Temporal and Spatial Variations of the Biochemical Composition of Phytoplankton and Potential Food Material (FM) in Jaran Bay, South Korea

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
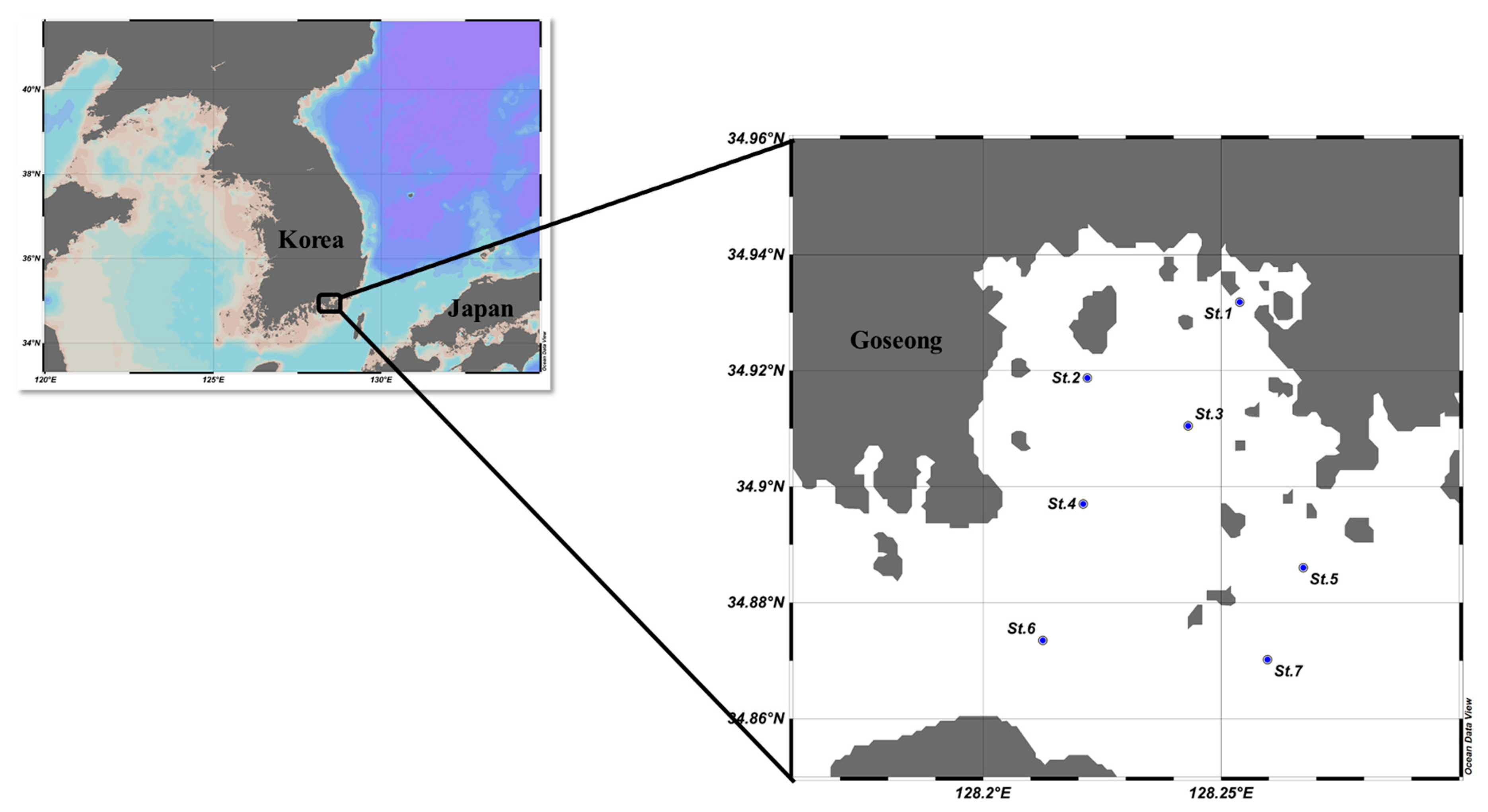
2.1. Water Sampling and Analysis
2.2. Statistical Analysis
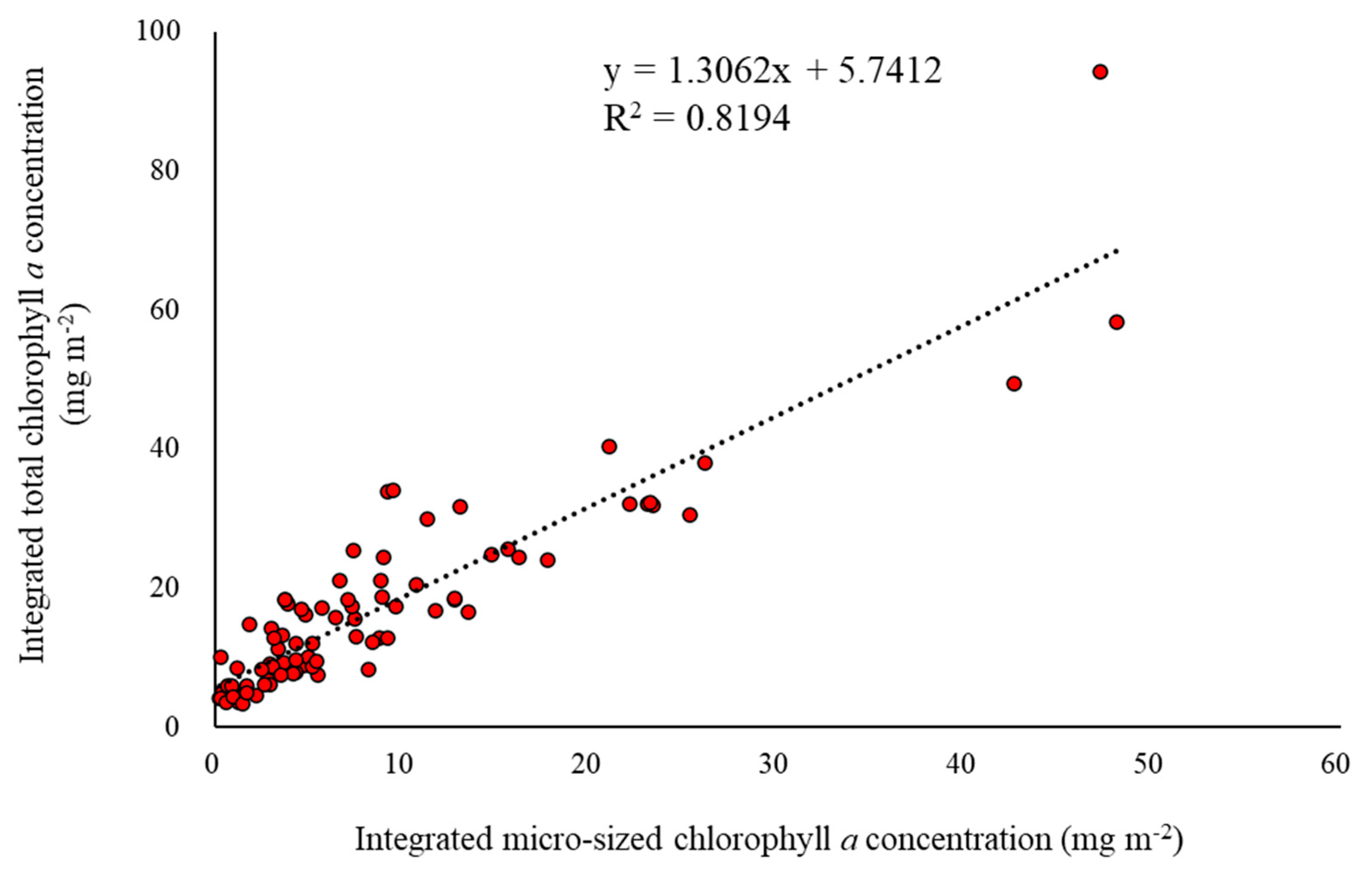
3. Results
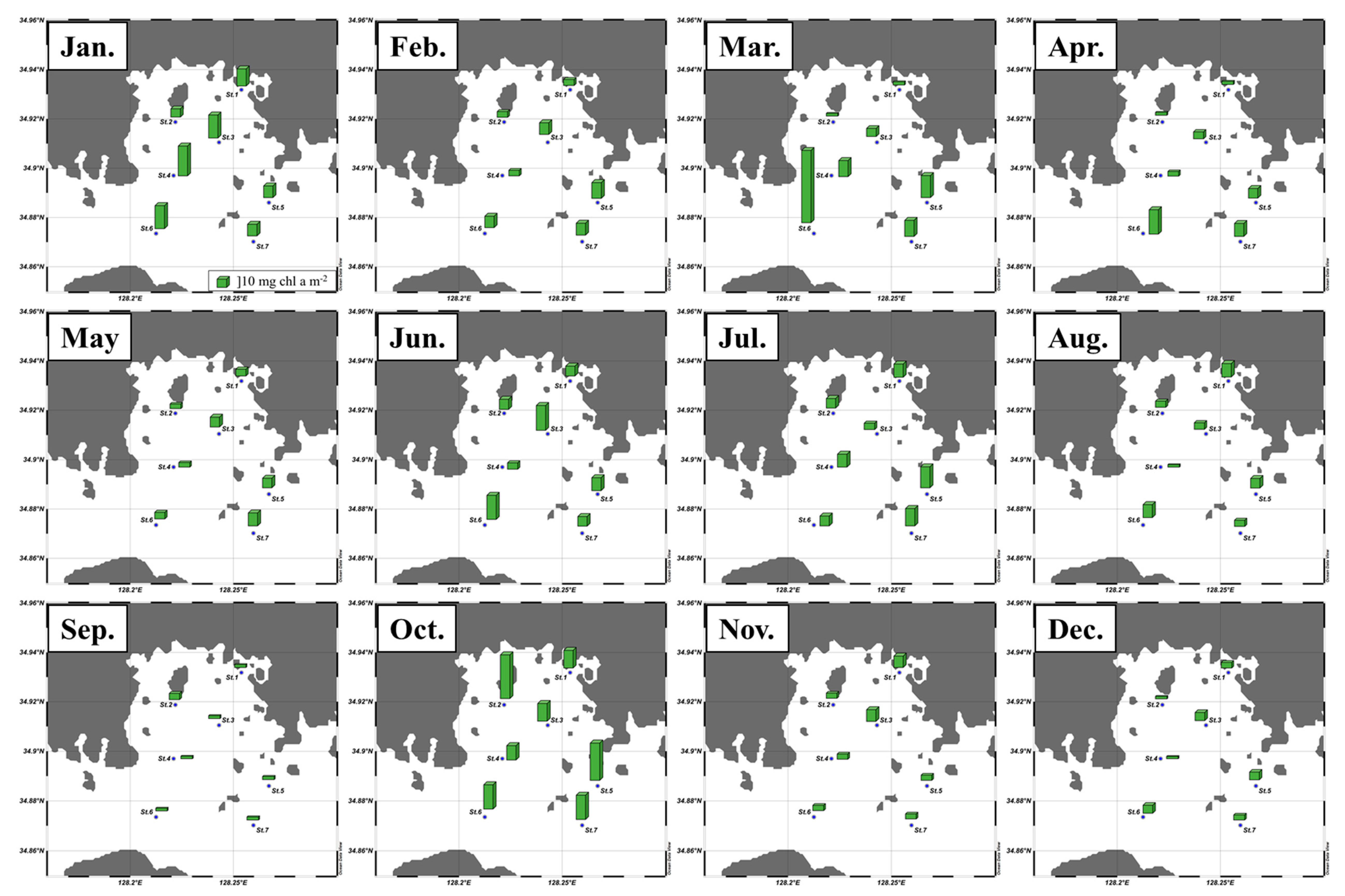
3.1. Monthly Concentrations of Nutrients and Chlorophyll a
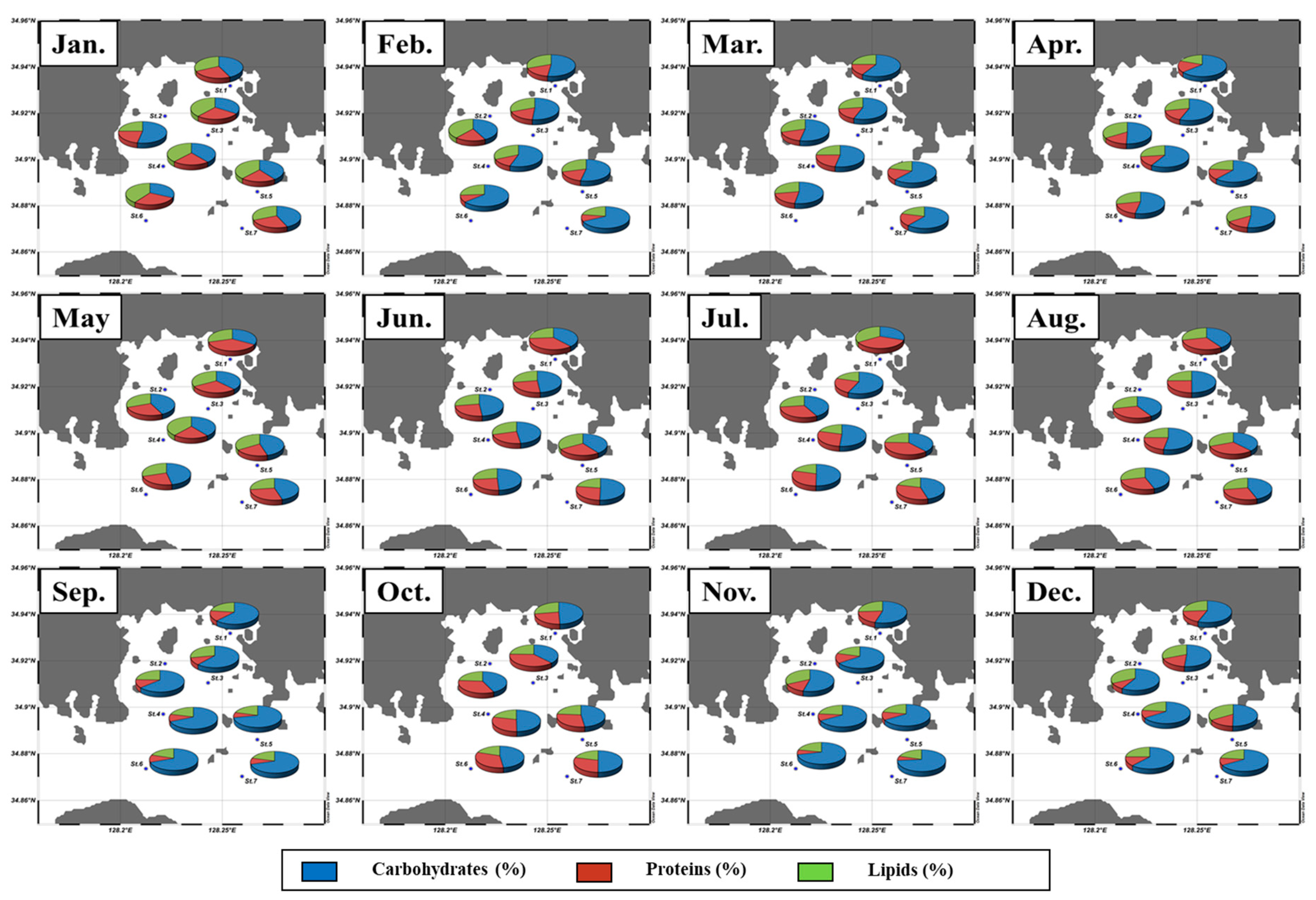
3.2. Spatial and Temporal Variations of the Macromolecular Compositions of POM
3.3. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mathiesen, A.M. The state of the World Fisheries and Aquaculture 2012; Food and Agriculture Organization: Rome, Italy, 2012; p. 290. [Google Scholar]
- Cho, C.H.; Park, K. Eutrophication of bottom mud in shellfish farms, the Goseong-Jaran Bay. Korean J. Fish. Aquat. Sci. 1983, 16, 260–264. [Google Scholar]
- Xu, Q.; Yang, H. Food sources of three bivalves living in two habitats of Jiaozhou Bay (Qingdao, China): Indicated by lipid biomarkers and stable isotope analysis. J. Shellfish Res. 2007, 26, 561–567. [Google Scholar] [CrossRef]
- Umehara, A.; Asaoka, S.; Fujii, N.; Otani, S.; Yamamoto, H.; Nakai, S.; Okuda, T.; Nishijima, W. Biological productivity evaluation at lower trophic levels with intensive Pacific oyster farming of Grassostrea gigas in Hiroshima Bay, Japan. Aquaculture 2018, 495, 311–319. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, D.; Kang, J.J.; Joo, H.; Lee, J.H.; Lee, H.W.; Ahn, S.H.; Kang, C.K.; Lee, S.H. The effects of different environmental factors on the biochemical composition of particulate organic matter in Gwanyang Bay, South Korea. Biogeosciences 2017, 14, 1903–1917. [Google Scholar] [CrossRef]
- Kim, H.C.; Lee, J.H.; Lee, W.C.; Hong, S.; Kang, J.J.; Lee, D.; Jo, N.; Bhavya, P.S. Decoupling of macromolecular compositions of particulate organic matters between the water columns and the sediment in Geoje-Hansan Bay, South Korea. Ocean Sci. J. 2018, 53, 735–743. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, J.H.; Kang, J.J.; Lee, J.H.; Lee, H.W.; Kang, C.K.; Lee, S.H. River discharge effects on the contribution of small-sized phytoplankton to the total biochemical composition of POM in Gwangyang Bay, Korea. Estuar. Coast. Shelf Sci. 2019, 226, 106293. [Google Scholar] [CrossRef]
- Morris, I.; Glover, H.E.; Yentsch, C.S. Products of photosynthesis by marine phytoplankton: The effect of environmental factors on the relative rates of protein synthesis. Mar. Biol. 1974, 21, 1–9. [Google Scholar] [CrossRef]
- Kowallik, W. Blue light effects on carbohydrate and protein metabolism. In Blue Light Responses: Phenomena and Occurrence in Plants; Senger, H., Ed.; CRC Press: Boca Raton, FL, USA, 1978; Volume 1, pp. 8–13. [Google Scholar]
- Suárez, I.; Maranón, E. Photosynthate allocation in a temperate sea over an annual cycle: The relationship between protein synthesis and phytoplankton physiological state. J. Sea Res. 2003, 50, 285–299. [Google Scholar] [CrossRef]
- Mortensen, S.H.; Børsheim, K.Y.; Rainuzzo, J.; Knutsen, G. Fatty acid and elemental composition of the marine diatom Chaetoceros gracilis Schütt. Effects of silicate deprivation, temperature and light intensity. J. Exp. Mar. Biol. Ecol. 1988, 122, 173–185. [Google Scholar] [CrossRef]
- Liebezeit, G. Particulate carbohydrate in relation to phytoplankton in the euphotic zone of the Bransfield Strait. Polar Biol. 1984, 2, 225–228. [Google Scholar] [CrossRef]
- Moal, J.; Jezequel, V.M.; Harris, R.P.; Samain, J.F.; Poulet, S.A. Interspecific and intraspecific variability of the chemical-composition of marine-phytoplankton. Oceanol. Acta 1987, 10, 339–346. [Google Scholar]
- Kilham, S.S.; Kreeger, D.A.; Goulden, C.E.; Lynn, S.G. Effects of nutrient limitation on biochemical constituents of Ankistrodesmus falcatus. Freshw. Biol. 1997, 38, 591–596. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, H.J.; Whitledge, T.E. High incorporation of carbon into proteins by the phytoplankton of the Bering Strait and Chukchi Sea. Cont. Shelf Res. 2009, 29, 1689–1696. [Google Scholar] [CrossRef]
- Yun, M.S.; Lee, D.B.; Kim, B.K.; Kang, J.J.; Lee, J.H.; Yang, E.J.; Park, W.G.; Chung, K.H.; Lee, S.H. Comparison of phytoplankton macromolecular compositions and zooplankton proximate compositions in the northern Chukchi Sea. Deep Sea Res. Part II 2015, 120, 82–90. [Google Scholar] [CrossRef]
- Jo, N.; Kang, J.J.; Park, W.G.; Lee, B.R.; Yun, M.S.; Lee, J.H.; Kim, S.M.; Lee, D.; Joo, H.; Lee, J.H.; et al. Seasonal variation in the biochemical compositions of phytoplankton and zooplankton communities in the southwestern East/Japan Sea. Deep Sea Res. Part II 2017, 143, 82–90. [Google Scholar] [CrossRef]
- Navarro, J.M.; Clasing, E.; Urrutia, G.; Asencio, G.; Stead, R.; Herrera, C. Biochemical composition and nutritive value of suspended particulate matter over a tidal flat of Southern Chile. Estuar. Coast. Shelf Sci. 1993, 37, 59–73. [Google Scholar] [CrossRef]
- Danovaro, R.; Dell’Anno, A.; Pusceddu, A.; Marrale, D.; Croce, N.D.; Fabiano, M.; Tselepides, A. Biochemical composition of pico-, nano- and microparticulate organic matter and bacterioplankton biomass in the oligotrophic Cretan Sea (NE Mediterranean). Prog. Oceanogr. 2000, 46, 279–310. [Google Scholar] [CrossRef]
- Navarro, J.M.; Thompson, R.J. Seasonal fluctuations in the size spectra, biochemical-composition and nutritive-value of the seston available to a suspension-feeding bivalve ina asub-arctic environment. Mar. Ecol. Prog. Ser. 1995, 125, 95–106. [Google Scholar] [CrossRef]
- Kang, J.J.; Joo, H.; Lee, J.H.; Lee, J.H.; Lee, H.W.; Lee, D.; Kang, C.K.; Yun, M.S.; Lee, S.H. Comparison of biochemical compositions of phytoplankton during spring and fall seasons in the northern East/Japan Sea. Deep Sea Res. Part II 2017, 143, 73–81. [Google Scholar] [CrossRef]
- Lee, S.H.; Whitledge, T.E.; Kang, S. Recent carbon and nitrogen uptake rates of phytoplankton in Bering Strait and the Chukchi Sea. Cont. Shelf Res. 2007, 27, 2231–2249. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, H.C.; Lee, T.; Lee, W.C.; Kang, J.J.; Jo, N.; Lee, D.; Kim, K.; Min, J.; Kang, S.; et al. Monthly variations in the intracellular nutrient pools of phytoplankton in Jaran Bay, Korea. J. Coast. Res. 2018, 85, 331–335. [Google Scholar] [CrossRef]
- Welschmeyer, N.A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and phaeopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colormetric method for determination of sugars and related substances. Anal. Chem. 1994, 28, 350–356. [Google Scholar] [CrossRef]
- Stevens, J. Applied Multivariate Statistics for the Social Science; Lawrence Erlbaum: New York, NY, USA, 1986; p. 515. [Google Scholar]
- Camdevyren, H.; Demyr, N.; Kanik, A.; Keskyn, S. Use principal component scores in multiple linear regression models for prediction of Chlorophyll-a in reservoirs. Ecol. Model. 2005, 181, 581–589. [Google Scholar] [CrossRef]
- Wang, Y.S.; Lou, Z.P.; Sun, C.C.; Wu, M.L.; Han, S.H. Multivariate statistical analysis of water quality and phytoplankton characteristics in Daya Bay, China, from 1999 to 2002. Oceanologia 2006, 48, 193–211. [Google Scholar]
- Deutsch, C.; Sarmiento, J.L.; Sigman, D.M.; Gruber, N.; Dunne, J.P. Spatial coupling of nitrogen inputs and losses in the ocean. Nature 2007, 445, 163–167. [Google Scholar] [CrossRef]
- Brun, P.; Vogt, M.; Payne, M.R.; Gruber, N.; O’Brien, C.J.; Buitenhuis, E.T.; Le Quere, C.; Leblanc, K.; Luo, Y.W. Ecological niches of open ocean phytoplankton taxa. Limnol. Oceanogr. 2015, 60, 1020–1038. [Google Scholar] [CrossRef]
- Pedhazur, E. Multiple regression in behavioral science; Holt Rinehart & Winston: Fort Worth, TX, USA, 1982; p. 135. [Google Scholar]
- Lee, Y.S.; Kang, C.K.; Kwon, K.Y.; Kim, S.Y. Organic and inorganic matter increase related to eutrophication in Gamak Bay, South Korea. J. Environ. Biol. 2009, 30, 373–380. [Google Scholar]
- Lee, Y.S.; Kang, C.K. Causes of COD increases in Gwangyang Bay, South Korea. J. Environ. Monit. 2010, 12, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Lim, W.A.; Jung, C.S.; Park, J. Spatial distributions and monthly variations of water quality in coastal seawater of Tongyeong, Korea. J. Korean Soc. Mar. Environ. Eng. 2011, 14, 154–162. [Google Scholar] [CrossRef]
- Jeong, D.H.; Shin, H.H.; Jung, S.W.; Lim, D.I. Variations and characters of water quality during flood and dry seasons in the Eastern coast of South Sea, Korea. Korean J. Environ. Biol. 2013, 31, 19–36. [Google Scholar] [CrossRef]
- Park, J.S.; Yoon, Y.H.; Oh, S.J. Variational characteristics of phytoplankton community in the mouth parts of Gamak Bay, Southern Korea. Korean J. Environ. Biol. 2009, 27, 205–215. [Google Scholar]
- Lee, J.H.; Kang, J.J.; Jang, H.K.; Jo, N.; Lee, D.; Yun, M.S.; Lee, S.H. Major controlling factors for spatio-temporal variations in the macromolecular composition and primary production by phytoplankton in Garolim and Asan bays in the Yellow Sea. Reg. Stud. Mar. Sci. 2020, 36, 101269. [Google Scholar] [CrossRef]
- Desortová, B. Relationship between chlorophyll-a concentration and phytoplankton biomass in several reservoirs in Czechoslovakia. Hydrobiology 1981, 66, 153–169. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Boss, E.; Siegel, D.A.; Shea, D.M. Carbon-based ocean productivity and phytoplankton physiology from space. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Boss, E. Beam attenuation and chlorophyll concentration as alternative optical indices of phytoplankton biomass. J. Mar. Res. 2006, 64, 431–451. [Google Scholar] [CrossRef]
- Kruskopf, M.; Flynn, K.J. Chlorophyll content and fluorescence responses cannot be used to gauge reliably phytoplankton biomass, nutrient status or growth rate. New Phytol. 2006, 169, 525–536. [Google Scholar] [CrossRef]
- Kim, T.W.; Lee, K.; Lee, C.K.; Jeong, H.D.; Suh, Y.S.; Lim, W.A.; Kim, K.Y.; Jeong, H.J. Interannual nutrient dynamics in Korean coastal waters. Harmful Algae 2013, 30, 15–27. [Google Scholar] [CrossRef]
- Son, Y.B.; Ryu, J.H.; Noh, J.H.; Ju, S.J.; Kim, S.-H. Climatological variability of satellite-derived sea surface temperature and chlorophyll in the South Sea of Korea and East China Sea. Ocean Polar Res. 2012, 34, 201–218. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Integrated Nutrients | ||||
|---|---|---|---|---|
| NH4+ | NO2− + NO3− | DIP | SiO2-Si | |
| mmol m−2 | ||||
| Jan. | 8 ± 5 | 21 ± 19 | 3 ± 2 | 64 ± 46 |
| Feb. | 4 ± 3 | 8 ± 6 | 2 ± 1 | 21 ± 11 |
| Mar. | 4 ± 2 | 7 ± 6 | 1 ± 1 | 22 ± 7 |
| Apr. | 7 ± 2 | 12 ± 5 | 2 ± 1 | 47 ± 5 |
| May | 6 ± 1 | 7 ± 2 | 0.5 ± 0.2 | 74 ± 14 |
| Jun. | 8 ± 3 | 12 ± 8 | 1 ± 1 | 167 ± 48 |
| Jul. | 11 ± 4 | 15 ± 10 | 2 ± 1 | 161 ± 31 |
| Aug. | 9 ± 4 | 8 ± 4 | 2 ± 1 | 90 ± 46 |
| Sep. | 48 ± 19 | 33 ± 13 | 6 ± 2 | 146 ± 37 |
| Oct. | 6 ± 2 | 22 ± 23 | 1 ± 2 | 72 ± 70 |
| Nov. | 11 ± 4 | 44 ± 24 | 4 ± 2 | 102 ± 46 |
| Dec. | 9 ± 4 | 46 ± 31 | 4 ± 2 | 106 ± 66 |
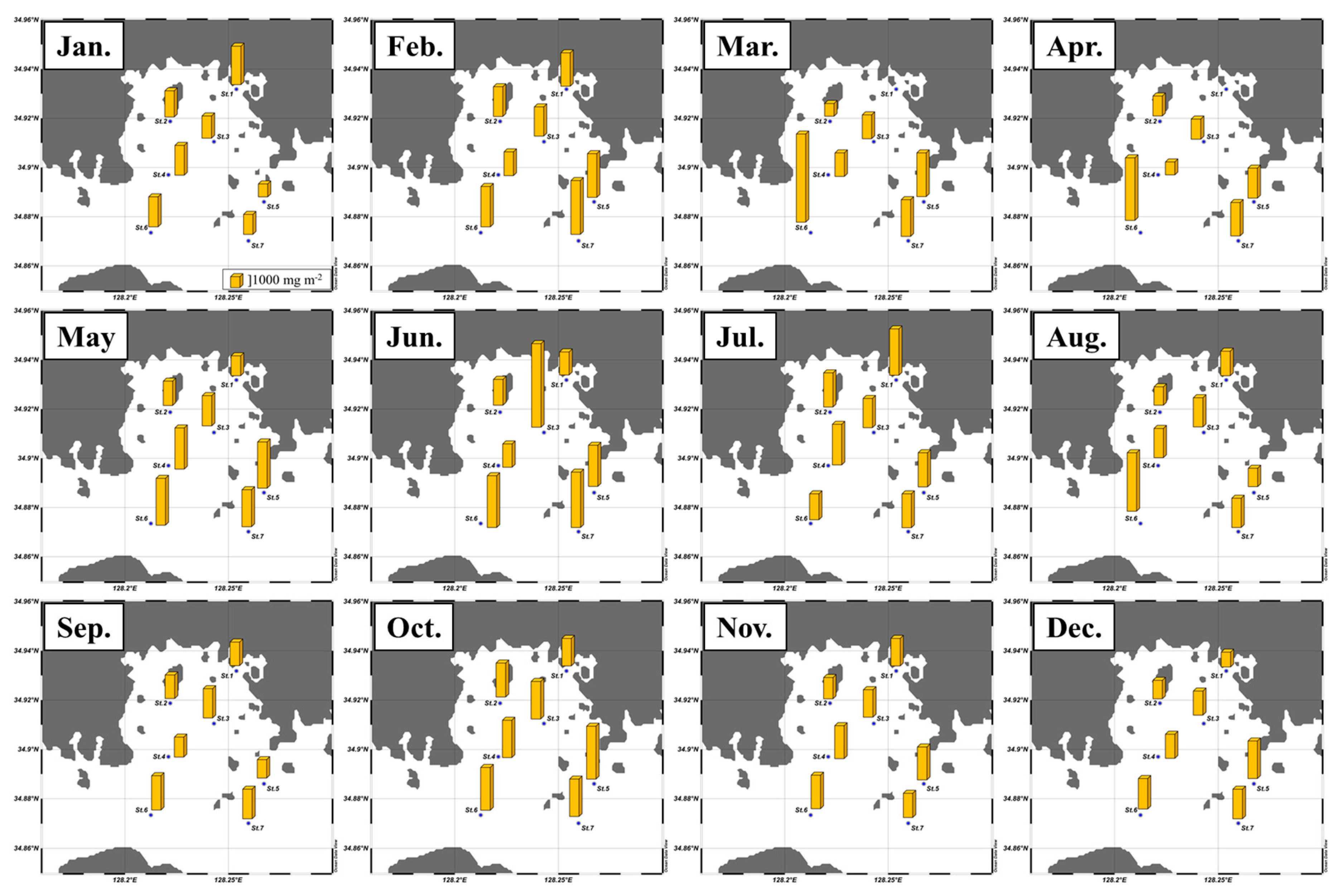
| Total chl a | Micro | Nano | Pico | CHO | PRT | LIP | FM | CHO | PRT | LIP | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (μg L−1) | (%) | (%) | (%) | (μg L−1) | (μg L−1) | (μg L−1) | (μg L−1) | (%) | (%) | (%) | |
| Jan. | 3.2 ± 1.0 | 78 ± 7 | 14 ± 7 | 8 ± 4 | 145 ± 64 | 81 ±19 | 119 ± 34 | 345 ± 84 | 41 ± 9 | 24 ± 4 | 35 ± 8 |
| Feb. | 1.7 ± 0.6 | 72 ± 19 | 15 ± 8 | 13 ± 18 | 243 ± 56 | 65 ± 21 | 134 ± 44 | 442 ± 56 | 55 ± 10 | 15 ± 5 | 30 ± 8 |
| Mar. | 2.5 ± 1.6 | 45 ± 22 | 19 ± 5 | 37 ± 18 | 206 ± 48 | 68 ± 21 | 92 ± 26 | 368 ± 78 | 56 ± 6 | 18 ± 3 | 25 ± 5 |
| Apr. | 1.4 ± 0.7 | 24 ± 19 | 30 ± 6 | 46 ± 18 | 183 ± 37 | 53 ± 17 | 96 ± 31 | 332 ± 57 | 56 ± 8 | 16 ± 3 | 29 ± 7 |
| May | 1.4 ± 0.8 | 32 ± 16 | 38 ± 12 | 29 ± 12 | 163 ± 44 | 103 ± 37 | 129 ± 50 | 395 ± 101 | 42 ± 7 | 26 ± 7 | 32 ± 5 |
| Jun. | 2.5 ± 1.4 | 32 ± 19 | 50 ± 21 | 18 ± 6 | 239 ± 182 | 122 ± 64 | 131 ± 82 | 492 ± 317 | 47 ± 8 | 26 ± 5 | 27 ± 8 |
| Jul. | 3.2 ± 1.8 | 40 ± 15 | 41 ± 15 | 19 ± 6 | 269 ± 94 | 202 ± 108 | 158 ± 99 | 630 ± 250 | 45 ± 11 | 31 ± 8 | 24 ± 6 |
| Aug. | 1.6 ± 1.5 | 62 ± 16 | 21 ± 11 | 16 ± 8 | 168 ± 71 | 101 ± 34 | 101 ± 32 | 370 ± 125 | 45 ± 6 | 28 ± 5 | 28 ± 4 |
| Sep. | 0.8 ± 0.2 | 32 ± 15 | 45 ± 14 | 23 ± 18 | 255 ± 48 | 42 ± 11 | 85 ± 14 | 382 ± 53 | 66 ± 6 | 11 ± 3 | 23 ± 4 |
| Oct. | 4.9 ± 1.6 | 75 ± 11 | 17 ± 8 | 8 ± 3 | 239 ± 48 | 157 ± 43 | 117 ± 18 | 513 ± 83 | 47 ± 6 | 30 ± 6 | 23 ± 3 |
| Nov. | 1.4 ± 0.7 | 51 ± 20 | 28 ± 12 | 20 ± 9 | 240 ± 36 | 45 ± 19 | 89 ± 22 | 375 ± 45 | 64 ± 8 | 12 ± 5 | 24 ± 5 |
| Dec. | 0.9 ± 0.4 | 55 ± 14 | 23 ±6 | 22 ± 13 | 172 ± 23 | 41 ± 14 | 85 ± 32 | 297 ± 49 | 58 ± 7 | 14 ± 4 | 28 ± 7 |
| Variables in | Standardized Weight of Variables in Selected | Loading of Variables (vik) | Communalities | ||||
|---|---|---|---|---|---|---|---|
| PC (tik; I = 1, 2, …, 12 and k = 1, 2) | |||||||
| PC 1 | PC 2 | PC 3 | PC 1 | PC 2 | PC 3 | ||
| Temperature | −0.103 | 0.18 | 0.373 | 0.109 | 0.506 | 0.587 | 0.612 |
| NH4+ | −0.002 | 0.35 | −0.06 | −0.087 | 0.874 | −0.065 | 0.775 |
| NO3− | 0.054 | 0.217 | −0.028 | 0.154 | 0.543 | 0.047 | 0.321 |
| P* | −0.052 | 0.289 | −0.141 | −0.371 | 0.708 | −0.289 | 0.723 |
| SiO2-Si | −0.017 | 0.273 | 0.222 | 0.221 | 0.719 | 0.43 | 0.75 |
| Micro | −0.102 | −0.133 | 0.416 | 0.171 | −0.276 | 0.624 | 0.495 |
| Nano | 0.204 | 0.046 | 0.033 | 0.772 | 0.118 | 0.33 | 0.719 |
| Pico | −0.107 | 0.023 | 0.335 | 0.048 | 0.105 | 0.489 | 0.253 |
| CHO | 0.357 | 0.108 | −0.357 | 0.818 | 0.219 | −0.19 | 0.754 |
| PRT | 0.094 | −0.068 | 0.277 | 0.694 | −0.133 | 0.627 | 0.892 |
| LIP | 0.252 | −0.053 | −0.075 | 0.805 | −0.147 | 0.177 | 0.702 |
| FM | 0.304 | 0.012 | −0.099 | 0.961 | 0.013 | 0.209 | 0.967 |
| Included Independent | Regression | Standard | Standardized Regression | t Statics | p Value | Adjusted R2 (%) |
|---|---|---|---|---|---|---|
| Variables | Coefficient (bk) | Error of bk | Coefficient | |||
| Constant | 337.872 | 12.08 | 27.969 | 0.000 ** | ||
| Nano-chlorophyll a concentration | 112.476 | 8.16 | 0.617 | 13.784 | 0.000 ** | 0.544 |
| Micro-chlorophyll a concentration | 20.115 | 5.412 | 0.156 | 3.716 | 0.000 ** | 0.57 |
| P* | −230.321 | 49.425 | −0.305 | −4.66 | 0.000 ** | 0.582 |
| NH4+ concentration | 19.321 | 5.362 | 0.225 | 3.603 | 0.000 ** | 0.602 |
| Region | Period | Total Chlorophyll a Concentration | FM Concentration | Reference |
|---|---|---|---|---|
| (μg L−1) | (mg m−3) | |||
| Gwangyang Bay, Korea | Seasonally, 2012–2013 | 3.45 (±2.81) | 434.5 (±175.5) | [5] |
| Geoje-Hansan Bay, Korea | Monthly, 2015 | 4.34 (±2.42) | 615.5 (±291.7) | [6] |
| Garolim-Asan Bay, Korea | Seasonally, 2015–2016 | 2.81 (±2.12) | 781.4 (±228.2) | [37] |
| Jaran Bay, Korea | Monthly, 2016 | 2.13 (±1.18) | 411.7 (±93.0) | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Lee, W.-C.; Kim, H.C.; Jo, N.; Kim, K.; Lee, D.; Kang, J.J.; Sim, B.-R.; Kwon, J.-I.; Lee, S.H. Temporal and Spatial Variations of the Biochemical Composition of Phytoplankton and Potential Food Material (FM) in Jaran Bay, South Korea. Water 2020, 12, 3093. https://doi.org/10.3390/w12113093
Lee JH, Lee W-C, Kim HC, Jo N, Kim K, Lee D, Kang JJ, Sim B-R, Kwon J-I, Lee SH. Temporal and Spatial Variations of the Biochemical Composition of Phytoplankton and Potential Food Material (FM) in Jaran Bay, South Korea. Water. 2020; 12(11):3093. https://doi.org/10.3390/w12113093
Chicago/Turabian StyleLee, Jae Hyung, Won-Chan Lee, Hyung Chul Kim, Naeun Jo, Kwanwoo Kim, Dabin Lee, Jae Joong Kang, Bo-Ram Sim, Jae-Il Kwon, and Sang Heon Lee. 2020. "Temporal and Spatial Variations of the Biochemical Composition of Phytoplankton and Potential Food Material (FM) in Jaran Bay, South Korea" Water 12, no. 11: 3093. https://doi.org/10.3390/w12113093
APA StyleLee, J. H., Lee, W.-C., Kim, H. C., Jo, N., Kim, K., Lee, D., Kang, J. J., Sim, B.-R., Kwon, J.-I., & Lee, S. H. (2020). Temporal and Spatial Variations of the Biochemical Composition of Phytoplankton and Potential Food Material (FM) in Jaran Bay, South Korea. Water, 12(11), 3093. https://doi.org/10.3390/w12113093