Drought or Severe Drought? Hemiparasitic Yellow Mistletoe (Loranthus europaeus) Amplifies Drought Stress in Sessile Oak Trees (Quercus petraea) by Altering Water Status and Physiological Responses

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Design Description
2.2. Leaf Water Potential
2.3. Gas Exchange Measurements
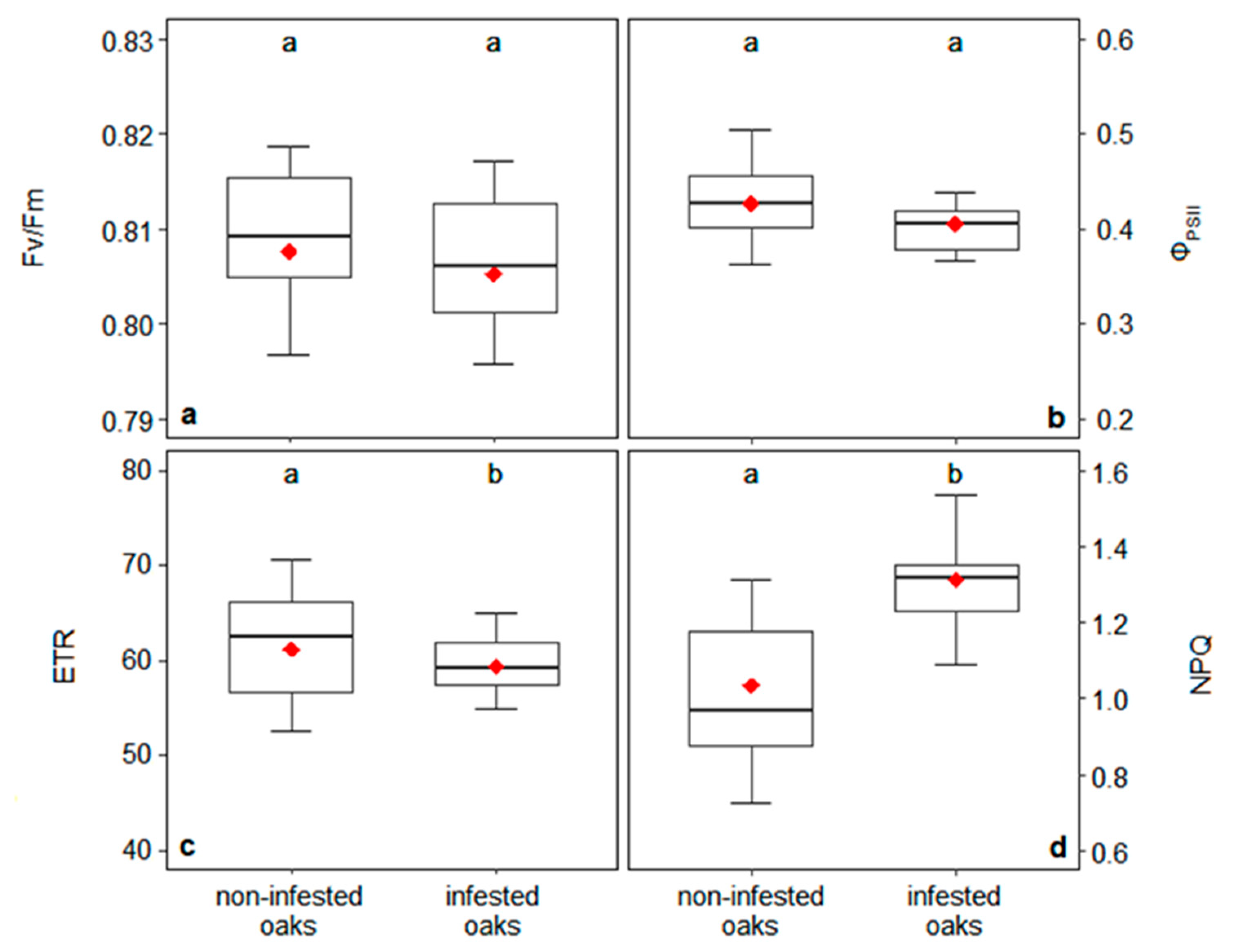
2.4. Chlorophyll a Fluorescence Measurements
2.5. Content of Nutrients in Plants and Soil
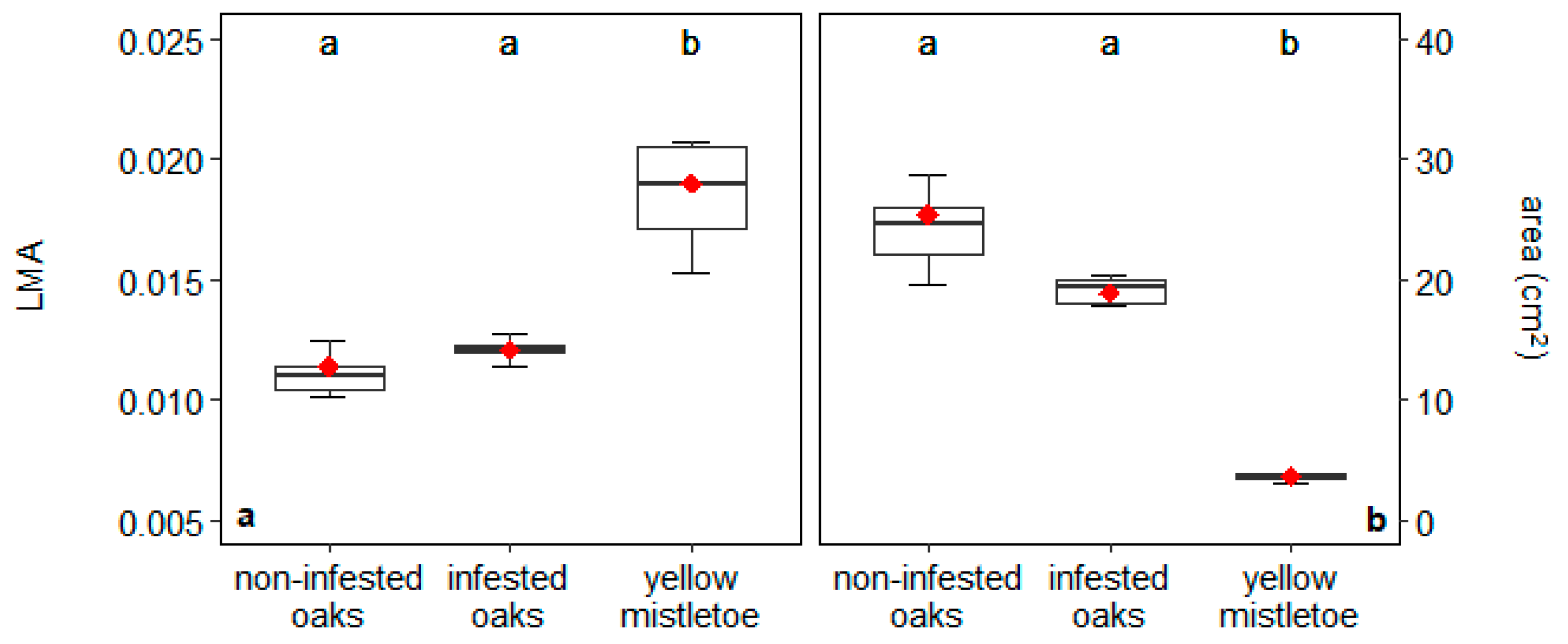
2.6. Leaf Morphological Traits
2.7. Statistical Analysis
3. Results
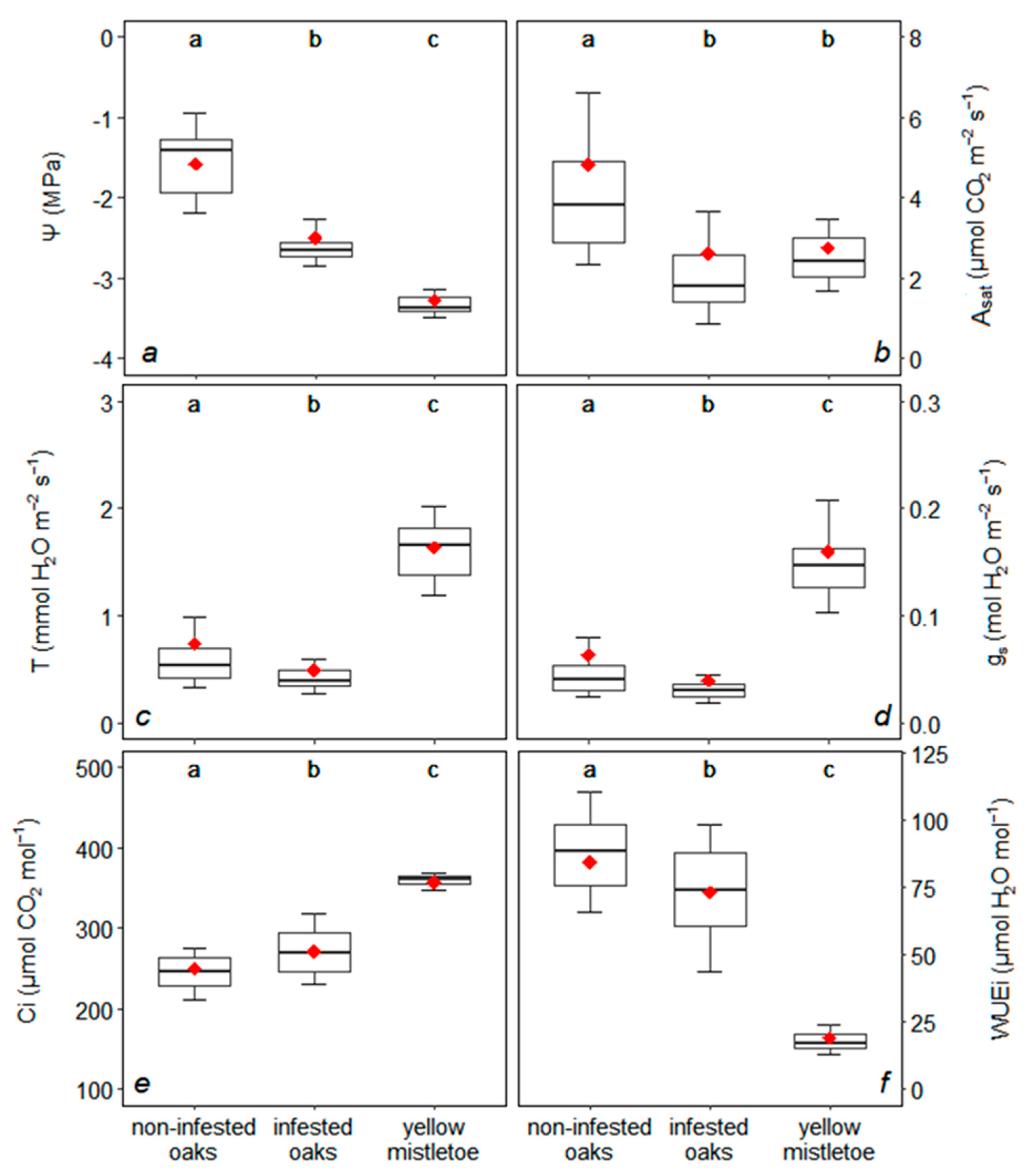
3.1. Leaf Water Potential and Photosynthesis
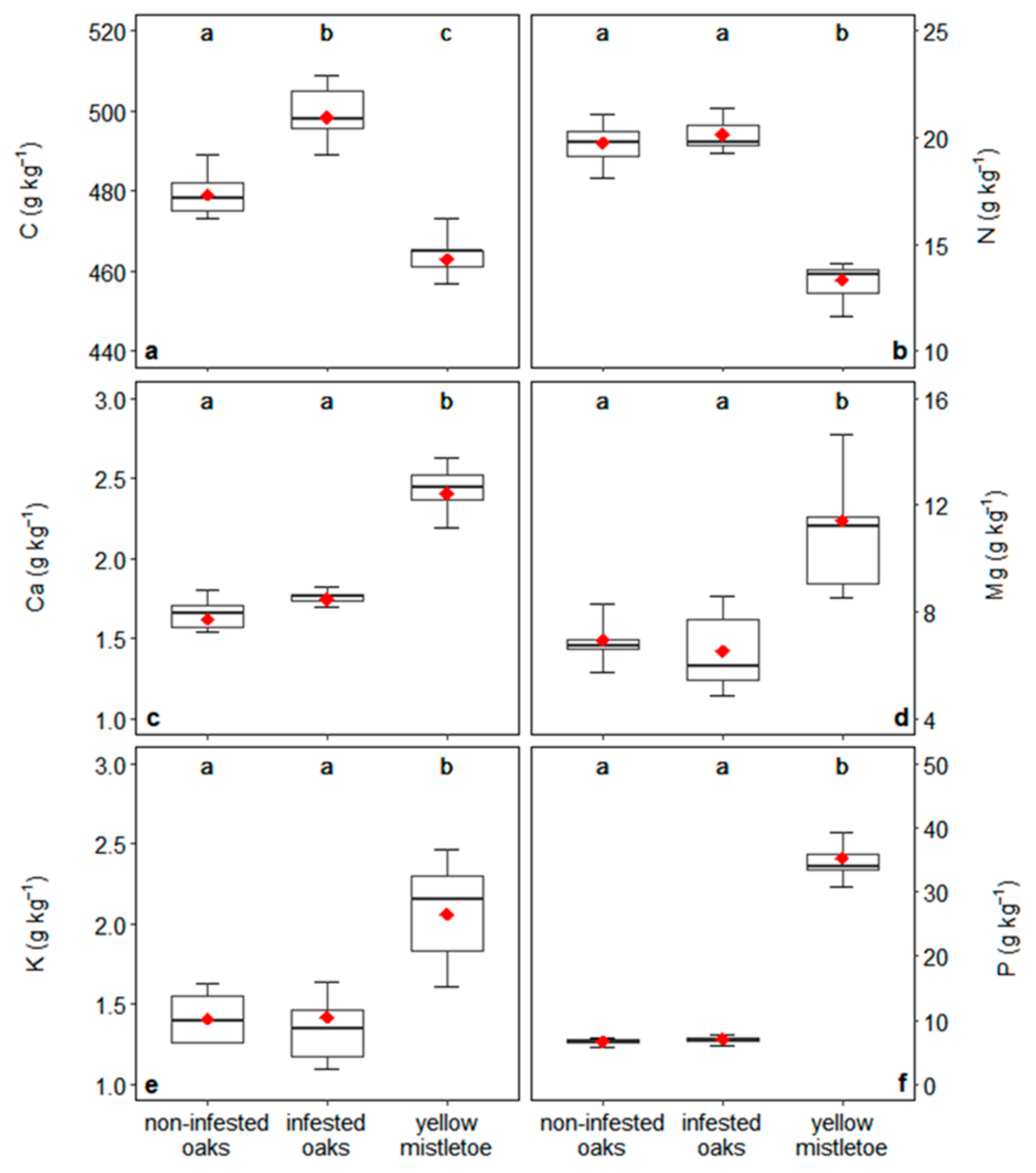
3.2. Content of Nutrients in Plants and Their Absorption
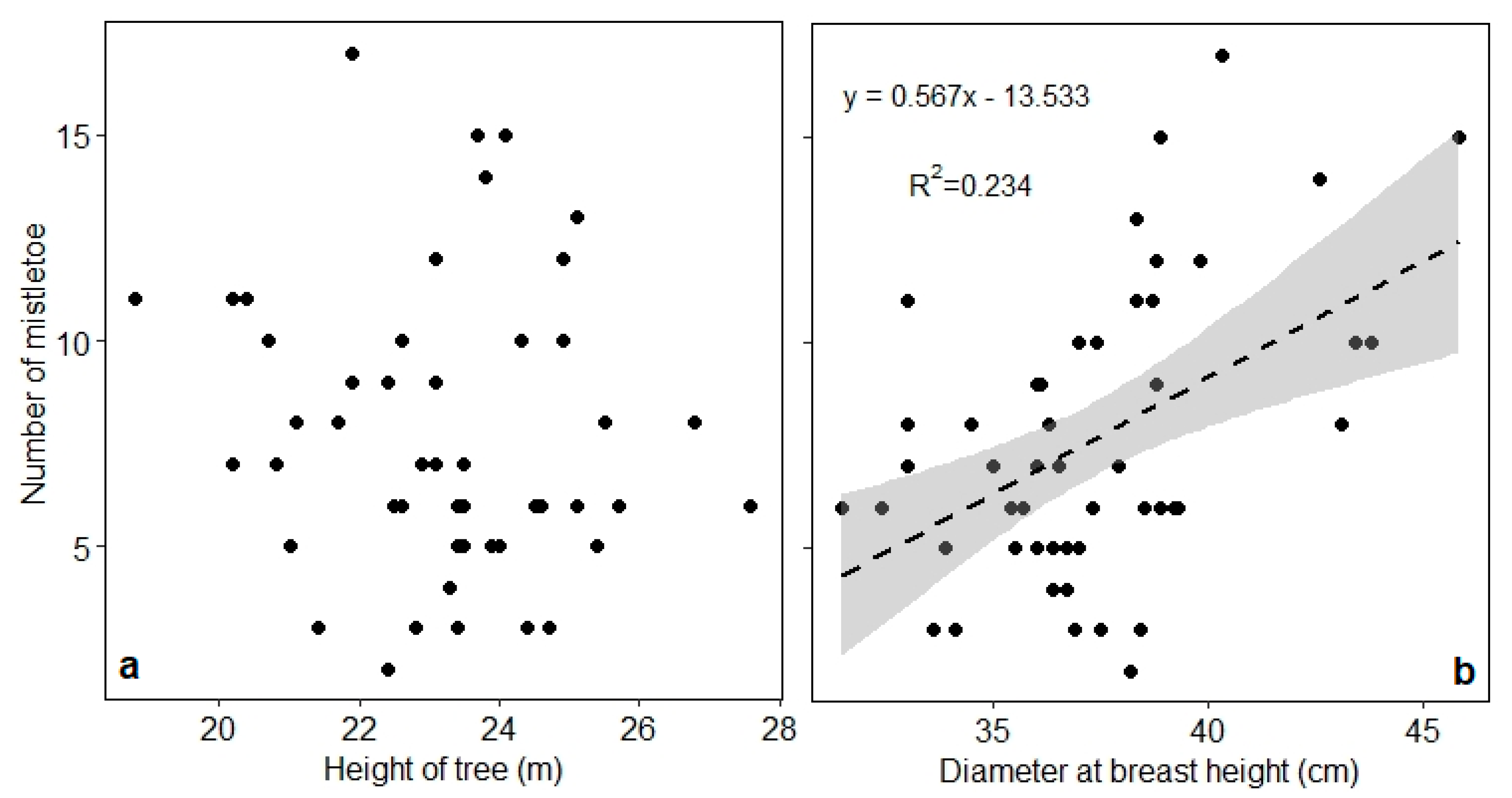
3.3. Growth and Rate of Infestation
4. Discussion
4.1. Impact of Mistletoe on Photosynthesis and Water Relationships
4.2. Relationships between Oaks and Mistletoes: Growth and Mineral Nutrition
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dimopoulos, P.; Bergmeier, E.; Chytrý, M.; Rodwell, J.; Schaminée, J.H.J.; Sykora, K.V. European oak woodlands: Past, present and future. Bot. Chron. Patra 2005, 18, 1–316. [Google Scholar]
- Rock, J.; Gockel, H.; Schulte, A. Vegetationsdiversität in Eichen-Jungwüchsen aus unterschiedlichen Pflanzschemata. Beitr. Für Forstwirtsch. Landschaftsökologie 2003, 37, 11–17. [Google Scholar]
- Kunz, J.; Löffler, G.; Bauhus, J. Minor European broadleaved tree species are more drought-tolerant than Fagus sylvatica but not more tolerant than Quercus petraea. For. Ecol. Manag. 2018, 414, 15–27. [Google Scholar] [CrossRef]
- Breda, N.; Cochard, H.; Dreyer, E.; Granier, A. Water transfer in a mature oak stand (Quercus petraea): Seasonal evolution and effects of a severe drought. Can. J. Forest Res. 1993, 23, 1136–1143. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, R.K.; Mayer, L.; Core Writing Team (Eds.) IPCC Climate Change 2014: Synthesis Report; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Mert, A.; Özkan, K.; Şentürk, Ö.; Negiz, M.G. Changing the potential distribution of Turkey oak (IL.) under climate change in Turkey. Pol. J. Environ. Stud. 2016, 25, 1633–1638. [Google Scholar] [CrossRef]
- Sallé, A.; Nageleisen, L.-M.; Lieutier, F. Bark and wood boring insects involved in oak declines in Europe: Current knowledge and future prospects in a context of climate change. For. Ecol. Manag. 2014, 328, 79–93. [Google Scholar] [CrossRef]
- Olano, J.M.; Laskurain, N.A.; Escudero, A.; De La Cruz, M. Why and where do adult trees die in a young secondary temperate forest? The role of neighbourhood. Ann. For. Sci. 2009, 66, 105. [Google Scholar] [CrossRef]
- Thomas, F.M.; Blank, R.; Hartmann, G. Abiotic and biotic factors and their interactions as causes of oak decline in Central Europe. For. Pathol. 2002, 32, 277–307. [Google Scholar] [CrossRef]
- Bréda, N. Water shortage as a key factor in the case of the oak dieback in the Harth Forest (Alsatian plain, France) as demonstrated by dendroecological and ecophysiological study. In Recent Advances on Oak Health in Europe. Selected Papers from a Conference Held in Warsaw, Poland, 22–24 November 1999; Instytut Badawczy Leśnictwa (Forest Research Institute): Warsaw, Poland, 2000; pp. 157–159. [Google Scholar]
- Doležal, J.; Mazůrek, P.; Klimešová, J. Oak decline in southern Moravia: The association between climate change and early and late wood formation in oaks. Preslia 2010, 82, 289–306. [Google Scholar]
- Sohar, K.; Helama, S.; Läänelaid, A.; Raisio, J.; Tuomenvirta, H. Oak decline in a southern Finnish forest as affected by a drought sequence. Geochronometria 2014, 41, 92–103. [Google Scholar] [CrossRef]
- Sonesson, K.; Drobyshev, I. Recent advances on oak decline in southern Sweden. Ecol. Bull. 2010, 11, 197–208. [Google Scholar]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Marçis, B.; Caël, O.; Delatour, C. Relationship between presence of basidiomes, above-ground symptoms and root infection by Collybia fusipes in oaks. For. Pathol. 2000, 30, 7–17. [Google Scholar] [CrossRef][Green Version]
- Hajji, M.; Dreyer, E.; Marçais, B. Impact of Erysiphe alphitoides on transpiration and photosynthesis in Quercus robur leaves. Eur. J. Plant Pathol. 2009, 125, 63–72. [Google Scholar] [CrossRef]
- Galko, J.; Nikolov, C.; Kimoto, T.; Kunca, A.; Gubka, A.; Vakula, J.; Zúbrik, M.; Ostrihoň, M. Attraction of ambrosia beetles to ethanol baited traps in a Slovakian oak forest. Biologia (Bratisl.) 2014, 69, 1376–1383. [Google Scholar] [CrossRef]
- Millaku, F.; Sahiti, G.; Abdullahu, K.; Krasniqi, E. The spread and the infection frecuency of the Golliak (Kosovo) forest with the species hemipariastoc mistletoe (Loranthus europaeus L.). Int. Multidiscip. Sci. GeoConf. SGEM 2011, 2011, 1035–1040. [Google Scholar]
- Kumbasli, M.; Keten, A.; Beskardes, V.; Makineci, E.; Özdemir, E.; Yilmaz, E.; Zengin, H.; Sevgi, O.; Yilmaz, H.C.; Caliskan, S. Hosts and distribution of yellow mistletoe (Loranthus europaeus Jacq. (Loranthaceae) on Northern Strandjas Oak Forests-Turkey. Sci. Res. Essays 2011, 6, 2970–2975. [Google Scholar]
- Gebauer, R.; Volařík, D.; Urban, J. Seasonal variations of sulphur, phosphorus and magnesium in the leaves and current-year twigs of hemiparasitic mistletoe Loranthus europaeus Jacq. and its host Quercus pubescens Willd. J. For. Sci. 2018, 64, 66–73. [Google Scholar]
- Schulze, E.D.; Ehleringer, J.R. The effect of nitrogen supply on growth and water-use efficiency of xylem-tapping mistletoes. Planta 1984, 162, 268–275. [Google Scholar] [CrossRef]
- Matula, R.; Svátek, M.; Pálková, M.; Volařík, D.; Vrška, T. Mistletoe infection in an oak forest is influenced by competition and host size. PLoS ONE 2015, 10, e0127055. [Google Scholar] [CrossRef]
- Zweifel, R.; Bangerter, S.; Rigling, A.; Sterck, F.J. Pine and mistletoes: How to live with a leak in the water flow and storage system? J. Exp. Bot. 2012, 63, 2565–2578. [Google Scholar] [CrossRef]
- Rigling, A.; Eilmann, B.; Koechli, R.; Dobbertin, M. Mistletoe-induced crown degradation in Scots pine in a xeric environment. Tree Physiol. 2010, 30, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Glatzel, G.; Geils, B.W. Mistletoe ecophysiology: Host-parasite interactions. Botany 2009, 87, 10–15. [Google Scholar] [CrossRef]
- Press, M.C.; Phoenix, G.K. Impacts of parasitic plants on natural communities. New Phytol. 2005, 166, 737–751. [Google Scholar] [CrossRef]
- Glatzel, G. Mineral nutrition and water relations of hemiparasitic mistletoes: A question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur. Oecologia 1983, 56, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Mathiasen, R.L.; Nickrent, D.L.; Shaw, D.C.; Watson, D.M. Mistletoes - ecology, systematics, ecology and management. Plant Dis. 2008, 92, 987–1006. [Google Scholar] [CrossRef]
- Szmidla, H.; Tkaczyk, M.; Plewa, R.; Tarwacki, G.; Sierota, Z. Impact of Common Mistletoe (Viscum album L.) on Scots Pine Forests—A Call for Action. Forests 2019, 10, 847. [Google Scholar] [CrossRef]
- Eliáš, P. Úhyn imelovca (Loranthus europaeus Jacq.) na severnej hranici rozšírenia v Európe: Slovensko (in Slovak)/Mistletoe (Loranthus europaeus Jacq.) mortality at northern geographical distribution in Europe: Slovakia. In Dreviny v Mestskom Prostredí a v Krajine. Aktualne Trendy Dendrologického Výskumu a Praxe; Slovak University of Agriculture: Nitra, Slovakia, 2007. [Google Scholar]
- Kubíček, J.; Martinková, M.; Špinlerová, Z. Hemiparazité a provozní bezpečnost jimi napadených stromů (in Czech)/Hemiparasites and operating safety of infested trees. In Provozní Bezpečnost Stromů; Mendel University in Brno: Brno, Czech Republic, 2011; pp. 44–50. [Google Scholar]
- McDowell, N.G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Doležal, J.; Lehečková, E.; Sohar, K.; Altman, J. Oak decline induced by mistletoe, competition and climate change: A case study from central Europe. Preslia 2016, 88, 323–346. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.R. Biological Methods of Prospecting for Minerals; Wiley: Chichester, UK; New York, NY, USA, 1983. [Google Scholar]
- Griebel, A.; Watson, D.; Pendall, E. Mistletoe, friend and foe: Synthesizing ecosystem implications of mistletoe infection. Environ. Res. Lett. 2017, 12, 115012. [Google Scholar] [CrossRef]
- Cochard, H.; Bréda, N.; Granier, A. Whole tree hydraulic conductance and water loss regulation in Quercus during drought: Evidence for stomatal control of embolism? Ann. Sci. For. 1996, 53, 197–206. [Google Scholar] [CrossRef]
- Zuber, D. Biological flora of Central Europe: Viscum album L. Flora—Morphol. Distrib. Funct. Ecol. Plants 2004, 199, 181–203. [Google Scholar] [CrossRef]
- Schulze, E.D.; Turner, N.C.; Glatzel, G. Carbon, water and nutrient relations of two mistletoes and their hosts: A hypothesis. Plant Cell Environ. 1984, 7, 293–299. [Google Scholar]
- Davidson, N.J.; True, K.C.; Pate, J.S. Water relations of the parasite: Host relationship between the mistletoe Amyema linophyllum (Fenzl) Tieghem and Casuarina obesa Miq. Oecologia 1989, 80, 321–330. [Google Scholar] [CrossRef]
- Steckel, M.; del Río, M.; Heym, M.; Aldea, J.; Bielak, K.; Brazaitis, G.; Černý, J.; Coll, L.; Collet, C.; Ehbrecht, M.; et al. Species mixing reduces drought susceptibility of Scots pine (Pinus sylvestris L.) and oak (Quercus robur L., Quercus petraea (Matt.) Liebl.)—Site water supply and fertility modify the mixing effect. For. Ecol. Manag. 2020, 461, 117908. [Google Scholar] [CrossRef]
- Sade, N.; Gebremedhin, A.; Moshelion, M. Risk-taking plants: Anisohydric behavior as a stress-resistance trait. Plant Signal. Behav. 2012, 7, 767–770. [Google Scholar] [CrossRef]
- Johnson, J.M.; Choinski, J.S. Photosynthesis in the Tapinanthus-Diplorhynchus Mistletoe-Host Relationship. Ann. Bot. 1993, 72, 117–122. [Google Scholar] [CrossRef]
- Ullmann, I.; Lange, O.L.; Ziegler, H.; Ehleringer, J.; Schulze, E.-D.; Cowan, I.R. Diurnal courses of leaf conductance and transpiration of mistletoes and their hosts in Central Australia. Oecologia 1985, 67, 577–587. [Google Scholar] [CrossRef]
- Küppers, M.; Küppers, B.I.L.; Neales, T.F.; Swan, A.G. Leaf gas exchange characteristics, daily carbon and water balances of the host/mistletoe pair Eucalyptus behriana F. Muell. and Amyema miquelii (Lehm. ex Miq.) Tiegh. at permanently low plant water status in the field. Trees 1992, 7, 1–7. [Google Scholar]
- Urban, J.; Gebauer, R.; Nadezhdina, N.; Čermák, J. Transpiration and stomatal conductance of mistletoe (Loranthus europaeus) and its host plant, downy oak (Quercus pubescens). Biologia (Bratisl.) 2012, 67, 917–926. [Google Scholar] [CrossRef]
- Orcutt, D.M.; Nilsen, E.T. Physiology of Plants under Stress: Soil and Biotic Factors; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000. [Google Scholar]
- Goldstein, G.; Rada, F.; Sternberg, L.; Burguera, J.L.; Burguera, M.; Orozco, A.; Montilla, M.; Zabala, O.; Azocar, A.; Canales, M.J.; et al. Gas exchange and water balance of a mistletoe species and its mangrove hosts. Oecologia 1989, 78, 176–183. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Schulze, E.-D.; Ziegler, H.; Lange, O.L.; Farquhar, G.D.; Cowar, I.R. Xylem-tapping mistletoes: Water or nutrient parasites? Science 1985, 227, 1479–1481. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef]
- Queijeiro-Bolaños, M.E.; Malda-Barrera, G.X.; Carrillo-Angeles, I.G.; Suzán-Azpiri, H. Contrasting gas exchange effects on the interactions of two mistletoe species and their host Acacia schaffneri. J. Arid Environ. 2020, 173, 104041. [Google Scholar] [CrossRef]
- Epron, D.; Dreyer, E.; Bréda, N. Photosynthesis of oak trees [Quercus petraea (Matt.) Liebl.] during drought under field conditions: Diurnal course of net CO2 assimilation and photochemical efficiency of photosystem II. Plant Cell Environ. 1992, 15, 809–820. [Google Scholar] [CrossRef]
- Morecroft, M.D.; Roberts, J.M. Photosynthesis and stomatal conductance of mature canopy oak (Quercus robur) and sycamore (Acer pseudoplatanus) trees throughout the growing season. Funct. Ecol. 1999, 13, 332–342. [Google Scholar] [CrossRef]
- Osuna, J.L.; Baldocchi, D.D.; Kobayashi, H.; Dawson, T.E. Seasonal trends in photosynthesis and electron transport during the Mediterranean summer drought in leaves of deciduous oaks. Tree Physiol. 2015, 35, 485–500. [Google Scholar] [CrossRef]
- Strong, G.L.; Bannister, P.; Burritt, D. Are mistletoes shade plants? CO2 assimilation and chlorophyll fluorescence of temperate mistletoes and their hosts. Ann. Bot. 2000, 85, 511–519. [Google Scholar] [CrossRef]
- Marshall, J.D.; Dawson, T.E.; Ehleringer, J.R. Integrated nitrogen, carbon, and water relations of a xylem-tapping mistletoe following nitrogen fertilization of the host. Oecologia 1994, 100, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Lüttge, U.; Haridasan, M.; Fernandes, G.W.; de Mattos, E.A.; Trimborn, P.; Franco, A.C.; Caldas, L.S.; Ziegler, H. Photosynthesis of mistletoes in relation to their hosts at various sites in tropical Brazil. Trees 1998, 12, 167–174. [Google Scholar] [CrossRef]
- Tennakoon, K.U.; Pate, J.S. Heterotrophic gain of carbon from hosts by the xylem-tapping root hemiparasite Olax phyllanthi (Olacaceae). Oecologia 1996, 105, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.D.; Ehleringer, J.R. Are xylem-tapping mistletoes partially heterotrophic? Oecologia 1990, 84, 244–248. [Google Scholar] [CrossRef]
- Schulze, E.D.; Gebauer, G.; Ziegler, H.; Lange, O.L. Estimates of nitrogen fixation by trees on an aridity gradient in Namibia. Oecologia 1991, 88, 451–455. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Linares, J.C.; Julio Camarero, J. Drought and mistletoe reduce growth and water-use efficiency of Scots pine. For. Ecol. Manag. 2013, 296, 64–73. [Google Scholar] [CrossRef]
- Cornic, G.; Massacci, A. Leaf photosynthesis under drought stress. In Photosynthesis and the Environment; Advances in photosynthesis and respiration; Baker, N.R., Ed.; Springer: Dordrecht, The Netherlands, 1996; pp. 347–366. [Google Scholar]
- Flexas, J.; Medrano, H. Energy dissipation in C3 plants under drought. Funct. Plant Biol. 2002, 29, 1209. [Google Scholar] [CrossRef]
- Flexas, J.; Escalona, J.M.; Medrano, H. Down-regulation of photosynthesis by drought under field conditions in grape vine leaves. Aust. J. Plant Physiol. 1998, 25, 893–900. [Google Scholar]
- Björkman, O.; Demmig-Adams, B. Regulation of photosynthetic light energy capture, conversion, and dissipation in leaves of higher plants. In Ecophysiology of Photosynthesis; Schulze, E.-D., Caldwell, M.M., Eds.; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 1995; pp. 17–47. [Google Scholar]
- Aziz, A.; Larher, F. Osmotic stress induced changes in lipid composition and peroxidation in leaf discs of Brassica napus L. J. Plant Physiol. 1998, 153, 754–762. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M. PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: Protocols and applications. In Molecular Stress Physiology of Plants; Rout, G.R., Das, A.B., Eds.; Springer India: New Delhi, India, 2013; pp. 87–131. [Google Scholar]
- Konôpková, A.; Húdoková, H.; Ježík, M.; Kurjak, D.; Jamnická, G.; Ditmarová, Ľ.; Gömöry, D.; Longauer, R.; Tognetti, R.; Pšidová, E. Origin rather than mild drought stress influenced chlorophyll a fluorescence in contrasting silver fir (Abies alba Mill.) provenances. Photosynthetica 2020, 58, 549–559. [Google Scholar] [CrossRef]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Berger, K.H.M.; Nayyar, N. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. FPB 2014, 41, 1148–1167. [Google Scholar] [CrossRef] [PubMed]
- Forner, A.; Valladares, F.; Aranda, I. Mediterranean trees coping with severe drought: Avoidance might not be safe. Environ. Exp. Bot. 2018, 155, 529–540. [Google Scholar] [CrossRef]
- Galmés, J.; Ribas-Carbó, M.; Medrano, H.; Flexas, J. Rubisco activity in Mediterranean species is regulated by the chloroplastic CO2 concentration under water stress. J. Exp. Bot. 2011, 62, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Galmés, J.; Flexas, J.; Savé, R.; Medrano, H. Water relations and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: Responses to water stress and recovery. Plant Soil 2007, 290, 139–155. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. Xanthophyll cycle and light stress in nature: Uniform response to excess direct sunlight among higher plant species. Planta 1996, 198, 460–470. [Google Scholar] [CrossRef]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 182–193. [Google Scholar] [CrossRef]
- Kościelniak, J.; Filek, W.; Biesaga-Kościelniak, J. The effect of drought stress on chlorophyll fluorescence in Lolium-Festuca hybrids. Acta Physiol. Plant. 2006, 28, 149–158. [Google Scholar] [CrossRef]
- Aukema, J.E.; Martinéz del Rio, C. Where does a fruit-eating bird deposit mistletoe seeds? Seed deposition patterns and an experiment. Ecology 2002, 83, 3489–3496. [Google Scholar] [CrossRef]
- Monteiro, R.F.; Martins, R.P.; Yamamoto, K. Host specificity and seed dispersal of Psittacanthus robustus (Loranthaceae) in Southeast Brazil. J. Trop. Ecol. 1992, 8, 307–314. [Google Scholar] [CrossRef]
- Skórka, P.; Wójcik, J.D. Winter bird communities in a managed mixed oak-pine forest (Niepolomice Forest, Southern Poland). Acta Zool. Crac. 2003, 46, 29–41. [Google Scholar]
- Eliáš, P. Quantitative ecological analysis of a mistletoe Loranthus europaeus jacq. population in an oak hornbeam forest space continuum approach. Ekologia 1987, 6, 359–372. [Google Scholar]
- Overton, J.M.C. Dispersal and infection in mistletoe metapopulations. J. Ecol. 1994, 82, 711–723. [Google Scholar] [CrossRef]
- Arruda, R.; Fadini, R.F.; Carvalho, L.N.; Del Claro, K.; Mourão, F.A.; Jacobi, C.M.; Teodoro, G.S.; Berg, E.; Caires, C.S.; Dettke, G.A. Ecology of neotropical mistletoes: An important canopy-dwelling component of Brazilian ecosystems. Acta Bot. Bras. 2012, 26, 264–274. [Google Scholar] [CrossRef]
- Gressler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought-induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Michopoulos, P.; Kaoukis, K.; Karetsos, G.; Grigoratos, T.; Samara, C. Nutrients in litterfall, forest floor and mineral soils in two adjacent forest ecosystems in Greece. J. For. Res. 2020, 31, 291–301. [Google Scholar] [CrossRef]
- Nilsson, L.-O. Forest biogeochemistry interactions among greenhouse gases and N deposition. Water. Air. Soil Pollut. 1995, 85, 1557–1562. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Kartoolinejad, D.; Mirnia, S.K.; Tabibzadeh, Z.; Akbarinia, M.; Shayanmehr, F. The effects of Viscum album L. on foliar weight and nutrients content of host trees in Caspian forests (Iran). Pol. J. Ecol. 2007, 55, 579–583. [Google Scholar]
- Hosseini, S.M.; Kartoolinejad, D.; Mirnia, S.K.; Tabibzadeh, Z.; Akbarinia, M.; Shayanmehr, F. The European mistletoe effects on leaves and nutritional elements of two host species in Hyrcanian forests. Silva Luisiana 2008, 16, 9. [Google Scholar]
- Lamont, B. Mineral nutrition of mistletoes. In The Biology of Mistletoes; Calder, M., Bernhardt, P., Eds.; Academic Press Australia: Sydney, Australia, 1983; pp. 185–204. [Google Scholar]
- Kutbay, H.; Karaer, F.; Kilinç, M. The Relationships of Some Nutrients between Cuscuta epithymum (L.) L. var. epithymum and Heliotropium europaeum L. Available online: /paper/The-Relationships-of-Some-Nutrients-Between-Cuscuta-Kutbay-Karaer/cf9b77e40ef40542f6de9a39ef0e5a4f1d69a1ca (accessed on 1 September 2020).
- Malicki, L.; Berbeciowa, C. Content of basic macroelements in common parasitical weeds. Acta Agrobot. 1986, 39, 123–128. [Google Scholar] [CrossRef][Green Version]
- Türe, C.; Böcük, H.; Aşan, Z. Nutritional relationships between hemi-parasitic mistletoe and some of its deciduous hosts in different habitats. Biologia (Bratisl.) 2010, 65, 859–867. [Google Scholar] [CrossRef]
- Bowie, M.; Ward, D. Water and nutrient status of the mistletoe Plicosepalus acaciae parasitic on isolated Negev Desert populations of Acacia raddiana differing in level of mortality. J. Arid Environ. 2004, 56, 487–508. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling and Measurements | Oak Trees Per Variant/Repetitions | Yellow Mistletoes/Repetitions |
|---|---|---|
| Diameter at breast height | 50/1 | - |
| Height of tree | 50/1 | - |
| Height of the first branch | 50/1 | - |
| Stem increment | 50/1 | - |
| Leaf water potential | 13/2–4 | 10/1 |
| Gas exchange | 14/8 | 14/6 |
| Chlorophyll a florescence | 14/5 | - |
| Content of nutrients in leaves | 14/1 (ca. 30 leaves) | 14/1 (ca. 100 leaves) |
| Leaf morphological traits | 14/1 (ca. 20 leaves) | 14/1 (ca. 100 leaves) |
| Carbon | Nitrogen | Calcium | Magnesium | Potassium | Phosphorus | |
|---|---|---|---|---|---|---|
| Non-infested tree | 21.7 ± 0.92 b | 17.3 ± 2.31 a | 12.3 ± 1.96 a | 190.9 ± 37.11 a | 18.4 ± 4.77 a | 55.7 ± 9.57 a |
| Infested tree | 22.6 ± 0.60 a | 17.7 ± 1.77 a | 13.2 ± 1.28 a | 179.8 ± 69.83 a | 18.5 ± 5.46 a | 59.2 ± 12.71 a |
| Mistletoe | 0.9 ± 0.02 | 0.7 ± 0.13 | 1.4 ± 0.23 | 1.9 ± 0.58 | 1.5 ± 0.27 | 5.2 ± 1.44 |
| DBH (cm) | Height (m) | Height of the First Branch (m) | Stem Increment (mm) | |
|---|---|---|---|---|
| Non-infested trees | 36.6 ± 3.5 a | 25.2 ± 1.7 a | 9.1 ± 3.3 a | 1.75 ± 1.4 a |
| Infested trees | 39.2 ± 5.1 a | 23.2 ± 1.9 b | 6.7 ± 2.6 b | 2.38 ± 1.7 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubov, M.; Fleischer, P., Jr.; Rozkošný, J.; Kurjak, D.; Konôpková, A.; Galko, J.; Húdoková, H.; Lalík, M.; Rell, S.; Pittner, J.; et al. Drought or Severe Drought? Hemiparasitic Yellow Mistletoe (Loranthus europaeus) Amplifies Drought Stress in Sessile Oak Trees (Quercus petraea) by Altering Water Status and Physiological Responses. Water 2020, 12, 2985. https://doi.org/10.3390/w12112985
Kubov M, Fleischer P Jr., Rozkošný J, Kurjak D, Konôpková A, Galko J, Húdoková H, Lalík M, Rell S, Pittner J, et al. Drought or Severe Drought? Hemiparasitic Yellow Mistletoe (Loranthus europaeus) Amplifies Drought Stress in Sessile Oak Trees (Quercus petraea) by Altering Water Status and Physiological Responses. Water. 2020; 12(11):2985. https://doi.org/10.3390/w12112985
Chicago/Turabian StyleKubov, Martin, Peter Fleischer, Jr., Jozef Rozkošný, Daniel Kurjak, Alena Konôpková, Juraj Galko, Hana Húdoková, Michal Lalík, Slavomír Rell, Ján Pittner, and et al. 2020. "Drought or Severe Drought? Hemiparasitic Yellow Mistletoe (Loranthus europaeus) Amplifies Drought Stress in Sessile Oak Trees (Quercus petraea) by Altering Water Status and Physiological Responses" Water 12, no. 11: 2985. https://doi.org/10.3390/w12112985
APA StyleKubov, M., Fleischer, P., Jr., Rozkošný, J., Kurjak, D., Konôpková, A., Galko, J., Húdoková, H., Lalík, M., Rell, S., Pittner, J., & Fleischer, P. (2020). Drought or Severe Drought? Hemiparasitic Yellow Mistletoe (Loranthus europaeus) Amplifies Drought Stress in Sessile Oak Trees (Quercus petraea) by Altering Water Status and Physiological Responses. Water, 12(11), 2985. https://doi.org/10.3390/w12112985