Bacterial Productivity in a Ferrocyanide-Contaminated Aquifer at a Nuclear Waste Site

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Sampling Procedure
2.2. Tritiated Leucine Incorporation Assay
2.3. Microbial Cell Numbers
2.4. Aqueous Speciation Modeling
2.5. Statistical and Geospatial Analyses
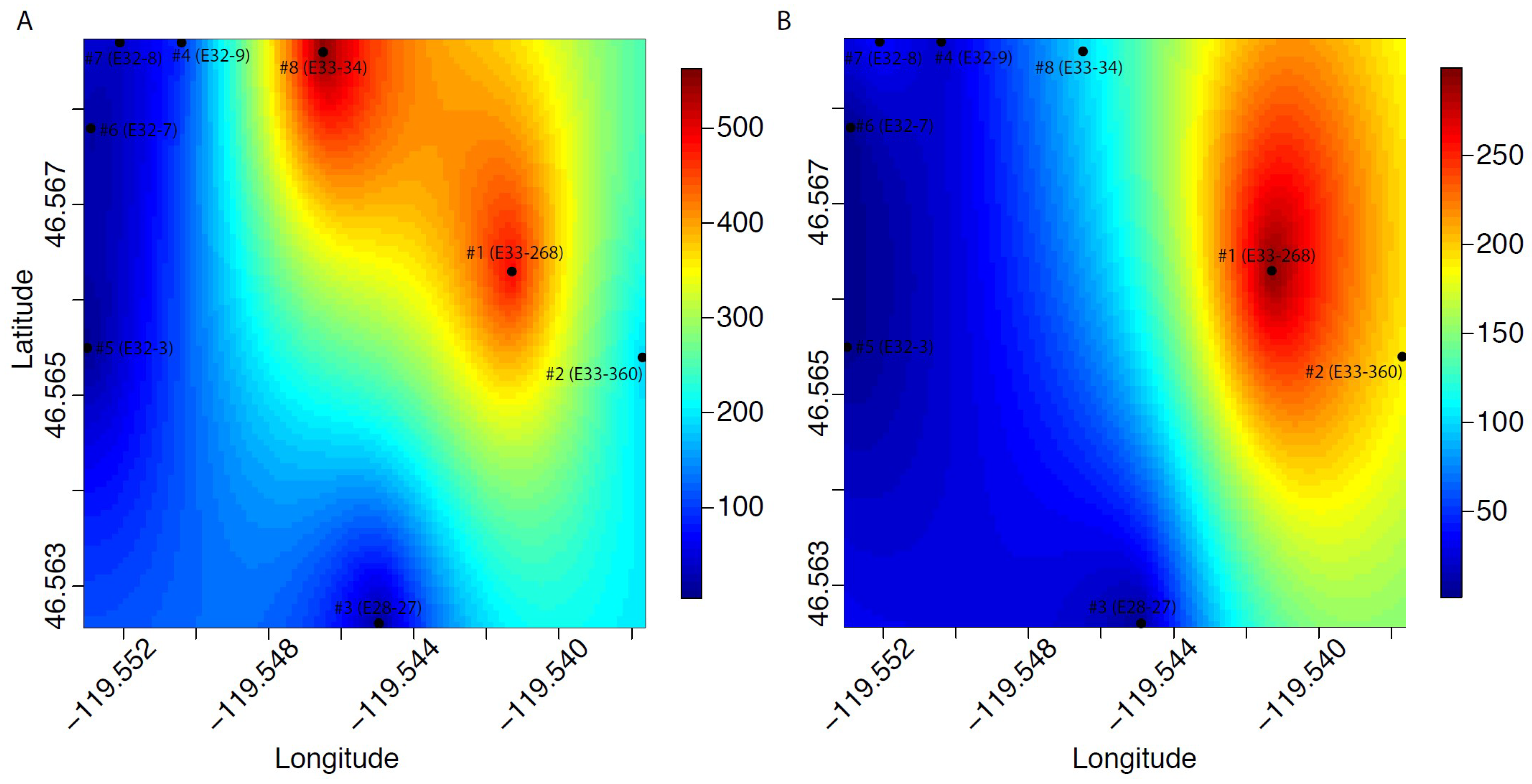
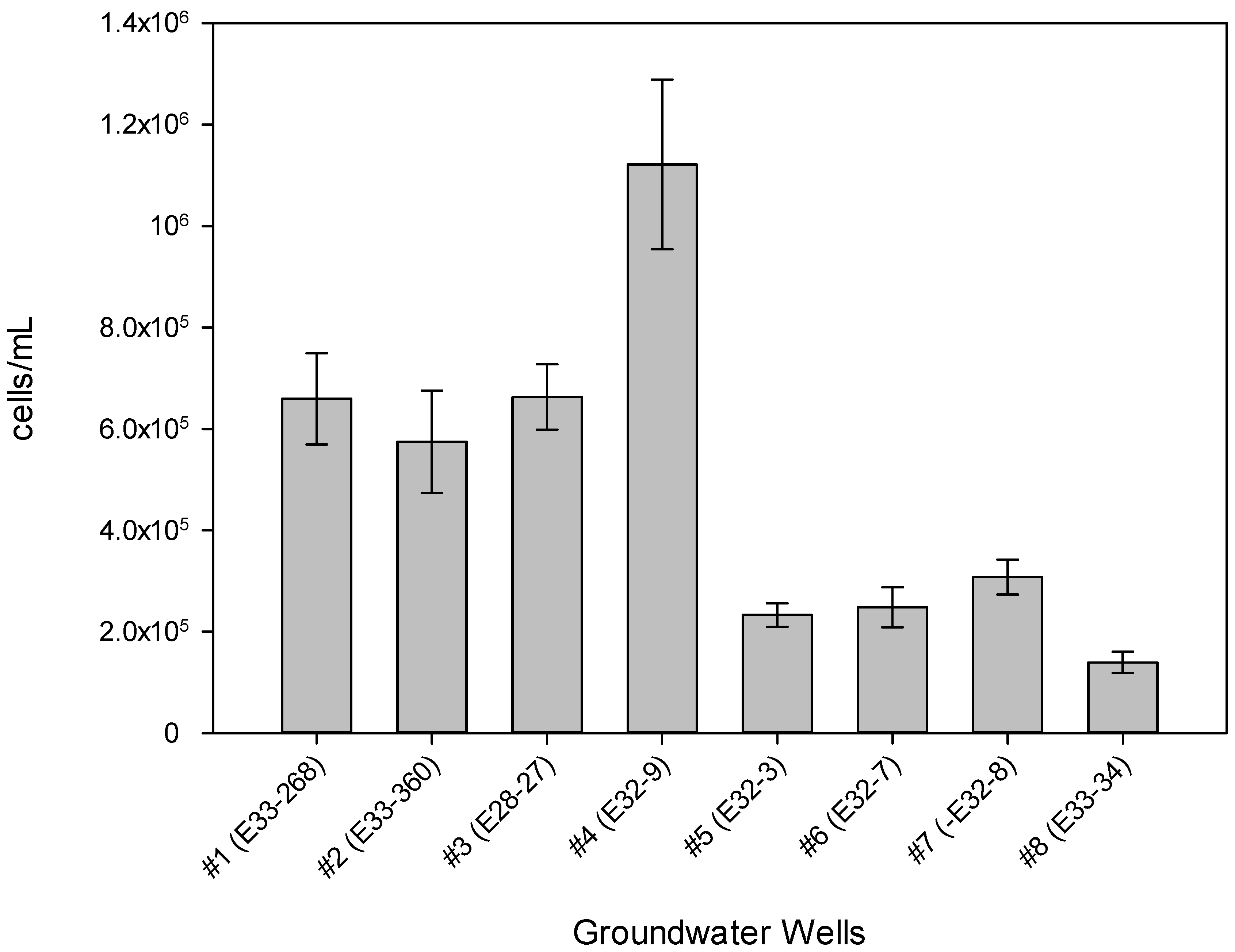
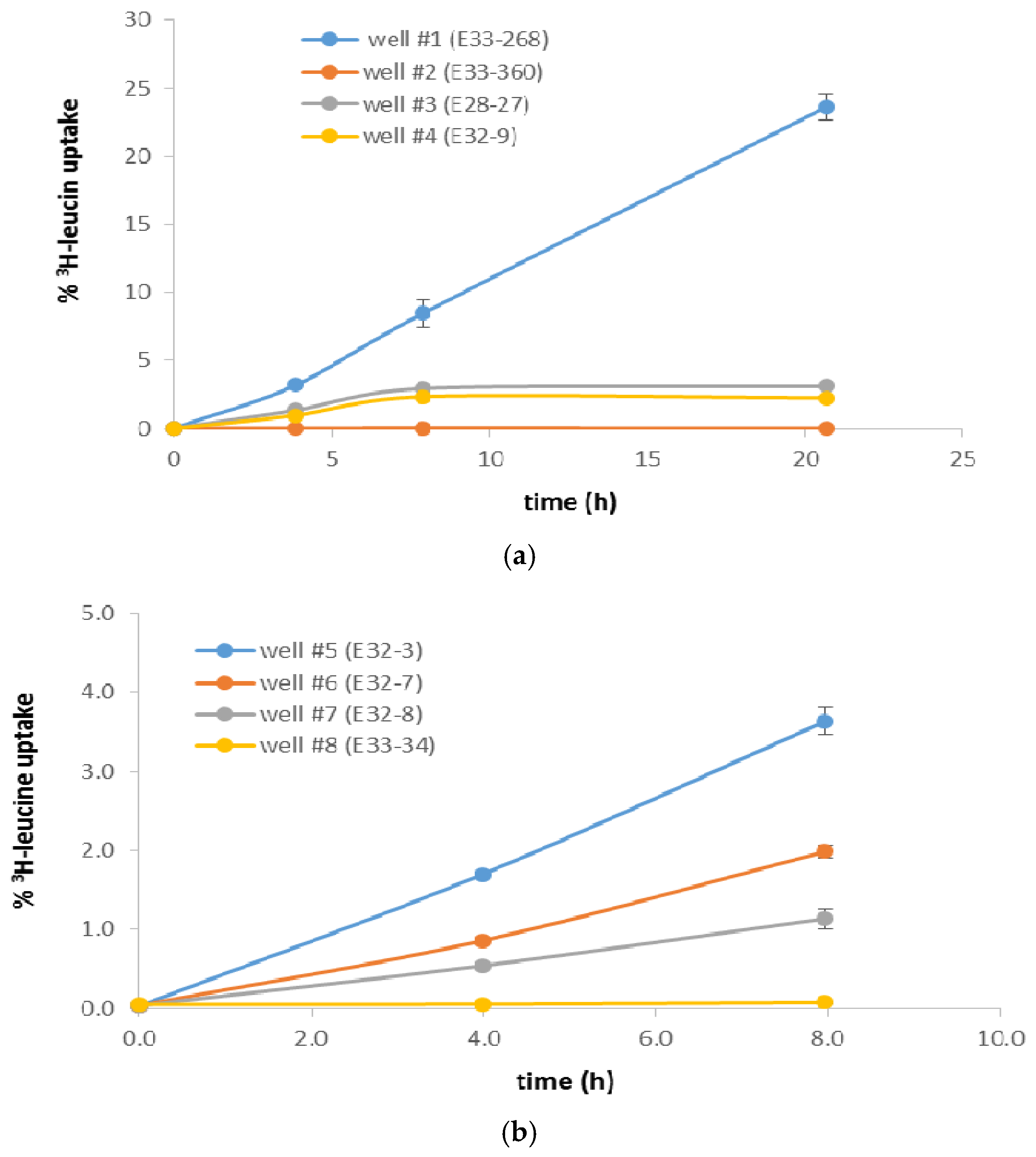
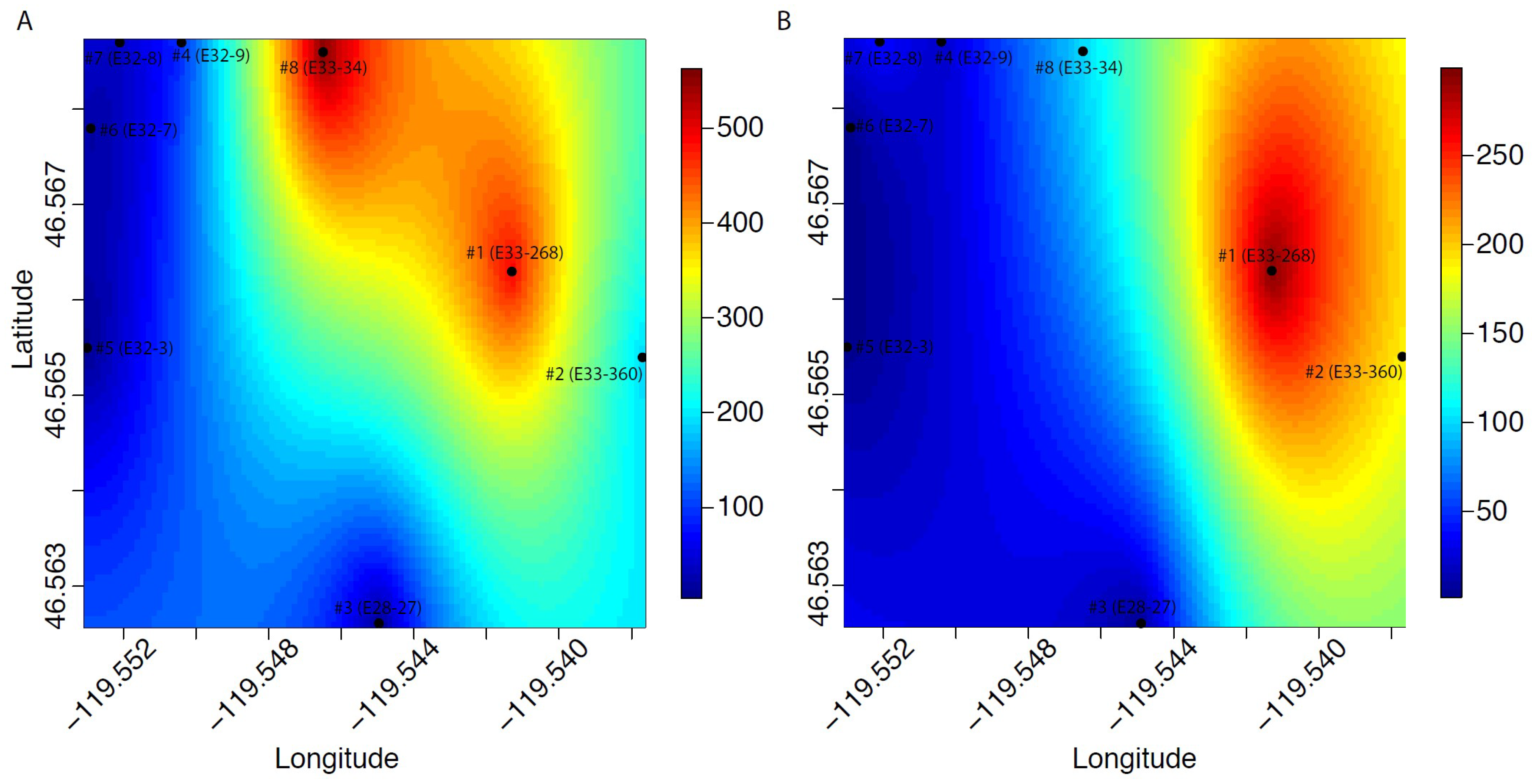
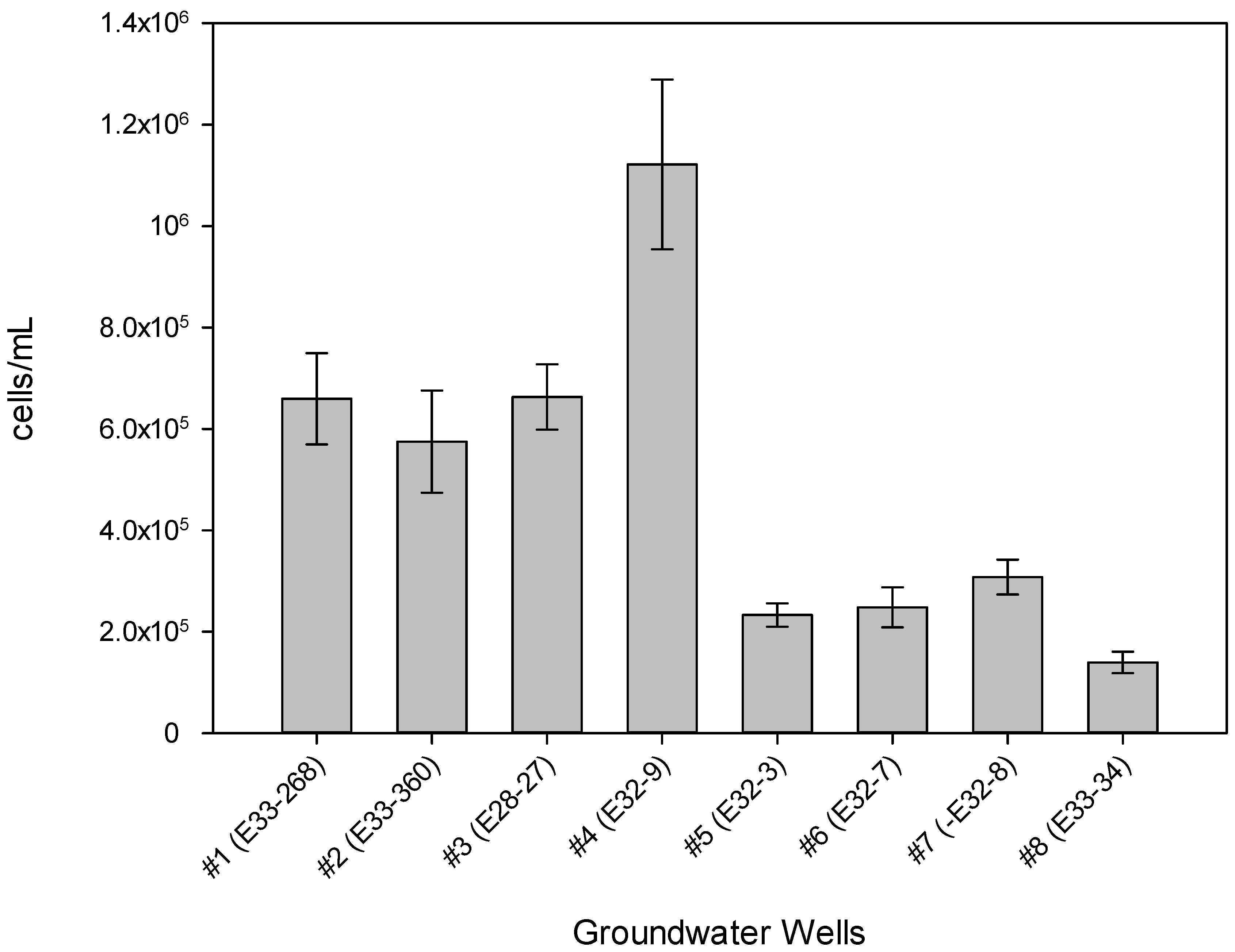
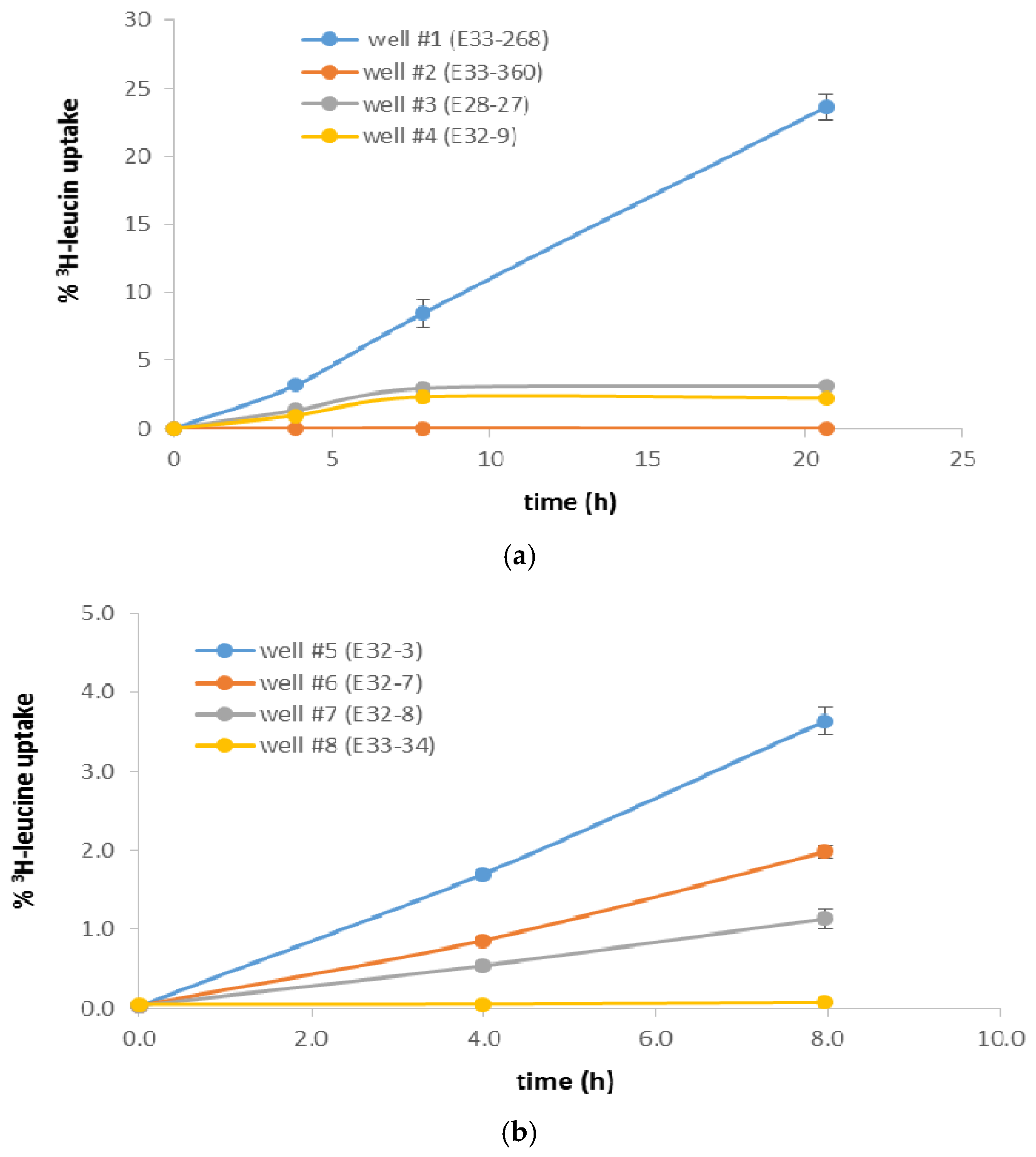
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gephart, R.E. A short history of waste management at the Hanford Site. Phys. Chem. Earth. 2010, 35, 298–306. [Google Scholar] [CrossRef]
- Hoitink, D.J.; Burk, K.W.; Ramsdell, J.V., Jr.; Shaw, W.J. Hanford Site Climatological Summary 2004 with Historical Data; Pacific Northwest National Laboratory: Richland, WA, USA, 2005; pp. 1–382. [Google Scholar]
- Fayer, M.J.; Keller, J.M. Recharge Data Package for Hanford Single-Shell Tank Waste Management Areas; PNNL-16688; Pacific Northwest National Laboratory: Richland, WA, USA, 2007. [Google Scholar]
- Hanford Groundwater Monitoring Report for 2016; DOE/RL-2016-67; CH2M Hill Plateau Remediation Company: Richland, WA, USA, 2017.
- Remedial Investigation Report for the 200-BP-5 Groundwater Operable Unit; DOE/RL-2009-127; CH2M Hill Plateau Remediation Company: Richland, WA, USA, 2015; pp. 1–424.
- Cash, R.J.; Meacham, J.E.; Lilga, M.A.; Babad, H. Resolution of the Hanford Site Ferrocyanide Safety Issue; HNF-SA-3126-FP; DE&S Hanford, Inc., Pacific Northwest National Laboratory, Babad Technical Services: Richland, WA, USA, 1997; pp. 1–10. [Google Scholar]
- What Are EPA’s Drinking Water Regulations for Cyanide? Available online: https://safewater.zendesk.com/hc/en-us/articles/212077077-4-What-are-EPA-s-drinking-water-regulations-for-cyanide- (accessed on 10 August 2018).
- Dzombak, D.; Ghosh, R.; Young, T. Physical-Chemical Properties and Reactivity of Cyanide in Water and Soil. In Cyanide in Water and Soil: Chemistry, Risk, and Management; Dzombak, D., Ghosh, R., Young, T., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 57–92. ISBN 9781566706667. [Google Scholar]
- Bringmann, G.; Kuehn, R. Comparative results of the damaging effects of water pollutants against bacteria (Pseudomonas putida) and blue algae (Microcystis aeruginosa). GWF Wasser/Abwasser 1976, 117, 410–413. (In Germany) [Google Scholar]
- Kapoor, V.; Elk, M.; Li, X.; Santo Domingo, J.W. Inhibitory effect of cyanide on wastewater nitrification determined using SOUR and RNA-based gene-specific assays. Lett. Appl. Microbiol. 2016, 63, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Cho, H.U.; Lee, D.S.; Park, D.; Park, J.M. Comparative study of free cyanide inhibition on nitrification and denitrification in batch and continuous flow systems. Desalin. 2011, 279, 439–444. [Google Scholar] [CrossRef]
- Kim, Y.M.; Cho, H.U.; Lee, D.S.; Park, D.; Park, J.M. Influence of operational parameters on nitrogen removal efficiency and microbial communities in a full-scale activated sludge process. Water Res. 2011, 45, 5785–5795. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Lee, D.S.; Park, C.; Park, D.; Park, J.M. Effects of free cyanide on microbial communities and biological carbon and nitrogen removal performance in the industrial activated sludge process. Water Res. 2010, 45, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Konopka, A.; Plymale, A.E.; Carvajal, D.A.; Lin, X.; McKinley, J.P. Environmental controls on the activity of aquifer microbial communities in the 300 area of the Hanford Site. Microb. Ecol. 2013, 66, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Gensemer, R.W.; DeForest, D.K.; Stenhouse, A.J.; Higgins, C.J.; Cardwell, R.D. Aquatic toxicity of cyanide. In Cyanide in Water and Soil: Chemisty, Risk, and Management; Dzombak, D., Ghosh, R., Young, T., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006; pp. 251–284. ISBN 9781566706667. [Google Scholar]
- Bååth, E.; Pettersson, M.; Söderberg, K.H. Adaptation of a rapid and economical microcentrifugation method to measure thymidine and leucine incorporation by soil bacteria. Soil Biol. Biochem. 2001, 33, 1571–1574. [Google Scholar] [CrossRef]
- Demoling, F.; Figueroa, D.; Bååth, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar] [CrossRef]
- Velasco Ayuso, S.; López-Archilla, A.; Montes, C.; Guerrero, M.C. Microbial activities in a coastal, sandy aquifer system (Doñana Natural Protected Area, SW Spain). Geomicrobiol. J. 2010, 27, 409–423. [Google Scholar] [CrossRef]
- Wilhartitz, I.C.; Kirschner, A.K.T.; Stadler, H.; Herndl, G.J.; Dietzel, M.; Latal, C.; Mach, R.L.; Farnleitner, A.H. Heterotrophic prokaryotic production in ultra-oligotrophic alpine karst aquifers and ecological implications. FEMS Microbiol. Ecol. 2009, 68, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D. Measuring bacterial biomass production and growth rates from leucine incorporation in natural aquatic environments. Methods Microbiol. 2001, 30, 227–237. [Google Scholar]
- Kepner, R.L.; Pratt, J.R. Use of fluorochromes for direct enumeration of total bacteria in environmental samples: past and present. Microbiol. Rev. 1994, 58, 603–615. [Google Scholar] [PubMed]
- The Geochemist's Workbench; 12.0.1; Aqueous Solutions LLC: Champaign, IL, USA, 2015.
- Sehmel, G.A. Cyanide and Antimony Thermodynamic Database for the Aqueous Species and Solids for the EPA-Minteq Geochemical Code; PNNL-6835; Pacific Northwest National Laboratory: Richland, WA, USA, 1989; pp. 1–224. [Google Scholar]
- The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 10 August 2018).
- Ribeiro, P.J., Jr.; Diggle, P.J. geoR: A package for geostatistical analysis. R News 2001, 1, 14–18. [Google Scholar]
- Furrer, R.; Nychka, D.; Sain, S. Fields: Tools for Spatial Data; 6.11. R Foundation, 2009. Available online: http://citeseerx.ist.psu.edu/viewdoc/summary?doi=10.1.1.304.1555 (accessed on 10 August 2018).
- Dunning, J.C.; Ma, Y.; Marquis, R.E. Anaerobic killing of oral streptococci by reduced, transition metal cations. Appl. Environ. Microbiol. 1998, 64, 27–33. [Google Scholar] [PubMed]
- Graham, E.B.; Crump, A.R.; Resch, C.T.; Fansler, S.; Arntzen, E.; Kennedy, D.W.; Fredrickson, J.K.; Stegen, J.C. Deterministic influences exceed dispersal effects on hydrologically-connected microbiomes. Environ. Microbiol. 2017, 19, 1552–1567. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Fredrickson, J.K.; Plymale, A.E.; Dohnalkova, A.C.; Resch, C.T.; McKinley, J.P.; Shi, L. An Autotrophic H2-oxidizing, Nitrate-Respiring, Tc(VII)-Reducing Acidovorax sp. Isolated from a Subsurface Oxic-Anoxic Transition Zone. Environ. Microbiol. Rep. 2015, 7, 395–403. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Well # | Depth to Ground-Water when Sampled (ft bgs) 1 | Date of Well Construction (year) 1 | Elevation (m) 1 | Depth (ft) 1 | Screened Interval(s) (ft bgs) 1 |
|---|---|---|---|---|---|
| #1 (E33-268) | - | 2012 | 198.0 | 263.5 | 241.9–252.4 |
| #2 (E33-360) | - | 2014 | 199.7 | 272.8 | 251.8–271.7 |
| #3 (E28-27) | 284.86 | 1987 | 208.5 | 301.5 | 269.8–289.8, |
| - | - | - | - | - | 291.2–301.5 |
| #4 (E32-9) | 247.52 | 1991 | 197.1 | 254.6 | - |
| #5 (E32-3) | 281.01 | 1987 | 207.3 | 304.0 | 266.2–286.2, |
| - | - | - | - | - | 291–301 |
| #6 (E32-7) | 262.54 | 1991 | 201.7 | 273.8 | 245.6–266.3 |
| #7 (E32-8) | 249.80 | 1990 | 197.8 | 256.7 | 234.7–255.0 |
| #8 (E33-34) | 237.71 | 1990 | 194.1 | 240.0 | 219.0–239.3 |
| Well # | % 3H-Leu Uptake 1 | Cells/mL 2 | Most Recent Eh (mV) 3 | Most Recent Total CN− (µg/L) 3,4 | Historical Maximum Total CN− (µg/L) 3 |
|---|---|---|---|---|---|
| #1 (E33-268) | 8.42 | 6.6 × 10+5 | +16 | 297 | 492 |
| #2 (E33-360) | 0.0110 | 5.8 × 10+5 | +304 | 190 | 190 |
| #3 (E28-27) | 2.92 | 6.6 × 10+5 | +8 | 4 | 12 |
| #4 (E32-9) | 2.33 | 1.1 × 10+6 | +382 | 22 | 91 |
| #5 (E32-3) | 3.60 | 2.3 × 10+5 | +202 | 4 | 9 |
| #6 (E32-7) | 1.95 | 2.5 × 10+5 | +233 | 4 | 20 |
| #7 (E32-8) | 1.10 | 3.1 × 10+5 | +169 | 40 | 40 |
| #8 (E33-34) | 0.0256 | 1.4 × 10+5 | +369 | 100 | 558 |
| Aqueous Species | Concentration (µM) | CN−% |
|---|---|---|
| 1 Fe(CN)64− | 1.296 | 67.07 |
| 1 NaFe(CN)63− | 0.464 | 24.04 |
| 2 HCN(aq) | 0.723 | 6.24 |
| 1 KFe(CN)63− | 0.038 | 1.95 |
| 2 CN− | 0.030 | 0.26 |
| 1 Na2Fe(CN)62− | 0.005 | 0.25 |
| 3 Ni(CN)42− | 0.004 | 0.13 |
| 3 Cu(CN)2− | 0.005 | 0.08 |
| Aqueous Species | Concentration (µM) | CN−% |
|---|---|---|
| 1 Fe(CN)64− | 0.64 | 52.76 |
| 2 HCN(aq) | 1.56 | 21.44 |
| 1 NaFe(CN)63− | 0.24 | 19.88 |
| 1 Fe(CN)63− | 0.04 | 3.14 |
| 1 KFe(CN)63− | 0.02 | 1.44 |
| 2 CN− | 0.05 | 0.64 |
| 3 Ni(CN)42− | 0.01 | 0.39 |
| 1 Na2Fe(CN)62− | 0.00 | 0.22 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plymale, A.; Wells, J.; Graham, E.; Qafoku, O.; Brooks, S.; Lee, B. Bacterial Productivity in a Ferrocyanide-Contaminated Aquifer at a Nuclear Waste Site. Water 2018, 10, 1072. https://doi.org/10.3390/w10081072
Plymale A, Wells J, Graham E, Qafoku O, Brooks S, Lee B. Bacterial Productivity in a Ferrocyanide-Contaminated Aquifer at a Nuclear Waste Site. Water. 2018; 10(8):1072. https://doi.org/10.3390/w10081072
Chicago/Turabian StylePlymale, Andrew, Jacqueline Wells, Emily Graham, Odeta Qafoku, Shelby Brooks, and Brady Lee. 2018. "Bacterial Productivity in a Ferrocyanide-Contaminated Aquifer at a Nuclear Waste Site" Water 10, no. 8: 1072. https://doi.org/10.3390/w10081072
APA StylePlymale, A., Wells, J., Graham, E., Qafoku, O., Brooks, S., & Lee, B. (2018). Bacterial Productivity in a Ferrocyanide-Contaminated Aquifer at a Nuclear Waste Site. Water, 10(8), 1072. https://doi.org/10.3390/w10081072