Knowledge to Predict Pathogens: Legionella pneumophila Lifecycle Critical Review Part I Uptake into Host Cells

Abstract
:1. Introduction
1.1. Rationale: Human Health Impacts and Level of Concern
1.2. Microbiology of Legionella spp.
1.3. Epidemiology of Legionellosis
1.4. Objectives of this Review Article
2. Materials and Methods
Literature Review Method
3. Results
3.1. Legionella pneumophila Life Cycle
3.2. Effects of Engineered System Stressors on L. pneumophila
3.3. Engineered System Stressors within a Biofilm
3.4. The Effects of Genetic Knockdowns
3.5. Postulated Modeling Framework
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Glick, T.H.; Gregg, M.B.; Berman, B.; Mallison, G.; Rhodes, W.W.; Kassanoff, I. Pontiac fever an epidemic of unknown etiology in a health department: I. Clinical and epidemiologic aspects. Am. J. Epidemiol. 1978, 107, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, A.F.; McDade, J.E.; Patton, C.M.; Bennett, J.V.; Skaliy, P.; Feeley, J.C.; Anderson, D.C.; Potter, M.E.; Newhouse, V.F.; Gregg, M.B.; et al. Pontiac Fever: Isolation of the Etiologic Agent (Legionella pneumophila) and Demonstration of its Mode of Transmission. Am. J. Epidemiol. 1981, 114, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.W.; Tsai, T.R.; Orenstein, W.; Parkin, W.E.; Beecham, H.J.; Sharrar, R.G.; Harris, J.; Mallison, G.F.; Martin, S.M.; McDade, J.E.; et al. Legionnaires’ disease. N. Engl. J. Med. 1977, 297, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Winn, W.C. Legionnaires disease: Historical perspective. Clin. Microbiol. Rev. 1988, 1, 60–81. [Google Scholar] [CrossRef] [PubMed]
- McDade, J.E.; Shepard, C.C.; Fraser, D.W.; Tsai, T.R.; Redus, M.A.; Dowdle, W.R. Legionnaires’ disease. N. Engl. J. Med. 1977, 297, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Boshuizen, H.C.; Neppelenbroek, S.E.; van Vliet, H.; Schellekens, J.F.; den Boer, J.W.; Peeters, M.F.; Conyn-van Spaendonck, M.A. Subclinical Legionella infection in workers near the source of a large outbreak of Legionnaires disease. J. Infect. Dis. 2001, 184, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Kirby, B.D.; Snyder, K.M.; Meyer, R.D.; Finegold, S.M. Legionnaires’ disease: Clinical features of 24 cases. Ann. Intern. Med. 1978, 89, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Lattimer, G.L.; Rhodes, L.V.; Salventi, J.S.; Galgon, J.P.; Stonebraker, V.; Boley, S.; Haas, G. The Philadelphia epidemic of Legionnaire’s disease: Clinical, pulmonary and serologic findings two years later. Ann. Intern. Med. 1979, 90, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, P.H.; Meyer, R.D. Legionnaires’ disease. Chest 1984, 85, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Marston, B.J.; Lipman, H.B.; Breiman, R.F. Surveillance for Legionnaires’ disease: Risk factors for morbidity and mortality. Arch. Intern. Med. 1994, 154, 2417–2422. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, K.; Klein, T.W.; Friedman, H.; Yamamoto, Y. Involvement of nicotinic acetylcholine receptors in suppression of antimicrobial activity and cytokine responses of alveolar macrophages to Legionella pneumophila infection by nicotine. J. Immunol. 2001, 167, 6518–6524. [Google Scholar] [CrossRef] [PubMed]
- Straus, W.L.; Plouffe, J.F.; File, T.M.; Lipman, H.B.; Hackman, B.H.; Salstrom, S.J.; Benson, R.F.; Breiman, R.F. Risk factors for domestic acquisition of legionnaires disease. Ohio Legionnaires’ Disease Group. Arch. Intern. Med. 1996, 156, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Office of the Surgeon General (US); Office on Smoking and Health (US). The Health Consequences of Smoking: A Report of the Surgeon General; Reports of the Surgeon General; U.S. Public Health Service: Atlanta, GA, USA, 2004.
- Neil, K.; Berkelman, R. Increasing incidence of legionellosis in the United States, 1990–2005: Changing epidemiologic trends. Clin. Infect. Dis. 2008, 47, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Legionellosis—United States, 2000–2009. MMWR Morb. Mortal. Wkly. Rep. 2011, 60, 1083–1086. [Google Scholar]
- Choffnes, E.R.; Mack, A. Global Issues in Water, Sanitation, and Health: Workshop Summary; National Academies Press: Washington, DC, USA, 2009. [Google Scholar]
- Yoder, J.; Roberts, V.; Craun, G.F.; Hill, V.; Hicks, L.; Alexander, N.T.; Radke, V.; Calderon, R.L.; Hlavsa, M.C.; Beach, M.J.; et al. Surveillance for waterborne disease and outbreaks associated with drinking water and water not intended for drinking–United States, 2005–2006. Morb. Mortal. Wkly. Rep. Surveill. Summ. 2008, 57, 39–62. [Google Scholar]
- National Research Council (US) Committee on Drinking Water Contaminants. Review of Methods for Assessing Microbial Pathogens; National Academies Press (US): Washington, DC, USA, 1999. [Google Scholar]
- US EPA. Contaminant Candidate List 3-CCL 3. Available online: https://www.epa.gov/ccl/contaminant-candidate-list-3-ccl-3 (accessed on 13 January 2017).
- Craun, G.F.; Brunkard, J.M.; Yoder, J.S.; Roberts, V.A.; Carpenter, J.; Wade, T.; Calderon, R.L.; Roberts, J.M.; Beach, M.J.; Roy, S.L. Causes of Outbreaks Associated with Drinking Water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010, 23, 507–528. [Google Scholar] [CrossRef] [PubMed]
- Benin, A.L.; Benson, R.F.; Arnold, K.E.; Fiore, A.E.; Cook, P.G.; Williams, L.K.; Fields, B.; Besser, R.E. An outbreak of travel-associated Legionnaires disease and Pontiac fever: The need for enhanced surveillance of travel-associated legionellosis in the United States. J. Infect. Dis. 2002, 185, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.B. Future health assessment and risk-management integration for infectious diseases and biological weapons for deployed U.S. Forces. In Strategies to Protect the Health of Deployed U.S. Forces: Assessing Health Risks to Deployed U.S. Forces; The National Academies Press: Washington, DC, USA, 2000; pp. 59–112. [Google Scholar]
- Muder, R.R.; Victor, L.Y.; Woo, A.H. Mode of transmission of Legionella pneumophila: A critical review. Arch. Intern. Med. 1986, 146, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Osterholm, M.T.; Chin, T.D.; Osborne, D.O.; Dull, H.B.; Dean, A.G.; Fraser, D.W.; Hayes, P.S.; Hall, W.N. A 1957 Outbreak of Legionnaires’ disease associated with a meat packing plant. Am. J. Epidemiol. 1983, 117, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, F.J.; Hoge, C.W.; Farley, T.A.; Barbaree, J.M.; Breiman, R.F.; Benson, R.F.; McFarland, L.M. Communitywide outbreak of Legionnaires’ disease associated with a grocery store mist machine. J. Infect. Dis. 1992, 165, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Den Boer, J.; Yzerman, E.P.; Schellekens, J.; Lettinga, K.D.; Boshuizen, H.C.; Steenbergen, J.; Bosman, A.; Hof, S.; Vliet, H.; Peeters, M.F.; et al. A large outbreak of Legionnaires’ disease at a flower show, the Netherlands, 1999. Emerg. Infect. Dis. 2002, 8, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Pastoris, M.C.; Ciceroni, L.; Monaco, R.L.; Goldoni, P.; Mentore, B.; Flego, G.; Cattani, L.; Ciarrocchi, S.; Pinto, A.; Visca, P. Molecular epidemiology of an outbreak of Legionnaires’ disease associated with a cooling tower in Genova-Sestri Ponente, Italy. Eur. J. Clin. Microbiol. Infect. Dis. 1997, 16, 883–892. [Google Scholar] [CrossRef]
- Brown, C.M.; Nuorti, P.J.; Breiman, R.F.; Hathcock, A.L.; Fields, B.S.; Lipman, H.B.; Llewellyn, G.C.; Hofmann, J.; Cetron, M. A community outbreak of Legionnaires’ disease linked to hospital cooling towers: An epidemiological method to calculate dose of exposure. Int. J. Epidemiol. 1999, 28, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Bhopal, R.S.; Fallon, R.J.; Buist, E.C.; Black, R.J.; Urquhart, J.D. Proximity of the home to a cooling tower and risk of non-outbreak Legionnaires’ disease. Br. Med. J. 1991, 302, 378–383. [Google Scholar] [CrossRef]
- Schwarzenbach, R.P.; Egli, T.; Hofstetter, T.B.; von Gunten, U.; Wehrli, B. Global water pollution and human health. Annu. Rev. Environ. Resour. 2010, 35, 109–136. [Google Scholar] [CrossRef]
- Dondero, T.J.; Rendtorff, R.C.; Mallison, G.F.; Weeks, R.M.; Levy, J.S.; Wong, E.W.; Schaffner, W. An outbreak of Legionnaires’ disease associated with a contaminated air-conditioning cooling tower. N. Engl. J. Med. 1980, 302, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Berglund, B.; Brunekreef, B.; Knöppe, H.; Lindvall, T.; Maroni, M.; Mølhave, L.; Skov, P. Effects of indoor air pollution on human health. Indoor Air 1992, 2, 2–25. [Google Scholar] [CrossRef]
- Armstrong, T.W.; Haas, C.N. Legionnaires’ disease: Evaluation of a quantitative microbial risk assessment model. J. Water Health 2008, 6, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Jernigan, D.; Hofmann, J.; Cetron, M.; Nuorti, J.; Fields, B.; Benson, R.; Breiman, R.; Lipman, H.; Carter, R.; Genese, C.; et al. Outbreak of Legionnaires’ disease among cruise ship passengers exposed to a contaminated whirlpool spa. Lancet 1996, 347, 494–499. [Google Scholar] [CrossRef]
- Lück, P.C.; Leupold, I.; Hlawitschka, M.; Helbig, J.; Carmienke, I.; Jatzwauk, L.; Guderitz, T. Prevalence of Legionella species, serogroups, and monoclonal subgroups in hot water systems in south-eastern Germany. Zentralbl. Hyg. Umweltmed. Int. J. Hyg. Environ. Med. 1993, 193, 450–460. [Google Scholar]
- Lück, P.; Lau, B.; Seidel, S.; Postl, U. Legionellae in dental units–a hygienic risk? Dtsch. Zahn Mund Kieferheilkd. Zentralbl. 1991, 80, 341–346. [Google Scholar]
- Steele, T.W.; Lanser, J.; Sangster, N. Isolation of Legionella longbeachae serogroup 1 from potting mixes. Appl. Environ. Microbiol. 1990, 56, 49–53. [Google Scholar] [PubMed]
- Van Heijnsbergen, E.; de Roda Husman, A.M.; Lodder, W.J.; Bouwknegt, M.; Docters van Leeuwen, A.E.; Bruin, J.P.; Euser, S.M.; den Boer, J.W.; Schalk, J.A.C. Viable Legionella pneumophila bacteria in natural soil and rainwater puddles. J. Appl. Microbiol. 2014, 117, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Van Heijnsbergen, E.; van Deursen, A.; Bouwknegt, M.; Bruin, J.P.; de Roda Husman, A.M.; Schalk, J.A.C. Presence and persistence of viable, clinically relevant Legionella pneumophila bacteria in garden soil in the Netherlands. Appl. Environ. Microbiol. 2016, 82, 5125–5131. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Dowsett, A.; Dennis, P.; Lee, J.; Keevil, C. Influence of temperature and plumbing material selection on biofilm formation and growth of Legionella pneumophila in a model potable water system containing complex microbial flora. Appl. Environ. Microbiol. 1994, 60, 1585–1592. [Google Scholar] [PubMed]
- Dennis, R. Legionnaires’ disease–preventative maintenance. J. Inst. Hosp. Eng. 1988, 42, 14–15. [Google Scholar] [PubMed]
- Dennis, P. Environmental Factors Affecting the Survival and Pathogenicity of Legionella pneumophila. Ph.D. Thesis, Open University, Hong Kong, China, 1986. [Google Scholar]
- Gast, R.J.; Moran, D.M.; Dennett, M.R.; Wurtsbaugh, W.A.; Amaral-Zettler, L.A. Amoebae and Legionella pneumophila in saline environments. J. Water Health 2011, 9, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Barbaree, J.M.; Sanchez, A.; Sanden, G.N. Tolerance of Legionella species to sodium chloride. Curr. Microbiol. 1983, 9, 1–5. [Google Scholar] [CrossRef]
- Ohno, A.; Kato, N.; Yamada, K.; Yamaguchi, K. Factors influencing survival of Legionella pneumophila serotype 1 in hot spring water and tap water. Appl. Environ. Microbiol. 2003, 69, 2540–2547. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Nour, M.; Duncan, C.; Low, D.E.; Guyard, C. Biofilms: The stronghold of Legionella pneumophila. Int. J. Mol. Sci. 2013, 14, 21660–21675. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Viscogliosi, P.; Solignac, L.; Delattre, J.-M. Viability PCR, a culture-independent method for rapid and selective quantification of viable Legionella pneumophila cells in environmental water samples. Appl. Environ. Microbiol. 2009, 75, 3502–3512. [Google Scholar] [CrossRef] [PubMed]
- Harb, O.S.; Gao, L.Y.; Abu Kwaik, Y. From protozoa to mammalian cells: A new paradigm in the life cycle of intracellular bacterial pathogens. Environ. Microbiol. 2000, 2, 251–265. [Google Scholar] [CrossRef] [PubMed]
- De Felipe, K.S.; Pampou, S.; Jovanovic, O.S.; Pericone, C.D.; Ye, S.F.; Kalachikov, S.; Shuman, H.A. Evidence for acquisition of Legionella type IV secretion substrates via interdomain horizontal gene transfer. J. Bacteriol. 2005, 187, 7716–7726. [Google Scholar] [CrossRef] [PubMed]
- Bodet, C.; Sahr, T.; Dupuy, M.; Buchrieser, C.; Héchard, Y. Legionella pneumophila transcriptional response to chlorine treatment. Water Res. 2012, 46, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired legionellosis: An international collaborative survey. J. Infect. Dis. 2002, 186, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Yu, V.L.; Vickers, R.M. Disease due to the Legionellaceae (other than Legionella pneumophila): Historical, microbiological, clinical, and epidemiological review. Medicine 1989, 68, 116–132. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, B.A.; Carman, J.; Eckert, K.; Tucker, G.; Givney, R.; Cameron, S. Does using potting mix make you sick? Results from a Legionella longbeachae case-control study in South Australia. Epidemiol. Infect. 2007, 135, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Cazalet, C.; Gomez-Valero, L.; Rusniok, C.; Lomma, M.; Dervins-Ravault, D.; Newton, H.J.; Sansom, F.M.; Jarraud, S.; Zidane, N.; Ma, L.; et al. Analysis of the Legionella longbeachae genome and transcriptome uncovers unique strategies to cause Legionnaires’ disease. PLoS Genet. 2010, 6, e1000851. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Legionnaires’ disease associated with potting soil—California, Oregon, and Washington, May–June 2000. MMWR Morb. Mortal. Wkly. Rep. 2000, 49, 777–778. [Google Scholar]
- Graham, F.; White, P.; Harte, D.; Kingham, S. Changing epidemiological trends of legionellosis in New Zealand, 1979–2009. Epidemiol. Infect. 2012, 140, 1481–1496. [Google Scholar] [CrossRef] [PubMed]
- Fliermans, C.B.; Cherry, W.B.; Orrison, L.H.; Smith, S.J.; Tison, D.L.; Pope, D.H. Ecological distribution of Legionella pneumophila. Appl. Environ. Microbiol. 1981, 41, 9–16. [Google Scholar] [PubMed]
- Blackburn, B.G.; Craun, G.F.; Yoder, J.S.; Hill, V.; Calderon, R.L.; Chen, N.; Lee, S.H.; Levy, D.A.; Beach, M.J. Surveillance for waterborne-disease outbreaks associated with drinking water—United States, 2001–2002. Morb. Mortal. Wkly. Rep. Surveill. Summ. 2004, 53, 23–45. [Google Scholar]
- National Research Council (US) Committee. Ecology and evolution of waterborne pathogens and indicator organisms. In Indicators for Waterborne Pathogens; National Academies Press (US): Washington, DC, USA, 2004. [Google Scholar]
- World Health Organization. Guide to Ship Sanitation, 3rd ed.; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Flemming, H.-C. Biofilms in drinking water systems-part I: Overview. Gas Wasserfach Wasser Abwasser 1998, 139, S65–S72. [Google Scholar]
- Jjemba, P.K.; Johnson, W.; Bukhari, Z.; LeChevallier, M.W. Occurrence and control of Legionella in recycled water systems. Pathogens 2015, 4, 470–502. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.Y.; Ashbolt, N.J. The role of biofilms and protozoa in Legionella pathogenesis: Implications for drinking water. J. Appl. Microbiol. 2009, 107, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Carrington, C.B. Pathology of Legionnaires’ disease. Ann. Intern. Med. 1979, 90, 496–499. [Google Scholar] [CrossRef] [PubMed]
- Balows, A.; Fraser, D.; Swartz, M.N.; Carrington, C.B.; Eickhoff, T.C.; Isenberg, H.D.; Ward, P.A.; Tsai, T.F.; Finn, D.R.; Pilkaytis, B.D.; et al. International symposium on Legionnaires’ disease, 13–15 November 1978, Atlanta, Georgia. Ann. Intern. Med. 1979, 90, 489–714. [Google Scholar]
- Swartz, M.N. Clinical aspects of Legionnaires’ disease. Ann. Intern. Med. 1979, 90, 492–495. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, H.D. Microbiology of Legionnaires’ disease bacterium. Ann. Intern. Med. 1979, 90, 499–502. [Google Scholar] [CrossRef]
- Eickhoff, T.C. Epidemiology of Legionnaires’ disease. Ann. Intern. Med. 1979, 90, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Hubber, A.; Roy, C.R. Modulation of host cell function by Legionella pneumophila type IV effectors. Annu. Rev. Cell Dev. Biol. 2010, 26, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Isberg, R.R.; O’Connor, T.; Heidtman, M. The Legionella pneumophila replication vacuole: Making a cozy niche inside host cells. Nat. Rev. Microbiol. 2009, 7, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Anderson, J.E.; Mueller, S.A.; Gaines, W.A.; Kendall, A.M. Literature review—Efficacy of various disinfectants against Legionella in water systems. Water Res. 2002, 36, 4433–4444. [Google Scholar] [CrossRef]
- Taylor, M.; Ross, K.; Bentham, R. Legionella, protozoa, and biofilms: Interactions within complex microbial systems. Microb. Ecol. 2009, 58, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Swanson, M.; Hammer, B. Legionella pneumophila pathogenesis: A fateful journey from amoebae to macrophages. Annu. Rev. Microbiol. 2000, 54, 567–613. [Google Scholar] [CrossRef] [PubMed]
- Albert-Weissenberger, C.; Cazalet, C.; Buchrieser, C. Legionella pneumophila—A human pathogen that co-evolved with fresh water protozoa. Cell. Mol. Life Sci. CMLS 2007, 64, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Declerck, P.; Behets, J.; De Keersmaecker, B.; Ollevier, F. Receptor-mediated uptake of Legionella pneumophila by Acanthamoeba castellanii and Naegleria lovaniensis. J. Appl. Microbiol. 2007, 103, 2697–2703. [Google Scholar] [CrossRef] [PubMed]
- Harb, O.S.; Venkataraman, C.; Haack, B.J.; Gao, L.Y.; Kwaik, Y.A. Heterogeneity in the attachment and uptake mechanisms of the Legionnaires’ disease bacterium, Legionella pneumophila, by protozoan hosts. Appl. Environ. Microbiol. 1998, 64, 126–132. [Google Scholar] [PubMed]
- Declerck, P.; Behets, J.; Delaedt, Y.; Margineanu, A.; Lammertyn, E.; Ollevier, F. Impact of non-Legionella bacteria on the uptake and intracellular replication of Legionella pneumophila in Acanthamoeba castellanii and Naegleria lovaniensis. Microb. Ecol. 2005, 50, 536–549. [Google Scholar] [CrossRef] [PubMed]
- Amer, A.O. Modulation of caspases and their non-apoptotic functions by Legionella pneumophila. Cell. Microbiol. 2010, 12, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Stone, B.J.; Abu Kwaik, Y. Expression of multiple pili by Legionella pneumophila: Identification and characterization of a type IV pilin gene and its role in adherence to mammalian and protozoan cells. Infect. Immun. 1998, 66, 1768–1775. [Google Scholar] [PubMed]
- Bozue, J.A.; Johnson, W. Interaction of Legionella pneumophila with Acanthamoeba castellanii: Uptake by coiling phagocytosis and inhibition of phagosome-lysosome fusion. Infect. Immun. 1996, 64, 668–673. [Google Scholar] [PubMed]
- Horwitz, M.A. The Legionnaires’ disease bacterium (Legionella pneumophila) inhibits phagosome-lysosome fusion in human monocytes. J. Exp. Med. 1983, 158, 2108–2126. [Google Scholar] [CrossRef] [PubMed]
- Rittig, M.G.; Krause, A.; Häupl, T.; Schaible, U.E.; Modolell, M.; Kramer, M.D.; Lütjen-Drecoll, E.; Simon, M.M.; Burmester, G.R. Coiling phagocytosis is the preferential phagocytic mechanism for Borrelia burgdorferi. Infect. Immun. 1992, 60, 4205–4212. [Google Scholar] [PubMed]
- Kwaik, Y.A.; Gao, L.-Y.; Stone, B.J.; Venkataraman, C.; Harb, O.S. Invasion of protozoa by Legionella pneumophila and its role in bacterial ecology and pathogenesis. Appl. Environ. Microbiol. 1998, 64, 3127–3133. [Google Scholar] [PubMed]
- Clemens, D.L.; Horwitz, M.A. Membrane sorting during phagocytosis: Selective exclusion of major histocompatibility complex molecules but not complement receptor CR3 during conventional and coiling phagocytosis. J. Exp. Med. 1992, 175, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Steinert, M.; Ott, M.; Lück, P.C.; Tannich, E.; Hacker, J. Studies on the uptake and intracellular replication of Legionella pneumophila in protozoa and in macrophage-like cells. FEMS Microbiol. Ecol. 1994, 15, 299–307. [Google Scholar] [CrossRef]
- Mintz, C.S.; Arnold, P.I.; Johnson, W.; Schultz, D.R. Antibody-independent binding of complement component C1q by Legionella pneumophila. Infect. Immun. 1995, 63, 4939–4943. [Google Scholar] [PubMed]
- Mintz, C.S.; Schultz, D.R.; Arnold, P.I.; Johnson, W. Legionella pneumophila lipopolysaccharide activates the classical complement pathway. Infect. Immun. 1992, 60, 2769–2776. [Google Scholar] [PubMed]
- Bellinger-Kawahara, C.; Horwitz, M. Legionella pneumophila fixes complement component C3 to its surface-demonstration by ELISA. In Program of the 1987 Annual Meeting of the American Society of Microbiology, Atlanta, GA, 1–6 March; American Society for Microbiology: Washington, DC, USA, 1987; pp. 1–6. [Google Scholar]
- Cianciotto, N.P.; Fields, B.S. Legionella pneumophila mip gene potentiates intracellular infection of protozoa and human macrophages. Proc. Natl. Acad. Sci. USA 1992, 89, 5188–5191. [Google Scholar] [CrossRef] [PubMed]
- Fields, B.S. The molecular ecology of Legionella. Trends Microbiol. 1996, 4, 286–290. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Eisenstein, B.I.; Mody, C.H.; Engleberg, N.C. A mutation in the mip gene results in an attenuation of Legionella pneumophila virulence. J. Infect. Dis. 1990, 162, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Cianciotto, N.P.; Eisenstein, B.I.; Mody, C.H.; Toews, G.B.; Engleberg, N.C. A Legionella pneumophila gene encoding a species-specific surface protein potentiates initiation of intracellular infection. Infect. Immun. 1989, 57, 1255–1262. [Google Scholar] [PubMed]
- Ratcliff, R.M.; Lanser, J.A.; Manning, P.A.; Heuzenroeder, M.W. Sequence-based classification scheme for the genus Legionella targeting the mip Gene. J. Clin. Microbiol. 1998, 36, 1560–1567. [Google Scholar] [PubMed]
- Fischer, G.; Bang, H.; Ludwig, B.; Mann, K.; Hacker, J. Mip protein of Legionella pneumophila exhibits peptidyl-prolyl-cis/trans isomerase (PPlase) activity. Mol. Microbiol. 1992, 6, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Lundemose, A.G.; Kay, J.E.; Pearce, J.H. Chlamydia trachomatis Mip-like protein has peptidyl-prolyl cis/trans isomerase activity that is inhibited by FK506 and rapamycin and is implicated in initiation of chlamydial infection. Mol. Microbiol. 1993, 7, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Moro, A.; Ruiz-Cabello, F.; Fernández-Cano, A.; Stock, R.P.; González, A. Secretion by Trypanosoma cruzi of a peptidyl-prolyl cis-trans isomerase involved in cell infection. EMBO J. 1995, 14, 2483–2490. [Google Scholar] [PubMed]
- Wintermeyer, E.; Ludwig, B.; Steinert, M.; Schmidt, B.; Fischer, G.; Hacker, J. Influence of site specifically altered Mip proteins on intracellular survival of Legionella pneumophila in eukaryotic cells. Infect. Immun. 1995, 63, 4576–4583. [Google Scholar] [PubMed]
- Helbig, J.H.; Kurtz, J.B.; Pastoris, M.C.; Pelaz, C.; Lück, P.C. Antigenic lipopolysaccharide components of Legionella pneumophila recognized by monoclonal antibodies: Possibilities and limitations for division of the species into serogroups. J. Clin. Microbiol. 1997, 35, 2841–2845. [Google Scholar] [PubMed]
- Köhler, R.; Fanghänel, J.; König, B.; Lüneberg, E.; Frosch, M.; Rahfeld, J.-U.; Hilgenfeld, R.; Fischer, G.; Hacker, J.; Steinert, M. Biochemical and functional analyses of the Mip protein: Influence of the N-terminal half and of peptidylprolyl isomerase activity on the virulence of Legionella pneumophila. Infect. Immun. 2003, 71, 4389–4397. [Google Scholar] [CrossRef] [PubMed]
- Wieland, H.; Faigle, M.; Lang, F.; Northoff, H.; Neumeister, B. Regulation of the Legionella mip-promotor during infection of human monocytes. FEMS Microbiol. Lett. 2002, 212, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Garduño, R.A.; Quinn, F.D.; Hoffman, P.S. HeLa cells as a model to study the invasiveness and biology of Legionella pneumophila. Can. J. Microbiol. 1998, 44, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Moffat, J.F.; Tompkins, L. A quantitative model of intracellular growth of Legionella pneumophila in Acanthamoeba castellanii. Infect. Immun. 1992, 60, 296–301. [Google Scholar] [PubMed]
- Köhler, R.; Bubert, A.; Goebel, W.; Steinert, M.; Hacker, J.; Bubert, B. Expression and use of the green fluorescent protein as a reporter system in Legionella pneumophila. Mol. Gen. Genet. MGG 2000, 262, 1060–1069. [Google Scholar] [CrossRef] [PubMed]
- Abu Kwaik, Y.; Venkataraman, C.; Harb, O.S.; Gao, L.-Y. Signal transduction in the protozoan host Hartmannella vermiformis upon attachment and invasion by Legionella micdadei. Appl. Environ. Microbiol. 1998, 64, 3134–3139. [Google Scholar] [PubMed]
- Lucas, C.E.; Brown, E.; Fields, B.S. Type IV pili and type II secretion play a limited role in Legionella pneumophila biofilm colonization and retention. Microbiol. Read. Engl. 2006, 152, 3569–3573. [Google Scholar] [CrossRef] [PubMed]
- Mampel, J.; Spirig, T.; Weber, S.S.; Haagensen, J.A.J.; Molin, S.; Hilbi, H. Planktonic replication is essential for biofilm formation by Legionella pneumophila in a complex medium under static and dynamic flow conditions. Appl. Environ. Microbiol. 2006, 72, 2885–2895. [Google Scholar] [CrossRef] [PubMed]
- Berk, S.G.; Ting, R.S.; Turner, G.W.; Ashburn, R.J. Production of respirable vesicles containing live Legionella pneumophila cells by two Acanthamoeba spp. Appl. Environ. Microbiol. 1998, 64, 279–286. [Google Scholar] [PubMed]
- Cao, Z.; Jefferson, D.M.; Panjwani, N. Role of Carbohydrate-mediated adherence in cytopathogenic mechanisms of Acanthamoeba. J. Biol. Chem. 1998, 273, 15838–15845. [Google Scholar] [CrossRef] [PubMed]
- Hales, L.M.; Shuman, H.A. The Legionella pneumophila rpoS gene is required for growth within Acanthamoeba castellanii. J. Bacteriol. 1999, 181, 4879–4889. [Google Scholar] [PubMed]
- Gal-Mor, O.; Segal, G. The Legionella pneumophila GacA homolog (LetA) is involved in the regulation of icm virulence genes and is required for intracellular multiplication in Acanthamoeba castellanii. Microb. Pathog. 2003, 34, 187–194. [Google Scholar] [CrossRef]
- Escoll, P.; Rolando, M.; Gomez-Valero, L.; Buchrieser, C. From Amoeba to Macrophages: Exploring the Molecular Mechanisms of Legionella pneumophila Infection in Both Hosts. Curr. Top. Microbiol. Immunol. 2013, 376, 1–34. [Google Scholar] [PubMed]
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S. Growth of Legionella pneumophila in Acanthamoeba castellanii enhances invasion. Infect. Immun. 1994, 62, 3254–3261. [Google Scholar] [PubMed]
- Gomez-Valero, L.; Rusniok, C.; Buchrieser, C. Legionella pneumophila: Population genetics, phylogeny and genomics. Infect. Genet. Evol. 2009, 9, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Legionella: From environmental habitats to disease pathology, detection and control. Environ. Microbiol. 1999, 1, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Berjeaud, J.-M.; Chevalier, S.; Schlusselhuber, M.; Portier, E.; Loiseau, C.; Aucher, W.; Lesouhaitier, O.; Verdon, J. Legionella pneumophila: The paradox of a highly sensitive opportunistic waterborne pathogen able to persist in the environment. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Declerck, P. Biofilms: The environmental playground of Legionella pneumophila. Environ. Microbiol. 2010, 12, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Emtiazi, F.; Schwartz, T.; Marten, S.M.; Krolla-Sidenstein, P.; Obst, U. Investigation of natural biofilms formed during the production of drinking water from surface water embankment filtration. Water Res. 2004, 38, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Yu-sen, E.L.; Vidic, R.D.; Stout, J.E.; Victor, L.Y. Legionella in water distribution systems. Am. Water Works Assoc. J. 1998, 90, 112–121. [Google Scholar]
- Haas, C.N. Disinfection. In Water Quality and Treatment, A Handbook of Community Water Supplies; Pontius, F., Ed.; McGraw Hill: New York, NY, USA, 1990. [Google Scholar]
- Miyamoto, M.; Yamaguchi, Y.; Sasatu, M. Disinfectant effects of hot water, ultraviolet light, silver ions and chlorine on strains of Legionella and nontuberculous mycobacteria. Microbios 1999, 101, 7–13. [Google Scholar]
- Kilvington, S.; Price, J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. J. Appl. Bacteriol. 1990, 68, 519–525. [Google Scholar] [CrossRef] [PubMed]
- European Working Group for Legionella Infections (EWGLI). ESGLI/EWGLI Technical Guidelines for the Investigation, Control and Prevention of Travel Associated Legionnaires’ Disease; The European Commission: Brussels, Belgium, 2011. [Google Scholar]
- Cooper, I.R.; Hanlon, G.W. Resistance of Legionella pneumophila serotype 1 biofilms to chlorine-based disinfection. J. Hosp. Infect. 2010, 74, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Gião, M.S.; Wilks, S.; Azevedo, N.F.; Vieira, M.J.; Keevil, C.W. Incorporation of natural uncultivable Legionella pneumophila into potable water biofilms provides a protective niche against chlorination stress. Biofouling 2009, 25, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Murga, R.; Forster, T.S.; Brown, E.; Pruckler, J.M.; Fields, B.S.; Donlan, R.M. Role of biofilms in the survival of Legionella pneumophila in a model potable-water system. Microbiology 2001, 147, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Allegra, S.; Berger, F.; Berthelot, P.; Grattard, F.; Pozzetto, B.; Riffard, S. Use of flow cytometry to monitor Legionella viability. Appl. Environ. Microbiol. 2008, 74, 7813–7816. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-W.; Hwang, Y.-H.; Cheng, W.-Y.; Chang, C.-P. Effects of chlorination and heat disinfection on long-term starved Legionella pneumophila in warm water. J. Appl. Microbiol. 2007, 102, 1636–1644. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, T.; Ghosh, A.; Pazhani, G.P.; Shinoda, S. Current perspectives on viable but non-culturable (VBNC) pathogenic bacteria. Front. Public Health 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Cook, K.L.; Bolster, C.H. Survival of Campylobacter jejuni and Escherichia coli in groundwater during prolonged starvation at low temperatures. J. Appl. Microbiol. 2007, 103, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Um, H.Y.; Kong, H.G.; Lee, H.J.; Choi, H.K.; Park, E.J.; Kim, S.T.; Murugiyan, S.; Chung, E.; Kang, K.Y.; Lee, S.-W. Altered gene expression and intracellular changes of the viable but nonculturable state in Ralstonia solanacearum by copper treatment. Plant Pathol. J. 2013, 29, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Alleron, L.; Khemiri, A.; Koubar, M.; Lacombe, C.; Coquet, L.; Cosette, P.; Jouenne, T.; Frere, J. VBNC Legionella pneumophila cells are still able to produce virulence proteins. Water Res. 2013, 47, 6606–6617. [Google Scholar] [CrossRef] [PubMed]
- Epalle, T.; Girardot, F.; Allegra, S.; Maurice-Blanc, C.; Garraud, O.; Riffard, S. Viable but not culturable forms of Legionella pneumophila generated after heat shock treatment are infectious for macrophage-like and alveolar epithelial cells after resuscitation on Acanthamoeba polyphaga. Microb. Ecol. 2015, 69, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Steinert, M.; Emödy, L.; Amann, R.; Hacker, J. Resuscitation of viable but nonculturable Legionella pneumophila Philadelphia JR32 by Acanthamoeba castellanii. Appl. Environ. Microbiol. 1997, 63, 2047–2053. [Google Scholar] [PubMed]
- Casini, B.; Valentini, P.; Baggiani, A.; Torracca, F.; Frateschi, S.; Nelli, L.C.; Privitera, G. Molecular epidemiology of Legionella pneumophila serogroup 1 isolates following long-term chlorine dioxide treatment in a university hospital water system. J. Hosp. Infect. 2008, 69, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Långmark, J.; Storey, M.V.; Ashbolt, N.J.; Stenström, T.-A. Accumulation and fate of microorganisms and microspheres in biofilms formed in a pilot-scale water distribution system. Appl. Environ. Microbiol. 2005, 71, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Codony, F.; Morató, J.; Mas, J. Role of discontinuous chlorination on microbial production by drinking water biofilms. Water Res. 2005, 39, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- De Beer, D.; Srinivasan, R.; Stewart, P.S. Direct measurement of chlorine penetration into biofilms during disinfection. Appl. Environ. Microbiol. 1994, 60, 4339–4344. [Google Scholar] [PubMed]
- White, G.C. Handbook of chlorination for potable water, wastewater, cooling water, industrial processes, and swimming pools. In Handbook of Chlorination for Potable Water, Wastewater, Cooling Water, Industrial Processes, and Swimming Pools; Van Nostrand Reinhold: New York, NY, USA, 1972. [Google Scholar]
- Kool, J.L.; Carpenter, J.C.; Fields, B.S. Effect of monochloramine disinfection of municipal drinking water on risk of nosocomial Legionnaires’ disease. Lancet Lond. Engl. 1999, 353, 272–277. [Google Scholar] [CrossRef]
- Chen, X.; Stewart, P.S. Chlorine penetration into artificial biofilm is limited by a reaction-diffusion interaction. Environ. Sci. Technol. 1996, 30, 2078–2083. [Google Scholar] [CrossRef]
- LeChevallier, M.W.; Cawthon, C.D.; Lee, R.G. Inactivation of biofilm bacteria. Appl. Environ. Microbiol. 1988, 54, 2492–2499. [Google Scholar] [PubMed]
- Shen, Y.; Huang, C.; Lin, J.; Wu, W.; Ashbolt, N.J.; Liu, W.-T.; Nguyen, T.H. Effect of disinfectant exposure on Legionella pneumophila associated with simulated drinking water biofilms: Release, inactivation, and infectivity. Environ. Sci. Technol. 2017, 51, 2087–2095. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.M.; Braun-Howland, E.B. Growth of Escherichia coli in model distribution system biofilms exposed to hypochlorous Acid or monochloramine. Appl. Environ. Microbiol. 2003, 69, 5463–5471. [Google Scholar] [CrossRef] [PubMed]
- Chiao, T.-H.; Clancy, T.M.; Pinto, A.; Xi, C.; Raskin, L. Differential resistance of drinking water bacterial populations to monochloramine disinfection. Environ. Sci. Technol. 2014, 48, 4038–4047. [Google Scholar] [CrossRef] [PubMed]
- Backer, H.; Hollowell, J. Use of iodine for water disinfection: Iodine toxicity and maximum recommended dose. Environ. Health Perspect. 2000, 108, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Cargill, K.L.; Pyle, B.H.; Sauer, R.L.; McFeters, G.A. Effects of culture conditions and biofilm formation on the iodine susceptibility of Legionella pneumophila. Can. J. Microbiol. 1992, 38, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.M.; Eccles, J.; Fricker, C. Laboratory observations of biocide efficiency against Legionella in model cooling tower systems. ASHRAE Trans. 1999, 105, 607–623. [Google Scholar]
- Farooq, S.; Chian, E.; Engelbrecht, R. Basic concepts in disinfection with ozone. J. Water Pollut. Control Fed. 1977, 49, 1818–1831. [Google Scholar] [PubMed]
- Hamelin, C. Production of single-and double-strand breaks in plasmid DNA by ozone. Int. J. Radiat. Oncol. Biol. Phys. 1985, 11, 253–257. [Google Scholar] [CrossRef]
- Domingue, E.L.; Tyndall, R.; Mayberry, W.; Pancorbo, O. Effects of three oxidizing biocides on Legionella pneumophila serogroup 1. Appl. Environ. Microbiol. 1988, 54, 741–747. [Google Scholar] [PubMed]
- McGrane, W. Ozone, a study of the effects of biocides on Legionella pneumophila. Ind. Water Treat. 1995, 27, 28–32. [Google Scholar]
- Bartram, J. Legionella and the Prevention of Legionellosis; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Proctor, C.R.; Hammes, F. Drinking water microbiology—From measurement to management. Curr. Opin. Biotechnol. 2015, 33, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Murray, J. A study of the prevention of hot tapwater burns. Burns 1988, 14, 185–193. [Google Scholar] [CrossRef]
- Boccia, S.; Laurenti, P.; Borella, P.; Moscato, U.; Capalbo, G.; Cambieri, A.; Amore, R.; Quaranta, G.; Boninti, F.; Orsini, M.; et al. Prospective 3-year surveillance for nosocomial and environmental Legionella pneumophila: Implications for infection control. Infect. Control Hosp. Epidemiol. 2006, 27, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Dyck, A.; Exner, M.; Kramer, A. Experimental based experiences with the introduction of a water safety plan for a multi-located university clinic and its efficacy according to WHO recommendations. BMC Public Health 2007, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- King, C.H.; Shotts, E.B.; Wooley, R.E.; Porter, K.G. Survival of coliforms and bacterial pathogens within protozoa during chlorination. Appl. Environ. Microbiol. 1988, 54, 3023–3033. [Google Scholar] [PubMed]
- Storey, M.; Ashbolt, N.; Stenström, T. Biofilms, thermophilic amoebae and Legionella pneumophila—A quantitative risk assessment for distributed water. Water Sci. Technol. 2004, 50, 77–82. [Google Scholar] [PubMed]
- Szewzyk, U.; Szewzyk, R.; Manz, W.; Schleifer, K.-H. Microbiological safety of drinking water. Annu. Rev. Microbiol. 2000, 54, 81–127. [Google Scholar] [CrossRef] [PubMed]
- Wimpenny, J.; Manz, W.; Szewzyk, U. Heterogeneity in biofilms. FEMS Microbiol. Rev. 2000, 24, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.; Bouchez, T.; Nicolas, V.; Robert, S.; Loret, J.F.; Lévi, Y. Amoebae in domestic water systems: Resistance to disinfection treatments and implication in Legionella persistence. J. Appl. Microbiol. 2004, 97, 950–963. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Bova, G.; Ross, T.; Mackie, K.; Paquette, N.; Merz, W.; Perl, T.M. A 17-month evaluation of a chlorine dioxide water treatment system to control Legionella species in a hospital water supply. Infect. Control Hosp. Epidemiol. 2003, 24, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Cianciotto, N.P. Iron acquisition by Legionella pneumophila. BioMetals 2007, 20, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Orsi, N. The antimicrobial activity of lactoferrin: Current status and perspectives. BioMetals 2004, 17, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, G.A.; Rand, J.L.; O’Leary, K.C.; Rygel, A.C.; Chauret, C.; Andrews, R.C. Disinfectant efficacy of chlorite and chlorine dioxide in drinking water biofilms. Water Res. 2005, 39, 1809–1817. [Google Scholar] [CrossRef] [PubMed]
- Zusman, T.; Gal-Mor, O.; Segal, G. Characterization of a Legionella pneumophila relA Insertion Mutant and Roles of RelA and RpoS in Virulence Gene Expression. J. Bacteriol. 2002, 184, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Tiaden, A.; Spirig, T.; Sahr, T.; Wälti, M.A.; Boucke, K.; Buchrieser, C.; Hilbi, H. The autoinducer synthase LqsA and putative sensor kinase LqsS regulate phagocyte interactions, extracellular filaments and a genomic island of Legionella pneumophila. Environ. Microbiol. 2010, 12, 1243–1259. [Google Scholar] [CrossRef] [PubMed]
- Lynch, D.; Fieser, N.; Glöggler, K.; Forsbach-Birk, V.; Marre, R. The response regulator LetA regulates the stationary-phase stress response in Legionella pneumophila and is required for efficient infection of Acanthamoeba castellanii. FEMS Microbiol. Lett. 2003, 219, 241–248. [Google Scholar] [CrossRef]
- Wagner, C.; Khan, A.S.; Kamphausen, T.; Schmausser, B.; Ünal, C.; Lorenz, U.; Fischer, G.; Hacker, J.; Steinert, M. Collagen binding protein Mip enables Legionella pneumophila to transmigrate through a barrier of NCI-H292 lung epithelial cells and extracellular matrix. Cell. Microbiol. 2007, 9, 450–462. [Google Scholar] [CrossRef] [PubMed]
- Barbaree, J.M.; Fields, B.S.; Feeley, J.C.; Gorman, G.W.; Martin, W.T. Isolation of protozoa from water associated with a legionellosis outbreak and demonstration of intracellular multiplication of Legionella pneumophila. Appl. Environ. Microbiol. 1986, 51, 422–424. [Google Scholar] [PubMed]
- Ashbolt, N.J. Microbial contamination of drinking water and disease outcomes in developing regions. Toxicology 2004, 198, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Shevchuk, O.; Jäger, J.; Steinert, M. Virulence properties of the Legionella pneumophila cell envelope. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Ashbolt, N.J.; Schoen, M.E.; Soller, J.A.; Roser, D.J. Predicting pathogen risks to aid beach management: The real value of quantitative microbial risk assessment (QMRA). Water Res. 2010, 44, 4692–4703. [Google Scholar] [CrossRef] [PubMed]
- Weir, M.H.; Razzolini, M.T.P.; Rose, J.B.; Masago, Y. Water reclamation redesign for reducing Cryptosporidium risks at a recreational spray park using stochastic models. Water Res. 2011, 45, 6505–6514. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Alvarez, M.S.; Weir, M.H.; Pope, J.M.; Seghezzo, L.; Rajal, V.B.; Salusso, M.M.; Moraña, L.B. Development of a relative risk model for drinking water regulation and design recommendations for a peri urban region of Argentina. Int. J. Hyg. Environ. Health 2015, 218, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Petterson, S. Application of a QMRA framework to inform selection of drinking water interventions in the developing context. Risk Anal. 2016, 36, 203–214. [Google Scholar] [CrossRef] [PubMed]
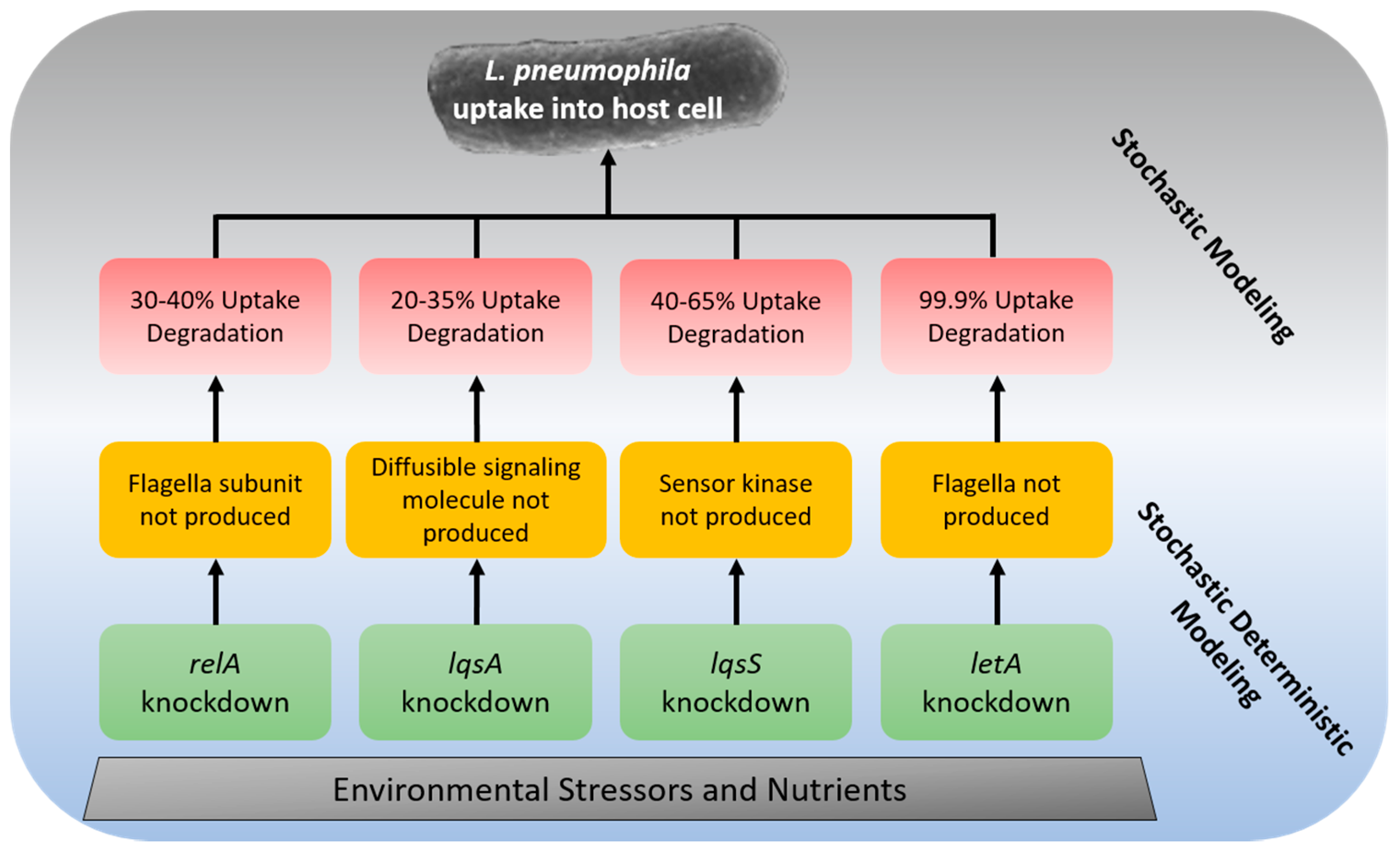

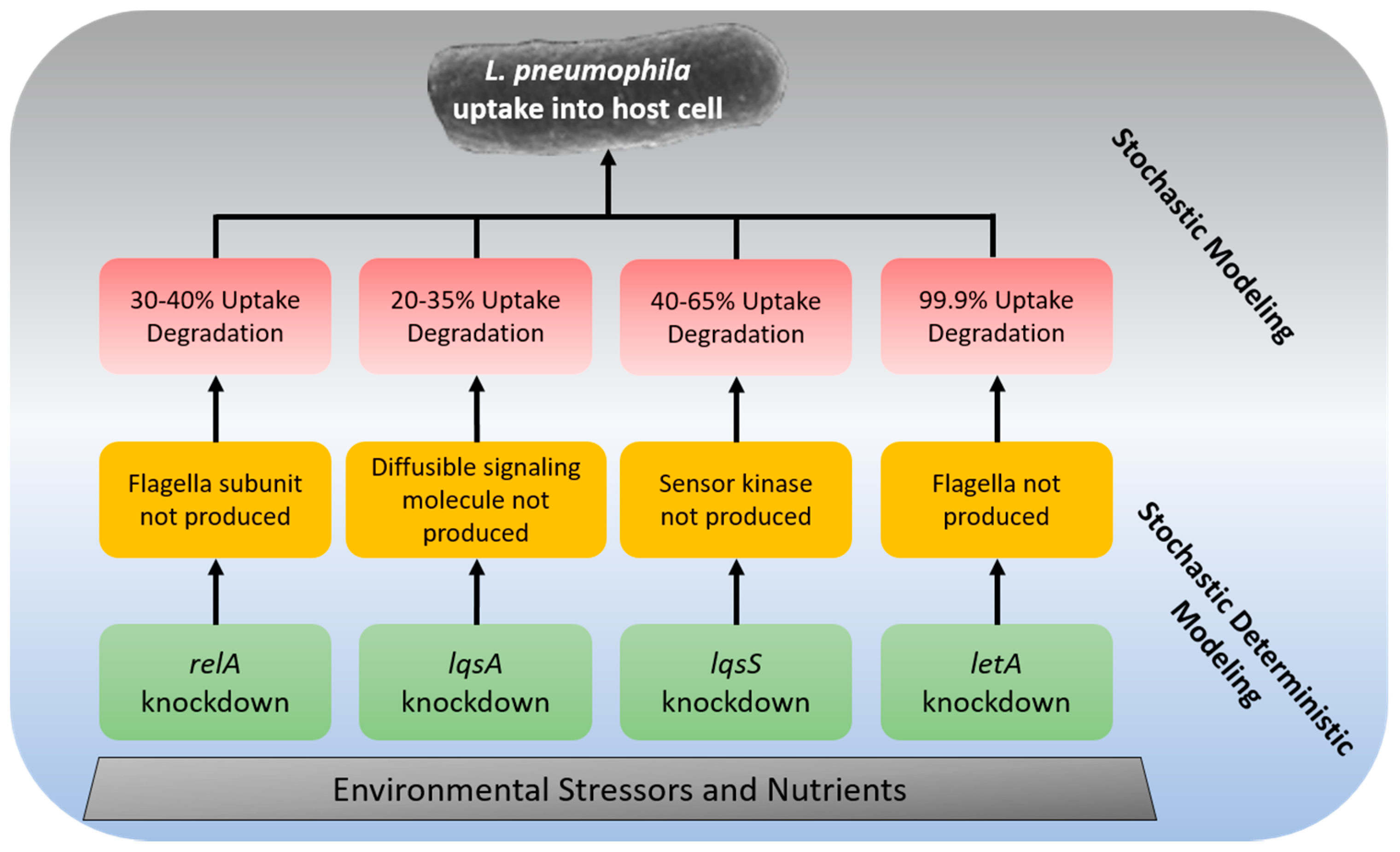{kind=link}
| Gene | Host Cell | Change in Uptake Efficiency * | Degradation of Uptake Rate ** | Process |
|---|---|---|---|---|
| relA | A. castellanii | 30–40% | 30–40% *** | The flagella subunit is not produced, causing reduced anchoring to the host cell [167] |
| pilE | U937 cells | 33.9% | 66.1% | Only the short pili is expressed in mutant [80] |
| pilE | HeLa cells | 34.0% | 66.0% | Only the short pili is expressed in mutant [80] |
| pilE | WI-26 VA4 | 48.7% | 51.3% | Only the short pili is expressed in mutant [80] |
| pilE | A. polyphaga | 53.4% | 46.4% | Only the short pili is expressed in mutant [80] |
| lqsA | A. castellanii | 20–35% | 20–35% | Diffusible signaling molecule (LA-1) is not produced during mutation [168] |
| lqsS | A. castellanii | 40–65% | 40–65% | Sensor kinase which recognizes LA-1 is not produced during mutation [168] |
| letA | A. castellanii | Log 3 CFU/L | 99.9% | The flagella is not produced in mutant [169] |
| Mip | NCI-H292 lung epithelial cells | Log 2 CFU/L | 99% | Mutant loses the inability to cross the extracellular matrix [170] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mraz, A.L.; Weir, M.H. Knowledge to Predict Pathogens: Legionella pneumophila Lifecycle Critical Review Part I Uptake into Host Cells. Water 2018, 10, 132. https://doi.org/10.3390/w10020132
Mraz AL, Weir MH. Knowledge to Predict Pathogens: Legionella pneumophila Lifecycle Critical Review Part I Uptake into Host Cells. Water. 2018; 10(2):132. https://doi.org/10.3390/w10020132
Chicago/Turabian StyleMraz, Alexis L., and Mark H. Weir. 2018. "Knowledge to Predict Pathogens: Legionella pneumophila Lifecycle Critical Review Part I Uptake into Host Cells" Water 10, no. 2: 132. https://doi.org/10.3390/w10020132
APA StyleMraz, A. L., & Weir, M. H. (2018). Knowledge to Predict Pathogens: Legionella pneumophila Lifecycle Critical Review Part I Uptake into Host Cells. Water, 10(2), 132. https://doi.org/10.3390/w10020132