Integrating Hydrogeological and Microbiological Data and Modelling to Characterize the Hydraulic Features and Behaviour of Coastal Carbonate Aquifers: A Case in Western Cuba


 ,
,  ,
, 


 ,
, 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
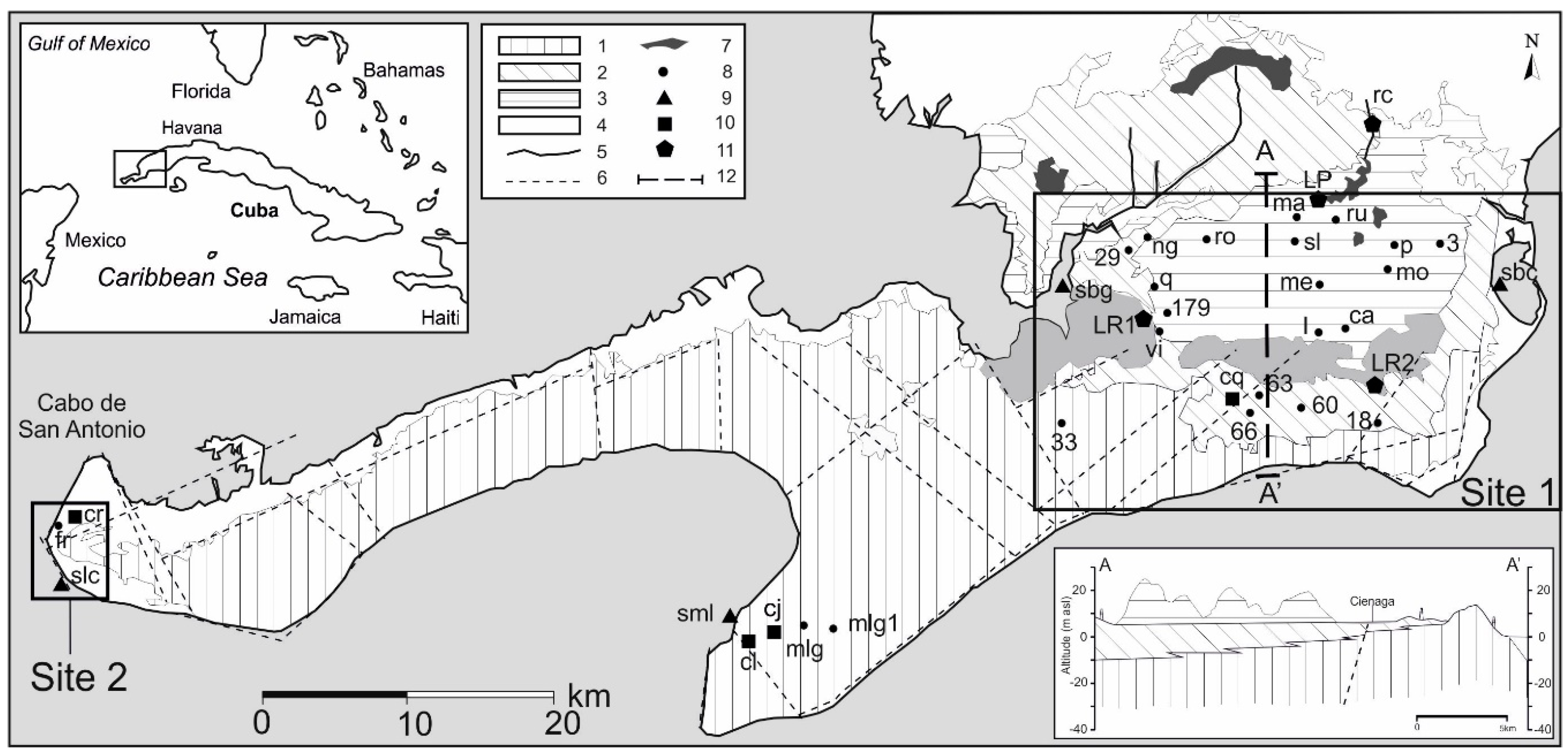
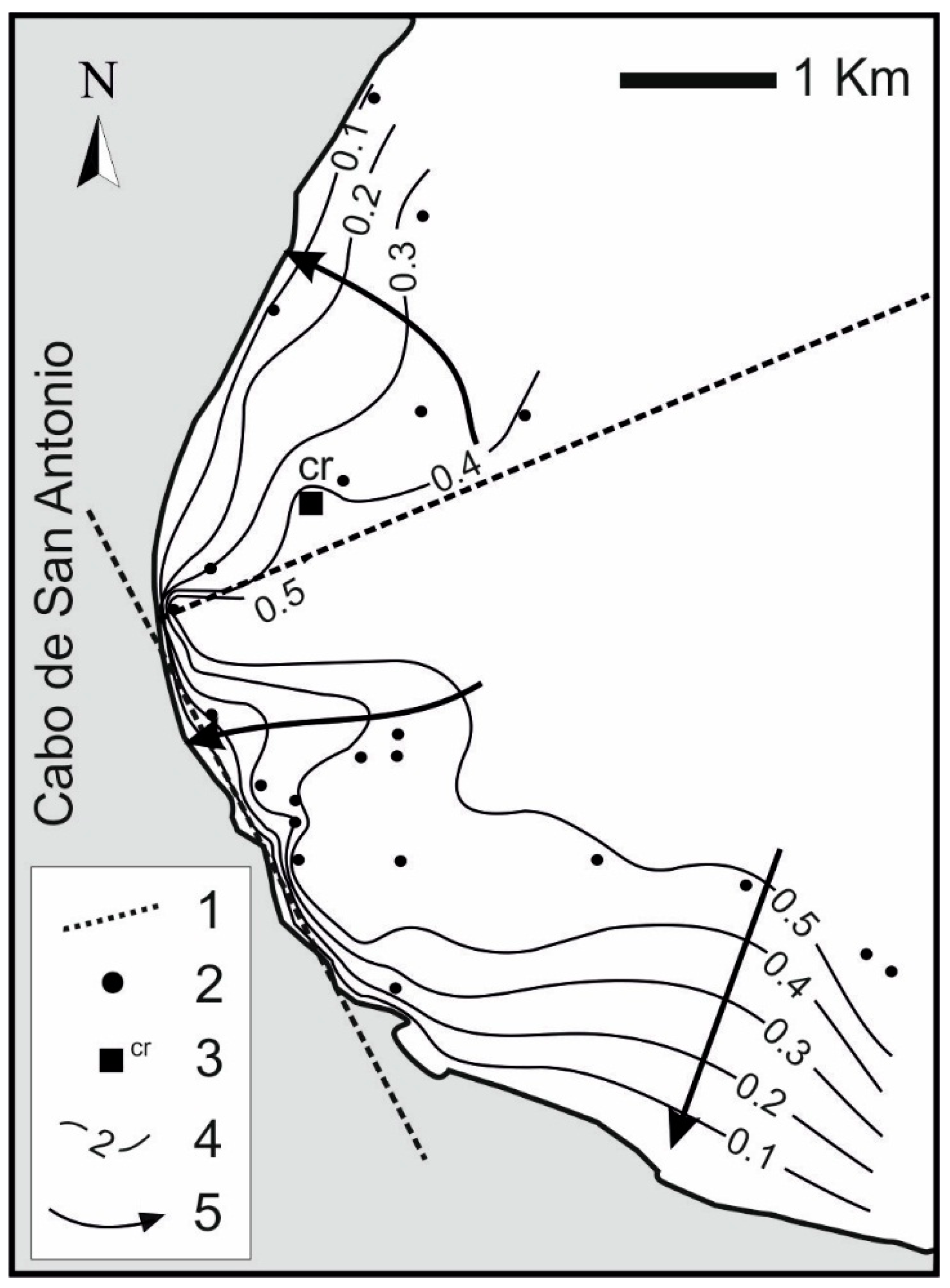
2. Study Area
3. Materials and Methods
3.1. Hydraulic Head Measurements
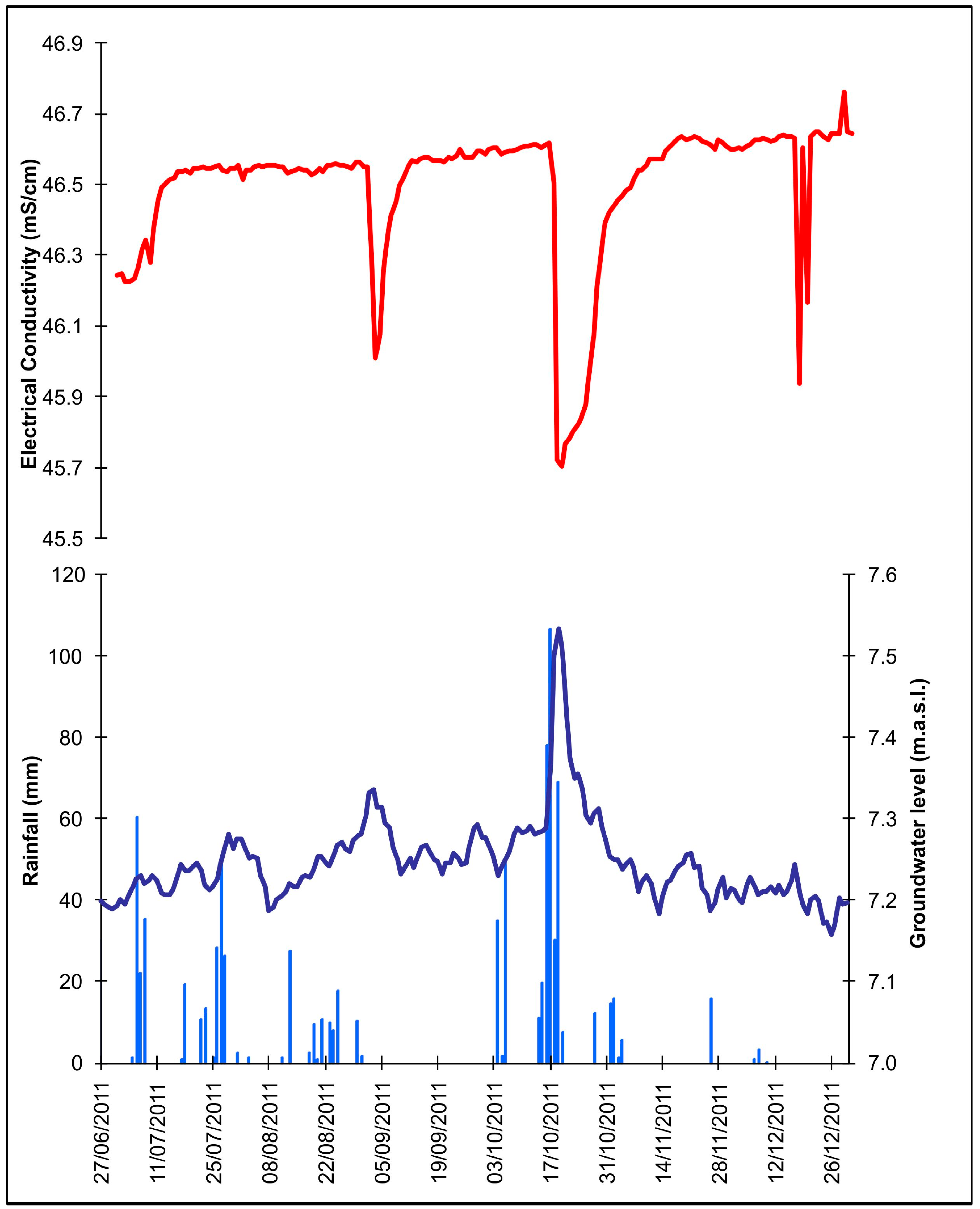
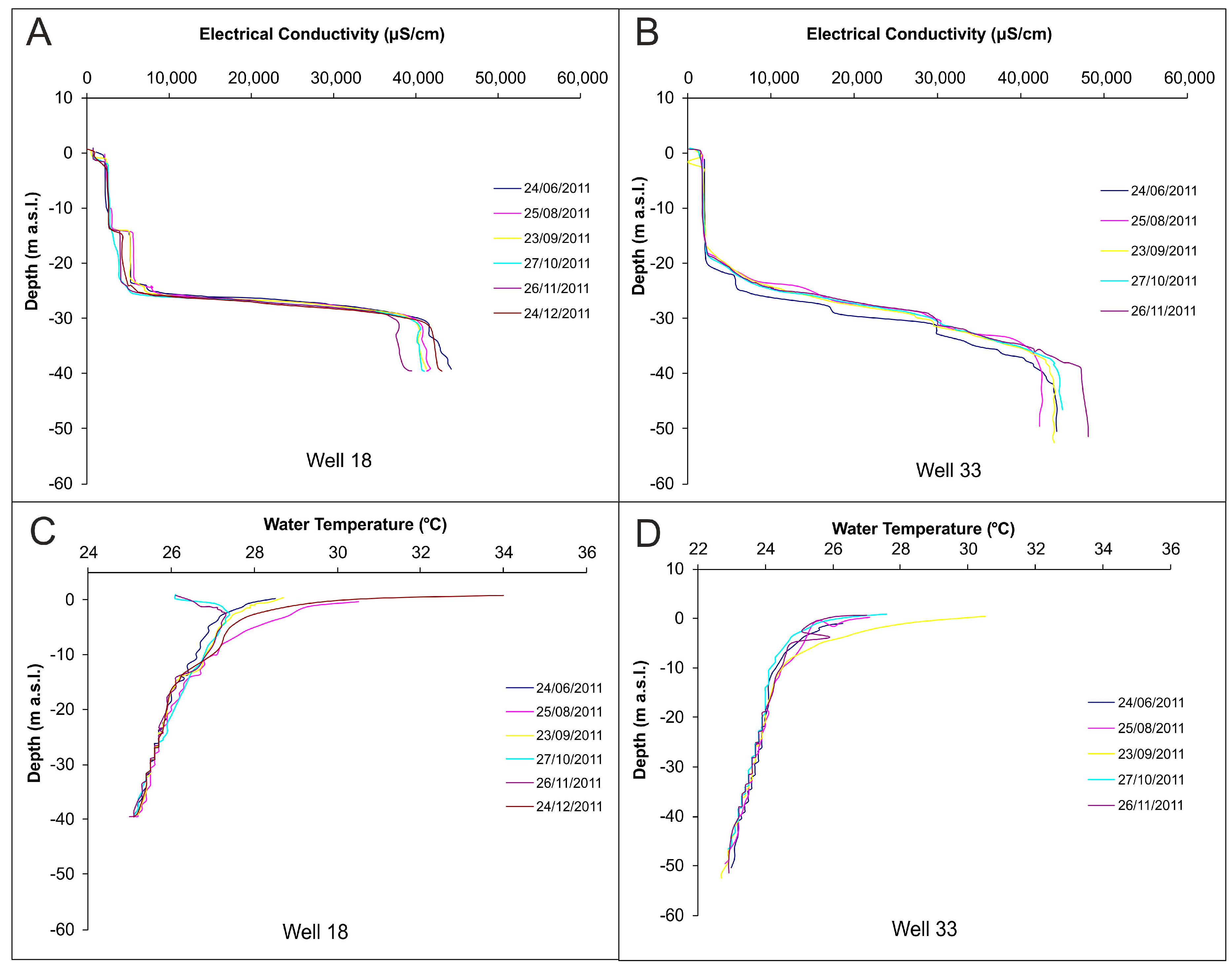
3.2. Water Sampling, Temperature and Electrical Conductivity Profiles
3.3. Biomolecular Investigations
3.3.1. DNA Extraction
3.3.2. 16S rDNA Polymerase Chain Reaction-Denaturing Gradient Gel Electrophoresis (PCR-DGGE)
3.3.3. Dual Index 16S rRNA Gene Amplicon Library Preparation and Bioinformatics Analysis
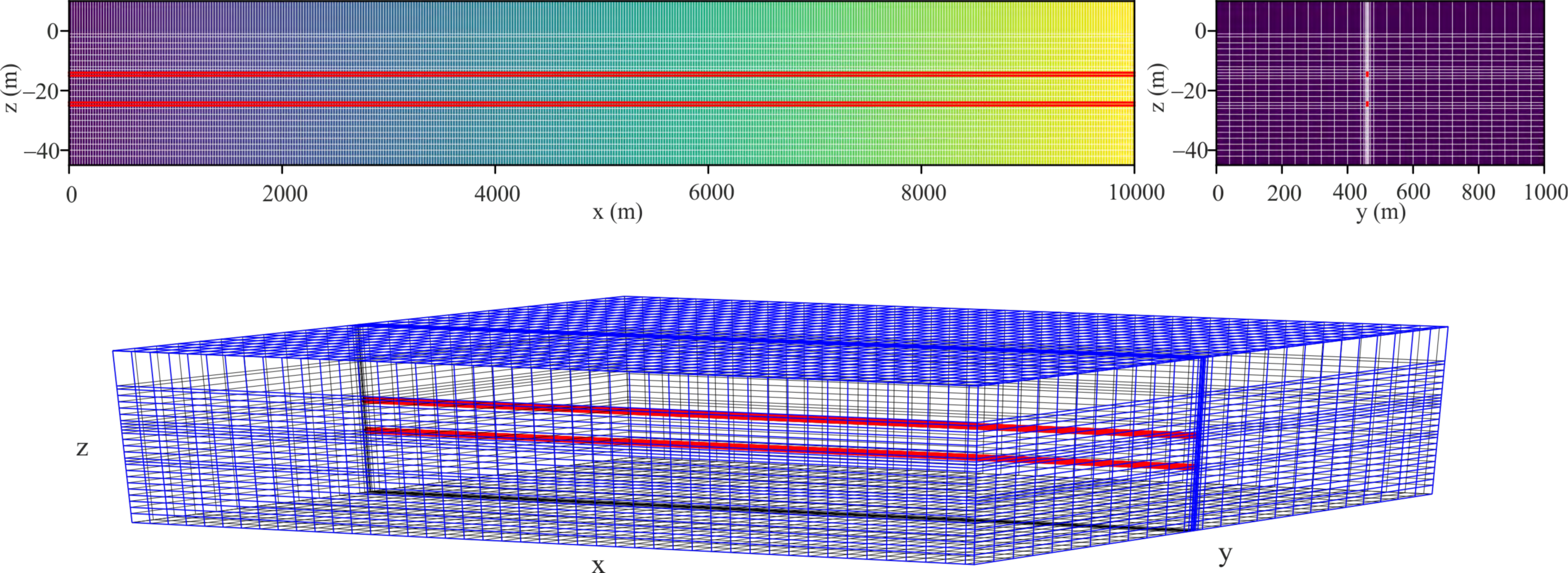
3.4. Groundwater Numerical Model
4. Results
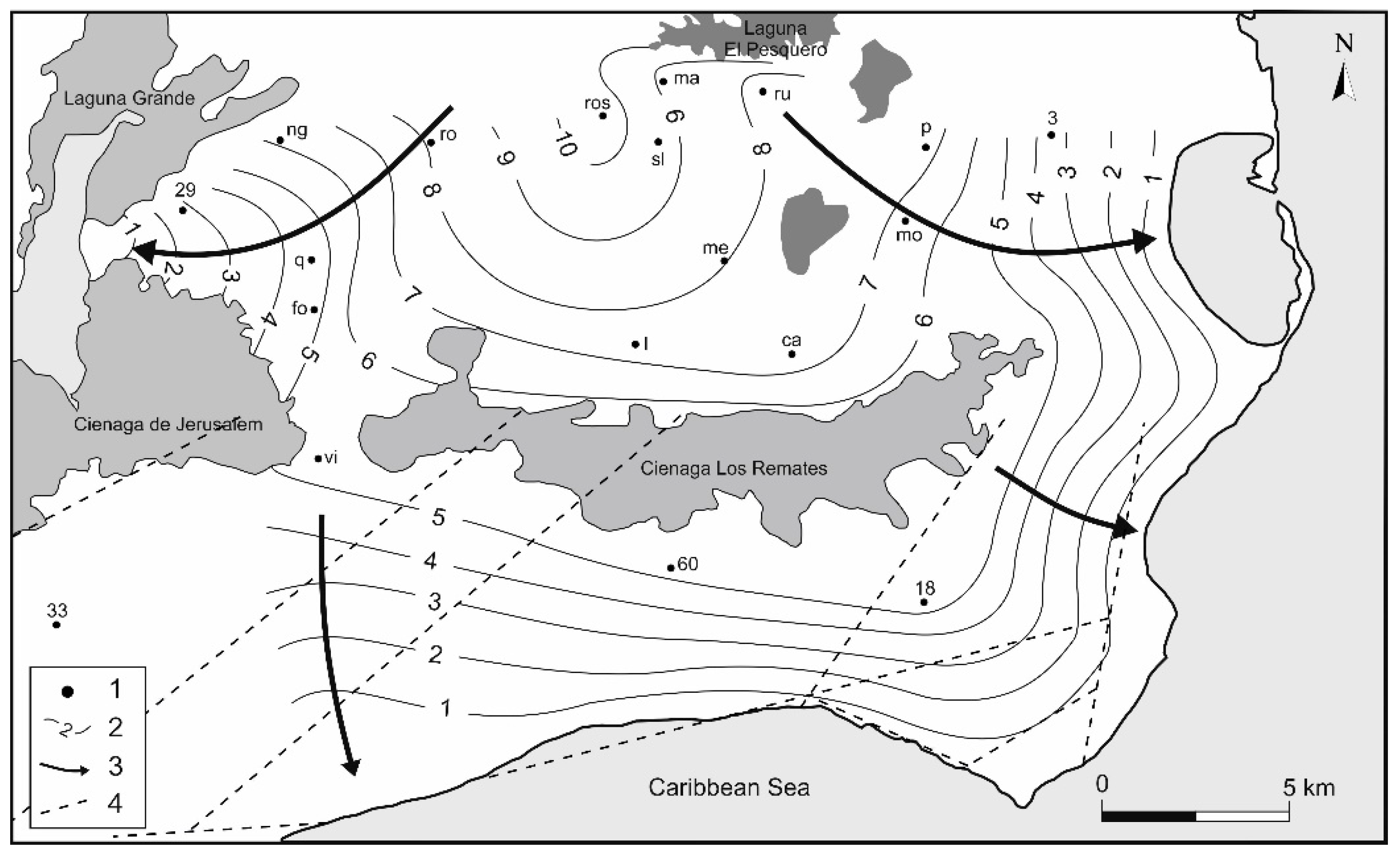
4.1. Groundwater Flow Pathway and Regime
4.2. Biomolecular Investigations
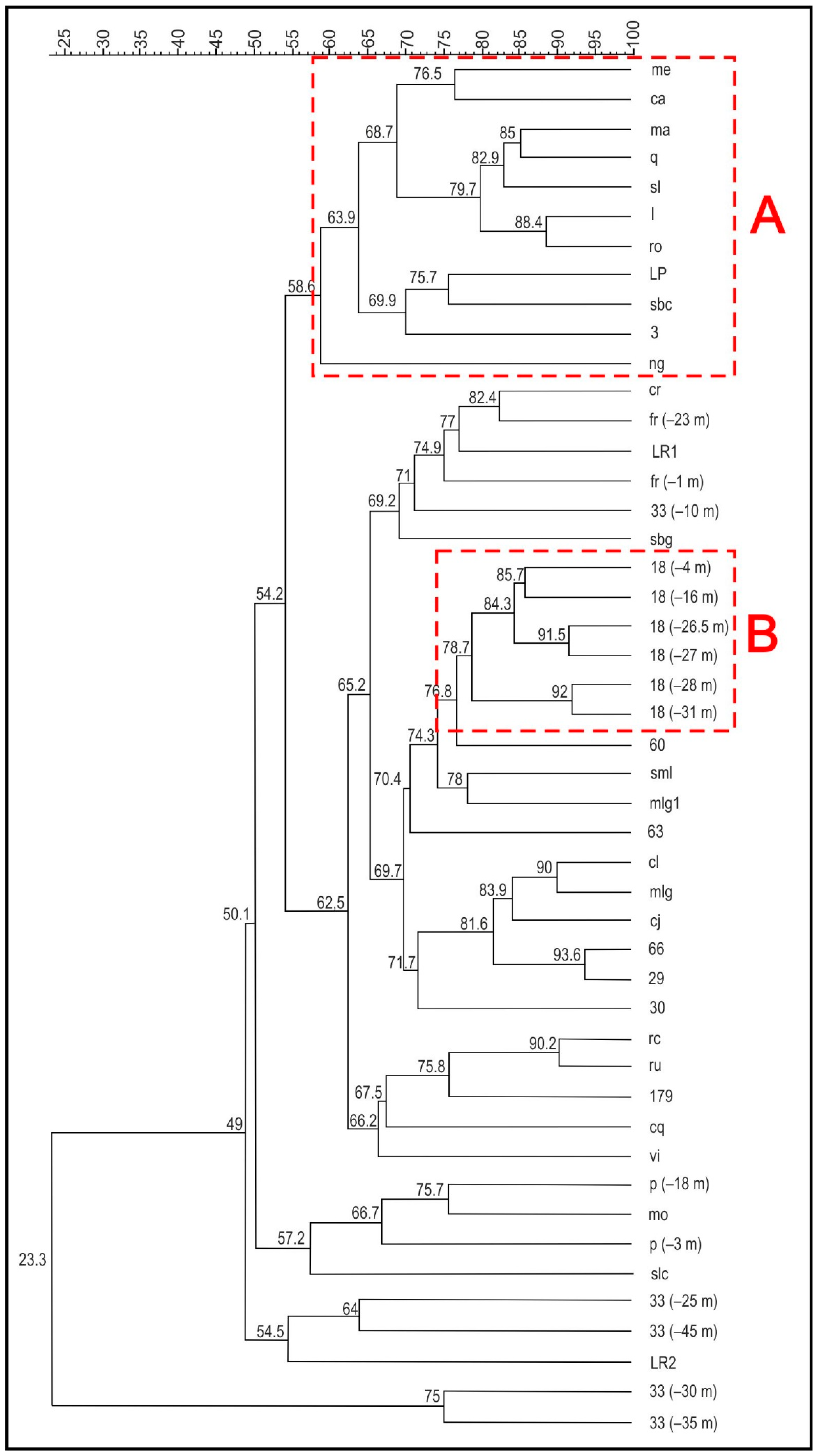
4.2.1. 16S rDNA PCR-DGGE Analysis of Sea-, Surface-, and Groundwater Microbial Communities
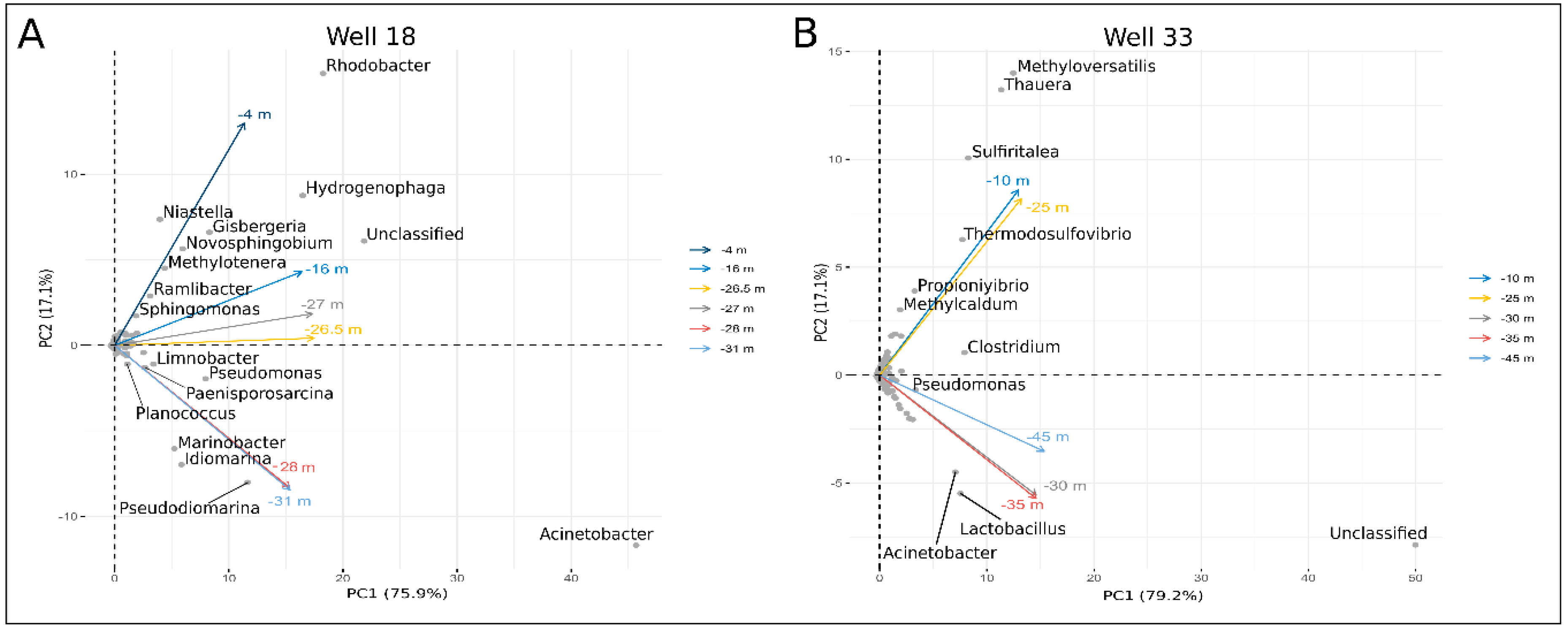
4.2.2. Next-Generation Sequencing Results
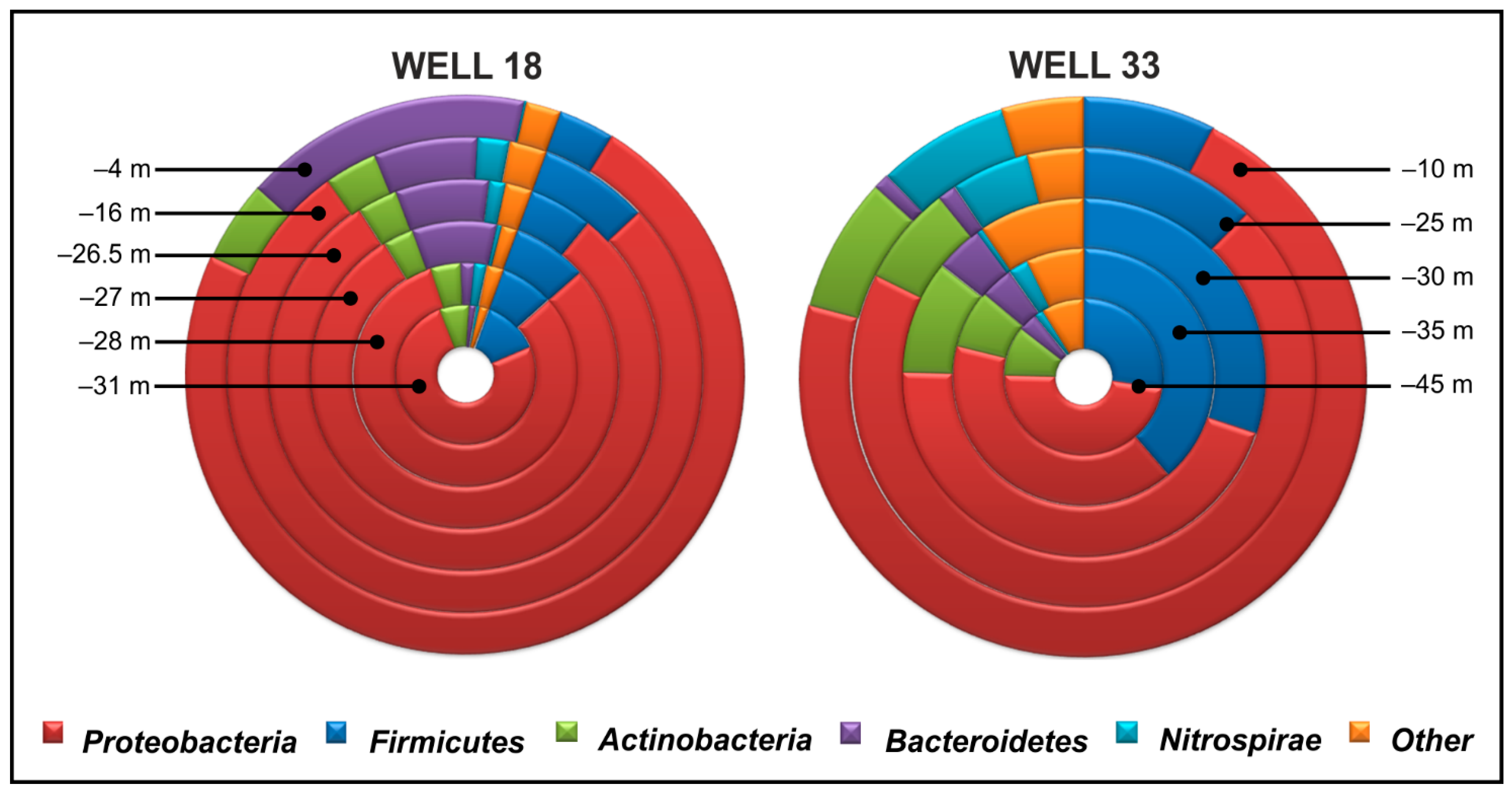
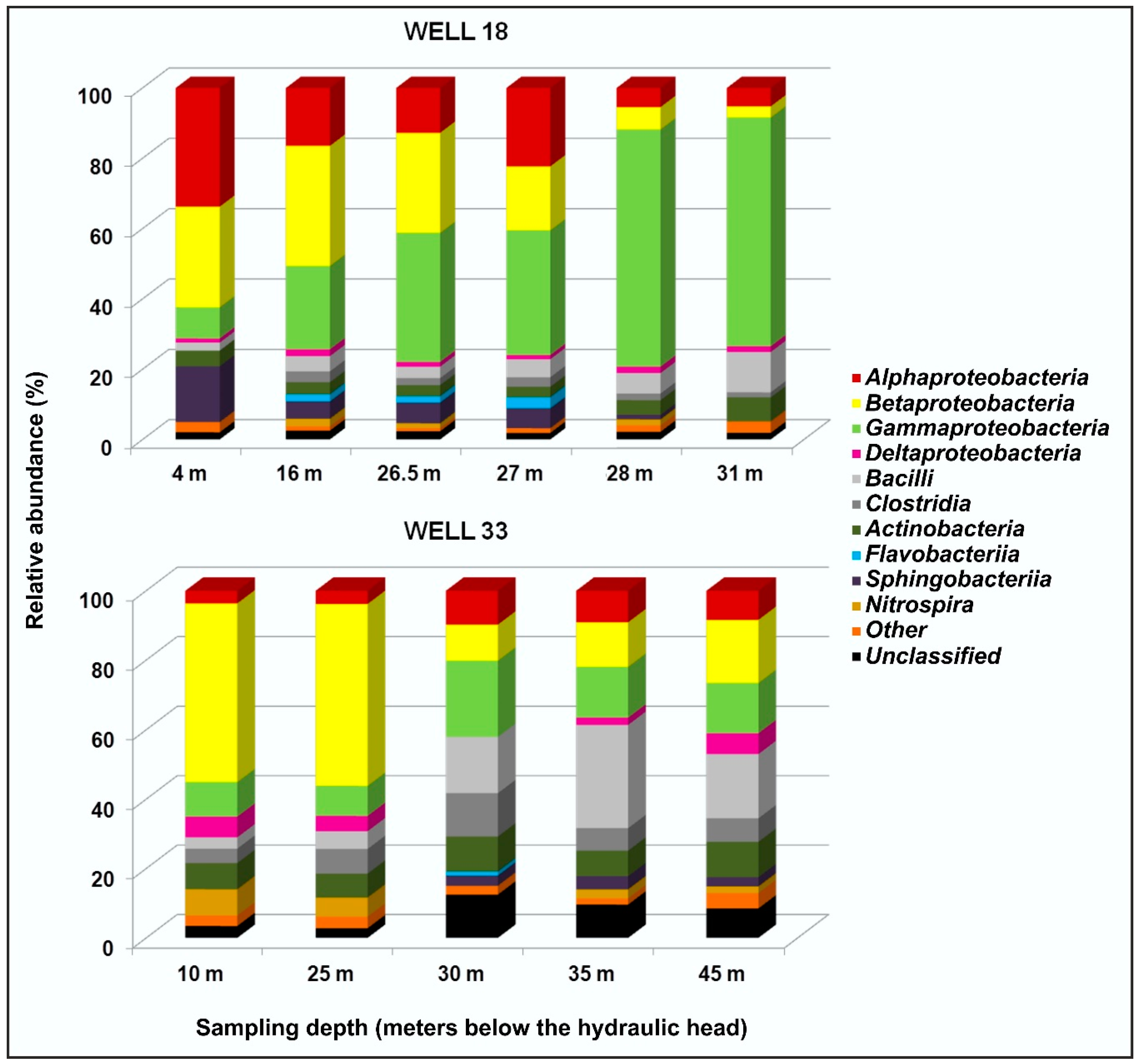
Microbial Communities in Well 18
Microbial Communities in Well 33
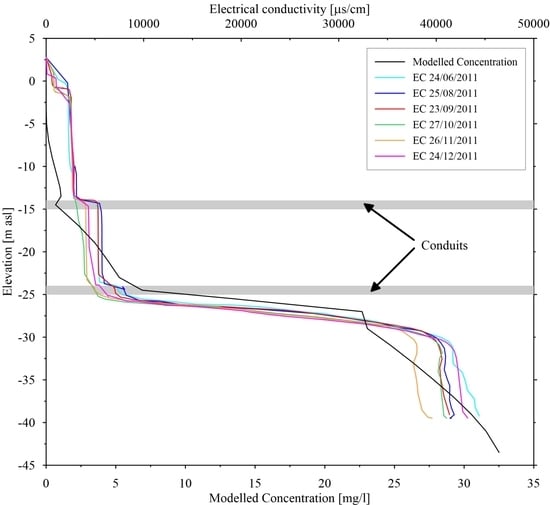
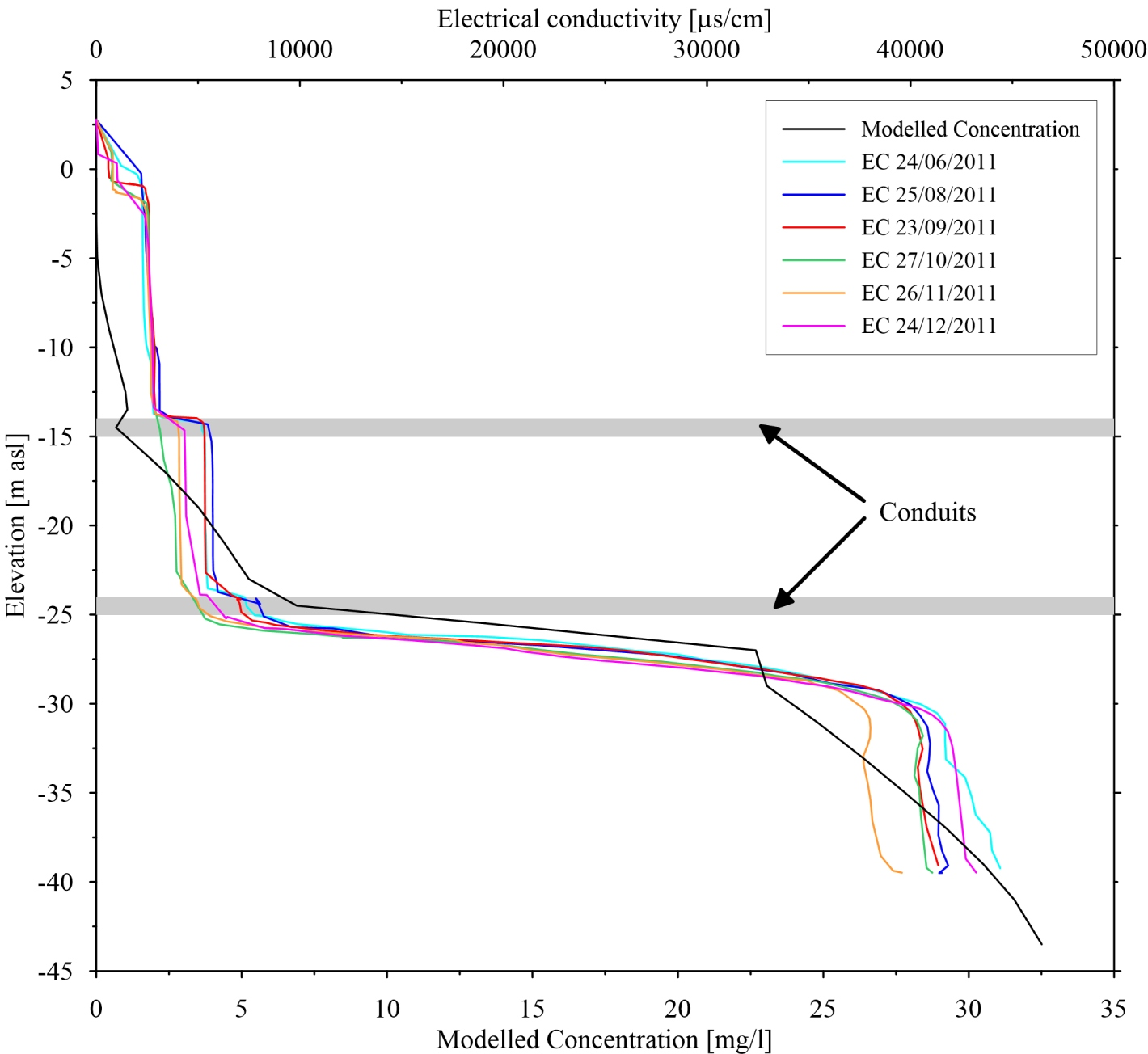
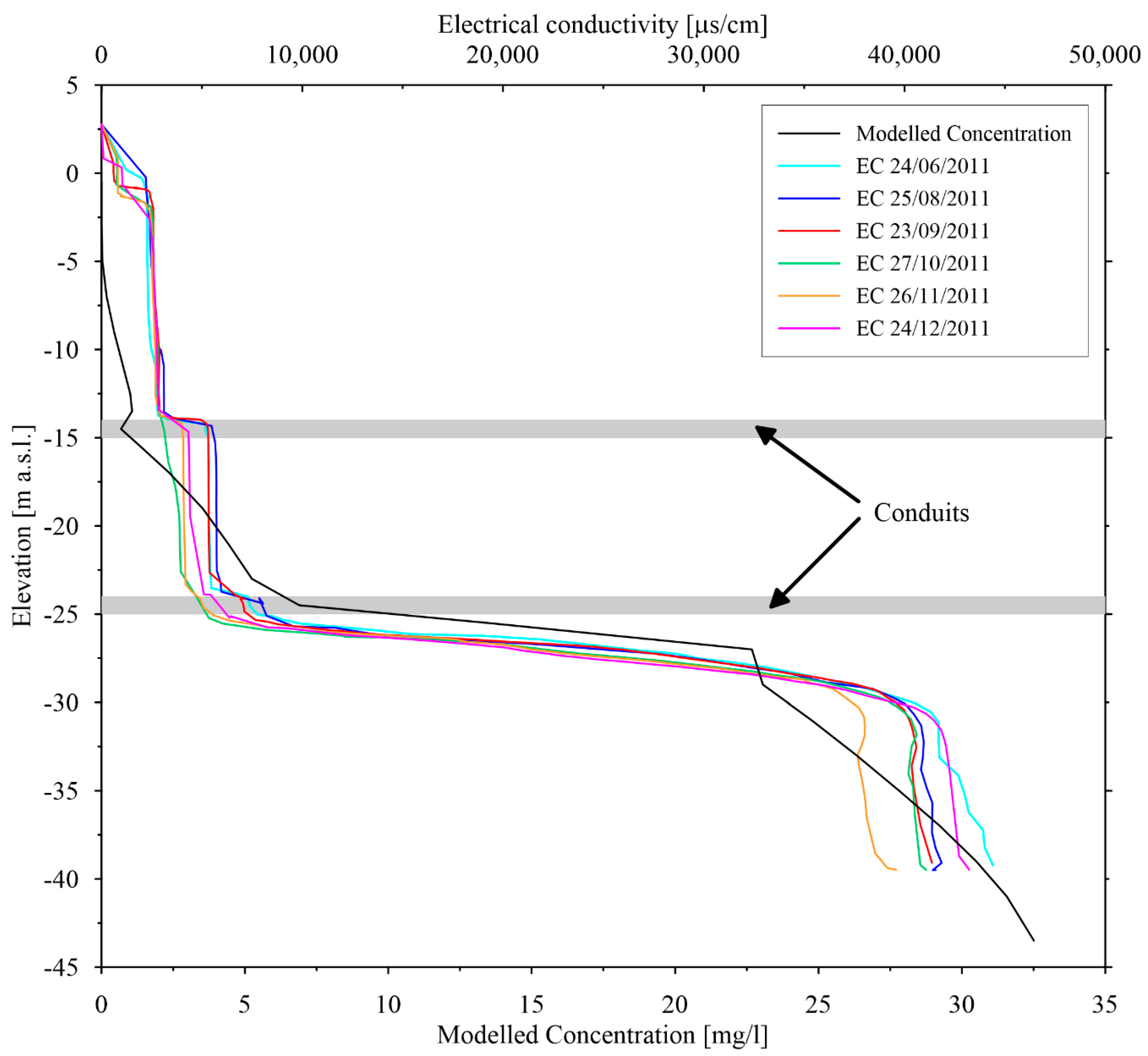
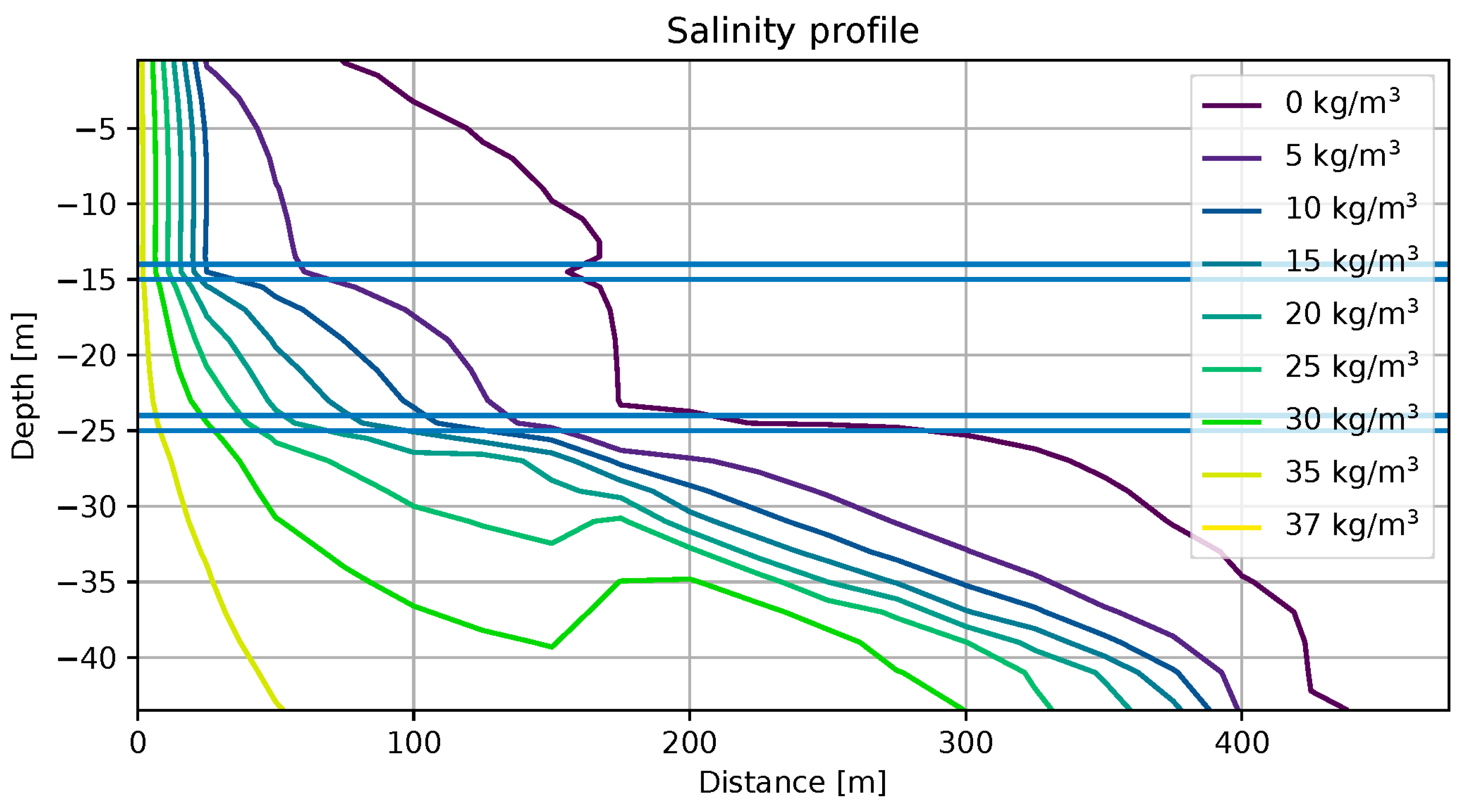
4.3. Saltwater Wedge Shape
4.4. Numerical Results
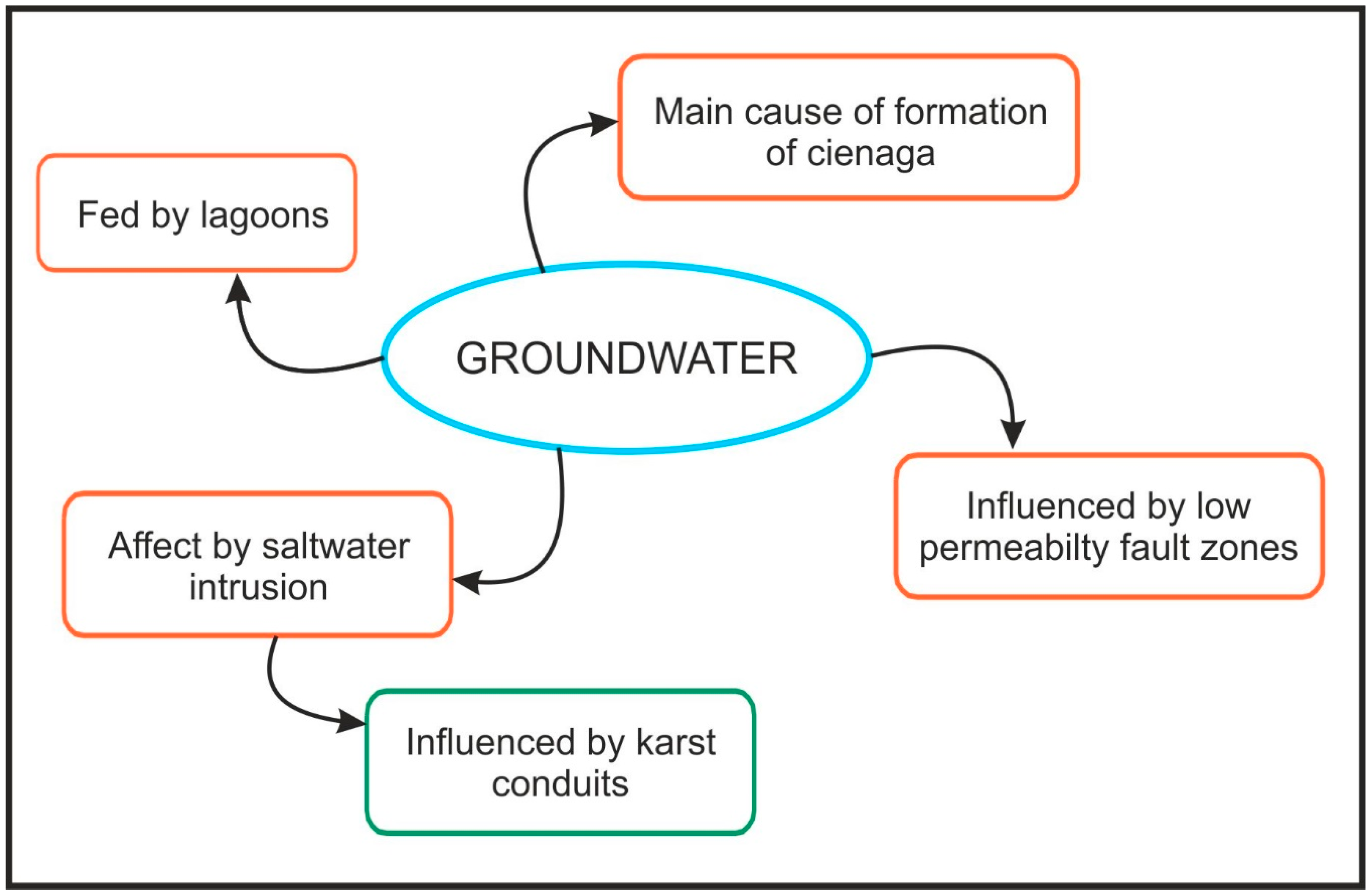
5. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Iturralde-Vinent, M.A.; Gutiérrez Domech, M.R. Some examples of karst development in Cuba. Bol. Inform. Com. Geoespeleol. 1999, 14, 1–4. [Google Scholar]
- Seale, L.D.; Soto, L.R.; Florea, L.J.; Fratesi, B. Karst of western Cuba: Observations, geomorphology and diagenesis. In Proceedings of the 12th Symposium on the Geology of the Bahamas and Other Carbonate Regions, San Salvador, Bahamas, 3–7 June 2004; pp. 1–9. [Google Scholar]
- Fagundo Castillo, J.R.; González Hernandez, P. Agricultural use and water quality at karstic Cuban western plain. Int. J. Speleol. 1999, 28, 175–185. [Google Scholar] [CrossRef][Green Version]
- Hernández, R.; Ramírez, R.; López-Portilla, M.; González, P.; Antigüedad, I.; Díaz, S. Seawater Intrusion in the Coastal Aquifer of Guanahacabibes, Pinar del Río, Cuba. In Management of Water Resources in Protected Areas; Farfán González, H., Corvea Porras, J.L., de Bustamante Gutiérrez, I., LaMoreaux, J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 301–308. [Google Scholar]
- Molerio León, L.; Parise, M. Managing environmental problems in Cuban karstic aquifers. Environ. Geol. 2009, 58, 275–283. [Google Scholar] [CrossRef]
- Brkić, Ž.; Kuhta, M.; Hunjak, T. Groundwater flow mechanism in the well-developed karst aquifer system in the western Croatia: Insights from spring discharge and water isotopes. Catena 2018, 161, 14–26. [Google Scholar] [CrossRef]
- Fleury, P.; Bakalowicz, M.; de Marsily, G. Submarine springs and coastal karst aquifers: A review. J. Hydrol. 2007, 339, 79–92. [Google Scholar] [CrossRef]
- Beddows, P.A.; Smart, P.L.; Whitaker, F.F.; Smith, S.L. Decoupled fresh–saline groundwater circulation of a coastal carbonate aquifer: Spatial patterns of temperature and specific electrical conductivity. J. Hydrol. 2007, 346, 18–32. [Google Scholar] [CrossRef]
- Boschetti, T.; Gonzales-Hernandez, P.; Hernandez-Diaz, R.; Naclerio, G.; Celico, F. Seawater intrusion in the Guanahacabibes Peninsula (Pinar del Rio Province, western Cuba): Effects on karst development and water isotope composition. Environ. Earth Sci. 2015, 73, 5703–5719. [Google Scholar] [CrossRef]
- Bucci, A.; Petrella, E.; Naclerio, G.; Allocca, V.; Celico, F. Microorganisms as contaminants and natural tracers: A 10-year research in some carbonate aquifers (southern Italy). Environ. Earth Sci. 2015, 74, 173–184. [Google Scholar] [CrossRef]
- Bucci, A.; Naclerio, G.; Allocca, V.; Celico, P.; Celico, F. Potential use of microbial community investigations to analyse hydrothermal systems behaviour: The case of Ischia Island, Southern Italy. Hydrol. Process. 2011, 25, 1866–1873. [Google Scholar] [CrossRef]
- Cabrera, M.; Peñalver, L.L. Contribución a la estratigrafía de los depósitos cuaternarios de Cuba. Cuatern. Geomorfol. 2001, 15, 37–49. [Google Scholar]
- Denis, R.; Díaz Guanche, C. Características Geologicas y Geomorfologicas de la Península Guanahacabibes; Dirección Provincial de Planificación Física: Pinar del Río, Cuba, 1993. [Google Scholar]
- Pulina, M.; Fagundo, J.R. Tropical karst and chemical denudation of Western Cuba. Geog. Pol. 1992, 60, 195–216. [Google Scholar]
- Chelli, A.; Pappalardo, M. Geomorphological mapping, a basic tool for interpreting rock coasts landforms: An example from Eastern Liguria (Italy). GFDQ 2008, 31, 13–19. [Google Scholar]
- Chelli, A.; Aringoli, D.; Aucelli, P.P.; Baldassarre, M.A.; Bellotti, P.; Bini, M.; Biolchi, S.; Bontempi, S.; Brandolini, P.; Davoli, L.; et al. Morphodynamics of coastal areas represented in the new geomorphologic map of Italy: Draw the landforms of the past to outline the future. Alp. Medit. Quat. 2018, 31, 17–21. [Google Scholar] [CrossRef]
- Mastronuzzi, G.; Aringoli, D.; Aucelli, P.; Baldassarre, M.A.; Bellotti, P.; Bini, M.; Biolchi, S.; Bontempi, S.; Brandolini, P.; Chelli, A.; et al. Geomorphological map of the Italian coast: From a descriptive to a morphodynamic approach. GFDQ 2017, 40, 161–195. [Google Scholar] [CrossRef]
- Ionin, A.S.; Pavlidis, Y.A. Condiciones de formación del relieve y los sedimentos de la Isla de la Juventud (Cuba). In Nuevas Investigaciones de los Procesos Costeros; Nauka: Moscow, Russia, 1977. [Google Scholar]
- Bucci, A.; Allocca, V.; Naclerio, G.; Capobianco, G.; Divino, F.; Fiorillo, F.; Celico, F. Winter survival of microbial contaminants in soil: An in situ verification. J. Environ. Sci. 2015, 27, 131–138. [Google Scholar] [CrossRef]
- Crescenzo, R.; Mazzoli, A.; Cancelliere, R.; Bucci, A.; Naclerio, G.; Baccigalupi, L.; Cutting, S.M.; Ricca, E.; Iossa, S. Beneficial effects of carotenoid-producing cells of Bacillus indicus HU16 in a rat model of diet-induced metabolic syndrome. Benef. Microbes 2017, 8, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Di Luccia, B.; Mazzoli, A.; Cancelliere, R.; Crescenzo, R.; Ferrandino, I.; Monaco, A.; Bucci, A.; Naclerio, G.; Iossa, S.; Ricca, E.; et al. Lactobacillus gasseri SF1183 protects the intestinal epithelium and prevents colitis symptoms in vivo. J. Funct. Foods 2018, 42, 195–202. [Google Scholar] [CrossRef]
- Pietrangelo, L.; Bucci, A.; Maiuro, L.; Bulgarelli, D.; Naclerio, G. Unraveling the composition of the root-associated bacterial microbiota of Phragmites australis and Typha latifolia. Front. Microbiol. 2018, 9, 1650. [Google Scholar] [CrossRef]
- Bucci, A.; Petrella, E.; Celico, F.; Naclerio, G. Use of molecular approaches in hydrogeological studies: The case of carbonate aquifers in southern Italy. Hydrogeol. J. 2017, 25, 1017–1031. [Google Scholar] [CrossRef]
- Chong, C.W.; Dunn, M.J.; Convey, P.; Tan, G.Y.A.; Wong, R.C.S.; Tan, I.K.P. Environmental influences on bacterial diversity of soils on Signy Island, maritime Antarctic. Polar Biol. 2009, 32, 1571–1582. [Google Scholar] [CrossRef]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 2002, 148, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Microbiol. Biotechnol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Harbaugh, A.W.; Banta, E.R.; Hill, M.C.; McDonald, M.G. MODFLOW-2000, the U.S. Geological Survey Modular Ground-Water Model-User Guide to Modularization Concepts and the Groundwater Flow Process; U.S. Geological Survey Open-File Report 00-92; U.S. Geological Survey: Reston, VA, USA, 2000; p. 121.
- Langevin, C.; Shoemaker, W.B.; Guo, W. MODFLOW-2000, the US Geological Survey Modular Ground-Water Model Documentation of the SEAWAT-2000 Version with the Variable-Density Flow Process (VDF) and the Integrated MT3DMS Transport Process (IMT); US Department of the Interior: Washington, DC, USA; US Geological Survey: Reston, VA, USA, 2003.
- Zheng, C. MT3DMS v5.2 Supplemental User’s Guide: Technical Report to the U.S. Army; Engineer Research and Development Center, Department of Geological Sciences, University of Alabama: Tuscaloosa, AL, USA, 2006; 24p. [Google Scholar]
- Zheng, C.; Wang, P.P. MT3DMS—A Modular Three-Dimensional Multispecies Transport Model for Simulation of Advection, Dispersion and Chemical Reactions of Contaminants in Ground-Water Systems: Documentation and User’s Guide: U.S. Army; Corps of Engineers Contract Report SERDP-99-1; Corps of Engineers: Washington, DC, USA, 1999.
- Petrella, E.; Falasca, A.; Celico, F. Natural-gradient tracer experiment in epikarst: A test study in the Acqua dei Faggi experimental site, southern Italy. Geofluids 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Xue, Y.Q.; Wu, J.C. Groundwater Hydraulics, 2nd ed.; Geological Publishing House: Beijing, China, 2010; pp. 14–15. [Google Scholar]
- Xu, Z.; Hu, B.X.; Ye, M. Numerical modeling and sensitivity analysis of seawater intrusion in a dual-permeability coastal karst aquifer with conduit networks. Hydrol. Earth Syst. Sci. 2018, 22, 221–239. [Google Scholar] [CrossRef]
- Bakker, M.; Post, V.; Langevin, C.D.; Hughes, J.D.; White, J.T.; Starn, J.J.; Fienen, M.N. Scripting MODFLOW Model Development Using Python and FloPy. Groundwater 2016, 54, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Drogue, C. Hydrodynamics of karst aquifers: Experimental sites in the Mediterranean karst, Southern France. Int. Contrib. Hydrogeol. IAH 1992, 13, 133–149. [Google Scholar]
- Aquino, D.; Petrella, E.; Florio, T.; Celico, P.; Celico, F. Complex hydraulic interactions between compartmentalized carbonate aquifers and heterogeneous siliciclastic successions: A case study in southern Italy. Hydrol. Process. 2015, 29, 4252–4263. [Google Scholar] [CrossRef]
- Bucci, A.; Petrella, E.; Naclerio, G.; Gambatese, S.; Celico, F. Bacterial migration through low-permeability fault zones in compartmentalised aquifer systems: A case study in Southern Italy. Int. J. Speleol. 2014, 43, 273–281. [Google Scholar] [CrossRef]
- Celico, F.; Petrella, E.; Celico, P. Hydrogeological behaviour of some fault zones in a carbonate aquifer of Southern Italy: An experimentally based model. Terra Nova 2006, 18, 308–313. [Google Scholar] [CrossRef]
- Giuffrida, A.; La Bruna, V.; Castelluccio, P.; Panza, E.; Rustichelli, A.; Tondi, E.; Giorgini, M.; Agosta, F. Fracture simulation parameters of fractured reservoirs: Analogy with outcropping carbonates of the Inner Apulian Platform, southern Italy. J. Struct. Geol. 2019, 123, 18–41. [Google Scholar] [CrossRef]
- Petrella, E.; Capuano, P.; Carcione, M.; Celico, F. A high-altitude temporary spring in a compartmentalized carbonate aquifer: The role of low-permeability faults and karst conduits. Hydrol. Process. 2009, 23, 3354–3364. [Google Scholar] [CrossRef]
- Petrella, E.; Aquino, D.; Fiorillo, F.; Celico, F. The effect of low-permeability fault zones on groundwater flow in a compartmentalized system. Experimental evidence from a carbonate aquifer (Southern Italy). Hydrol. Process. 2015, 29, 1577–1587. [Google Scholar] [CrossRef]
- Storti, F.; Billi, A.; Salvini, F. Particle size distributions in natural carbonate fault rocks: Insight for non-self-similar cataclasis. Earth Planet. Sci. Lett. 2003, 206, 173–186. [Google Scholar] [CrossRef]
- Storti, F.; Balsamo, F.; Cappanera, F.; Tosi, G. Sub-seismic scale fracture pattern and in situ permeability data in the chalk atop of the Krempe salt ridge at Lagerdorf, NW Germany: Inferences on synfolding stress field evolution and its impact on fracture connectivity. Mar. Pet. Geol. 2011, 28, 1315–1332. [Google Scholar] [CrossRef]
- Morris, R.M.; Rappé, M.S.; Connon, S.A.; Vergin, K.L.; Siebold, W.A.; Carlson, C.A.; Giovannoni, S.J. SAR11 clade dominates ocean surface bacterioplankton communities. Nature 2002, 420, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Gerea, M.; Queimaliños, C.; Schiaffino, M.R.; Izaguirre, I.; Forn, I.; Massana, R.; Unrein, F. In situ prey selection of mixotrophic and heterotrophic flagellates in Antarctic oligotrophic lakes: An analysis of the digestive vacuole content. J. Plankton Res. 2013, 35, 201–212. [Google Scholar] [CrossRef]
- Glöckner, F.O.; Fuchs, B.M.; Amann, R. Bacterioplankton compositions of lakes and oceans: A first comparison based on fluorescence in situ hybridization. Appl. Environ. Microbiol. 1999, 65, 3721–3726. [Google Scholar]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef]
- Salcher, M.M.; Pernthaler, J.; Frater, N.; Posch, T. Vertical and longitudinal distribution patterns of different bacterioplankton populations in a canyon-shaped, deep prealpine lake. Limnol. Oceanogr. 2011, 56, 2027–2039. [Google Scholar] [CrossRef]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling Expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef]
- Wu, Q.L.; Zwart, G.; Schauer, M.; Kamst-van Agterveld, M.P.; Hahn, M.W. Bacterioplankton community composition along a salinity gradient of sixteen high-mountain lakes located on the Tibetan Plateau, China. Appl. Environ. Microbiol. 2006, 72, 5478–5485. [Google Scholar] [CrossRef]
- Macagnan, D.; Romeiro, R.D.S.; de Souza, J.T.; Pomella, A.W.V. Isolation of actinomycetes and endospore-forming bacteria from the cacao pod surface and their antagonistic activity against the witches’ broom and black pod pathogens. Phytoparasitica 2006, 34, 122–132. [Google Scholar] [CrossRef]
- Henry, E.A.; Devereux, R.; Maki, J.S.; Gilmour, C.C.; Woese, C.R.; Mandelco, L.; Schauder, R.; Remsen, C.C.; Mitchell, R. Characterization of a new thermophilic sulfate-reducing bacterium Thermodesulfovibrio yellowstonii, gen. nov. and sp. nov.: Its phylogenetic relationship to Thermodesulfobacterium commune and their origins deep within the bacterial domain. Arch. Microbiol. 1994, 161, 62–69. [Google Scholar] [CrossRef]
- Sonne-Hansen, J.; Ahring, B.K. Thermodesulfobacterium hveragerdense sp. nov., and Thermodesulfovibrio islandicus sp. nov., two thermophilic sulfate reducing bacteria isolated from a Icelandic hot spring. Syst. Appl. Microbiol. 1999, 22, 559–564. [Google Scholar] [CrossRef]
- Haouari, O.; Fardeau, M.L.; Cayol, J.L.; Fauque, G.; Casiot, C.; Elbaz-Poulichet, F.; Hamdi, M.; Ollivier, B. Thermodesulfovibrio hydrogeniphilus sp. nov., a new thermophilic sulphate-reducing bacterium isolated from a Tunisian hot spring. Syst. Appl. Microbiol. 2008, 31, 38–42. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Muramatsu, M.; Imachi, H.; Narihiro, T.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y. Thermodesulfovibrio aggregans sp. nov. and Thermodesulfovibrio thiophilus sp. nov., anaerobic, thermophilic, sulfate-reducing bacteria isolated from thermophilic methanogenic sludge, and emended description of the genus Thermodesulfovibrio. Int. J. Syst. Evol. Microbiol. 2008, 58, 2541–2548. [Google Scholar] [CrossRef]
- Kimura, H.; Sugihara, M.; Yamamoto, H.; Patel, B.K.; Kato, K.; Hanada, S. Microbial community in a geothermal aquifer associated with the subsurface of the Great Artesian Basin, Australia. Extremophiles 2005, 9, 407–414. [Google Scholar] [CrossRef]
- Nazina, T.N.; Shestakova, N.M.; Grigor’ian, A.A.; Mikhaĭlova, E.M.; Turova, T.P.; Poltaraus, A.B.; Feng, C.; Ni, F.; Beliaev, S.S. Phylogenetic diversity and activity of anaerobic microorganisms of high-temperature horizons of the Dagang Oilfield (China). Mikrobiologiia 2006, 75, 70–81. [Google Scholar] [CrossRef]
- Wang, L.Y.; Ke, W.J.; Sun, X.B.; Liu, J.F.; Gu, J.D.; Mu, B.Z. Comparison of bacterial community in aqueous and oil phases of water-flooded petroleum reservoirs using pyrosequencing and clone library approaches. Appl. Microbiol. Biotechnol. 2014, 98, 4209–4221. [Google Scholar] [CrossRef]
- Yang, G.C.; Zhou, L.; Mbadinga, S.M.; Liu, J.F.; Yang, S.Z.; Gu, J.D.; Mu, B.Z. Formate-dependent microbial conversion of CO2 and the dominant pathways of methanogenesis in production water of high-temperature oil reservoirs amended with bicarbonate. Front. Microbiol. 2016, 7, 365. [Google Scholar] [CrossRef]
- Konno, U.; Kouduka, M.; Komatsu, D.D.; Ishii, K.; Fukuda, A.; Tsunogai, U.; Ito, K.; Suzuki, Y. Novel microbial populations in deep granitic groundwater from Grimsel Test Site, Switzerland. Microb. Ecol. 2013, 65, 626–637. [Google Scholar] [CrossRef]
- Frank, Y.A.; Kadnikov, V.V.; Lukina, A.P.; Banks, D.; Beletsky, A.V.; Mardanov, A.V.; Sen’kina, E.I.; Avakyan, M.R.; Karnachuk, O.V.; Ravin, N.V. Characterization and genome analysis of the first facultatively alkaliphilic Thermodesulfovibrio isolated from the deep terrestrial subsurface. Front. Microbiol. 2016, 7, 2000. [Google Scholar] [CrossRef]
- Garrity, G.M.; Holt, J.G. Phylum BVIII. Nitrospirae phy. Nov. In Bergey’s Manual of Systematic Bacteriology; Boone, D.R., Castenholz, R.W., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2013; pp. 451–464. [Google Scholar]
- Cook, P.G.; Love, A.J.; Dighton, J.C. Inferring groundwater flow in fractured rock from dissolved radon. Ground Water 1999, 37, 606–610. [Google Scholar] [CrossRef]
- Morin, R.H.; Carleton, G.B.; Poirier, S. Fractured-aquifer hydrogeology from geophysical logs; the Passaic Formation, New Jersey. Ground Water 1997, 35, 328–338. [Google Scholar] [CrossRef]
- Petrella, E.; Naclerio, G.; Falasca, A.; Bucci, A.; Capuano, P.; De Felice, V.; Celico, F. Non-permanent shallow halocline in a fractured carbonate aquifer, southern Italy. J. Hydrol. 2009, 373, 267–272. [Google Scholar] [CrossRef]
- Petrella, E.; Celico, F. Mixing of water in a carbonate aquifer, southern Italy, analysed through stable isotope investigations. Int. J. Speleol. 2013, 42, 25–33. [Google Scholar] [CrossRef]













© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernàndez-Diaz, R.; Petrella, E.; Bucci, A.; Naclerio, G.; Feo, A.; Sferra, G.; Chelli, A.; Zanini, A.; Gonzalez-Hernandez, P.; Celico, F. Integrating Hydrogeological and Microbiological Data and Modelling to Characterize the Hydraulic Features and Behaviour of Coastal Carbonate Aquifers: A Case in Western Cuba. Water 2019, 11, 1989. https://doi.org/10.3390/w11101989
Hernàndez-Diaz R, Petrella E, Bucci A, Naclerio G, Feo A, Sferra G, Chelli A, Zanini A, Gonzalez-Hernandez P, Celico F. Integrating Hydrogeological and Microbiological Data and Modelling to Characterize the Hydraulic Features and Behaviour of Coastal Carbonate Aquifers: A Case in Western Cuba. Water. 2019; 11(10):1989. https://doi.org/10.3390/w11101989
Chicago/Turabian StyleHernàndez-Diaz, Rebeca, Emma Petrella, Antonio Bucci, Gino Naclerio, Alessandra Feo, Gabriella Sferra, Alessandro Chelli, Andrea Zanini, Patricia Gonzalez-Hernandez, and Fulvio Celico. 2019. "Integrating Hydrogeological and Microbiological Data and Modelling to Characterize the Hydraulic Features and Behaviour of Coastal Carbonate Aquifers: A Case in Western Cuba" Water 11, no. 10: 1989. https://doi.org/10.3390/w11101989
APA StyleHernàndez-Diaz, R., Petrella, E., Bucci, A., Naclerio, G., Feo, A., Sferra, G., Chelli, A., Zanini, A., Gonzalez-Hernandez, P., & Celico, F. (2019). Integrating Hydrogeological and Microbiological Data and Modelling to Characterize the Hydraulic Features and Behaviour of Coastal Carbonate Aquifers: A Case in Western Cuba. Water, 11(10), 1989. https://doi.org/10.3390/w11101989