Temporal Dynamics of Fish Assemblages as a Reflection of Policy Shift from Fishing Concession to Co-Management in One of the World’s Largest Tropical Flood Pulse Fisheries

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
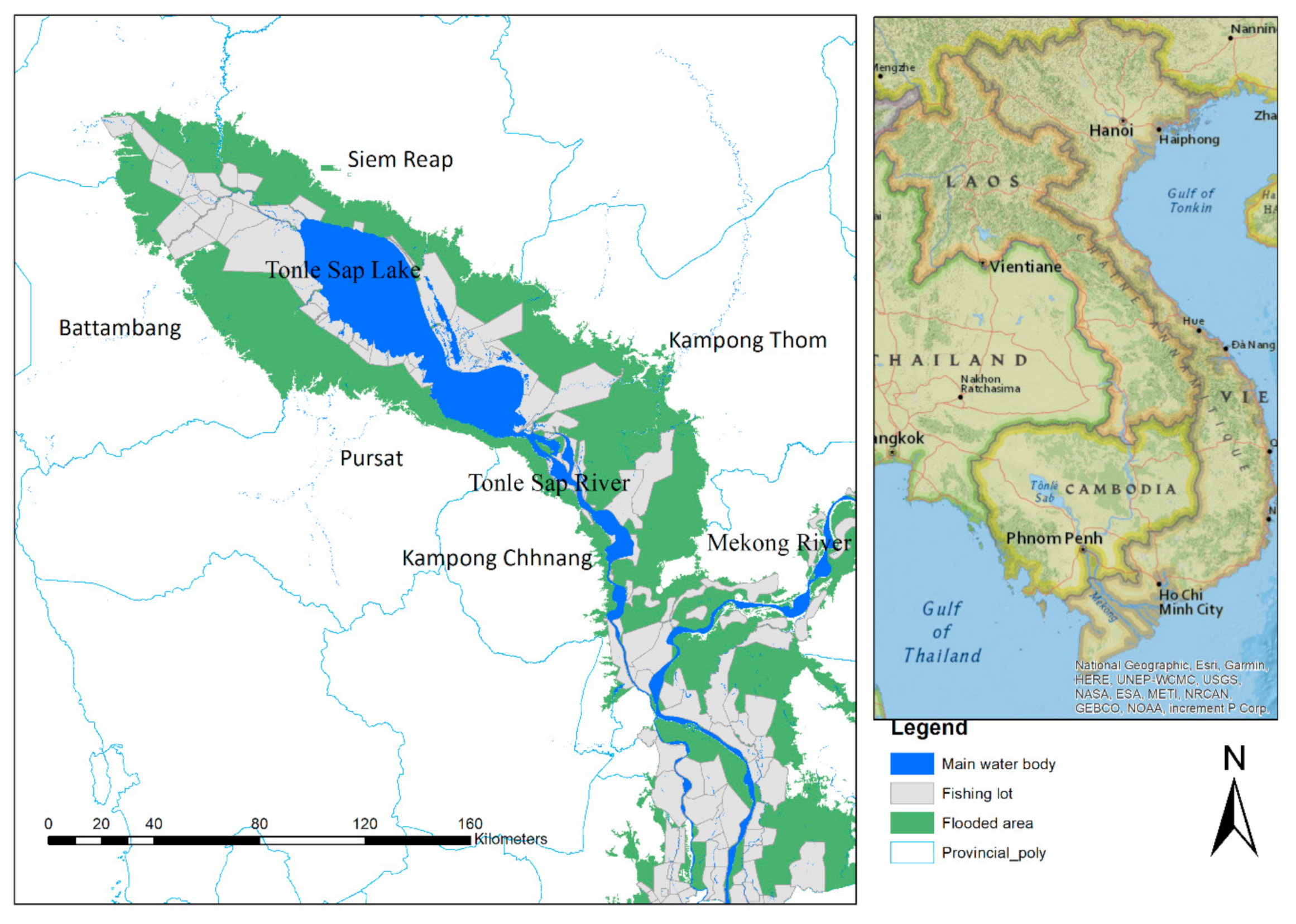
2.1. Sites Description
2.2. Data Collection
2.3. Data Preparation
2.4. Statistical Analyses
3. Results
3.1. Temporal Changes in Monthly Catch Weight
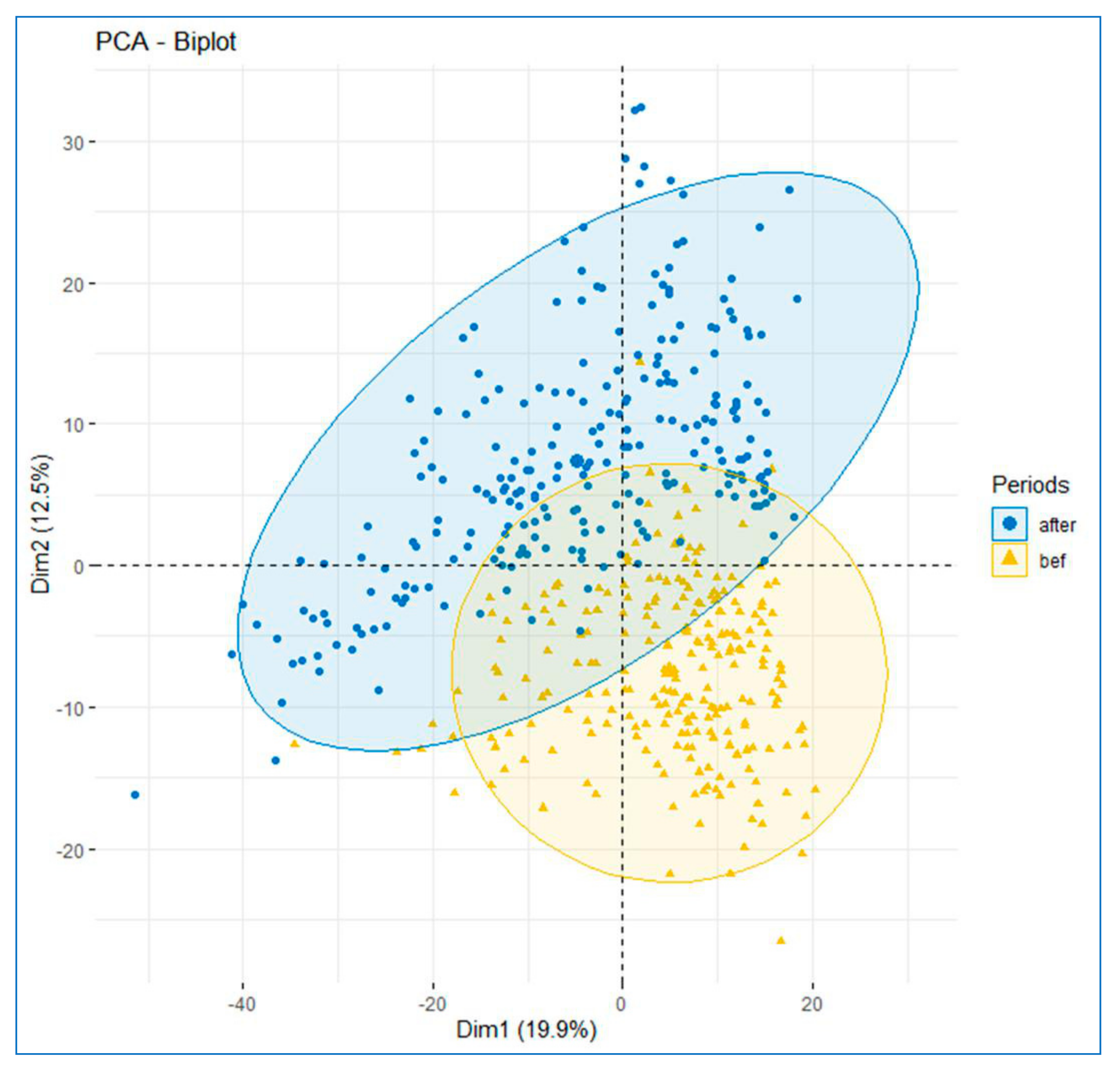
3.2. Temporal Changes in Assemblage Composition and Species’ Catch Weight
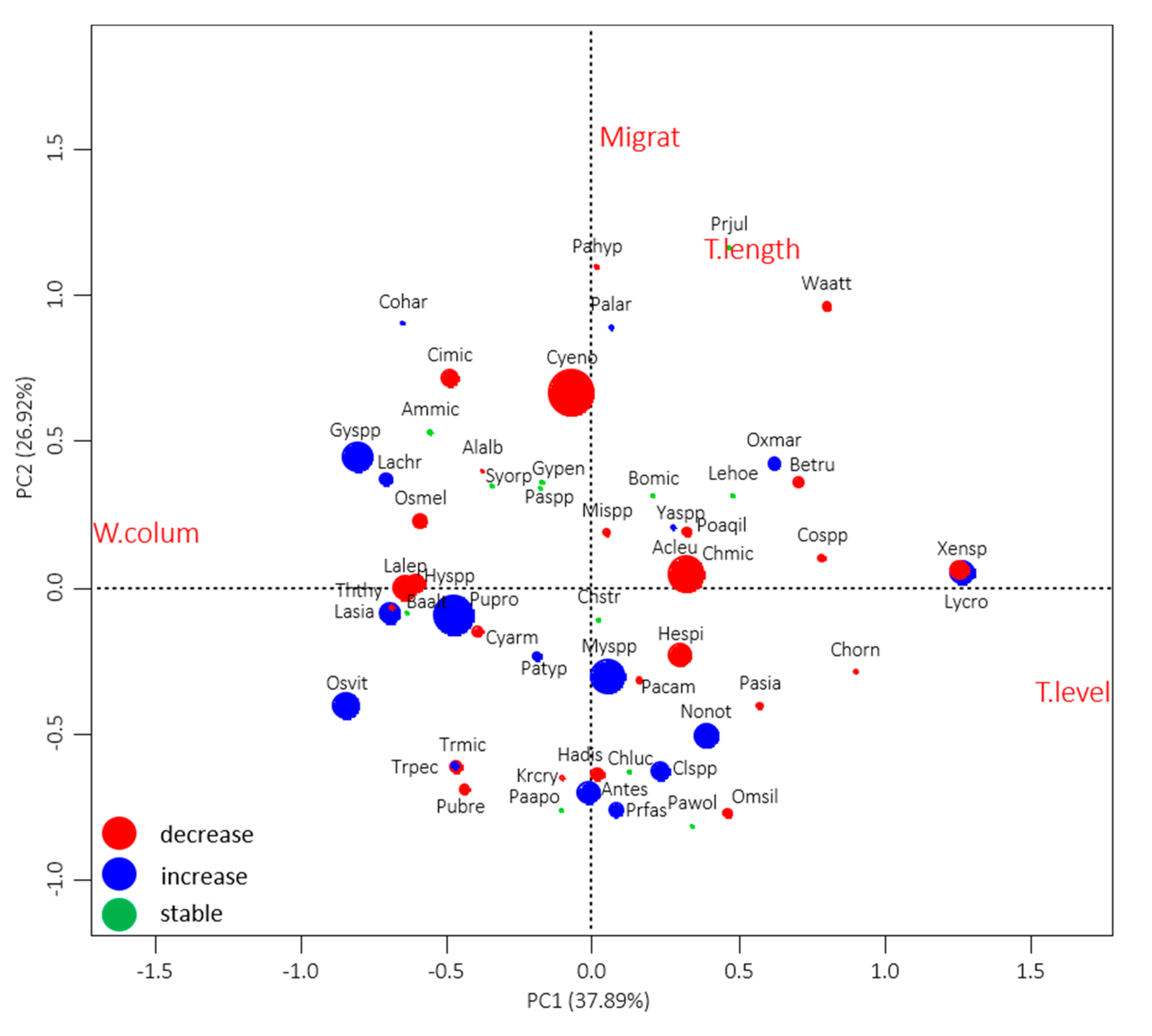
3.3. Relationship of Fish Species’ Status with Their Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mekong River Commission. Overview of the Hydrology of the Mekong Basin; Red Plough International Co. Ltd., Ed.; Mekong River Commission: Vientiane, Laos, 2005; p. 73. ISBN 17283248. [Google Scholar]
- Baran, E.; So, N.; Degen, P.; Chen, X.-Y.; Starr, P. Updated information on fish and fisheries in the Mekong Basin. Catch Cult. 2013, 19, 24–25. [Google Scholar]
- Hogan, Z.S.; Ngor, P.B.; Zalinge, N.V. Status and conservation of two endangered fish species, the Mekong giant catfish Pangasianodon gigas and the giant carp Catlocarpio siamensis, in Cambodia’s Tonle Sap River. Nat. Hist. Bull. Siam Soc. 2001, 49, 269–282. [Google Scholar]
- Ngor, P.B.; Grenouillet, G.; Phem, S.; So, N.; Lek, S. Spatial and temporal variation in fish community structure and diversity in the largest tropical flood-pulse system of South-East Asia. Ecol. Freshw. Fish 2018, 27, 1087–1100. [Google Scholar] [CrossRef]
- Baran, E. Fish Migration Triggers in the Lower Mekong Basin and Other Freshwater Tropical Systems; Mekong River Commission: Vientiane, Laos, 2006; p. 56. [Google Scholar]
- Poulsen, A.F.; Ouch, P.; Viravong, S.; Suntornratana, U.; Nguyen, T.T. Fish Migrations of the Lower Mekong River Basin: Implications for Development, Planning and Environmental Management; Mekong River Commission: Phnom Penh, Cambodia, 2002; p. 62. ISSN 1683-1489. [Google Scholar]
- Ngor, P.B.; McCann, K.S.; Grenouillet, G.; So, N.; McMeans, B.C.; Fraser, E.; Lek, S. Evidence of indiscriminate fishing effects in one of the world’s largest inland fisheries. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Ngor, P.B.; Oberdorff, T.; Phen, C.; Baehr, C.; Grenouillet, G.; Lek, S. Fish assemblage responses to flow seasonality and predictability in a tropical flood pulse system. Ecosphere 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Hortle, K.G.; Bamrungrach, P. Fisheries Habitat and Yield in the Lower Mekong Basin; Mekong River Commission: Phnom Penh, Cambodia, 2015; p. 80. ISSN 1683-1489. [Google Scholar]
- Zalinge, N.V.; Nao, T.; Touch, S.T. Where there is water, there is fish? Fisheries issues in the Lower Mekong Basin from a Cambodian perspective. In Proceedings of the Contribution to the Mekong Panel at the Seventh Common Property Conference of the International Association for the Study of Common Property, Vancouver, Canada, 10—14 June 1998. [Google Scholar]
- Vaidyanathan, G. Dam controversy: Remaking the Mekong. Nature 2011, 478, 305–307. [Google Scholar] [CrossRef]
- Allen, D.J.; Smith, K.G.; Darwall, W.R.T. The Status and Distribution of Freshwater Biodiversity in Indo-Burma; Complier, Ed.; IUCN: Cambridge, UK and Gland, Switzerland, 2012; ISBN 9782831714240. [Google Scholar]
- UNESCO Tonle Sap Biosphere Reserve. Available online: http://www.unesco.org/new/en/phnompenh/natural-sciences/biosphere-reserves/tonle-sap-biosphere-reserve/ (accessed on 12 January 2018).
- Degen, P.; Nao, T. Historical, cultural and legal perspectives on the fishing lot system in Cambodia. In Proceedings of the Common Property in the Mekong: Issues of Sustainability and Subsistence; Ahmed, M., Hirsch, P., Eds.; ICLARM Stud. Rev.: Phnom Penh, Cambodia, 2000; pp. 49–60. [Google Scholar]
- Lieng, S.; Yim, C.; Zalinge, N.P. Van Freshwater fisheries of Cambodia, I: The bagnet (Dai) fishery in the Tonle Sap River. Asian Fish. Sci. 1995, 8, 255–262. [Google Scholar]
- Degen, P.; Acker, F.V.; van Zalinge, N.V.; Nao, T.; Ly, V. Taken for granted: Conflicts over the Cambodia’s fresh water resources. In Proceedings of the Constituting the Commons: Crafting sustainable commons in the New Millennium, the Eighth Biennial Conference of the International Association for the Study of Common Property; The Internaitonal Association for the Study of the Commons (ASCP): Bloomington, IN, USA, 2000. [Google Scholar]
- Dina, T.; Sato, J. Is greater fishery access better for the poor? Explaining de-territorialisation of the Tonle Sap, Cambodia. J. Dev. Stud. 2014, 50, 962–976. [Google Scholar] [CrossRef]
- Ratner, B.D. Community management by decree? Lessons from Cambodia’s fisheries reform. Soc. Nat. Resour. 2006, 19, 79–86. [Google Scholar] [CrossRef]
- Cooperman, M.S.; So, N.; Arias, M.; Cochrane, T.A.; Elliott, V.; Hand, T.; Hannah, L.; Holtgrieve, G.W.; Kaufman, L.; Koning, A.A.; et al. A watershed moment for the Mekong: Newly announced community use and conservation areas for the Tonle Sap Lake may boost sustainability of the world’ s largest inland fishery. Cambodian J. Nat. Hist. 2012, 2012, 101–106. [Google Scholar]
- Hannah, L.; Costello, C.; Elliot, V.; Owashi, B.; Nam, S.; Oyanedel, R.; Chea, R.; Vibol, O.; Phen, C.; McDonald, G. Designing freshwater protected areas (FPAs) for indiscriminate fisheries. Ecol. Modell. 2019, 393, 127–134. [Google Scholar] [CrossRef]
- MRC. Cambodia abolishes fishing lots. Catch Cult. 2012, 18, 12–21. [Google Scholar]
- Chap, S.; Touch, P.; Diepart, J.-C. Fisheries Reforms and Right-based Fisheries: Insights from Community Fisheries across Cambodia; The Learning Institute: Phnom Penh, Cambodia, 2016. [Google Scholar]
- Oeur, I.; Kosal, M.; Sour, K.; Ratner, B.D. Innovations to Strengthen Aquatic Resource Governance on Cambodia’s Tonle Sap Lake. Rrogram Report; WorldFish: Penang, Malaysia, 2014. [Google Scholar]
- Van Acker, F. Cambodia’s Commons: Changing Governance, Shifting Entitlements? Centre for ASEAN Studies and Centre for International Management and Development Antwerp: Phnom Penh, Cambodia, 2003. [Google Scholar]
- Touch, S.; Todd, B. The Inland and Marine Fisheries Trade of Cambodia; Oxfam America: Phnom Penh, Cambodia, 2003. [Google Scholar]
- Degen, P.; Vuthy, L.; Thyda, P.C.; Dara, Y.; Piseth, C.; Saroeun, L. People, power and fishery policy: Fisheries management reform addressing community fisheries in Cambodia. In Proceedings of the 4th Technical Symposium on Mekong Fisheries; Inland Fisheries Research Development Institute of Cambodia & Mekong River Commission: Phnom Penh, Cambodia, 2002; pp. 11–28. [Google Scholar]
- Ngor, P.B. Informal aspects of fishing lot auctions in Cambodia: A case study in Prey Veng Province; A research paper submitted in partial fulfillment for the Degree of Master of Environmental Management and Development; The Australian National University: Canberra, Australia, 2007. [Google Scholar]
- KC, K.B.; Elliott, V.; Seng, R.; Pomeroy, R.S.; Schenkels, J.; Fraser, E.D.G. Evaluating community fishery management using Fishers’ perceptions in the Tonle Sap lake of Cambodia. Environ. Dev. 2020, 33, 100503. [Google Scholar] [CrossRef]
- Dina, T.; Sato, J. The cost of privatizing the commons: Overlapping property systems in Tonle Sap, Cambodia. Int. J. Commons 2015, 9, 261–280. [Google Scholar] [CrossRef]
- Ratner, B.D.; Mam, K.; Halpern, G. Collaborating for resilience: Conflict, collective action, and transformation on Cambodia’s Tonle Sap Lake. Ecol. Soc. 2014, 19, 31. [Google Scholar] [CrossRef]
- Ratner, B.D.; So, S.; Mam, K.; Oeur, I.; Kim, S. Conflict and collective action in Tonle Sap fisheries: Adapting governance to support community livelihoods. Nat. Resour. Forum 2017, 41, 71–82. [Google Scholar] [CrossRef]
- Say, S. Limited state and strong social forces: Fishing lot management in Cambodia. J. Southeast Asian Stud. 2014, 45, 174–193. [Google Scholar]
- Ngor, P.B.; Heng, K.; Thor, S.; Thomson, D. Pilot Participatory Policy Impact Assessment Fisheries Policy Related to Community Fisheries Development Stung Treng, Kampot/Kep, Koh Kong and Siem Reap Provinces; Fisheries Administration: Phnom Penh, Cambodia, 2010. [Google Scholar]
- Thay, S. Current perceptions of local fishers of community fisheries or co-management of fisheries: A case study. In Proceedings of the 4th Technical Symposium on Mekong Fisheries, 10–11 December 2001; MRC Conference Series No. 2. Hewitt, M.M., Ed.; Mekong River Commission: Phnom Penh, Cambodia, 2002; pp. 29–42. [Google Scholar]
- Teichert, N.; Lepage, M.; Sagouis, A.; Borja, A.; Chust, G.; Ferreira, M.T.; Pasquaud, S.; Schinegger, R.; Segurado, P.; Argillier, C. Functional redundancy and sensitivity of fish assemblages in European rivers, lakes and estuarine ecosystems. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Halls, A.S.; Paxton, B.R.; Hall, N.; Ngor, P.B.; Lieng, S.; Ngor, P.; So, N. The Stationary Trawl (Dai) Fishery of the Tonle Sap-Great Lake System, Cambodia; Mekong River Commission: Phnom Penh, Cambodia, 2013; p. 142. ISSN 1683-1489. [Google Scholar]
- MRC. State of the Basin Report: 2003. Executive Summary; Mekong River Commission: Phnom Penh, Cambodia, 2003; p. 50. ISSN 1728:3248. [Google Scholar]
- Lamberts, D. The Tonle Sap Lake as a productive ecosystem. Int. J. Water Resour. Dev. 2006, 22, 481–495. [Google Scholar] [CrossRef]
- Kummu, M.; Tes, S.; Yin, S.; Adamson, P.; Józsa, J.; Koponen, J.; Richey, J.; Sarkkula, J. Water balance analysis for the Tonle Sap Lake-floodplain system. Hydrol. Process. 2014, 28, 1722–1733. [Google Scholar] [CrossRef]
- Adamson, P.T.; Rutherfurd, I.D.; Peel, M.C.; Conlan, I.A. The hydrology of the Mekong River. In The Mekong Biophysical Environment of an International River Basin; Campbell, I.C., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 53–76. [Google Scholar]
- Kummu, M.; Penny, D.; Sarkkula, J.; Koponen, J. Sediment: Curse or blessing for Tonle Sap Lake? Ambio 2008, 37, 158–163. [Google Scholar] [CrossRef]
- Kummu, M.; Koponen, J.; Sarkkula, J. Modelling sediment and ecosystem in Tonle Sap Lake for impact assessment. Mater. Geoenvironment 2005, Volume 52, 87. [Google Scholar]
- Sarkkula, J.; Kiirikki, M.; Koponen, J.; Kummu, M. Ecosystem processes of the Tonle Sap Lake. In Proceedings of the 1st Workshop of Ecotone Phase II, Phnom Penh and Siem Reap, Cambodia, January 2003; pp. 1–14. [Google Scholar]
- Sarkkula, J.; Baran, E.; Chheng, P.; Keskinen, M.; Koponen, J.; Kummu, M. Tonle Sap pulsing system and fisheries productivity. In Proceedings of the XXIXe International Congress of Limnology, Lahti, Finland, 08–14 August 2004; pp. 1–5. [Google Scholar]
- MRC. Monitoring Fish Abundance and Diversity in the Lower Mekong Basin: Methodological Guidelines; Mekong River Commission: Phnom Penh, Cambodia, 2007. [Google Scholar]
- Ngor, P.B.; Hortle, K.G.; So, N. Standard Sampling Procedures for Fish Abundance and Diversity Monitoring in the Lower Mekong Basin; Mekong River Commission: Phnom Penh, Cambodia, 2016. [Google Scholar]
- Stamatopoulos, C. Sample-Based Fishery Surveys: A Technical Handbook; Food and Agriculture Oranization of the United Nations: Rome, Italy, 2002. [Google Scholar]
- Ly, S.; Van Zalinge, N.P. Catch Statistics of Cambodian Freshwater Fisheries 1994–1997, Report for Project: The Management of the Freshwater Capture Fisheries of Cambodia Phase 1; Mekong River Commission, Department of Fisheries and Danish International Development Agency: Phnom Penh, Cambodia, 1998. [Google Scholar]
- McCann, K.S.; Gellner, G.; McMeans, B.C.; Deenik, T.; Holtgrieve, G.; Rooney, N.; Hannah, L.; Cooperman, M.; Nam, S.; Giacomini, H. Food webs and the sustainability of indiscriminate fisheries 1. Can. J. Fish. Aquat. Sci. 2016, 665, 656–665. [Google Scholar] [CrossRef]
- Allan, J.D.; Abell, R.; Hogan, Z.; Revenga, C.; Taylor, B.W.; Welcomme, R.L.; Winemiller, K. Overfishing of inland waters. Bioscience 2005, 55, 1041–1051. [Google Scholar] [CrossRef]
- Rainboth, W.J. Fishes of the Cambodian Mekong; Food and Agriculture Organisation of the United Nations: Rome, Italy, 1996; ISBN 92-5-103743-4. [Google Scholar]
- MFD. Mekong Fish Database: A Taxonomic Fish Database for the Mekong Basin; Mekong River Commission: Phnom Penh, Cambodia, 2003. [Google Scholar]
- So, N.; Utsugi, K.; Shibukawa, K.; Thach, P.; Chhuoy, S.; Kim, S.; Chin, D.; Nen, P.; Chheng, P. Fishes of the Cambodian Freshwater Bodies; Inland Fisheries Research and Development Institute, Fisheries Administration: Phnom Penh, Cambodia, 2019; p. 197. [Google Scholar]
- Froese, R.; Pauly, D.; FishBase. World Wide Web electronic publication. Available online: www.fishbase.org (accessed on 10 February 2017).
- Poulsen, A.F.; Hortle, K.G.; Chan, S.; Chhuon, C.K.; Viravong, S.; Bouakhamvongsa, K.; Suntornratana, U.; Yoorong, N.; Nguyen, T.T.; Tran, B.Q. Distribution and Ecology of some Important Riverine Fish Species of the Mekong River Basin; Mekong River Commission: Phnom Penh, Cambodia, 2004; pp. 1–116. ISSN 1683-1489. [Google Scholar]
- Ngor, P.B.; Legendre, P.; Oberdorff, T.; Lek, S. Flow alterations by dams shaped fish assemblage dynamics in the complex Mekong-3S river system. Ecol. Indic. 2018, 88, 103–114. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. 2020. Available online: https://CRAN.R-project.org/package=factoextra.
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Fiendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Gavin, L.; Simpson, G.L.; et al. Vegan: Community Ecology Package. 2019; Available online: https://CRAN.R-project.org/package=vegan.
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- R Core Team R: A language and environment for statistical computing 2019, R version 1.2.1335; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/.
- Valbo-Jorgensen, J.; Poulsen, A.F. Using local knowledge as a research tool in the study of river fish biology: Experiences from the Mekong. Environ. Dev. Sustain. 2000, 2, 253–276. [Google Scholar] [CrossRef]
- Ngor, P.B. Dai fisheries in the Tonle Sap River of Phnom Penh and Kandal province (including a Review of the Census Data of 1996–1997). In Proceedings of the Management aspects of Cambodia’s Freshwater Capture Fisheries and Management Implications, Eleven Presentation Given at the Annual Meeting of the Department of Fisheries of the Ministry of Agriculture, Forestry and Fisheries, Phnom Penh, Cambodia., 27–27 January 2000. [Google Scholar]
- KC, K.B.; Bond, N.; Fraser, E.D.G.; Elliott, V.; Farrell, T.; McCann, K.; Rooney, N.; Bieg, C. Exploring tropical fisheries through fishers’ perceptions: Fishing down the food web in the Tonlé Sap, Cambodia. Fish. Manag. Ecol. 2017, 24, 452–459. [Google Scholar] [CrossRef]
- Deap, L.; Degen, P.; Zalinge, N.S. Fishing Gears of the Cambodian Mekong; Inland Fisheries Research and Development Institude of Cambodia (IFRdDI): Phnom Penh, Cambodia, 2003; p. 269. ISSN 1726-3972. [Google Scholar]
- Hardin, G. The tragedy of the commons. Science 1968, 162, 1243–1248. [Google Scholar]
- Sonneveld, B.; Thoto, F.; Houessou, D.; Wesenbeeck, L.V. The tragedy of the inland lakes. Int. J. Commons 2019, 13, 609–636. [Google Scholar] [CrossRef]
- Mahood, S.P.; Poole, C.M.; Watson, J.E.M.; Sharma, S.; Garnett, S.T.; Mackenzie, R.A. Agricultural intensification is causing rapid habitat change in the Tonle Sap Floodplain, Cambodia. Wetl. Ecol. Manag. 2020, 1. [Google Scholar] [CrossRef]
- Lohani, S.; Dilts, T.E.; Weisberg, P.J.; Null, S.E.; Hogan, Z.S. Rapidly accelerating deforestation in Cambodia’s Mekong River Basin: A comparative analysis of spatial patterns and drivers. Water 2020, 2191, 1–29. [Google Scholar]
- Daly, K.; Ahmad, S.K.; Bonnema, M.; Beveridge, C.; Hossain, F.; Nijssen, B.; Holtgrieve, G. Recent warming of Tonle Sap Lake, Cambodia: Implications for one of the world’s most productive inland fisheries. Lake Resevoirs 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V.; Guénette, S.; Pitcher, T.J.; Sumaila, U.R.; Walters, C.J.; Watson, R.; Zeller, D. Towards sustainability in world fisheries. Nature 2002, 418, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Welcomme, R.L.; Cowx, I.G.; Coates, D.; Béné, C.; Funge-Smith, S.; Halls, A.; Lorenzen, K. Inland capture fisheries. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2010, 365, 2881–2896. [Google Scholar] [CrossRef]
- Frank, K.T.; Petrie, B.; Choi, J.S.; Leggett, W.C. Trophic cascades in a formerly cod-dominated ecosystem. Science 2005, 308, 1621–1623. [Google Scholar] [CrossRef]
- Andersen, K.H.; Gislason, H. Unplanned ecological engineering. Proc. Natl. Acad. Sci. USA 2017, 114, 634–635. [Google Scholar] [CrossRef]
- CFDO. Policy Reform Impact Assessment, Cambodia: Impacts of the Fisheries Policy Reforms; Cambodian Department of Fisheries: Phnom Penh, Cambodia, 2004; p. 84. [Google Scholar]
- Hecht, J.S.; Lacombe, G.; Arias, M.E.; Dang, T.D.; Piman, T. Hydropower dams of the Mekong River basin: A review of their hydrological impacts. J. Hydrol. 2019, 568, 285–300. [Google Scholar] [CrossRef]
- Sokhem, P.; Sunada, K. The governance of the Tonle Sap Lake, Cambodia: Integration of local, national and international levels. Int. J. Water Resour. Dev. 2006, 22, 399–416. [Google Scholar] [CrossRef]
- Thay, S.; Schmidt, U. Aquatic Resources Management: The Tonle Sap Great Lake, Cambodia. In The International Conference on Sustainable Aquatic Resources are more than Managing Fish; The Eco-System Approach in Inland Fisheries and the Role of Intra-country Linkages: Penang, Malaysia, 2004. [Google Scholar]
- Kurien, J.; Kaing, K.; Pech, B. Cambodia: Towards a modern commons. Samudra Rep. 2016, 72, 15–19. [Google Scholar]
- MAFF. The Strategic Planning Framework for Fisheries: Update for 2015–2024 “Fishing for the Future”; Ministry of Agriculture, Forestry & Fisheries of the Royal Government of Cambodia: Phnom Penh, Cambodia, 2015; p. 52.
- Pokhrel, Y.; Burbano, M.; Roush, J.; Kang, H.; Sridhar, V.; Hyndman, D.W. A review of the integrated effects of changing climate, land use, and dams on Mekong river hydrology. Water 2018, 10, 266. [Google Scholar] [CrossRef]
- Baran, E.; Van Zalinge, N.P.; Ngor, P.B. Floods, floodplains and fish production in the Mekong Basin: Present and past trends. In Proceedings of the Second Asian Wetlands Symposium, Penang, Malaysia, 27–30 August 2001; Ali, A., Ed.; Penerbit Universiti Sains Malaysia: Pulau Pinang, Malaysia, 2001; pp. 920–932. [Google Scholar]
- Ziv, G.; Baran, E.; So, N.; Rodriguez-Iturbe, I.; Levin, S.A. Trading-off fish biodiversity, food security, and hydropower in the Mekong River Basin. Proc. Natl. Acad. Sci. USA 2012, 109, 5609–5614. [Google Scholar] [CrossRef] [PubMed]
- Sabo, J.L.; Ruhi, A.; Holtgrieve, G.W.; Elliott, V.; Arias, M.E.; Ngor, P.B.; Räsänen, T.A.; Nam, S. Designing river flows to improve food security futures in the Lower Mekong Basin. Science 2017, 358, eaao1053. [Google Scholar] [CrossRef]
- Winemiller, K.O.; McIntyre, P.B.; Castello, L.; Fluet-Chouinard, E.; Giarrizzo, T.; Nam, S.; Baird, I.G.; Darwall, W.; Lujan, N.K.; Harrison, I.; et al. Balancing hydropower and biodiversity in the Amazon, Congo, and Mekong. Science 2016, 351, 128–129. [Google Scholar] [CrossRef]
- Phomikong, P.; Fukushima, M.; Sricharoendham, B.; Nohara, S.; Jutagate, T. Diversity and community structure of fishes in the regulated versus unregulated tributaries of the mekong river. River Res. Appl. 2014, 31, 14. [Google Scholar] [CrossRef]
- Arias, M.E.; Cochrane, T.A.; Elliott, V. Modelling future changes of habitat and fauna in the Tonle Sap wetland of the Mekong. Environ. Conserv. 2013, 41, 165–175. [Google Scholar] [CrossRef]
- Daskalova, G.N.; Myers-smith, I.H.; Bjorkman, A.D.; Blowes, S.A. Landscape-scale forest loss as a catalyst of population and biodiversity change. Science 2020, 1347, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Ngor, P.B.; Lek, S.; McCann, K.S.; Hogan, Z.S. Dams threaten world’s largest inland fishery. Nature 2018, 563. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.E.; Holtgrieve, G.W.; Ngor, P.B.; Dang, T.D.; Piman, T. Maintaining perspective of ongoing environmental change in the Mekong floodplains. Curr. Opin. Environ. Sustain. 2019, 37, 1–7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Code | av. before | av. after | Cumsum | p-Value | Change |
|---|---|---|---|---|---|---|
| Gymnostomus spp. | Gyspp | 11.960 | 16.280 | 0.096 | 0.010 | + |
| Cyclocheilos enoplos | Cyeno | 13.397 | 2.791 | 0.181 | 0.010 | − |
| Puntioplites proctozystron | Pupro | 2.951 | 10.612 | 0.247 | 0.010 | + |
| Mystus spp. | Myspp | 3.575 | 8.897 | 0.301 | 0.010 | + |
| Channa micropeltes | Chmic | 7.044 | 0.696 | 0.351 | 0.010 | − |
| Trichopodus microlepis | Trmic | 5.163 | 4.168 | 0.398 | 0.010 | − |
| Osteochilus vittatus | Osvit | 4.503 | 7.930 | 0.445 | 0.010 | + |
| Hypsibarbus spp. | Hyspp | 6.358 | 4.640 | 0.483 | 0.010 | − |
| Channa striata | Chstr | 3.092 | 3.103 | 0.516 | 0.130 | N |
| Labiobarbus leptocheilus | Lalep | 3.991 | 0.939 | 0.547 | 0.010 | − |
| Hemibagrus spilopterus | Hespi | 4.172 | 1.743 | 0.578 | 0.010 | − |
| Notopterus notopterus | Nonot | 0.913 | 3.831 | 0.605 | 0.010 | + |
| Xenentodon sp. | Xensp | 0.358 | 3.284 | 0.631 | 0.010 | + |
| Anabas testudineus | Antes | 1.242 | 3.563 | 0.656 | 0.010 | + |
| Cyclocheilichthys armatus | Cyarm | 2.410 | 1.610 | 0.677 | 0.010 | − |
| Labeo chrysophekadion | Lachr | 1.653 | 2.760 | 0.696 | 0.010 | + |
| Labiobarbus siamensis | Lasia | 0.377 | 2.429 | 0.715 | 0.010 | + |
| Clarias spp. | Clspp | 0.516 | 2.233 | 0.733 | 0.010 | + |
| Thynnichthys thynnoides | Ththy | 1.894 | 1.543 | 0.749 | 0.010 | − |
| Osteochilus melanopleura | Osmel | 2.291 | 0.974 | 0.765 | 0.010 | − |
| Pangasianodon hypophthalmus | Pahyp | 1.528 | 1.355 | 0.780 | 0.010 | − |
| Paralaubuca typus | Patyp | 0.842 | 1.579 | 0.795 | 0.030 | + |
| Pristolepis fasciata | Prfas | 0.780 | 2.136 | 0.809 | 0.010 | + |
| Cirrhinus microlepis | Cimic | 1.872 | 0.304 | 0.823 | 0.010 | − |
| Parambassis wolffii | Pawol | 1.523 | 1.503 | 0.837 | 0.320 | N |
| Lycothrissa crocodilus | Lycro | 1.837 | 0.000 | 0.850 | 0.010 | − |
| Boesemania microlepis | Bomic | 0.748 | 0.974 | 0.860 | 0.260 | N |
| Hampala dispar | Hadis | 1.359 | 0.188 | 0.870 | 0.010 | − |
| Puntius brevis | Pubre | 1.136 | 0.243 | 0.879 | 0.010 | − |
| Coilia spp. | Cospp | 0.922 | 0.383 | 0.888 | 0.010 | − |
| Albulichthys albuloides | Alalb | 0.671 | 0.504 | 0.896 | 0.030 | − |
| Amblyrhynchichthys micracanthus | Ammic | 0.482 | 0.877 | 0.904 | 0.130 | N |
| Oxyeleotris marmorata | Oxmar | 0.043 | 1.077 | 0.912 | 0.010 | + |
| Belodontichthys truncatus | Betru | 0.966 | 0.145 | 0.919 | 0.010 | − |
| Pangasius larnaudii | Palar | 0.423 | 0.803 | 0.926 | 0.030 | + |
| Wallago attu | Waatt | 0.940 | 0.167 | 0.934 | 0.010 | − |
| Parachela siamensis | Pasia | 0.719 | 0.217 | 0.940 | 0.010 | − |
| Pangasius spp. | Paspp | 0.381 | 0.602 | 0.946 | 0.700 | N |
| Polynemus aquilonaris | Poaqi | 0.802 | 0.032 | 0.952 | 0.010 | − |
| Micronema spp. | Mispp | 0.639 | 0.105 | 0.957 | 0.010 | − |
| Ompok siluroides | Omsil | 0.634 | 0.010 | 0.961 | 0.010 | − |
| Systomus rubripinnis | Syorp | 0.368 | 0.267 | 0.966 | 0.100 | N |
| Chitala ornata | Chorn | 0.516 | 0.121 | 0.970 | 0.010 | − |
| Leptobarbus rubripinna | Lehoe | 0.279 | 0.429 | 0.974 | 0.140 | N |
| Trichopodus pectoralis | Trpec | 0.062 | 0.530 | 0.978 | 0.010 | + |
| Pao cambodgiensis | Pacam | 0.517 | 0.009 | 0.982 | 0.010 | − |
| Kryptopterus cryptopterus | Krcry | 0.462 | 0.028 | 0.986 | 0.010 | − |
| Cosmochilus harmandi | Cohar | 0.130 | 0.381 | 0.989 | 0.010 | + |
| Yasuhikotakia spp. | Yaspp | 0.067 | 0.397 | 0.992 | 0.010 | + |
| Probarbus jullieni | Prjul | 0.273 | 0.085 | 0.995 | 0.130 | N |
| Parambassis apogonoides | Paapo | 0.100 | 0.249 | 0.997 | 0.590 | N |
| Barbonymus altus | Baalt | 0.048 | 0.132 | 0.998 | 0.060 | N |
| Gyrinocheilus pennocki | Gypen | 0.002 | 0.133 | 0.999 | 0.070 | N |
| Achiroides leucorhynchos | Acleu | 0.066 | 0.010 | 1.000 | 0.010 | − |
| Channa lucius | Chluc | 0.002 | 0.002 | 1.000 | 0.710 | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, B.; Ngor, P.B.; Hogan, Z.S.; So, N.; Brosse, S.; Lek, S. Temporal Dynamics of Fish Assemblages as a Reflection of Policy Shift from Fishing Concession to Co-Management in One of the World’s Largest Tropical Flood Pulse Fisheries. Water 2020, 12, 2974. https://doi.org/10.3390/w12112974
Chan B, Ngor PB, Hogan ZS, So N, Brosse S, Lek S. Temporal Dynamics of Fish Assemblages as a Reflection of Policy Shift from Fishing Concession to Co-Management in One of the World’s Largest Tropical Flood Pulse Fisheries. Water. 2020; 12(11):2974. https://doi.org/10.3390/w12112974
Chicago/Turabian StyleChan, Bunyeth, Peng Bun Ngor, Zeb S. Hogan, Nam So, Sébastien Brosse, and Sovan Lek. 2020. "Temporal Dynamics of Fish Assemblages as a Reflection of Policy Shift from Fishing Concession to Co-Management in One of the World’s Largest Tropical Flood Pulse Fisheries" Water 12, no. 11: 2974. https://doi.org/10.3390/w12112974
APA StyleChan, B., Ngor, P. B., Hogan, Z. S., So, N., Brosse, S., & Lek, S. (2020). Temporal Dynamics of Fish Assemblages as a Reflection of Policy Shift from Fishing Concession to Co-Management in One of the World’s Largest Tropical Flood Pulse Fisheries. Water, 12(11), 2974. https://doi.org/10.3390/w12112974