Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland)


Abstract
1. Introduction
2. Materials and Methods
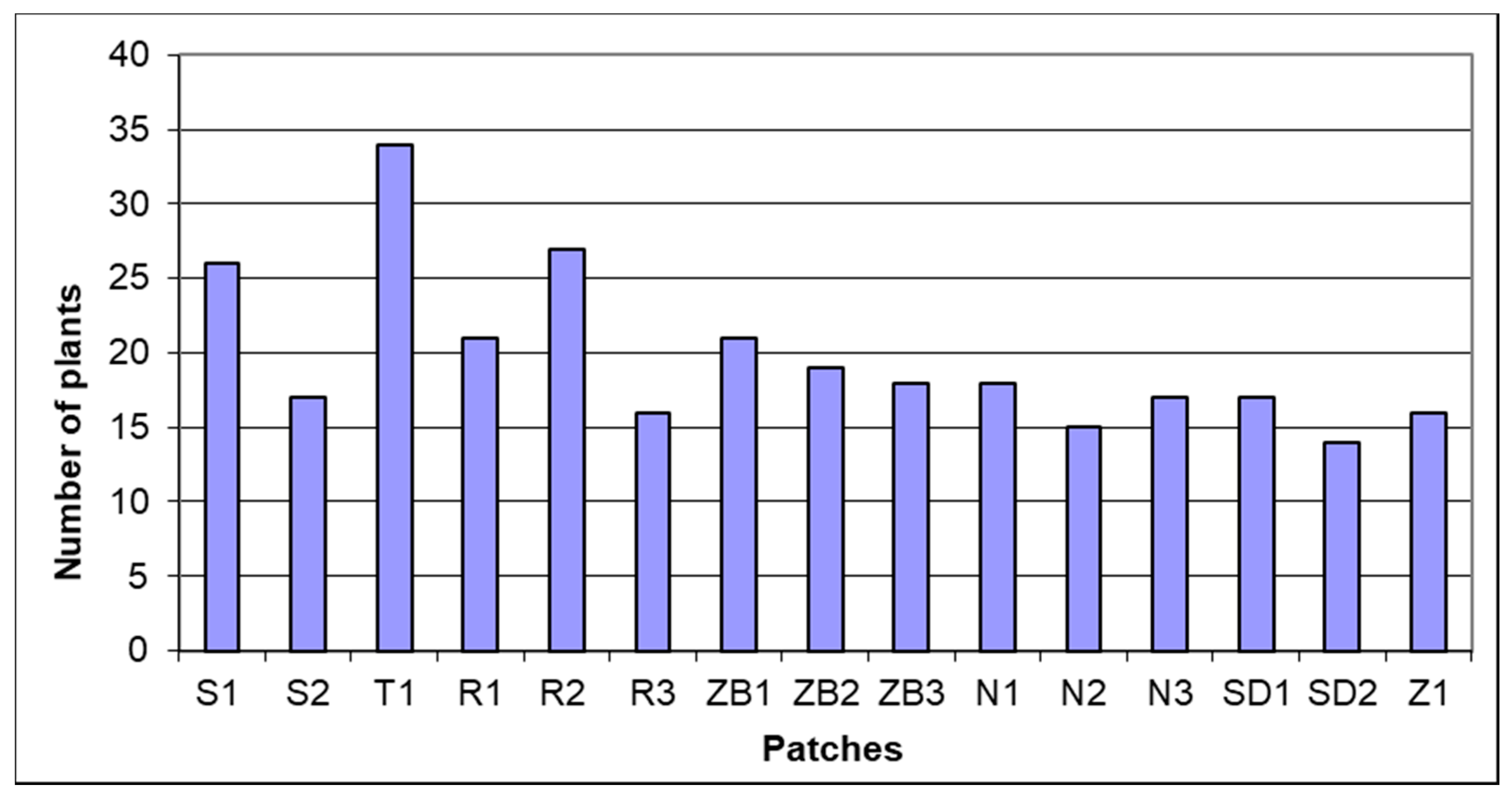
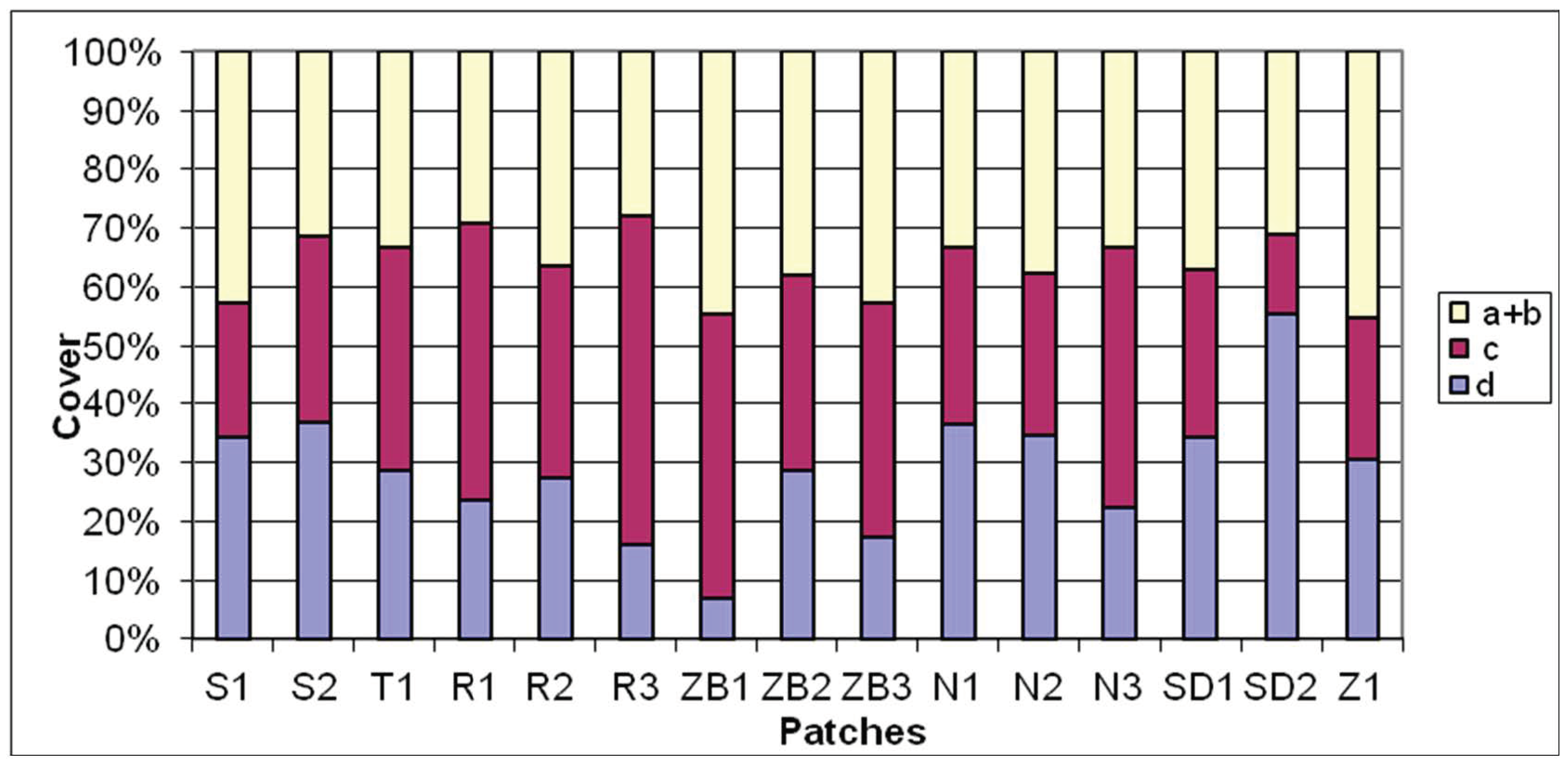
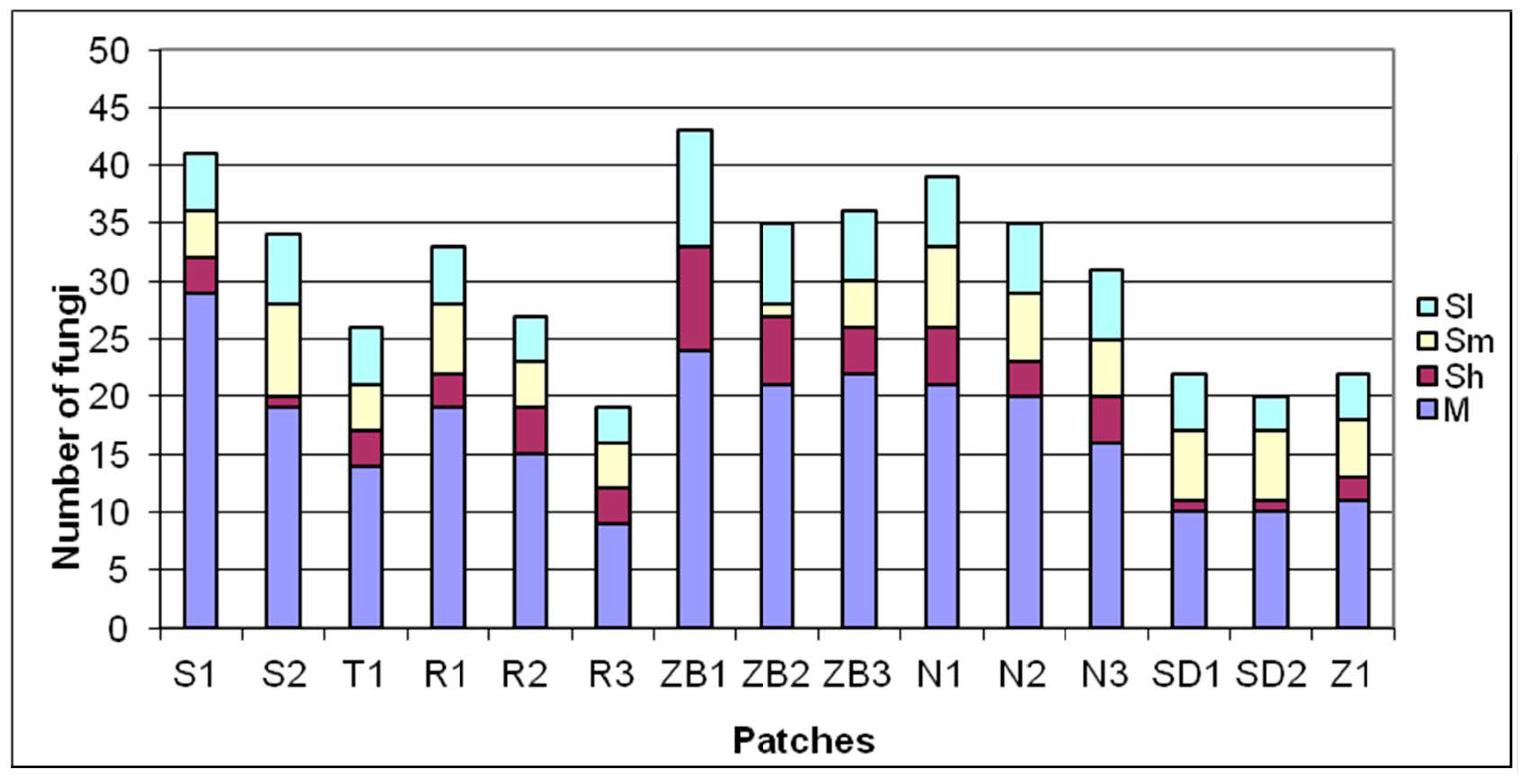
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Joosten, H.; Clarke, D. Wise Use of Mires and Peatlands; International Mire Conservation Group and International Peat Society: Saarijarvi, Finland, 2002. [Google Scholar]
- Dise, N.B.; Narasinha, J.S.; Weishampel, P.; Verma, S.B.; Verry, E.S.; Gorham, E.; Crill, P.M.; Harriss, R.C.; Kelley, C.A.; Yavitt, J.B.; et al. Carbon emissions from peatlands. In Peatland Biogeochemistry and Watershed Hydrology at the Marcell Experimental Forest; Kolka, R.K., Sebestyen, S.D., Verry, E.S., Brooks, K.N., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2011; pp. 297–347. [Google Scholar]
- Smith, L.C.; MacDonald, G.M.; Velichko, A.A.; Beilman, D.W.; Borisova, O.K.; Frey, K.E.; Kremenetski, K.V.; Sheng, Y. Siberian Peatlands a Net Carbon Sink and Global Methane Source Since Early Holocene. Science 2004, 303, 353–356. Available online: http://science.sciencemag.org/content/sci/303/5656/353 (accessed on 5 February 2019). [CrossRef] [PubMed]
- Franzén, L.G. Increased Decomposition of Subsurface Peat in Swedish Raised Bogs: Are Temperate Peatlands Still Net Sinks of Carbon? Mires Peat 2006, 1, 3. Available online: http://pixelrauschen.de/wbmp/media/map01/map_1_3.pdf accessed (accessed on 10 February 2019).
- Andersen, R.; Rochefort, L.; Poulin, M. Peat, Water and Plant Tissue Chemistry Monitoring: A Seven-Year Case-Study in a Restored Peatland. Wetlands 2010, 30, 159–170. Available online: https://link.springer.com/article/10.1007/s13157-009-0015-0 (accessed on 8 February 2019). [CrossRef]
- Minayeva, T.; Sirin, A.; Bragg, O. (Eds.) A Quick Scan of Peatlands in Central and Eastern Europe; Wetlands International: Wageningen, The Netherlands, 2009; 132p. [Google Scholar]
- Franzén, L.G.; Chen, D.; Klinger, L.F. Principles for a Climate Regulation Mechanism during the Late Phanerozoic era, Based on Carbon Fixation in Peat-Forming Wetlands. Ambio 1996, 25, 435–442. Available online: https://inis.iaea.org/search/search.aspx?orig_q=RN:28025142 (accessed on 9 February 2019).
- Strack, M. (Ed.) Peatlands and Climate Change; IPCC: Saarijärvi, Finland, 2008. [Google Scholar]
- Turunen, J. Development of FINNISH Peatland Area and Carbon Storage 1950–2000. Boreal Environ. Res. 2008, 13, 319–334. Available online: https://helda.helsinki.fi/bitstream/handle/10138/234741/ber13-4-319.pdf?sequence=1 (accessed on 9 February 2019).
- Sienkiewicz, J.; Kloss, M. Distribution and conservation of mires in Poland. In Regional Variation and Conservation of Mire Ecosystems; Moen, A., Ed.; NTNU University Museum, Gunneria: Trondheim, Norway, 1995; Volume 70, pp. 149–158. [Google Scholar]
- Tomassen, H.B.M.; Smolders, A.J.P.; Limpes, J.; Lamers, L.P.M.; Roelofs, J.G.M. Expansion of invasive species on ombrotrophic bogs: Desiccation or high N deposition? J. Appl. Ecol. 2004, 41, 139–150. [Google Scholar] [CrossRef]
- Kollmann, J.C.; Rasmussen, K.K. Succession of a degraded bog in NE Denmark over 164 years–monitoring one of the earliest restoration experiments. Tuexenia 2012, 32, 67–85. [Google Scholar]
- Sotek, Z. Distribution Paterns, History, and Dynamics of Peatland Vascular Plants in Pomerania (NW Poland). Biodiv. Res. Conserv. 2010, 18, 1–82. Available online: https://www.degruyter.com/downloadpdf/j/biorc.2010.18.issue--1/v10119-010-0020-4/v10119-010-0020-4.pdf (accessed on 8 February 2019).
- Herbichowa, M.; Pawlaczyk, P.; Stańko, R. Conservation of Baltic Raised Bogs in Pomerania. In Experience and Results of the LIFE04NAT/PL/000208 PLBALTBOGS Project; Wydawnictwo Klubu Przyrodników: Świebodzin, Poland, 2007. (In Polish) [Google Scholar]
- Smolders, A.J.P.; Lamers, L.P.M.; Lucassen, E.C.H.E.T.; Van der Velde, G.; Roelofs, J.G.M. Internal eutrophication: ‘how it works and what to do about it’, a review. Chem. Ecol. 2006, 22, 93–111. [Google Scholar] [CrossRef]
- Lamers, L.P.; Vile, M.A.; Grootjans, A.P.; Acreman, M.C.; van Diggelen, R.; Evans, M.G.; Richardson, C.J.; Rochefort, L.; Kooijman, A.M.; Roelofs, J.G. Ecological Restoration of Rich Fens in Europe and North America: From Trial and Error to an Evidence-Based Approach. Biol. Rev. 2014, 90, 182–203. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/brv.12102 (accessed on 8 February 2019). [CrossRef]
- Fay, E.; Lavoie, C. The Impact of Birch Seedlings on Evapotranspiration from a Mined Peatland: An Experimental Study in Southern Quebec, Canada. Mires Peat 2009, 5, 1–7. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.493.7184&rep=rep1&type=pdf (accessed on 8 February 2019).
- Talbot, J.; Richard, P.J.H.; Roulet, N.T.; Booth, R. KAssessing long-term hydrological and ecological responses to drainage in a raised bog using paleoecology and a hydrosequence. J. Veg. Sci. 2010, 21, 143–156. [Google Scholar] [CrossRef]
- Gunnarsson, U.; Malmer, N.; Rydin, H. Dynamics or constancy in Sphagnum dominated mire ecosystems? A 40-year study. Ecography 2002, 25, 685–704. [Google Scholar] [CrossRef]
- Directive, H. Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–15. [Google Scholar]
- Mucina, L.; Grabherr, G.; Wallnöver, S. Die Pflanzengesellschaften Österreichs; Wälder und Gebüsche. Gustav Fischer: Jena, Germany, 1993; Volume 3, pp. 1–350. [Google Scholar]
- Horodyská, E. Minerotrofní rašeliniště Českomoravské vrchoviny: Srovnání historických a současných dat; Ms., dipl. pr., PřF MU: Brno, Czech Republic, 2006. [Google Scholar]
- Dierßen, K. Vegetation Nordeuropas; Verlag Eugen Ulmer: Stuttgart, Germany, 1996. [Google Scholar]
- Oberdorfer, E. Süddeutsche Pflanzengesellschaften; Wälder und Gebüsche; Gustav Fischer: Jena, Germany, 1992; Volume 4, pp. 1–862. [Google Scholar]
- Preising, E.; Weber, H.E.; Vahle, H.-C. Die Pflanzengesellschaften Niedersachsens—Wälder und Gebüsche. Naturschutz Landschaftspflege Niedersachsen; Nds. Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz: Hannover, Germany, 2003; Volume 20, pp. 1–139. [Google Scholar]
- Kevey, B. Magyarország erdötársulásai. Tilia 2008, 14, 1–489. Available online: http://real-d.mtak.hu/152/1/Kevey.pdf (accessed on 5 February 2019).
- Kelly, D.L.; Iremonger, S.F. Irish wetland woods: The plant communities and their ecology. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 1997; Volume 97B, pp. 1–32. Available online: https://www.jstor.org/stable/20499982 (accessed on 7 February 2019).
- Michalko, J.; Berta, J.; Magic, D. Geobotanická mapa ČSSR; Slovenská socialistická republika: Veda, Bratislava, 1986. [Google Scholar]
- Didukh, Y.P.; Fitsailo, T.V.; Korotchenko, I.A.; Iakushenko, D.M.; Pashkevych, N.A. Biotopy lisovoï ta lïsostepovoï zon Ukraïny; TOV MAKROS: Kyïv, Ukraine, 2011. [Google Scholar]
- Herbichowa, M. Bory i lasy bagienne. B. Opis podtypów. Brzezina bagienna. In Lasy i bory. Poradnik ochrony siedlisk i gatunków Natura 2000—Podręcznik metodyczny; Herbich, J., Ed.; Ministerstwo Środowiska: Warszawa, Poland, 2004; Volume 5, pp. 175–178. (In Polish) [Google Scholar]
- Matuszkiewicz, W. A guide for marking Poland’s plant communities. In Biodiversity of Poland; Mirek, Z., Ed.; Inst. Bot. im. W. Szafera. PAN: Kraków, Poland, 2002; Volume 3, p. 537. (In Polish) [Google Scholar]
- Cross, J.R. Unusual stands of birch on bogs. Irish Nat. J. 1987, 22, 305–310. Available online: https://www.jstor.org/stable/25539176?seq=1#page_scan_tab_contents (accessed on 25 April 2019).
- Brzeg, A.; Kuświk, H.; Melosik, I.; Urbański, P. Flora and vegetation of the projected nature reserve “Torfowisko Toporzyk” in Drawski Landscape Park. Bad. Fizjogr. Pol. Zach. B 1995, 44, 51–76. (In Polish) [Google Scholar]
- Brzeg, A.; Kuświk, H.; Melosik, I.; Urbański, P. Flora and vegetation of the projected nature reserve “Zielone Bagna” in Drawski Landscape Park. Bad. Fizjogr. Pol. Zach. B 1996, 45, 121–145. (In Polish) [Google Scholar]
- Sotek, Z.; Stasińska, M. Diversity of macromycetes and the transformation of vegetation in the atlantic raised bog, Stramniczka”. Woda-Środowisko-Obszary Wiejskie 2010, 10, 257–270. (In Polish) [Google Scholar]
- Sotek, Z.; Grzejszczak, G.; Stasińska, M.; Malinowski, R. Synanthropisation of the Baltic-type raised bog “Roby” (NW Poland). Biodiv. Res. Conserv. 2015, 38, 51–56. [Google Scholar]
- Bešta, T.; Šafránková, J.; Pouzar, M.; Novák, J.; Nováková, K. Late Pleistocene—Early Holocene transition recorded in the sediments of a former shallow lake in the Czech Republic. Hydrobiologia 2009, 631, 107–120. [Google Scholar] [CrossRef]
- Gálová, A.; Hájková, P.; Petr, L.; Hájek, M.; Jamrichova, E. Origin of a boreal birch bog woodland and landscape development on a warm low mountain summit at the Carpathian-Pannonia interface. Holocene 2016, 26, 1112–1125. [Google Scholar] [CrossRef]
- Karpińska-Kołaczek, M.; Kołaczek, P.; Stachowicz-Rybka, R. Palaeobotanical studies on Late Glacial and Holocene vegetation development and transformations of the ‘Wielkie Błoto’ mire near Gołdap (north-eastern Poland). Acta Palaeobot. 2013, 53, 53–67. [Google Scholar] [CrossRef][Green Version]
- Karpińska-Kołaczek, M.; Kołaczek, P.; Stachowicz-Rybka, R. Pathways of woodland succession under low human impact during the last 13,000 years in northeastern Poland. Quat. Int. 2014, 328–329, 196–212. [Google Scholar] [CrossRef]
- Kołaczek, P.; Gałka, M.; Karpińska-Kołaczek, M. Succession of arboreal taxa during the Late Glacial in south-eastern Poland: Climatic implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 421, 1–14. [Google Scholar] [CrossRef]
- Filippova, N.V.; Bulyonkova, T.M. The diversity of larger fungi in the vicinities of Khanty-Mansiysk (middle taiga of West Siberia). Environ. Dyn. Glob. Clim. Chang. 2017, 8, 13–24. Available online: http://journals.eco-vector.com/EDGCC/article/view/6949 (accessed on 11 February 2019). [CrossRef]
- Stasińska, M. Macrofungi of raised and transitional bogs of Pomerania. Monogr. Bot. 2011, 101, 1–142. [Google Scholar] [CrossRef][Green Version]
- Thormann, M.N.; Rice, A.V. Fungi from Peatlands. Fungal Divers. 2007, 24, 241–299. Available online: http://www.cfs.nrcan.gc.ca/bookstore_pdfs/29484.pdf (accessed on 9 February 2019).
- Chinan, V. Macrofungi Identified in the Peat Bogs from the NATURA 2000 Site “Găina Lucina” (Eastern Carpathians, Romania). Analele ştiinţifice ale Universităţii “Al. I. Cuza” Iaşi. Biologie Vegetală 2010, 56, 61–64. Available online: http://www.bio.uaic.ro/publicatii/anale_vegetala/issue/2010F1/08-2010F1.pdf (accessed on 8 February 2019).
- Mapa topograficzna. 158. Czaplinek (Tempelburg) 1:100 000; Instytut Wojskowo-Geograficzny: Warszawa, Poland, 1921; Available online: http://maps.mapywig.org/m/WIG_maps/series/100K/158_CZAPLINEK_(Tempelburg)_1921_LoC_G6520_s100_.P6.jpg (accessed on 24 April 2019).
- Mapa topograficzna. 92. Treptow a. d. Rega (Trzebiatowo) 1:100 000. (Druk na podstawie mapy 1:100 000 wydania z r. 1929); Wojskowy Instytut Geograficzny: Warszawa, Poland, 1937; Available online: http://maps.mapywig.org/m/WIG_maps/series/100K_300dpi/P32_S20_92_TREPTOW_a.d._REGA_(TRZEBIATOWO)_1937_300dpi.jpg (accessed on 24 April 2019).
- Mapa topograficzna. 93. Kolberg (Kołobrzeg) 1:100 000. (Druk na podstawie mapy 1:100 000 wydania z r. 1929. Częściowo uzupełniono); Wojskowy Instytut Geograficzny: Warszawa, Poland, 1937; Available online: http://www.ziemiagryfa.org.pl/wp-content/uploads/2014/12/P32_S21_93_KOLBERG_KOLOBRZEG_1937_300dpi.jpg (accessed on 24 April 2019).
- Mapa topograficzna. 156. Naugard (Nowogard) 1:100 000. (Druk na podstawie mapy 1:100 000 wydania z r. 1929. Częściowo uzupełniono); Wojskowy Instytut Geograficzny: Warszawa, Poland, 1937; Available online: http://maps.mapywig.org/m/WIG_maps/series/100K_300dpi/P34_S20_156_NAUGARD_(NOWOGARD)_1936_300dpi.jpg (accessed on 24 April 2019).
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering plants and Pteridophytes of Poland—A checklist. In Biodiversity of Poland; Mirek, Z., Ed.; Inst. Bot. im. W. Szafera. PAN: Kraków, Poland, 2002; Volume 1, p. 442. [Google Scholar]
- Ochyra, R.; Żarnowiec, J.; Bednarek-Ochyra, H. Census catalogue of Polish mosses. In Biodiversity of Poland; Mirek, Z., Ed.; Inst. Bot. im. W. Szafera. PAN: Kraków, Poland, 2003; Volume 3, p. 372. [Google Scholar]
- Index Fungorum. Database 2016. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 15 February 2019).
- Hair, J.F.; Black, W.C.; Babin, B.J.; Anderson, R.E. Multivariate Data Analysis, 7th ed.; Pearson Education Limited: London, UK, 2014. [Google Scholar]
- Pawlaczyk, P.; Herbichowa, M.; Stańko, R. Protection of the Baltic Bogs: A Guide for Practitioners, Theorists and Officials; Wydawnictwo Klubu Przyrodnikow: Świebodzin, Poland, 2005. (In Polish) [Google Scholar]
- Salonen, V. Revegetation of harvested peat surfaces in relation to substrate quality. J. Veg. Sci. 1994, 5, 403–408. [Google Scholar] [CrossRef]
- Wind-Mulder, H.L.; Rochefort, L.; Vitt, D.H. Water and peat chemistry cutover of natural and post-harvested peatlands across Canada and their relevance to peatland restoration. Ecol. Eng. 1996, 7, 161–181. [Google Scholar] [CrossRef]
- Zając, E.; Zarzycki, J.; Ryczek, M. Substrate Quality and Spontaneous Revegetationof Extracted Peatland: Case Study of an Abandoned Polish Mountain Bog. Mires Peat 2018, 21, 1–14. Available online: https://www.researchgate.net/profile/Ewelina_Zajac/publication/326172848_Degradation_of_peat_surface_on_an_abandoned_post-extracted_bog_and_implications_for_re-vegetation/links/5b4c5d12a6fdccadaecf5c46/Degradation-of-peat-surface-on-an-abandoned-post-extracted-bog-and-implications-for-re-vegetation.pdf (accessed on 3 March 2019).
- Pawlaczyk, P. Bory i lasy bagienne. In Monitoring siedlisk przyrodniczych. Przewodnik metodyczny; Mróz, W., Ed.; Biblioteka Monitoringu Środowiska: Warszawa, Poland, 2010; Volume 1, pp. 216–235. [Google Scholar]
- Paavilainen, E.; Pälvänen, J. Peatland Foresty: Ecology and Principles; Ecological Studies 111; Springer: Berlin, Germany, 1995; pp. 1–256. [Google Scholar]
- Wieder, R.K.; Vitt, D.H. (Eds.) Boreal Peatland Ecosystems; Ecological Studies; Springer: Berlin/Heideberg, Germany, 2006; Volume 188, pp. 1–448. [Google Scholar]
- Aerts, R.; Ludwig, F. Water-table changes and nutritional status affect trace gas emissions from laboratory columns of peatland soils. Soil Biol. Biochem. 1997, 29, 1691–1698. [Google Scholar] [CrossRef]
- Gustavsen, H.G.; Päivänen, J. Luonnontilaisten soiden puustot kasvullisella metsämaalla 1950-Luvun alussa (Tree stands on virgin forested mires in the early 1950’s in Finland). Folia For. 1986, 673, 1–27. [Google Scholar]
- Hogg, P.; Squires, P.; Fitter, A.H. Acidification, nitrogen deposition and rapid vegetational change in a small valley mire in Yorkshire. Biol. Conserv. 1995, 71, 143–153. [Google Scholar] [CrossRef]
- Limpens, J. Prospects for Sphagnum Bogs Subject to High Nitrogen Deposition. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2003. Available online: https://library.wur.nl/WebQuery/wurpubs/fulltext/121392 (accessed on 5 March 2019).
- Hayward, P.M.; Clymo, R.S. The Growth of Sphagnum: Experiments on, and Simulation of, Some Effect of Light Flux and Water-Table Depth. J. Ecol. 1983, 71, 845–863. Available online: https://www.jstor.org/stable/2259597?read-now=1&seq=2#page_scan_tab_contents (accessed on 5 March 2019). [CrossRef]
- Heijmans, M.M.P.D.; Berendse, F.; Arp, W.J.; Masselink, A.K.; Klees, H.; De Visser, W.; Van Breemen, N. Effects of Elevated Carbon Dioxide and Increased Nitrogen Deposition on Bog Vegetation in the Netherlands. J. Ecol. 2001, 89, 268–279. [Google Scholar] [CrossRef]
- Tomassen, H.B.M.; Smolders, A.J.P.; Lamers, L.P.M.; Roelofs, J.G.M. Stimulated growth of Betula pubescens and Molinia caerulea on ombrotrophic bogs: Role of high levels of atmospheric nitrogen deposition. J. Ecol. 2003, 91, 357–370. [Google Scholar] [CrossRef]
- Takagi, K.; Tsuboya, T.; Takahashi, H.; Inoue, T. Effect of the invasion of vascular plants on heat and water balance in the Sarobetsu mire, Northern Japan. Wetlands 1999, 19, 246–254. [Google Scholar] [CrossRef]
- Arnolds, E. The analysis and classification of fungal communities with special reference to macrofungi. In Fungi in Vegetation Science; Winnterhoff, W., Ed.; Kluver Academic Publishers: Dordrecht, The Netherlands, 1992; pp. 7–47. [Google Scholar]
- Thormann, M.N. Diversity and function of fungi in peatlands: A carbon cycling perspective. Can. J. Soil Sci. 2006, 86, 281–293. [Google Scholar] [CrossRef]
- Thormann, M.N. The role of fungi in decomposition dynamics in peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Ecological Studies; Springer: Berlin, Heideberg, Germany, 2006; Volume 188, pp. 101–123. Available online: https://link.springer.com/content/pdf/10.1007%2F978-3-540-31913-9_6.pdf (accessed on 20 February 2019).
- Bujakiewicz, A. Share of macromycetes in plant communities found on the peat substrate in the Słowiński National Park. Poznań. Bad. Fizjogr. Pol. Zach. B 1986, 37, 101–129. (In Polish) [Google Scholar]
- Friedrich, S. Macromycetes in the plant associations of the Goleniowska Woods. Acta Mycol. 1986, 21, 43–76. (In Polish) [Google Scholar] [CrossRef][Green Version]
- Tedersoo, L.; Bahram, M.; Toots, M.; Diedhiou, A.G.; Henkel, T.W.; Kjoller, R.; Morris, M.H.; Nara, K.; Nouhra, E.; Peay, K.G.; et al. Towards global patterns in the diversity and community structure of ectomycorrhizal fungi. Mol. Ecol. 2012, 21, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Watling, R. Assessment of fungal diversity: Macromycetes, the problems. Can. J. Bot. 1995, 73 (Suppl. 1), S15–S24. [Google Scholar] [CrossRef]
- Ferris, R.; Peace, A.J.; Newton, A.C. Macrofungal communities of lowland Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karsten) plantations in England: Relationships with site factors and stand structure. For. Ecol. Manag. 2000, 131, 255–267. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Friesen, J.; Gamiet, S.; Kroeger, P. Epigeous Fruiting Bodies of Ectomycorrhizal Fungi as Indicators of Soil Fertility and Associated Nitrogen Status of Boreal Forests. Mycorrhiza 2009, 19, 535–548. Available online: https://link.springer.com/article/10.1007/s00572-009-0255-0 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Rudawska, M. Studies on the Factors Regulating the Mycorrhizal Symbiosis of Pine Seedlings; Instytut Dendrologii PAN: Kórnik, Poland, 1998; pp. 1–55. (In Polish) [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Baar, J.; Van Groenendael, J.M.; Roelofs, J.G.M. Are Ectomycorrhizal Fungi Associated with Alnus of Importance for Forest Development in Wet Environments? Plant Biol. 2000, 2, 505–511. Available online: https://www.thieme-connect.com/products/ejournals/html/10.1055/s-2000-7468 (accessed on 8 March 2019). [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. How Phosphorus Availability Affects the Impact of Nitrogen Deposition on Sphagnum and Vascular Plants in Bogs. Ecosystems 2004, 7, 793–804. Available online: https://link.springer.com/article/10.1007/s10021-004-0274-9 (accessed on 8 February 2019). [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bog Name | Geographical Coordinates | Area: Bog/Reserve (ha) | Dominant Communities | Kind of Protection |
|---|---|---|---|---|
| Roby | 54°6′33.6″ N 15°18′53.8″ E | 96.25/84.40 | Birch bog, willow thickets, community with Myrica gale and Erica tetralix, moss community, high sedge | Reserve; Natura 2000 PLH320017, PLB320010 |
| Stramniczka | 54°9′1.6″ N 15°41′25.2″ E | 94.49 | Boggy coniferous forests, birch bog, moss community | Reserve; Natura 2000 PLH320017 |
| Ziemomyśl | 53°11′33.7″ N 15°18′39.7″ E | 0.76 | Birch bog, community of Eriophorum vaginatum-Sphagnum fallax | – |
| Zielone Bagna | 53°40′23.7″ N 16°5′10.3″ E | 55.38 | Boggy coniferous forests, birch bog, willow thickets, moss community | Reserve; Natura 2000 PLH320039, PLB320019 |
| Mszar near Stara Dobrzyca * | 53°48′9.3″ N 15°31′58.0″ E | 11.17 | Boggy coniferous forests, birch bog, moss community | Reserve; Natura 2000 PLH320049 |
| Niewiadowo * | 53°39′25.20″ N 14°53′24.89″ E | 42 | Boggy coniferous forests, birch bog, moss community | Natura 2000 PLH320013 |
| Torfowisko Toporzyk | 53°42′40.2″ N 16°3′3.8″ E | 43.07 | Boggy coniferous forests, birch bog, moss community | Reserve; Natura 2000 PLB320019 |
| Bog Name | Starting Peat Extraction | Plot | Localization | Comments |
|---|---|---|---|---|
| Niewiadowo | before 1929 1 | N1 | middle part of the bog | secondary habitat—former peat extraction pit |
| N2 | middle part of the bog | habitat drying * | ||
| N3 | middle part of the bog | habitat drying *, secondary—former peat extraction pit | ||
| Roby | before 1929 2 | R1 | middle part of the bog | habitat drying *, about 30 m from the former peat extraction pit |
| R2 | middle part of the bog | habitat drying *, about 20 m from the former peat extraction pit | ||
| R3 | middle part of the bog | habitat drying * | ||
| Stramniczka | before 1929 3 | S1 | middle part of the bog | plot in the immediate vicinity of overgrowing former extraction with a moss community |
| S2 | middle part of the bog | secondary habitat—former peat extraction pit | ||
| Mszar near Stara Dobrzyca | – | SD1 | bog margin | plot on the bog, not drained and not exploited, in the depression of the area, natural habitat |
| SD2 | bog margin | plot on the bog, not drained and not exploited, less hydrated, natural habitat | ||
| Torfowisko Toporzyk | before 1921 4 | T1 | bog margin | plot in the depression of the area, exposed to surface runoff from the surrounding arable fields |
| Ziemomyśl | – | Z1 | bog margin | plot on the bog, not drained and not exploited, in the depression of the area, exposed to surface runoff from the surrounding arable fields |
| Zielone Bagna | before 1921 4 | ZB1 | middle part of the bog | secondary habitat—former peat extraction pit |
| ZB2 | middle part of the bog | secondary habitat—former peat extraction pit | ||
| ZB3 | middle part of the bog | habitat drying *, secondary—former peat extraction pit |
| Plot | N-NH4 mg/dm3 | N-NO3 mg/dm3 | N-NO2 mg/dm3 | P-PO4 mg/dm3 | pHH2O | pHKCl | Humidity % |
|---|---|---|---|---|---|---|---|
| Peat bog—Stramniczka | |||||||
| S1 | 3.25 abc | 2.63 ab | 0.062 abc | 1.60 ab | 4.16 cd | 2.95 ab | 86 abcd |
| S2 | 4.83 bcd | 2.41 ab | 0.058 abc | 1.36 a | 3.89 abc | 2.84 ab | 88 bcde |
| Peat bog—Zielone Bagno | |||||||
| ZB1 | 2.91 ab | 2.29 ab | 0.036 ab | 1.06 a | 4.02 bc | 3.01 ab | 90 de |
| ZB2 | 5.75 de | 2.50 ab | 0.054 ab | 1.07 a | 4.03 bc | 2.97 ab | 92 e |
| ZB3 | 3.23 abc | 2.03 a | 0.075 abc | 1.22 a | 3.78 abc | 2.74 ab | 84 abcd |
| Peat bog—Roby | |||||||
| R1 | 3.60 abcd | 3.83 abc | 0.088 abc | 1.34 a | 3.74 ab | 2.82 ab | 81 a |
| R2 | 3.64 abcd | 3.78 abc | 0.085 abc | 1.33 a | 3.70 ab | 2.81 ab | 81 a |
| R3 | 3.54 abcd | 3.70 abc | 0.078 abc | 1.33 a | 3.68 ab | 2.73 ab | 81 a |
| Peat bog—Niewiadowo | |||||||
| N1 | 3.62 abcd | 4.02 bc | 0.123 c | 2.61 bc | 4.42 de | 3.03 b | 90 de |
| N2 | 1.79 a | 2.34 ab | 0.025 a | 0.80 a | 3.60 a | 2.76 ab | 80 a |
| N3 | 5.37 cd | 5.24 c | 0.057 abc | 0.65 a | 3.77 ab | 2.67 a | 83 ab |
| Peat bog—Mszar near Stara Dobrzyca | |||||||
| SD1 | 4.22 bcd | 3.80 adc | 0.036 ab | 1.19 a | 4.60 e | 3.61 c | 92 e |
| SD2 | 2.94 ab | 2.07 a | 0.079 abc | 1.329 a | 4.49 de | 3.53 c | 84 abc |
| Peat bog—Toporzyk | |||||||
| T1 | 4.22 bcd | 3.61 abc | 0.034 ab | 1.22 a | 4.57 e | 3.67 c | 88 bcde |
| Peat bog—Ziemomyśl | |||||||
| Z1 | 7.74 e | 2.98 ab | 0.095 bc | 2.80 c | 4.03 bc | 3.03 b | 89 cde |
| Habitat | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Wet | 82.08 b | 3.82 b | 2.86 b | 3.28 | 0.070 | 3.44 b | 1.14 b |
| Wetter | 89.51 a | 4.21 a | 3.14 a | 3.03 | 0.077 | 4.57 a | 1.61 a |
| Parameter | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Surface runoff | 88.87 a | 4.30 a | 3.35 b | 3.30 | 0.064 | 5.60 a | 2.01 a |
| Natural | 88.12 a | 4.54 a | 3.57 a | 2.93 | 0.058 | 3.58 b | 1.26 b |
| Without surface runoff | 85.15 b | 3.88 b | 2.84 c | 3.15 | 0.080 | 3.77 b | 1.31 b |
| Plot Location | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Peat extraction pits | 87.59 a | 4.01 b | 2.89 b | 3.01 | 0.083 | 4.14 ab | 1.37 |
| Bog margin | 88.50 a | 4.22 a | 3.46 a | 3.11 | 0.061 | 4.78 a | 1.63 |
| Middle part of the bog | 80.89 b | 3.68 c | 2.77 b | 3.41 | 0.069 | 3.14 b | 1.20 |
| Parameter | Humidity % | pHH2O | pHKCl | N-NO3 mg/dm3 | N-NO2 mg/dm3 | N-NH4 mg/dm3 | P-PO4 mg/dm3 |
|---|---|---|---|---|---|---|---|
| Mid-field bogs | 85.86 | 3.96 b | 2.96 | 2.97 | 0.066 | 4.27 | 1.43 |
| Mid-forest bogs | 86.14 | 4.18 a | 3.12 | 3.49 | 0.087 | 3.59 | 1.31 |
| Parameter | Humidity | pHH2O | pHKCl | P-PO4 | N-NH4 | N-NO3 | N-NO2 |
|---|---|---|---|---|---|---|---|
| NP | 0.60 * | 0.31 | 0.29 | 0.33 | 0.35 | 0.35 | 0.35 |
| Vac-Pic | 0.52 * | 0.49 | 0.82 * | 0.73 * | 0.72 * | 0.68 * | 0.72 * |
| Ox-Sph | −0.42 | −0.30 | −0.53 * | −0.44 | −0.48 | −0.43 | −0.48 |
| a + b | 0.56 * | 0.51 * | 0.42 | 0.45 | 0.47 | 0.47 | 0.47 |
| c | 0.54 * | −0.12 | 0.24 | 0.21 | 0.25 | 0.20 | 0.25 |
| d | 0.19 | 0.56 * | −0.17 | 0.02 | 0.09 | 0.18 | 0.09 |
| NF | 0.57 * | 0.48 | 0.94 * | 0.83 * | 0.80 * | 0.73 * | 0.80 * |
| M | 0.64 * | 0.67 * | 0.92 * | 0.92 * | 0.84 * | 0.83 * | 0.84 * |
| Sh | 0.44 | 0.19 | 0.73 * | 0.58 * | 0.58 * | 0.48 | 0.58 * |
| Sm | −0.24 | 0.01 | −0.41 | −0.26 | −0.23 | −0.14 | −0.23 |
| Sl | 0.50 | 0.39 | 0.82 * | 0.73 * | 0.73 * | 0.67 * | 0.73 * |
| Parameter | NP | Vac-Pic | Ox-Sph | a + b | c | d |
|---|---|---|---|---|---|---|
| NF | 0.21 | 0.87 * | −0.60 * | 0.34 | 0.25 | −0.43 |
| M | 0.24 | 0.67 * | −0.30 | 0.37 | 0.11 | −0.14 |
| Sh | 0.19 | 0.72 * | −0.61 * | 0.22 | 0.43 | −0.59 * |
| Sm | −0.31 | −0.49 | 0.44 | −0.40 | −0.39 | 0.50 |
| Sl | 0.05 | 0.78 * | −0.46 | 0.26 | 0.27 | −0.42 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotek, Z.; Stasińska, M.; Malinowski, R.; Gamrat, R.; Gałczyńska, M. Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland). Water 2019, 11, 1224. https://doi.org/10.3390/w11061224
Sotek Z, Stasińska M, Malinowski R, Gamrat R, Gałczyńska M. Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland). Water. 2019; 11(6):1224. https://doi.org/10.3390/w11061224
Chicago/Turabian StyleSotek, Zofia, Małgorzata Stasińska, Ryszard Malinowski, Renata Gamrat, and Małgorzata Gałczyńska. 2019. "Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland)" Water 11, no. 6: 1224. https://doi.org/10.3390/w11061224
APA StyleSotek, Z., Stasińska, M., Malinowski, R., Gamrat, R., & Gałczyńska, M. (2019). Birch Bog on Anthropogenically Transformed Raised Bogs. A Case Study from Pomerania (Poland). Water, 11(6), 1224. https://doi.org/10.3390/w11061224