E.; Pena-Cortes, F.; Vergara, C.; Cerna, C.; Vargas-Chaoff, I. Effects of local land-use on riparian vegetation, water quality and the functional organisation of macroinvertebrate assemblages. Sci. Total Environ. 2017, 609, 724–734. [Google Scholar] [CrossRef]
- Lazorchak, J.M.; Klemm, D.J.; Peck, D.V. Environmental Monitoring and Assessment Program Surface Waters: Field Operations and Methods Manual for Measuring the Ecological Condition of Wadeable Streams; EPA 620/R-94/004F; Environmental Protection Agency: Washington, DC, USA, 1998. [Google Scholar]
- Merritt, R.W.; Cummins, K.W. An Introduction to the Aquatic Insects of North America; Kendall Hunt: St. Louis, MO, USA, 1996. [Google Scholar]
- Day, J.A.; Harrison, A.D.; De Moor, I.J. Guides to the Freshwater Fnvertebrates of Southern Africa: Diptera; Water Research Commission: Pretoria, South Africa, 2003. [Google Scholar]
- de Moor, I.J.; Day, J.A.; de Moor, F.C. Guides to the Freshwater Invertebrates of Southern Africa: Insecta I. Ephemeroptera, Odonata and Plecoptera; Water Research Commission: Pretoria, South Africa, 2003; Volume 7. [Google Scholar]
- Gordon, N.D.; McMahon, T.A.; Finlayson, B.L. Stream Hydrology, an Introduction for Ecologists; John Wiley & Sons Ltd.: New York, NY, USA, 1994; p. 526. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Murphy, J.F.; Davy-Bowker, J.; McFarland, B.; Ormerod, S.J. A diagnostic biotic index for assessing acidity in sensitive streams in Britain. Ecol. Indic. 2013, 24, 562–572. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Steven, M.H.M.; Szoecs, E.; Wagner, H. Vegan: Community Ecol. Package. 2019. Available online: https://cran.r-project.org (accessed on 11 February 2019).
- R Development Core Team. R Statistical Computing Environment; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISBN 3–900051–07-0. [Google Scholar]
- Lee, J.H.; Bang, K.W. Characterization of urban stormwater runoff. Water Res. 2000, 34, 1773–1780. [Google Scholar] [CrossRef]
- Hatt, B.E.; Flecther, T.D.; Walsh, C.J.; Taylor, S.L. The influence of urban density and drainage infrastructure on the concentrations and loads of pollutants in small streams. Environ. Manag. 2004, 34, 112–124. [Google Scholar] [CrossRef]
- Roy, A.H.; Freeman, M.C.; Freeman, B.J.; Wenger, S.J.; Ensign, W.E.; Meyer, J.L. Investigating hydrological alteration as a mechanism of fish assemblage shifts in urbanising streams. J. North Am. Benthol. Soc. 2005, 24, 656–678. [Google Scholar] [CrossRef]
- Dolédec, S.; Statzner, B. Invertebrate traits for the biomonitoring of large European rivers: An assessment of specific types of human impact. Freshw. Biol. 2008, 53, 617–634. [Google Scholar] [CrossRef]
- Ogren, S.A.; King, D.K. The effect of large woody debris on macroinvertebrate communities and epilithon detritus composition in a channelized headwater stream. J. Freshw. Ecol. 2008, 23, 65–77. [Google Scholar] [CrossRef]
- Jones, J.I.; Murphy, J.F.; Collins, A.L.; Sear, D.A.; Naden, P.S.; Armitage, P.D. The impact of fine sediment on macroinvertebrates. River Res. Appl. 2012, 28, 1055–1071. [Google Scholar] [CrossRef]
- Heino, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 2013, 88, 166–178. [Google Scholar] [CrossRef]
- Krynak, E.M.; Yates, A.G. Benthic invertebrates taxonomic and trait associations with land use intensively managed watershed: Implications for indicator identification. Ecol. Indic. 2018, 93, 1050–1059. [Google Scholar] [CrossRef]
- Bonada, N.; Doledec, S. Do Mediterranean genera not included in Tachet et al., 2002 have Mediterranean trait characteristics? Limnetica 2011, 30, 129–142. [Google Scholar]
- Chevenet, F.; Dolédec, S.; Chessel, D. A fuzzy coding approach for analysis of longterm ecological data. Freshw. Biol. 1994, 31, 295–309. [Google Scholar] [CrossRef]
- Dolédec, S.; Chessel, D.; ter Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Dray, S.; Choler, P.; Doledec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Milosevic, D.; Stojanovic, K.; Djurdjevic, A.; Markovic, Z.; Piperac, M.S.; Zivic, M.; Zivic, I. The response of chironomid taxonomy-and functional trait-based metric to fish effluent pollution in lotic systems. Environ. Pollut. 2018, 242, 1058–1066. [Google Scholar] [CrossRef]
- Statzner, B.; Beche, L. Can biological invertebrate traits resolve effects of multiple stressors on running water ecosystems? Freshw. Biol. 2010, 55, 80–199. [Google Scholar] [CrossRef]
- Townsend, C.R.; Hildrew, A.G. Species traits in relation to a habitat templet for river systems. Freshw. Biol. 1994, 31, 265–275. [Google Scholar] [CrossRef]
- Serra, R.Q.S.; Graca, M.A.S.; Doledec, S.; Feio, M.J. Chironomidae traits and life history strategies as indicators of anthropogenic disturbance. Environ. Monit. Assess. 2017, 189. [Google Scholar] [CrossRef]
- Castro, D.M.P.; Dolédec, S.; Callisto, M. Land cover disturbance homogenizes aquatic insect functional structure in neotropical savanna streams. Ecol. Indic. 2018, 84, 573–582. [Google Scholar] [CrossRef]
- Ramirez, A. Odonata. Rev. De Biol. Trop. 2010, 58, 97–136. [Google Scholar]
- Pilière, A.F.H.; Verberk, W.C.E.P.; Gräwe, M.; Breure, A.M.; Dyer, S.D.; Posthuma, L.; de Zwart, D.; Huijbregts, M.A.J.; Schipper, A.M. On the importance of trait interrelationships for understanding environmental responses of stream macroinvertebrates. Freshw. Biol. 2016, 61, 181–194. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Konddratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. North Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef]
- Ding, N.; Yang, W.; Zhou, Y.; Gonzalez-Bergonzoni, I.; Zhang, J.; Chen, K.; Vidal, N.; Jeppesen, E.; Liu, Z.; Wang, B. Different responses of functional traits and diversity of stream macroinvertebrates to environmental and spatial factors in the Xishuangbanna watershed of the upper Mekong River Basin, China. Sci. Total Environ. 2017, 574, 288–299. [Google Scholar] [CrossRef]
- Tomanova, S.; Moya, N.; Oberdorff, T. Using macroinvertebrate biological traits for assessing biotic integrity of neotropical streams. River Res. Appl. 2008, 24, 1230–1239. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Codes | Coordinate Values of Sampled Sites on the PCA Axis 1 | Inter-Site Distances | % Inter-Site Distances | Site Ecological Classes |
|---|---|---|---|---|
| Wa2 | −28.3 | 189 | 100 | LIS |
| Wa1 | −28 | 188.6 | 99.8 | LIS |
| Ad | −26.1 | 186.7 | 98.8 | LIS |
| Or | −24.2 | 184.8 | 97.8 | MIS |
| As2 | −22.5 | 183.1 | 96.9 | MIS |
| Iy3 | −22.4 | 183.0 | 96.9 | MIS |
| Iy1 | −21.2 | 181.8 | 96.2 | MIS |
| As1 | −20.7 | 181.4 | 96.0 | MIS |
| Iy2 | −20.3 | 181 | 95.8 | MIS |
| Be3 | −17.1 | 177.7 | 94.0 | MIS |
| Os2 | −14.8 | 173.5 | 91.8 | HIS |
| Be1 | −12.9 | 173.5 | 91.8 | HIS |
| Os1 | −12.2 | 172.9 | 91.5 | HIS |
| Oa | −8.6 | 169.3 | 89.6 | HIS |
| Um2 | −2.3 | 163 | 86.2 | HIS |
| Er | 19.0 | 142 | 75.0 | HIS |
| Ui2 | 24.7 | 136 | 71.9 | HIS |
| Um1 | 32.9 | 127.8 | 67.6 | HIS |
| Ui1 | 44.1 | 116.6 | 61.7 | HIS |
| Be2 | 160.7 | 0.0 | 0.0 | HIS |
| Traits | Physico-Chemical Variables | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Water Temp | Depth | Flow | Cond | DO | BOD5 | pH | Nitrate | Phosp | |
| Gills | |||||||||
| Tegument/cutaneous | + | ||||||||
| Aerial: spiracle | |||||||||
| Aerial/vegetation: breathing tube, strap/other apparatus | |||||||||
| Hardshell | - | + | - | ||||||
| Completely sclerotized | |||||||||
| Partly sclerotized | |||||||||
| Soft and exposed | |||||||||
| Cased/tubed | - | ||||||||
| Clear and transparent waters | |||||||||
| Silty waters | - | + | |||||||
| Turbid waters | |||||||||
| No preference for turbid waters | |||||||||
| 1 year (Univoltine) | + | ||||||||
| 2 years (Bivoltine) | + | - | + | ||||||
| D3 = >2 years (Multivoltine) | |||||||||
| D4 = longer than one year (Semivoltine) | |||||||||
| Free-living | |||||||||
| Temporary attachment | |||||||||
| Permanent attachment | |||||||||
| Climbing | |||||||||
| Crawling | + | ||||||||
| Sprawling | + | + | |||||||
| Swimming | - | + | - | - | |||||
| Skating | |||||||||
| Burrowing | + | + | - | ||||||
| Streamlined | - | ||||||||
| Flattened | - | ||||||||
| Spherical | |||||||||
| Cylindrical/tubular | |||||||||
| Detritus (FPOM) | |||||||||
| Detritus (CPOM) | |||||||||
| Macrophytes/algae | |||||||||
| Animal materials | |||||||||
| Highly sensitive to oxygen depletion | |||||||||
| Moderately sensitive to oxygen depletion | - | - | |||||||
| Moderately tolerant of oxygen depletion | |||||||||
| Highly tolerant of oxygen depletion | + | - | + | ||||||
| Very small body size (<5 mm) | |||||||||
| Small body size (>5–10 mm) | |||||||||
| Medium body size (>10–20 mm | |||||||||
| Large body size (>20–40 mm) | + | ||||||||
| Very large body size (>40–80 mm) | |||||||||
| Egg aquatic stage | |||||||||
| Larva aquatic stage | |||||||||
| Nymph aquatic stage | |||||||||
| Pupa aquatic stage | - | + | |||||||
| Predation | - | ||||||||
| Scraping | |||||||||
| Grazing | - | - | |||||||
| Filter feeding | |||||||||
| Deposit feeding | |||||||||
| Shredding | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edegbene, A.O.; Arimoro, F.O.; Odume, O.N. How does Urban Pollution Influence Macroinvertebrate Traits in Forested Riverine Systems? Water 2020, 12, 3111. https://doi.org/10.3390/w12113111
Edegbene AO, Arimoro FO, Odume ON. How does Urban Pollution Influence Macroinvertebrate Traits in Forested Riverine Systems? Water. 2020; 12(11):3111. https://doi.org/10.3390/w12113111
Chicago/Turabian StyleEdegbene, Augustine O., Francis O. Arimoro, and Oghenekaro N. Odume. 2020. "How does Urban Pollution Influence Macroinvertebrate Traits in Forested Riverine Systems?" Water 12, no. 11: 3111. https://doi.org/10.3390/w12113111
APA StyleEdegbene, A. O., Arimoro, F. O., & Odume, O. N. (2020). How does Urban Pollution Influence Macroinvertebrate Traits in Forested Riverine Systems? Water, 12(11), 3111. https://doi.org/10.3390/w12113111