Streams with Riparian Forest Buffers versus Impoundments Differ in Discharge and DOM Characteristics for Pasture Catchments in Southern Amazonia

 ,
,  , , , and
, , , and
Abstract
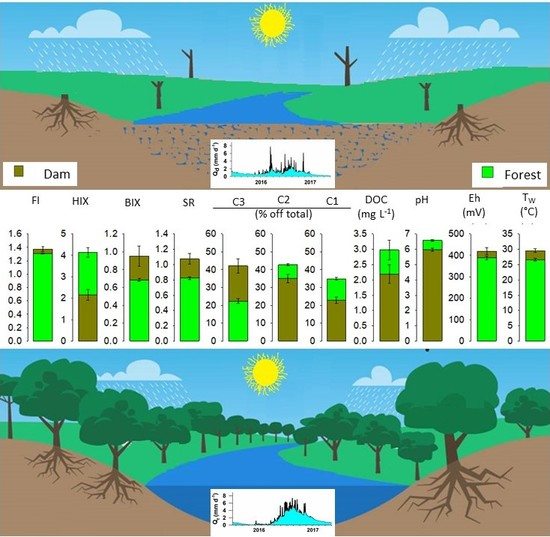
:
1. Introduction
2. Materials and Methods
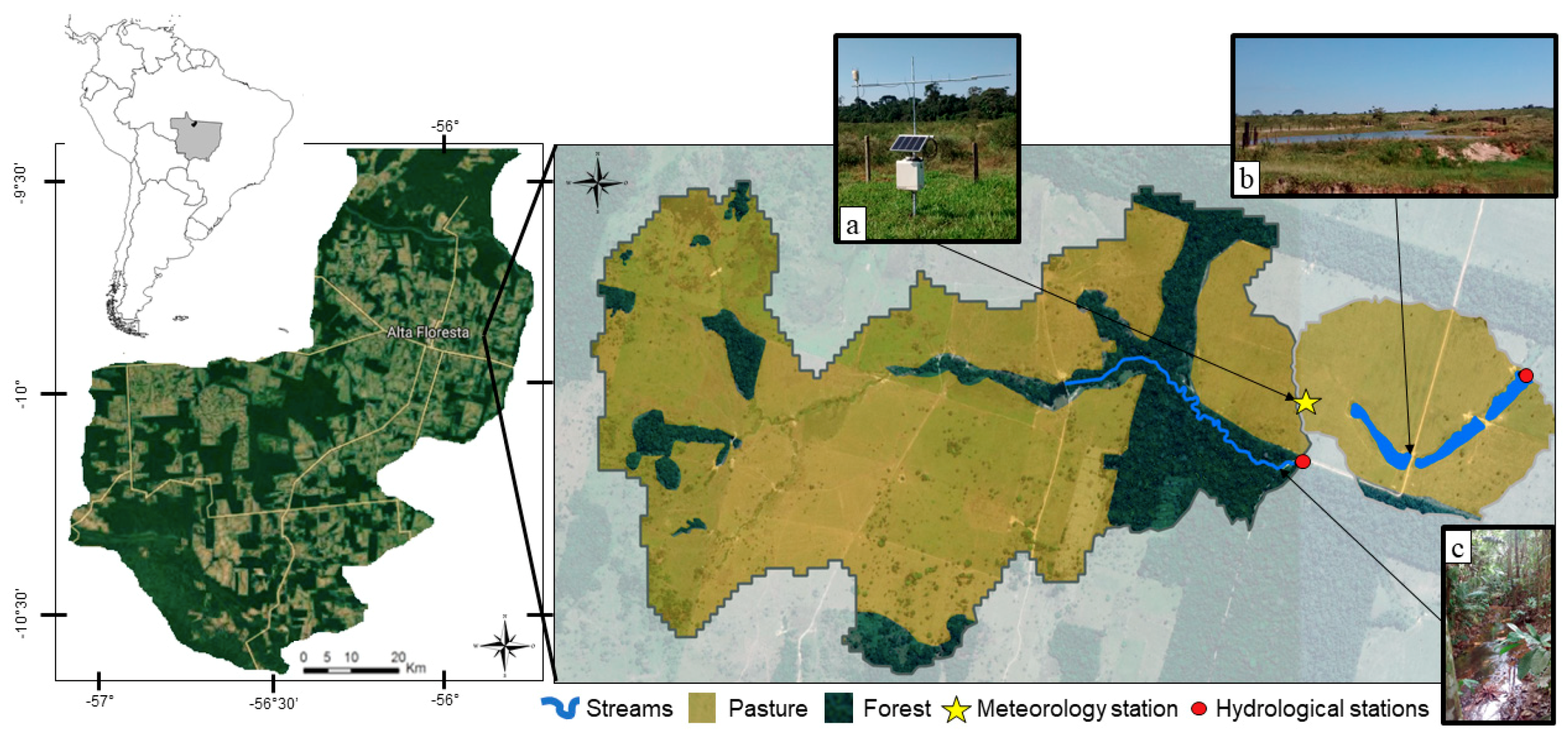
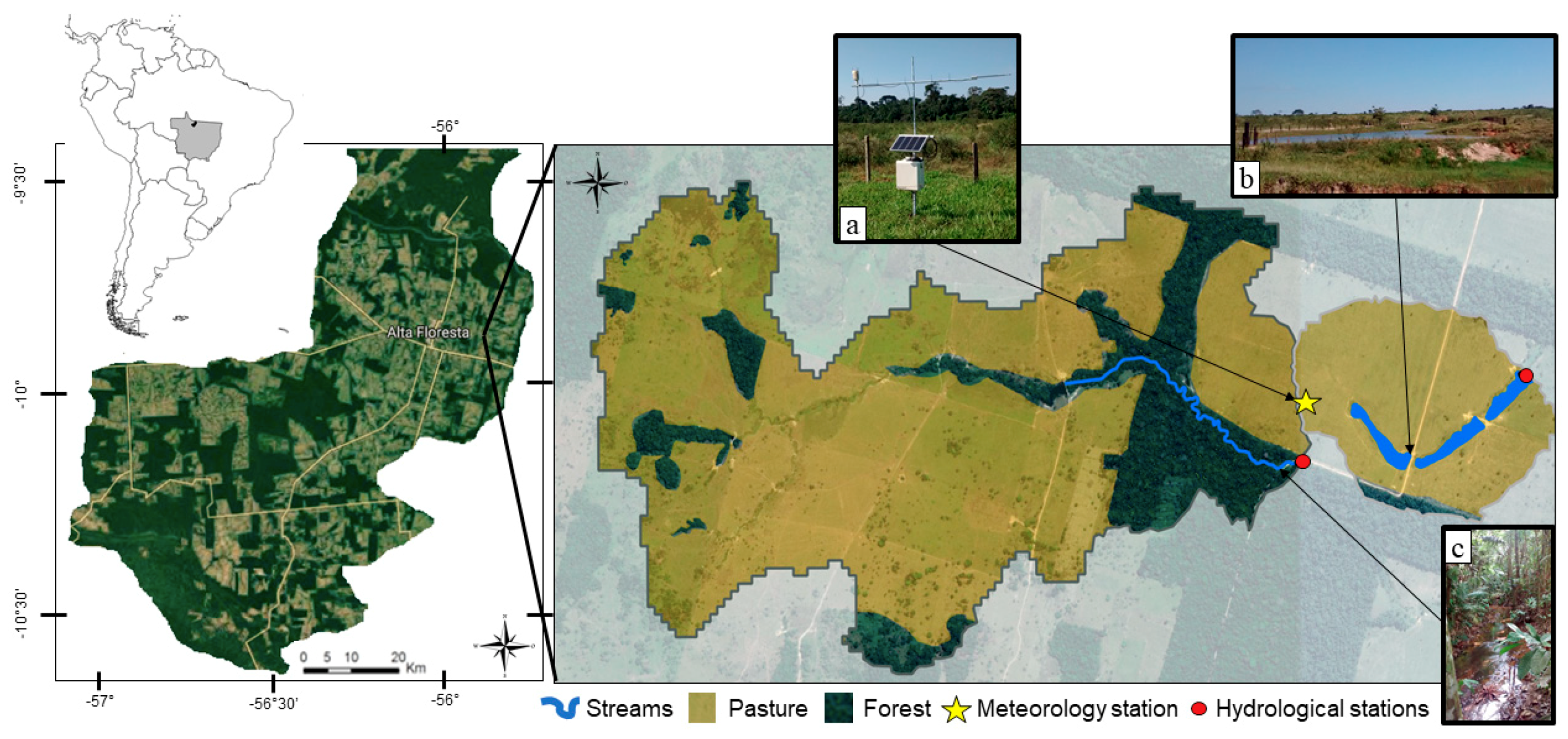
2.1. Site Description and Headwater Watersheds
2.2. Precipitation and Discharge Measurements
2.3. Water Sampling and Processing and Environment Variables Measurements
2.4. Measurements of DOC and Characterization of DOM Compositions
2.5. EEMs Pre-Processing and Parallel Factor Analysis (PARAFAC)
2.6. Optical Indices Calculations
2.7. Statistical Analyses
3. Results
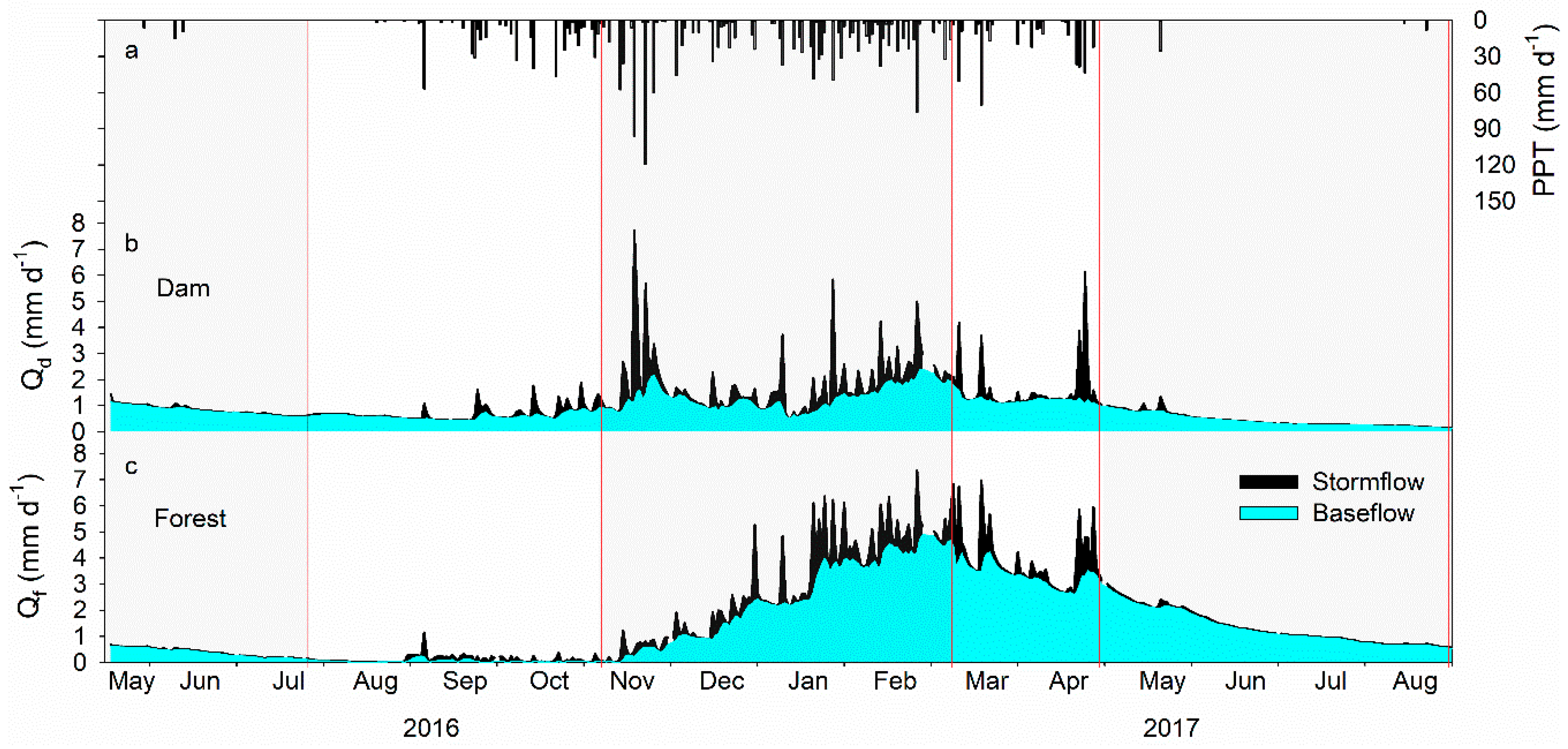
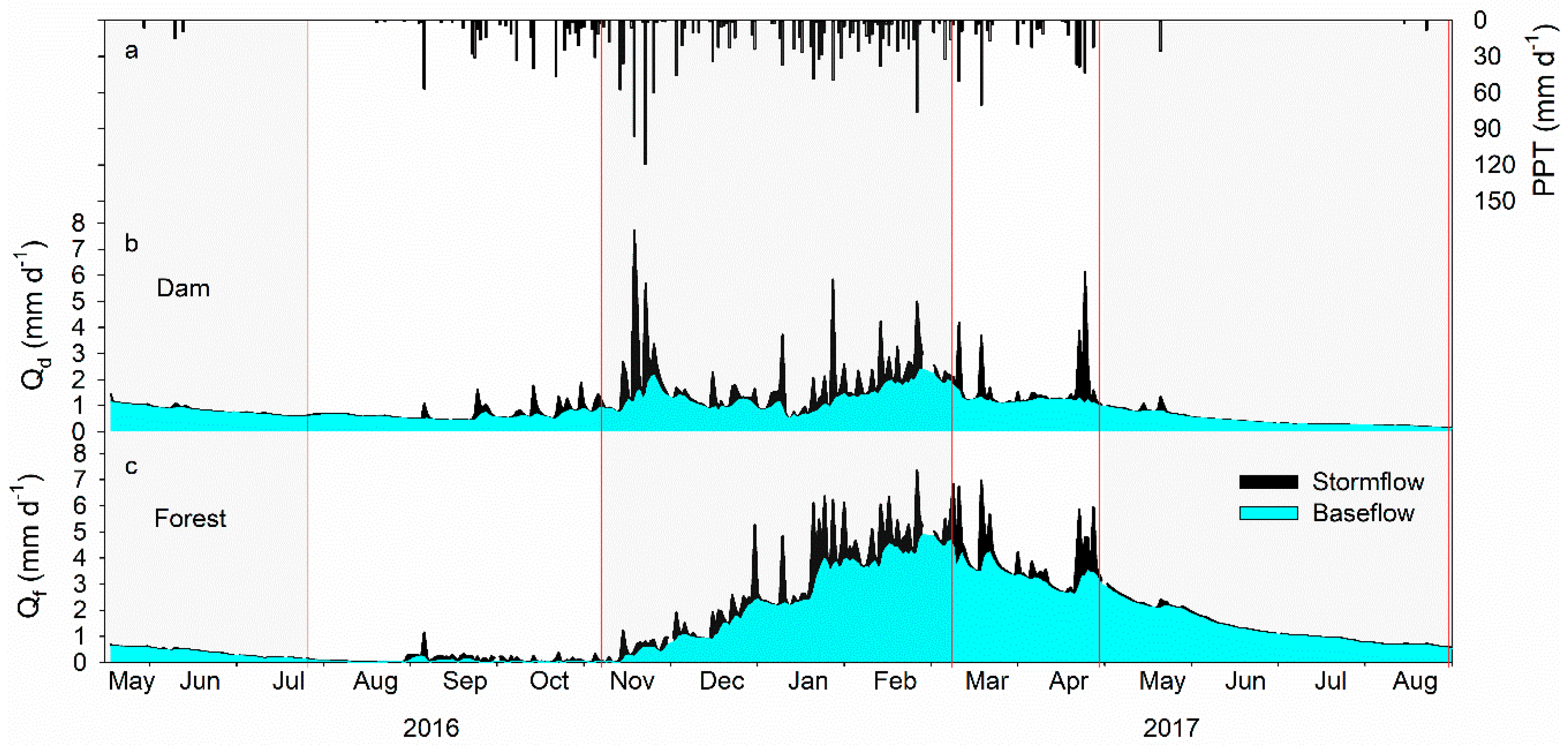
3.1. Hydrograph, Precipitation, and Stream Discharge
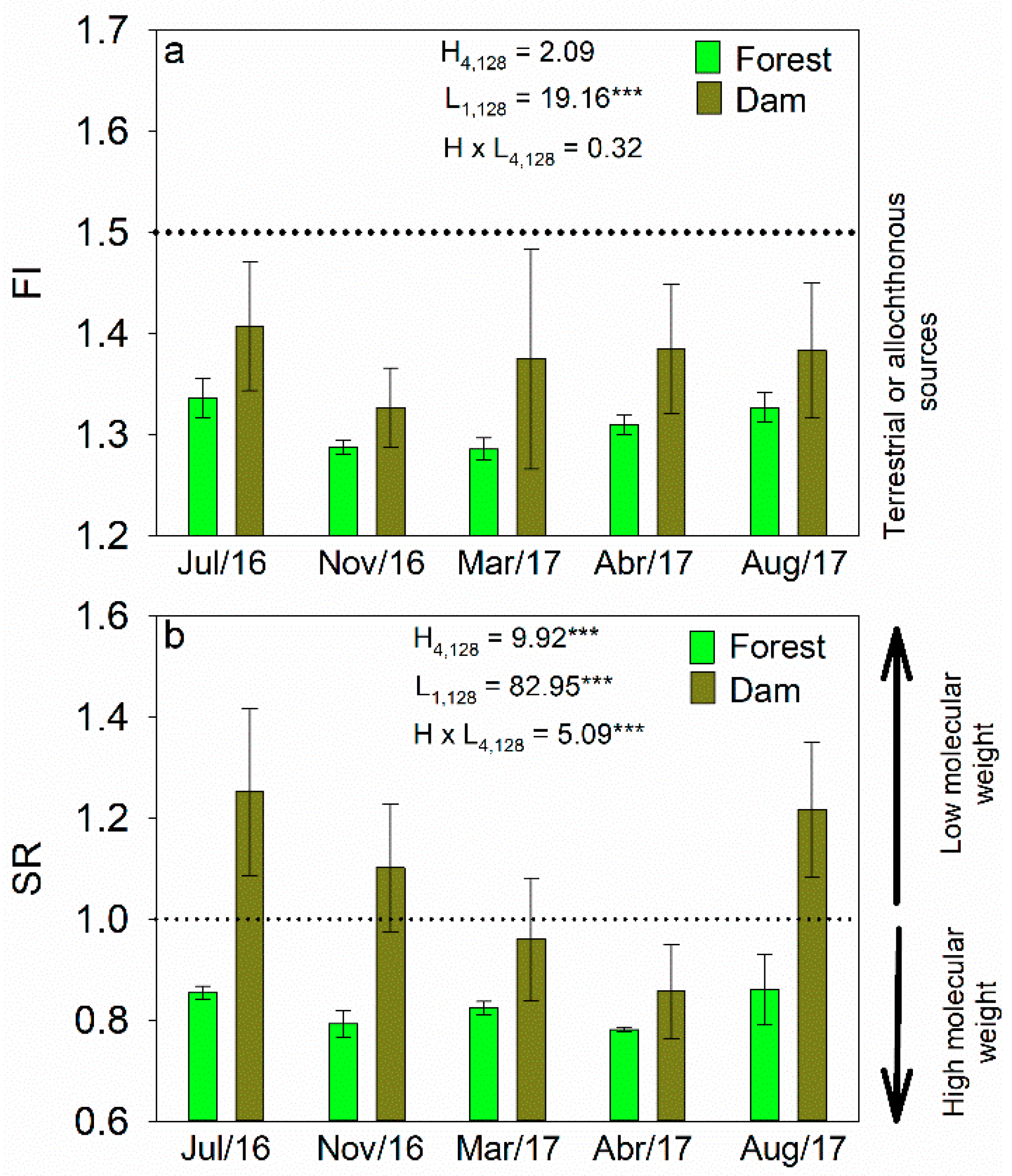
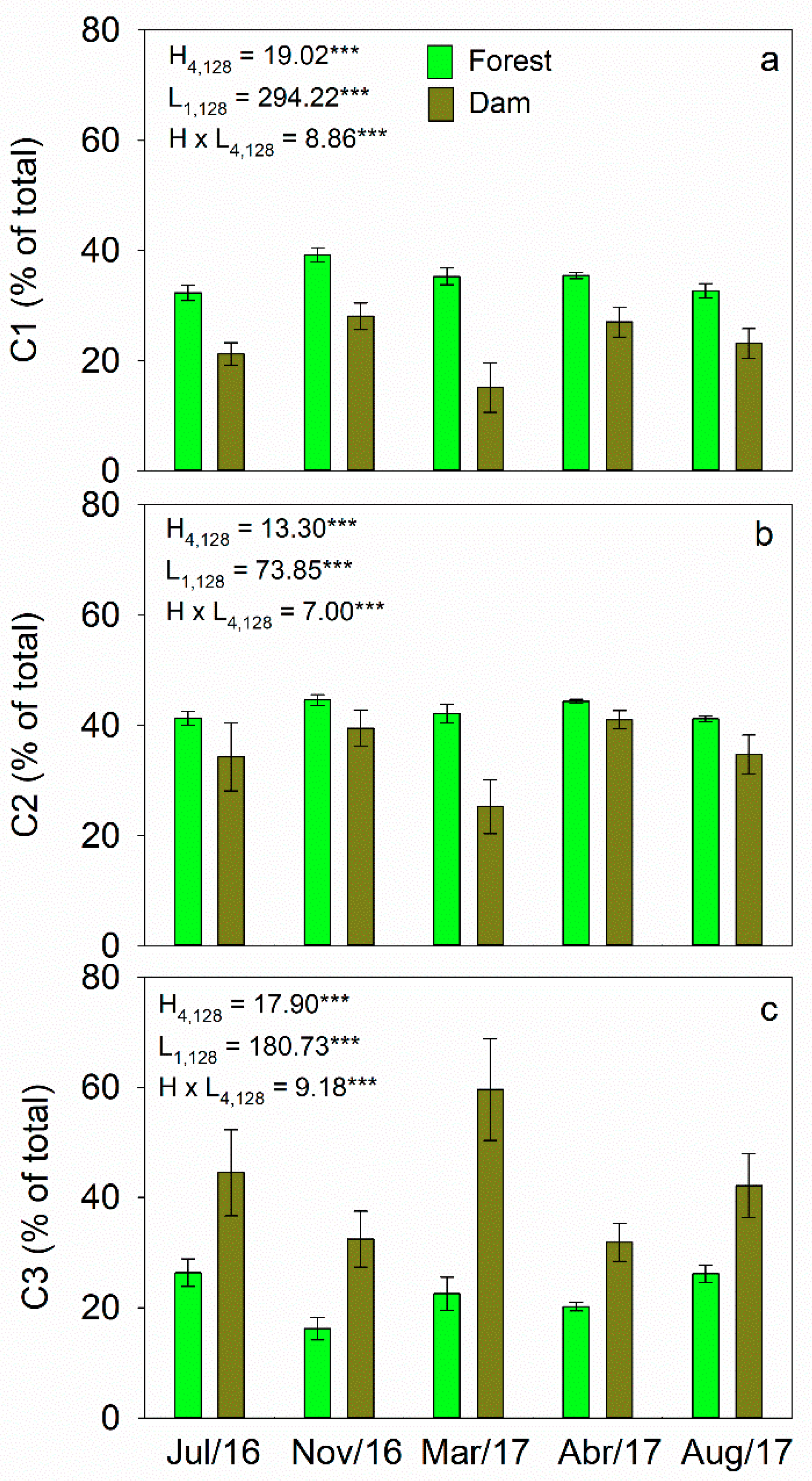
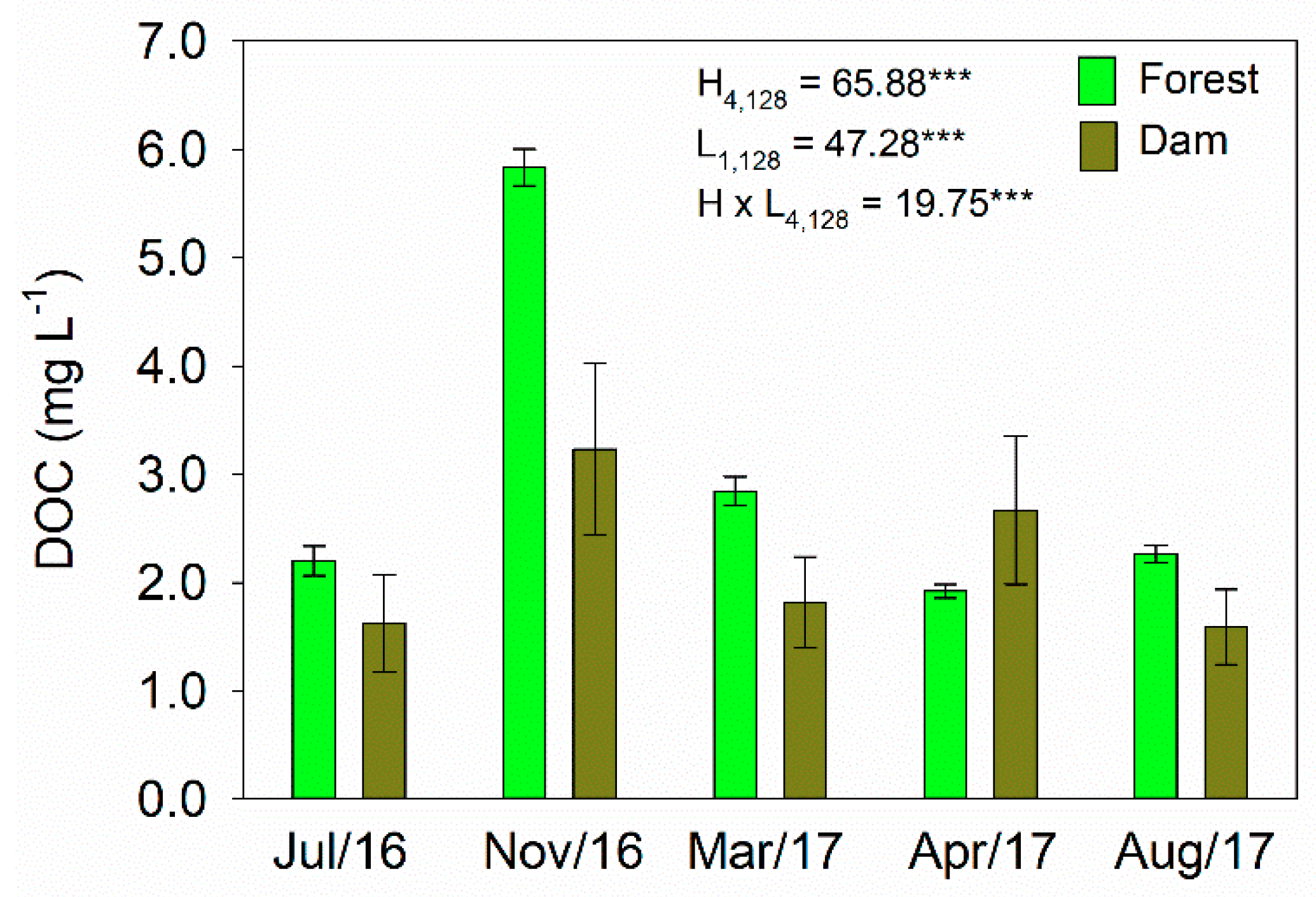
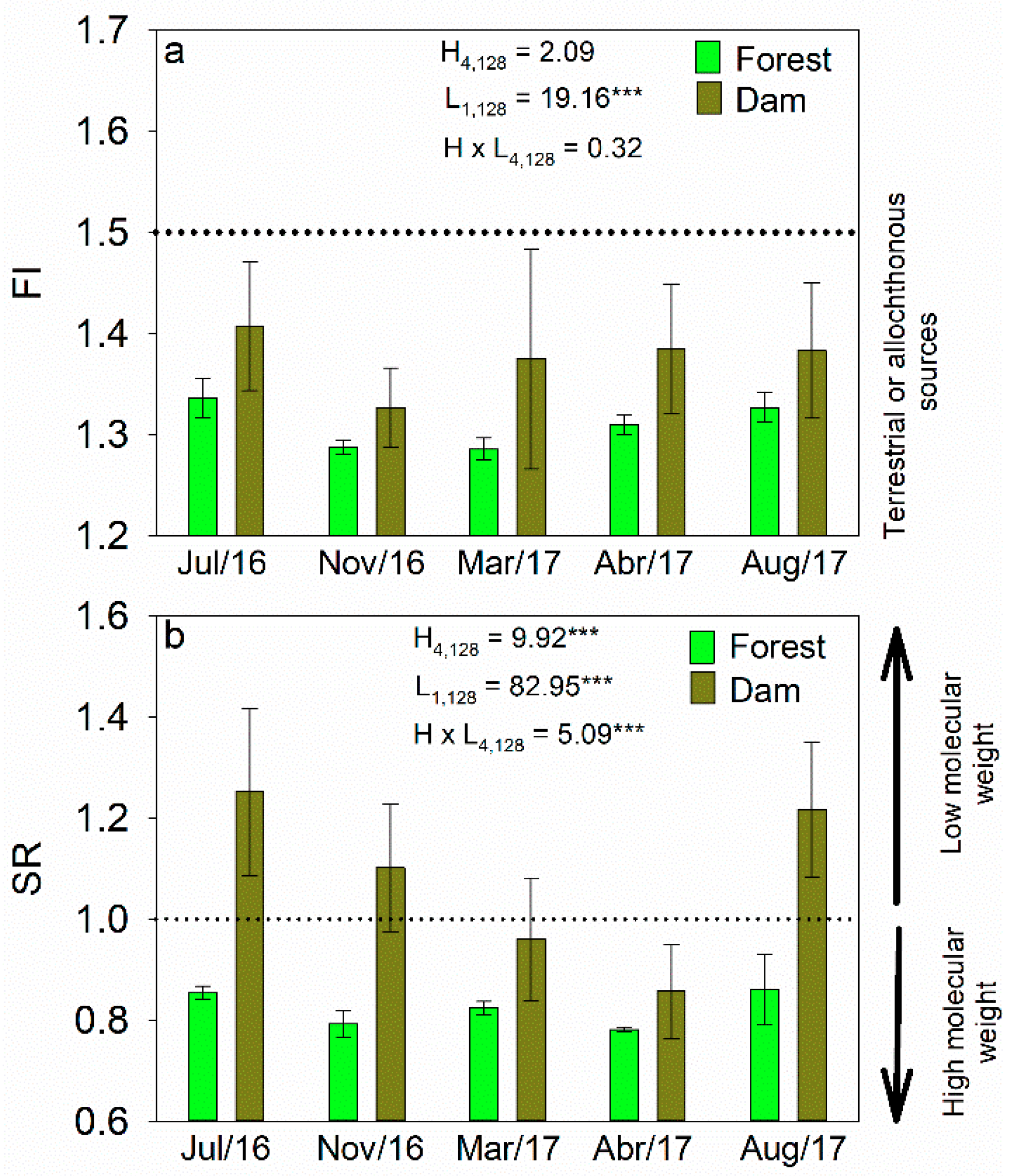
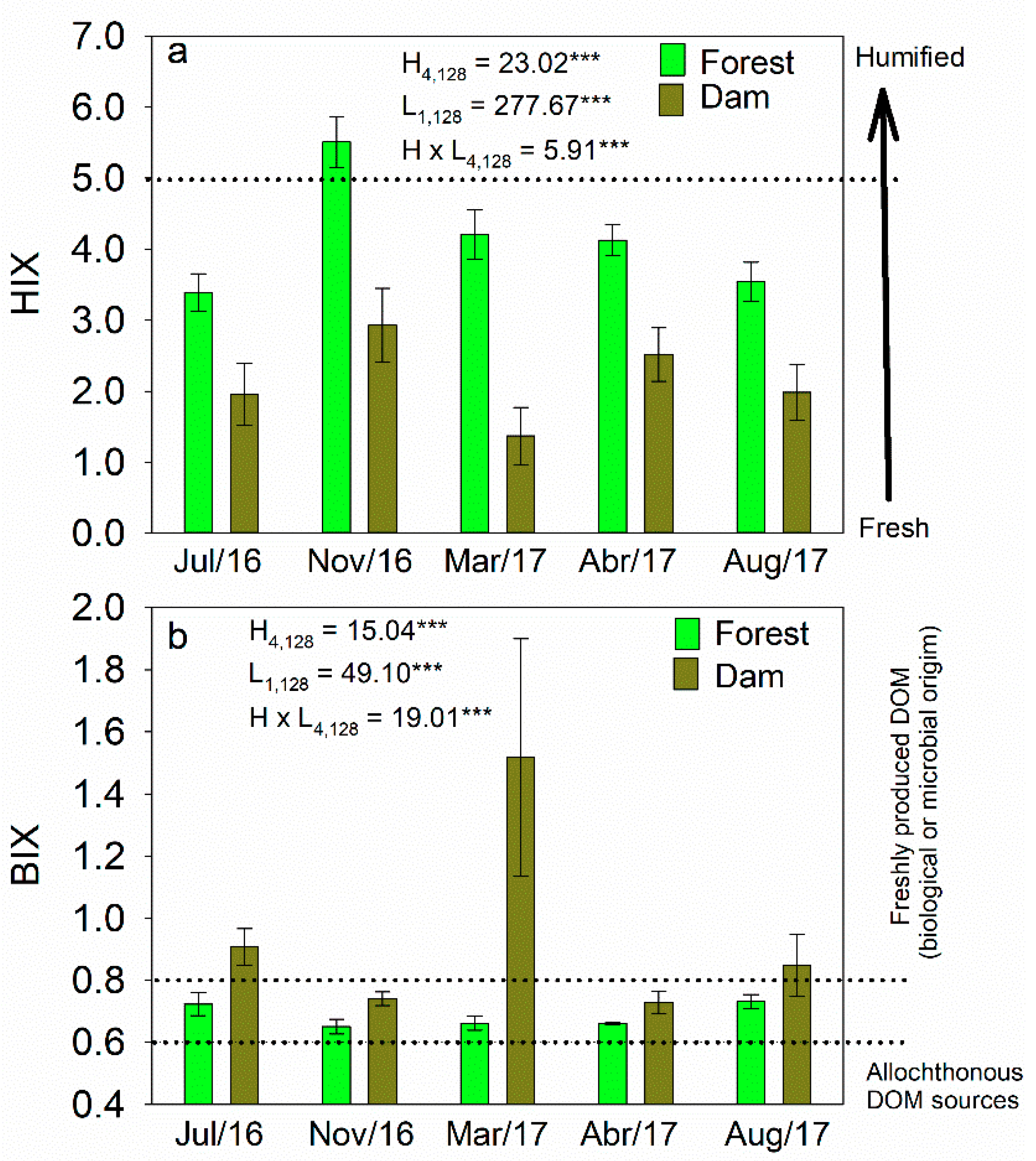
3.2. Concentrations of DOC and Optical Characteristics of DOM
3.3. Fluorescence Characterization by PARAFAC
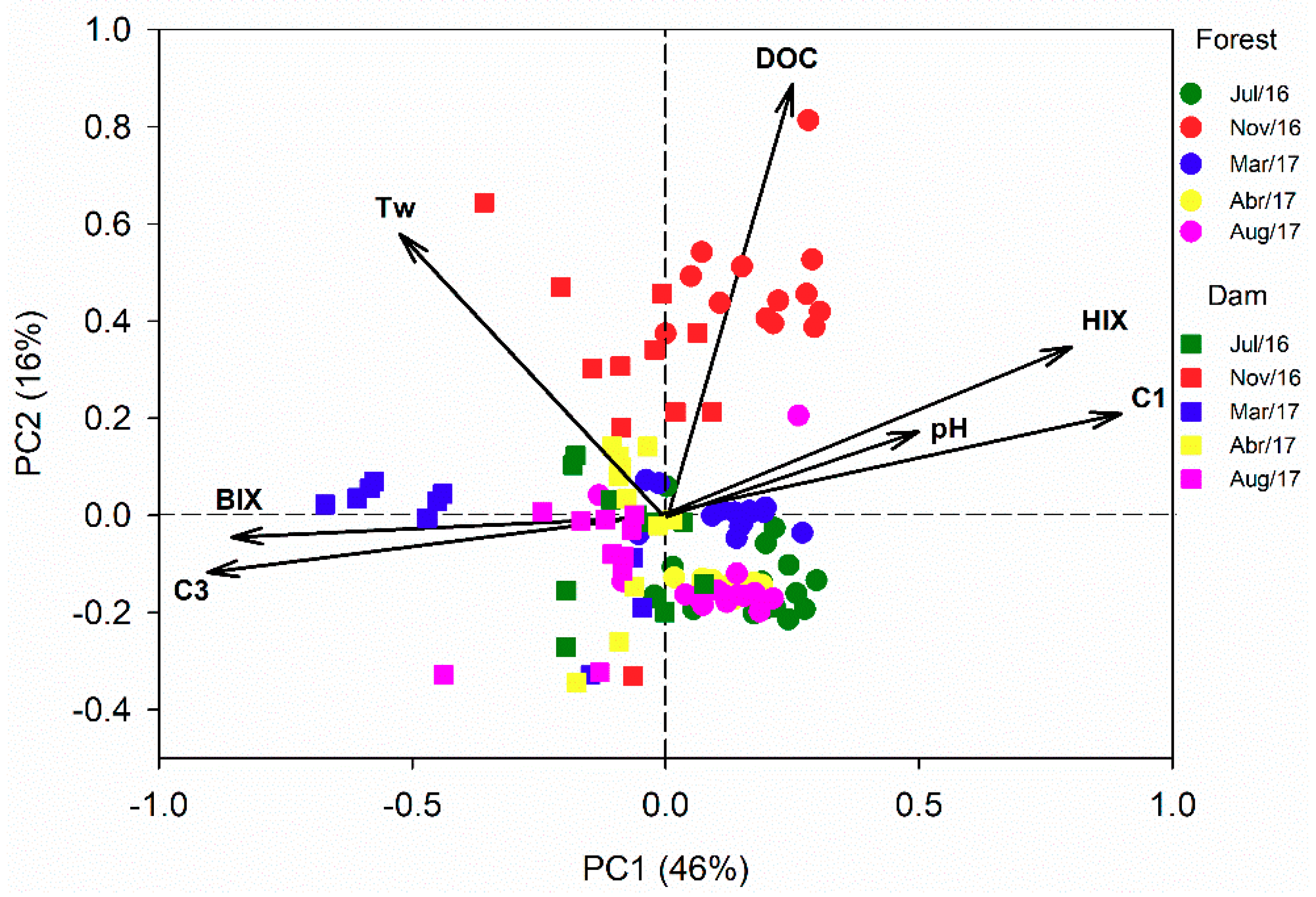
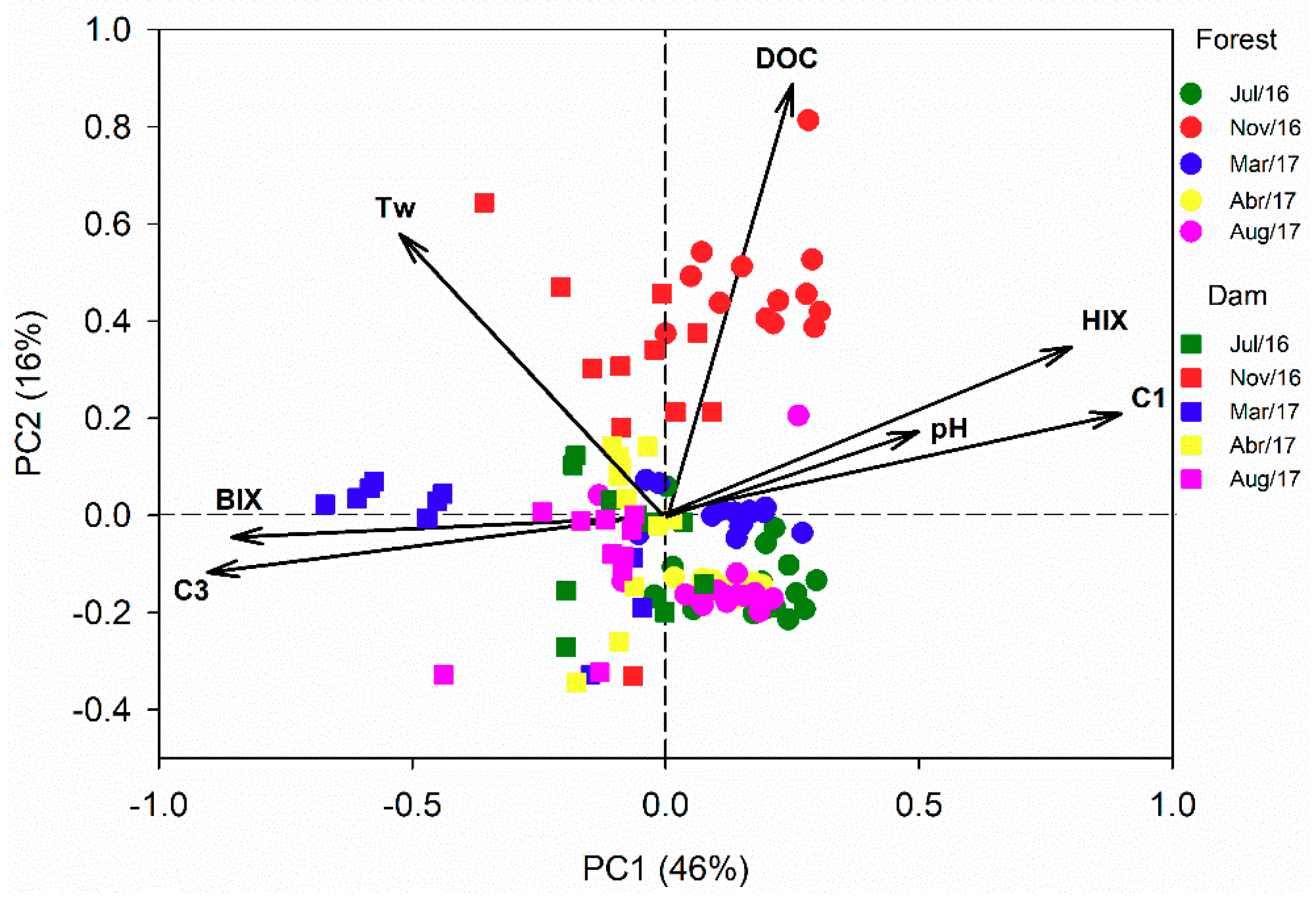
3.4. Principal Component Analysis (PCA)
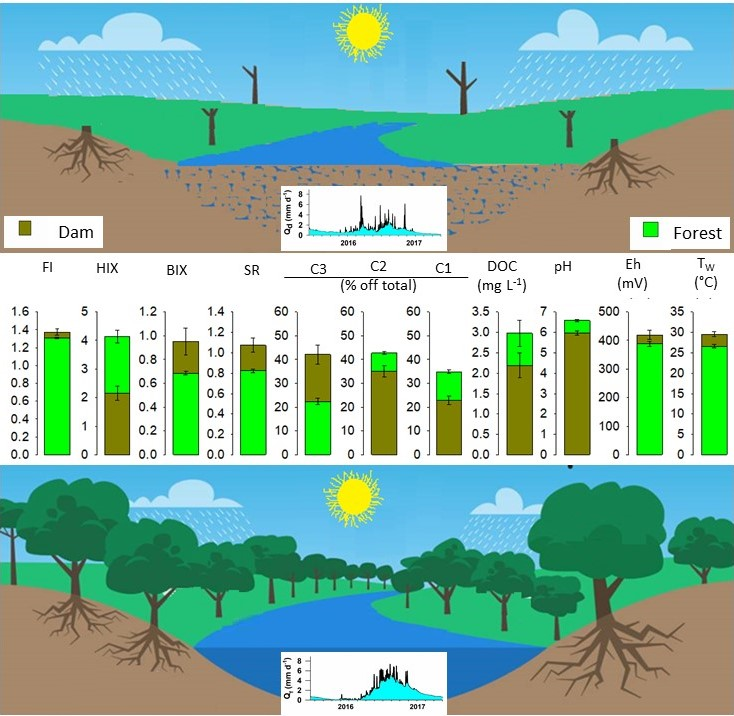
4. Discussion
4.1. Variations of Hydrology in Dam and Forest Catchments
4.2. Seasonal Variability in DOC and Optical Properties of DOM
4.3. Effects of Land Cover on Optical Properties of DOM in Headwater Catchments
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malhi, Y.; Roberts, J.; Betts, R.; Killeen, T.; Li, W.; Nobre, C. Climate change, deforestation, and the fate of the amazon. Science 2008, 319, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Neu, V.; Ward, N.D.; Krusche, A.V.; Neill, C. Dissolved organic and inorganic carbon flow paths in an Amazonian transitional forest. Front. Mar. Sci. 2016, 3, 114. [Google Scholar] [CrossRef]
- Lathuillière, M.J.; Coe, M.T.; Johnson, M.S. A review of green- and blue-water resources and their trade-offs for future agricultural production in the amazon basin: What could irrigated agriculture mean for Amazonia? Hydrol. Earth Syst. Sci. 2016, 20, 2179–2194. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Global Forest Resources Assessment 2015, 2nd ed.; FAO: Rome, Italy, 2015. [Google Scholar]
- Levy, M.; Lopes, A.; Cohn, A.; Larsen, L.; Thompson, S. Land use change increases streamflow across the arc of deforestation in Brazil. Geophys. Res. Lett. 2018, 45, 3520–3530. [Google Scholar] [CrossRef]
- INPE. Projeto Prodes, Monitoramente da Floresta Amazonica Brasileira por Satelite. Available online: http://www.obt.inpe.br/prodes/dashboard/prodes-rates.html (accessed on 14 October 2018).
- Macedo, M.N.; DeFries, R.S.; Morton, D.C.; Stickler, C.M.; Galford, G.L.; Shimabukuro, Y.E. Decoupling of deforestation and soy production in the southern Amazon during the late 2000s. Proc. Natl. Acad. Sci. USA 2012, 109, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Barona, E.; Ramankutty, N.; Hyman, G.; Coomes, O.T. The role of pasture and soybean in deforestation of the Brazilian Amazon. Environ. Res. Lett. 2010, 5, 024002. [Google Scholar] [CrossRef]
- Johnson, M.S.; Lehmann, J.; Riha, S.J.; Krusche, A.V.; Richey, J.E.; Ometto, J.P.H.; Couto, E.G. CO2 efflux from Amazonian headwater streams represents a significant fate for deep soil respiration. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef]
- Hayhoe, S.J.; Neill, C.; Porder, S.; McHorney, R.; Lefebvre, P.; Coe, M.T.; Elsenbeer, H.; Krusche, A.V. Conversion to soy on the Amazonian agricultural frontier increases streamflow without affecting stormflow dynamics. Glob. Chang. Biol. 2011, 17, 1821–1833. [Google Scholar] [CrossRef]
- Macedo, M.N.; Coe, M.T.; DeFries, R.; Uriarte, M.; Brando, P.M.; Neill, C.; Walker, W.S. Land-use-driven stream warming in southeastern Amazonia. Phil. Trans. R. Soc. B 2013, 368, 20120153. [Google Scholar] [CrossRef] [PubMed]
- Ågren, A.; Berggren, M.; Laudon, H.; Jansson, M. Terrestrial export of highly bioavailable carbon from small boreal catchments in spring floods. Freshw. Biol. 2008, 53, 964–972. [Google Scholar] [CrossRef]
- Laudon, H.; Berggren, M.; Ågren, A.; Buffam, I.; Bishop, K.; Grabs, T.; Jansson, M.; Köhler, S. Patterns and dynamics of dissolved organic carbon (DOC) in boreal streams: The role of processes, connectivity, and scaling. Ecosystems 2011, 14, 880–893. [Google Scholar] [CrossRef]
- McElmurry, S.P.; Long, D.T.; Voice, T.C. Stormwater dissolved organic matter: Influence of land cover and environmental factors. Environ. Sci. Technol. 2013, 48, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.D.; Reissig, M.; Queimaliños, C.P.; Garcia, P.E.; Dieguez, M.C. Climate-driven terrestrial inputs in ultraoligotrophic mountain streams of Andean Patagonia revealed through chromophoric and fluorescent dissolved organic matter. Sci. Total Environ. 2015, 521, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Dalmagro, H.J.; Johnson, M.S.; Musis, C.R.; Lathuillière, M.J.; Graesser, J.; Junior, O.B.; Couto, E.G. Spatial patterns of DOC concentration and DOM optical properties in a Brazilian tropical river-wetland system. J. Geophys. Res. Biogeosci. 2017, 122, 1883–1902. [Google Scholar] [CrossRef]
- Lambert, T.; Teodoru, C.R.; Nyoni, F.C.; Borges, A.V. Along-stream transport and transformation of dissolved organic matter in a large tropical river. Biogeosciences 2016, 13, 2727–2741. [Google Scholar] [CrossRef]
- Wilson, H.F.; Xenopoulos, M.A. Ecosystem and seasonal control of stream dissolved organic carbon along a gradient of land use. Ecosystems 2008, 11, 555–568. [Google Scholar] [CrossRef]
- Yamashita, Y.; Scinto, L.J.; Maie, N.; Jaffé, R. Dissolved organic matter characteristics across a subtropical wetland’s landscape: Application of optical properties in the assessment of environmental dynamics. Ecosystems 2010, 13, 1006–1019. [Google Scholar] [CrossRef]
- Williams, C.J.; Yamashita, Y.; Wilson, H.F.; Jaffé, R.; Xenopoulos, M.A. Unraveling the role of land use and microbial activity in shaping dissolved organic matter characteristics in stream ecosystems. Limnol. Oceanogr. 2010, 55, 1159–1171. [Google Scholar] [CrossRef]
- Jaffé, R.; McKnight, D.; Maie, N.; Cory, R.; McDowell, W.; Campbell, J. Spatial and temporal variations in DOM composition in ecosystems: The importance of long-term monitoring of optical properties. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- Fellman, J.B.; Hood, E.; Edwards, R.T.; D’Amore, D.V. Changes in the concentration, biodegradability, and fluorescent properties of dissolved organic matter during stormflows in coastal temperate watersheds. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Spencer, R.G.; Stubbins, A.; Hernes, P.J.; Baker, A.; Mopper, K.; Aufdenkampe, A.K.; Dyda, R.Y.; Mwamba, V.L.; Mangangu, A.M.; Wabakanghanzi, J.N. Photochemical degradation of dissolved organic matter and dissolved lignin phenols from the Congo River. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Singh, S.; Inamdar, S.; Scott, D. Comparison of two parafac models of dissolved organic matter fluorescence for a mid-atlantic forested watershed in the USA. J. Ecosyst. 2013, 2013, 532424. [Google Scholar] [CrossRef]
- Mariot, M.; Dudal, Y.; Furian, S.; Sakamoto, A.; Vallès, V.; Fort, M.; Barbiero, L. Dissolved organic matter fluorescence as a water-flow tracer in the tropical wetland of Pantanal of Nhecolândia, Brazil. Sci. Total Environ. 2007, 388, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Fellman, J.B.; Hood, E.; Spencer, R.G. Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: A review. Limnol. Oceanogr. 2010, 55, 2452–2462. [Google Scholar] [CrossRef]
- Yamashita, Y.; Kloeppel, B.D.; Knoepp, J.; Zausen, G.L.; Jaffé, R. Effects of watershed history on dissolved organic matter characteristics in headwater streams. Ecosystems 2011, 14, 1110–1122. [Google Scholar] [CrossRef]
- Stedmon, C.A.; Markager, S.; Bro, R. Tracing dissolved organic matter in aquatic environments using a new approach to fluorescence spectroscopy. Mar. Chem. 2003, 82, 239–254. [Google Scholar] [CrossRef]
- Larsen, L.G.; Aiken, G.R.; Harvey, J.W.; Noe, G.B.; Crimaldi, J.P. Using fluorescence spectroscopy to trace seasonal DOM dynamics, disturbance effects, and hydrologic transport in the Florida Everglades. J. Geophys. Res. Biogeosci. 2010, 115. [Google Scholar] [CrossRef]
- Fasching, C.; Behounek, B.; Singer, G.A.; Battin, T.J. Microbial degradation of terrigenous dissolved organic matter and potential consequences for carbon cycling in brown-water streams. Sci. Rep. 2014, 4, 4981. [Google Scholar] [CrossRef] [PubMed]
- Baker, A. Fluorescence properties of some farm wastes: Implications for water quality monitoring. Water Res. 2002, 36, 189–195. [Google Scholar] [CrossRef]
- Dubreuil, V.; Debortoli, N.; Funatsu, B.; Nédélec, V.; Durieux, L. Impact of land-cover change in the southern Amazonia climate: A case study for the region of Alta Floresta, Mato Grosso, Brazil. Environ. Monit. Assess. 2012, 184, 877–891. [Google Scholar] [CrossRef] [PubMed]
- International Union of Soil Sciences Working Group WRB. World Reference Base for Soil Resources 2014 (Update 2015), International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Report No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Santos, V. Análise florística e Estrutural de uma Floresta Ombrófila Aberta Primária no Parque Estadual Cristalino, Alta Floresta–MT. Master’s Dissertation, Universidade Estadual do Mato Grosso, Alta Floresta, Brazil, 2005. [Google Scholar]
- Hund, S.V.; Johnson, M.S.; Keddie, T. Developing a hydrologic monitoring network in data-scarce regions using open-source Arduino dataloggers. Agric. Environ. Lett. 2016, 1. [Google Scholar] [CrossRef]
- Cunha, S.B.D.; Guerra, A.J.T. Geomorfologia: Exercícios, Técnicas e Aplicações; Bertand Brazil: Rio de Janeiro, Brazil, 1996; p. 345. ISBN 8528605485. [Google Scholar]
- Ladson, A.; Brown, R.; Neal, B.; Nathan, R. A standard approach to baseflow separation using the Lyne and Hollick filter. Australas. J. Water Resour. 2013, 17, 25–34. [Google Scholar] [CrossRef]
- Lyne, V.; Hollick, M. Stochastic Time-Variable Rainfall-Runoff Modelling. In Proceedings of the Institute of Engineers Australia National Conference: Engineering Nanagement—update, Melbourne, Australia, 21–22 March 1979; pp. 89–93. [Google Scholar]
- Dalmagro, H.J.; Lathuillière, M.J.; Hawthorne, I.; de Morais, D.; Pinto Júnior, O.B.; Couto, E.G.; Johnson, M.S. Carbon biogeochemistry of a flooded Pantanal forest over three annual flood cycles. Biogeochemistry 2018, 139, 1–18. [Google Scholar] [CrossRef]
- Vepraskas, M.J.; Faulkner, S. Redox chemistry of hydric soils. In Wetland Soils: Genesis, Hydrology, Landscapes, and Classification; Vepraskas, M.J., Richardson, J.L., Vepraskas, M.J., Craft, C.B., Eds.; CRC Press: Boca Raton, MA, USA, 2000; p. 432. [Google Scholar]
- Dalmagro, H.J.; Lathuillière, M.J.; Vourlitis, G.L.; Campos, R.C.; Pinto, O.B.; Johnson, M.S.; Ortíz, C.E.; Lobo, F.D.A.; Couto, E.G. Physiological responses to extreme hydrological events in the Pantanal wetland: Heterogeneity of a plant community containing super-dominant species. J. Veg. Sci. 2016, 27, 568–577. [Google Scholar] [CrossRef]
- Eykelbosh, A.J.; Johnson, M.S.; Couto, E.G. Biochar decreases dissolved organic carbon but not nitrate leaching in relation to vinasse application in a Brazilian sugarcane soil. J. Environ. Manag. 2015, 149, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.R.; Stedmon, C.A.; Graeber, D.; Bro, R. Fluorescence spectroscopy and multi-way techniques. PARAFAC. Anal. Methods 2013, 5, 6557–6566. [Google Scholar] [CrossRef]
- Stedmon, C.A.; Markager, S. Resolving the variability in dissolved organic matter fluorescence in a temperate estuary and its catchment using PARAFAC analysis. Limnol. Oceanogr. 2005, 50, 686–697. [Google Scholar] [CrossRef]
- Massicotte, M.P. Package ‘eemr’. 2016. Available online: http://cran.fhcrc.org/web/packages/eemR/eemR.pdf (accessed on 10 May 2018).
- Lawaetz, A.J.; Stedmon, C.A. Fluorescence intensity calibration using the Raman scatter peak of water. Appl. Spectrosc. 2009, 63, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Stedmon, C.A.; Bro, R. Characterizing dissolved organic matter fluorescence with parallel factor analysis: A tutorial. Limnol. Oceanogr. Methods 2008, 6, 572–579. [Google Scholar] [CrossRef]
- Murphy, K.R.; Stedmon, C.A.; Wenig, P.; Bro, R. Openfluor—An online spectral library of auto-fluorescence by organic compounds in the environment. Anal. Methods 2014, 6, 658–661. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Shen, Z.; Chen, J.; Feng, C. Characterization and spacial distribution variability of chromophoric dissolved organic matter (CDOM) in the Yangtze estuary. Chemosphere 2014, 95, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Cory, R.M.; McKnight, D.M. Fluorescence spectroscopy reveals ubiquitous presence of oxidized and reduced quinones in dissolved organic matter. Environ. Sci. Technol. 2005, 39, 8142–8149. [Google Scholar] [CrossRef] [PubMed]
- McKnight, D.M.; Boyer, E.W.; Westerhoff, P.K.; Doran, P.T.; Kulbe, T.; Andersen, D.T. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol. Oceanogr. 2001, 46, 38–48. [Google Scholar] [CrossRef]
- Huguet, A.; Vacher, L.; Relexans, S.; Saubusse, S.; Froidefond, J.-M.; Parlanti, E. Properties of fluorescent dissolved organic matter in the Gironde Estuary. Org. Geochem. 2009, 40, 706–719. [Google Scholar] [CrossRef]
- Zsolnay, A.; Baigar, E.; Jimenez, M.; Steinweg, B.; Saccomandi, F. Differentiating with fluorescence spectroscopy the sources of dissolved organic matter in soils subjected to drying. Chemosphere 1999, 38, 45–50. [Google Scholar] [CrossRef]
- Ohno, T. Fluorescence inner-filtering correction for determining the humification index of dissolved organic matter. Environ. Sci. Technol. 2002, 36, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Bro, R. Dissolved organic matter characterization using multiway spectral decomposition of fluorescence landscapes. Soil Sci. Soc. Am. J. 2006, 70, 2028–2037. [Google Scholar] [CrossRef]
- Ohno, T.; Chorover, J.; Omoike, A.; Hunt, J. Molecular weight and humification index as predictors of adsorption for plant-and manure-derived dissolved organic matter to goethite. Eur. J. Soil Sci. 2007, 58, 125–132. [Google Scholar] [CrossRef]
- Hunt, J.F.; Ohno, T. Characterization of fresh and decomposed dissolved organic matter using excitation−emission matrix fluorescence spectroscopy and multiway analysis. J. Agric. Food Chem. 2007, 55, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Birdwell, J.E.; Engel, A.S. Characterization of dissolved organic matter in cave and spring waters using UV–vis absorbance and fluorescence spectroscopy. Org. Geochem. 2010, 41, 270–280. [Google Scholar] [CrossRef]
- Yamashita, Y.; Fichot, C.G.; Shen, Y.; Jaffé, R.; Benner, R. Linkages among fluorescent dissolved organic matter, dissolved amino acids and lignin-derived phenols in a river-influenced ocean margin. Front. Mar. Sci. 2015, 2, 92. [Google Scholar] [CrossRef]
- Helms, J.R.; Stubbins, A.; Ritchie, J.D.; Minor, E.C.; Kieber, D.J.; Mopper, K. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 2008, 53, 955–969. [Google Scholar] [CrossRef]
- Graeber, D.; Gelbrecht, J.; Pusch, M.T.; Anlanger, C.; von Schiller, D. Agriculture has changed the amount and composition of dissolved organic matter in central European headwater streams. Sci. Total Environ. 2012, 438, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Coble, P.G. Marine optical biogeochemistry: The chemistry of ocean color. Chem. Rev. 2007, 107, 402–418. [Google Scholar] [CrossRef] [PubMed]
- Osburn, C.L.; Wigdahl, C.R.; Fritz, S.C.; Saros, J.E. Dissolved organic matter composition and photoreactivity in prairie lakes of the US Great Plains. Limnol. Oceanogr. 2011, 56, 2371–2390. [Google Scholar] [CrossRef]
- Baker, A. Fluorescence excitation−emission matrix characterization of some sewage-impacted rivers. Environ. Sci. Technol. 2001, 35, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Coe, M.; Latrubesse, E.; Ferreira, M.; Amsler, M. The effects of deforestation and climate variability on the streamflow of the Araguaia River, Brazil. Biogeochemistry 2011, 105, 119–131. [Google Scholar] [CrossRef]
- von Randow, C.; Manzi, A.O.; Kruijt, B.; De Oliveira, P.; Zanchi, F.; Silva, R.; Hodnett, M.; Gash, J.; Elbers, J.; Waterloo, M. Comparative measurements and seasonal variations in energy and carbon exchange over forest and pasture in South West Amazonia. Theor. Appl. Climatol. 2004, 78, 5–26. [Google Scholar] [CrossRef]
- Lehner, B.; Liermann, C.R.; Revenga, C.; Vörösmarty, C.; Fekete, B.; Crouzet, P.; Döll, P.; Endejan, M.; Frenken, K.; Magome, J. High-resolution mapping of the world’s reservoirs and dams for sustainable river-flow management. Front. Ecol. Environ. 2011, 9, 494–502. [Google Scholar] [CrossRef]
- Shiklomanov, I.A. Appraisal and assessment of world water resources. Water Int. 2000, 25, 11–32. [Google Scholar] [CrossRef]
- Palhares, J. Consumo de água na produção animal. In Embrapa Pecuária Sudeste-Comun. Técnico (Infoteca-E); Embrapa Pecuária Sudeste: São Carlos, SP, Brazil, 2013. [Google Scholar]
- Ward, D.; McKague, K. Water Requirements of Livestock; Order No. 07-023; Ministry of Agriculture, Food and Rural Affairs: Ontario, Canada, 2007. [Google Scholar]
- Zimmermann, B.; Elsenbeer, H.; De Moraes, J.M. The influence of land-use changes on soil hydraulic properties: Implications for runoff generation. For. Ecol. Manag. 2006, 222, 29–38. [Google Scholar] [CrossRef]
- Germer, S.; Neill, C.; Krusche, A.V.; Elsenbeer, H. Influence of land-use change on near-surface hydrological processes: Undisturbed forest to pasture. J. Hydrol. 2010, 380, 473–480. [Google Scholar] [CrossRef]
- Neill, C.; Jankowski, K.; Brando, P.M.; Coe, M.T.; Deegan, L.A.; Macedo, M.N.; Riskin, S.H.; Porder, S.; Elsenbeer, H.; Krusche, A.V. Surprisingly modest water quality impacts from expansion and intensification of large-scale commercial agriculture in the Brazilian Amazon-Cerrado region. Trop. Conserv. Sci. 2017, 10, 1–5. [Google Scholar]
- Johnson, M.S.; Lehmann, J.; Couto, E.G.; Novaes Filho, J.P.; Riha, S.J. DOC and DIC in flowpaths of Amazonian headwater catchments with hydrologically contrasting soils. Biogeochemistry 2006, 81, 45–57. [Google Scholar] [CrossRef]
- Xu, N.; Saiers, J.E. Temperature and hydrologic controls on dissolved organic matter mobilization and transport within a forest topsoil. Environ. Sci. Technol. 2010, 44, 5423–5429. [Google Scholar] [CrossRef] [PubMed]
- Mann, P.; Davydova, A.; Zimov, N.; Spencer, R.; Davydov, S.; Bulygina, E.; Zimov, S.; Holmes, R. Controls on the composition and lability of dissolved organic matter in Siberia’s Kolyma river basin. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef]
- Lambert, T.; Darchambeau, F.; Bouillon, S.; Alhou, B.; Mbega, J.-D.; Teodoru, C.R.; Nyoni, F.C.; Massicotte, P.; Borges, A.V. Landscape control on the spatial and temporal variability of chromophoric dissolved organic matter and dissolved organic carbon in large African rivers. Ecosystems 2015, 18, 1224–1239. [Google Scholar] [CrossRef]
- Wilson, H.F.; Xenopoulos, M.A. Effects of agricultural land use on the composition of fluvial dissolved organic matter. Nat. Geosci. 2009, 2, 37–41. [Google Scholar] [CrossRef]
- Meyer, J.L.; Tate, C.M. The effects of watershed disturbance on dissolved organic carbon dynamics of a stream. Ecology 1983, 64, 33–44. [Google Scholar] [CrossRef]
- Cammack, W.L.; Kalff, J.; Prairie, Y.T.; Smith, E.M. Fluorescent dissolved organic matter in lakes: Relationships with heterotrophic metabolism. Limnol. Oceanogr. 2004, 49, 2034–2045. [Google Scholar] [CrossRef]
- Balcarczyk, K.L.; Jones Jr., J.; Jaffé, R.; Maie, N. Dissolved Organic Matter Bioavailability and Composition in Streams Draining Catchments with Discontinuous Permafrost. Doctoral Dissertation, University of Alaska Fairbanks, Fairbanks, AK, USA, 2008. [Google Scholar]
- Baker, A.; Spencer, R.G. Characterization of dissolved organic matter from source to sea using fluorescence and absorbance spectroscopy. Sci. Total Environ. 2004, 333, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Hudson, N.; Baker, A.; Reynolds, D. Fluorescence analysis of dissolved organic matter in natural, waste and polluted waters—A review. River Res. Appl. 2007, 23, 631–649. [Google Scholar] [CrossRef]
- Maie, N.; Scully, N.M.; Pisani, O.; Jaffé, R. Composition of a protein-like fluorophore of dissolved organic matter in coastal wetland and estuarine ecosystems. Water Res. 2007, 41, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Moran, M.A.; Zepp, R.G. Invited review role of photoreactions in the formation of biologically labile compounds from dissolved organic matter. Oceanography 1997, 42, 1307–1316. [Google Scholar]
- Ishii, S.K.; Boyer, T.H. Behavior of reoccurring parafac components in fluorescent dissolved organic matter in natural and engineered systems: A critical review. Environ. Sci. Technol. 2012, 46, 2006–2017. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | Dam | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Date | Month | PPT (mm) | Qf (BF) (mm) | Qf (mm d-1) | % BF | % SF | Qd Accumulated (mm) | Qd (mm d-1) | % BF | % SF | ||
| Mean | Median | Mean | Median | |||||||||
| 25/07/2016 | Jul/16 | 32 | 26.9 | 0.38 | 0.40 | 94.9 | 5.1 | 59.0 | 0.84 | 0.81 | 96.6 | 3.4 |
| 05/11/2016 | Nov/16 | 464 | 12.1 | 0.12 | 0.62 | 41.6 | 58.4 | 72.1 | 0.70 | 0.62 | 84.6 | 15.4 |
| 07/03/2017 | Mar/17 | 1414 | 339.2 | 2.78 | 2.36 | 83.4 | 16.6 | 217.9 | 1.79 | 1.47 | 71.5 | 28.5 |
| 29/04/2017 | Abr/17 | 403 | 205.5 | 3.88 | 3.55 | 86.5 | 13.5 | 83.5 | 1.57 | 1.25 | 77.4 | 22.6 |
| 28/08/2017 | Aug/17 | 36 | 166.5 | 1.38 | 1.11 | 97.0 | 3.0 | 54.7 | 0.45 | 0.34 | 94.7 | 8.7 |
| Total (study period) | 2350 | 750.8 | 487.3 | |||||||||
| Mean (study period) | 1.60 | 87 | 13 | 1.04 | 80 | 20 | ||||||
| Component 1 | Component 2 | Component 3 | ||||||
| Ex (nm) | Em (nm) | Contribution to total fDOM | Ex (nm) | Em (nm) | Contribution to total fDOM | Ex (nm) | Em (nm) | Contribution to total fDOM |
| <252 (357) | 474 | 40.1% | <252 (321) | 408 | 39.1% | 288 | 333 | 20.8% |
| UV Humic-Like | Ubiquitous Humic-Like | Protein-Like | ||||||
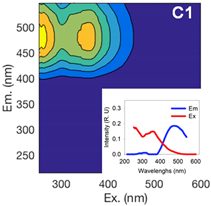 | 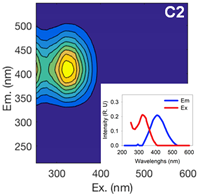 | 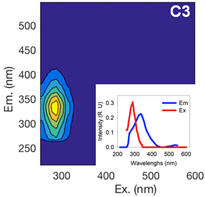 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalmagro, H.J.; Lathuillière, M.J.; Sallo, F.d.S.; Guerreiro, M.F.; Pinto, O.B., Jr.; de Arruda, P.H.Z.; Couto, E.G.; Johnson, M.S. Streams with Riparian Forest Buffers versus Impoundments Differ in Discharge and DOM Characteristics for Pasture Catchments in Southern Amazonia. Water 2019, 11, 390. https://doi.org/10.3390/w11020390
Dalmagro HJ, Lathuillière MJ, Sallo FdS, Guerreiro MF, Pinto OB Jr., de Arruda PHZ, Couto EG, Johnson MS. Streams with Riparian Forest Buffers versus Impoundments Differ in Discharge and DOM Characteristics for Pasture Catchments in Southern Amazonia. Water. 2019; 11(2):390. https://doi.org/10.3390/w11020390
Chicago/Turabian StyleDalmagro, Higo J., Michael J. Lathuillière, Fernando da S. Sallo, Maurel F. Guerreiro, Osvaldo B. Pinto, Jr., Paulo H.Z. de Arruda, Eduardo G. Couto, and Mark S. Johnson. 2019. "Streams with Riparian Forest Buffers versus Impoundments Differ in Discharge and DOM Characteristics for Pasture Catchments in Southern Amazonia" Water 11, no. 2: 390. https://doi.org/10.3390/w11020390
APA StyleDalmagro, H. J., Lathuillière, M. J., Sallo, F. d. S., Guerreiro, M. F., Pinto, O. B., Jr., de Arruda, P. H. Z., Couto, E. G., & Johnson, M. S. (2019). Streams with Riparian Forest Buffers versus Impoundments Differ in Discharge and DOM Characteristics for Pasture Catchments in Southern Amazonia. Water, 11(2), 390. https://doi.org/10.3390/w11020390