Canopy Light Utilization and Yield of Rice under Rain-Catching and Controlled Irrigation




Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Plant Material and Cultivation Practices
2.3. Water Regimes
2.4. Measurements
- (1)
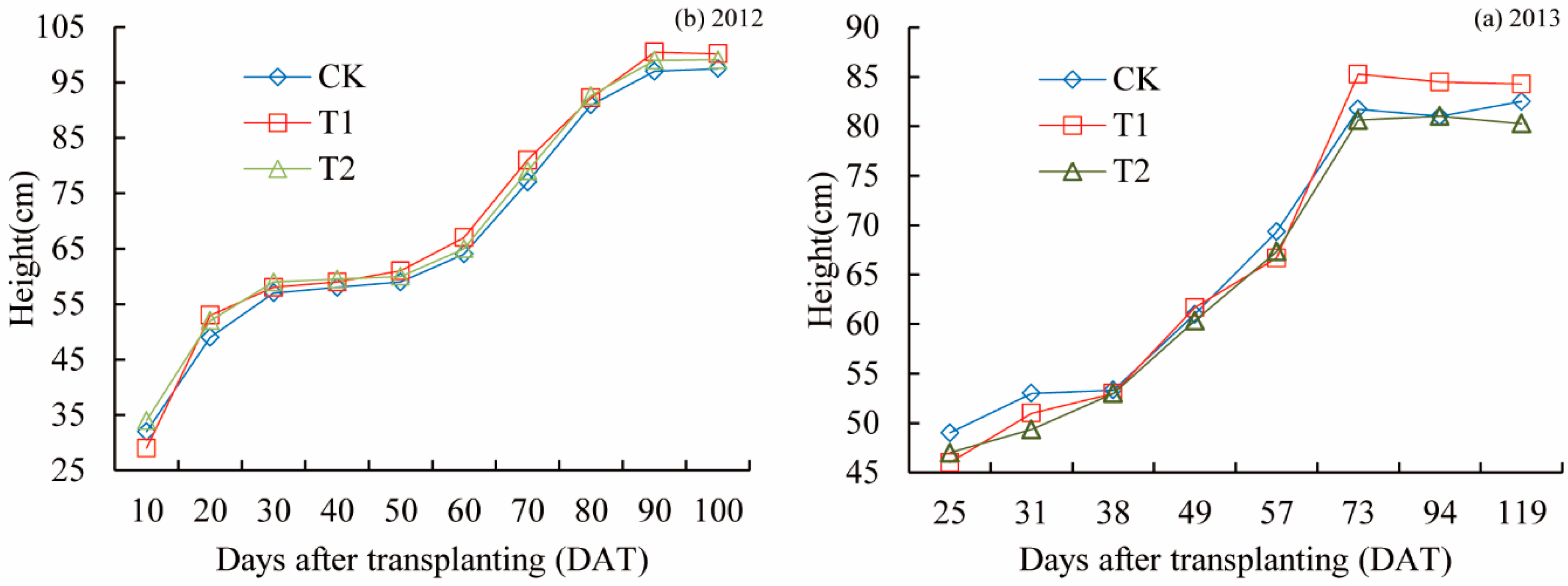
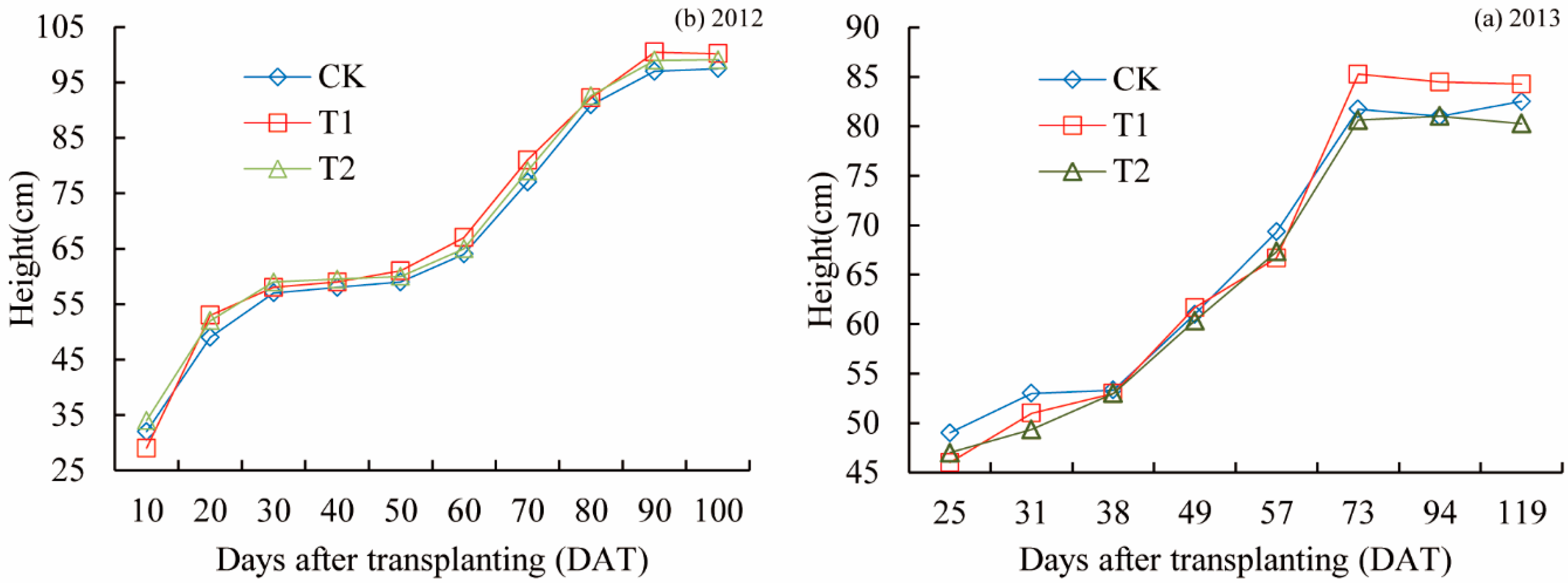
- The height of rice was measured using a ruler, and the tiller number per plot was calculated every 6–10 days in 2012 and 2013. The high tiller rate was also measured.
- (2)
- We used SUNSCAN (Delta-T Devices) to measure the PAR and LAI at 11:30~12:30 as the weather was clear and cloudless. The PAR interception rate was calculated based on the following equation:where FIPARh is the PAR interception rate of the canopy height (h); PARh is the PAR of the canopy height (μmol/(m2∙s)); and PART is the PAR of the top (μmol/(m2∙s)).FIPARh = 1 − PARh/PART
- (3)
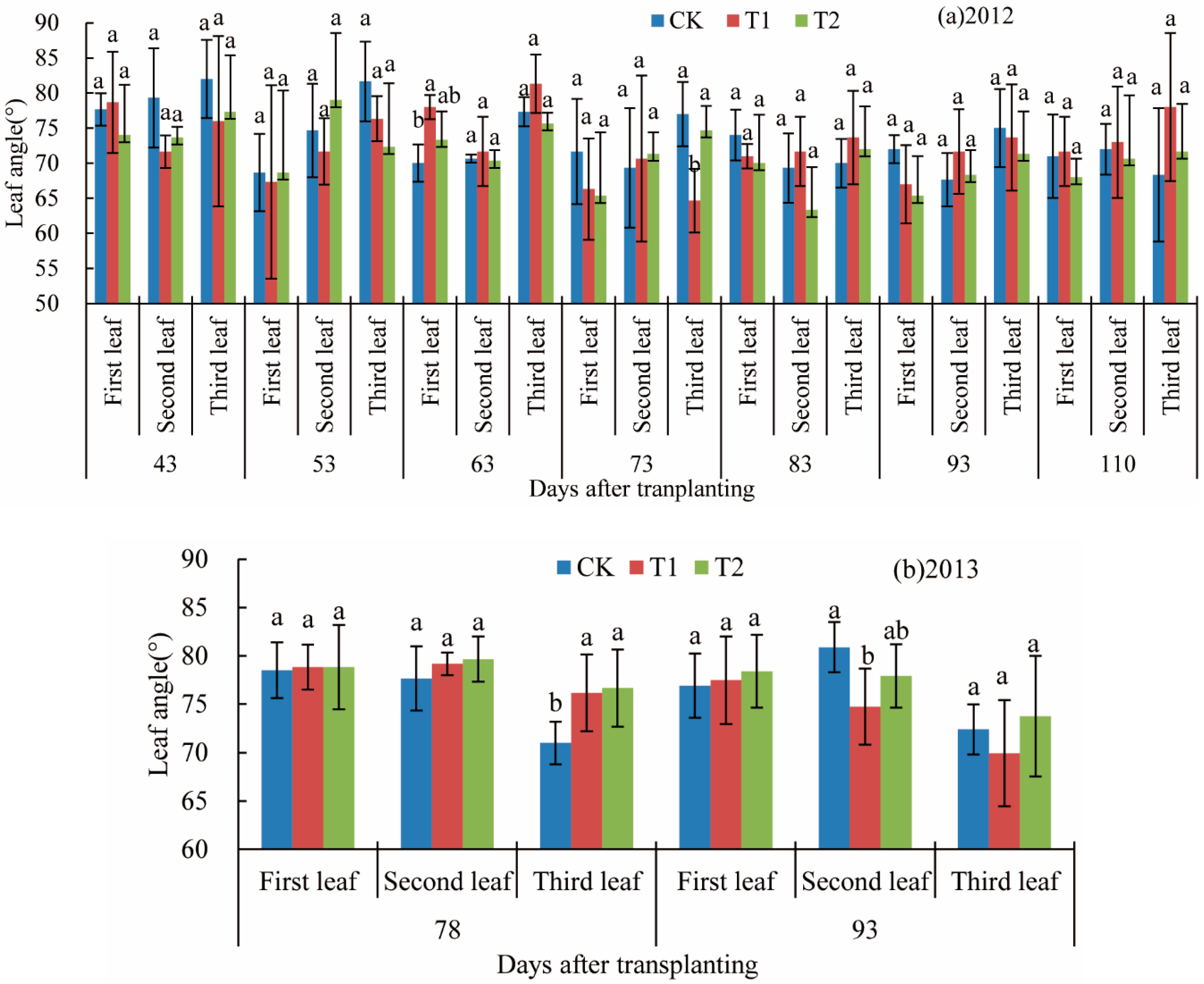
- Leaf angle. We dug a soil block (5 cm × 5 cm × 5 cm) around the root of each plant at about 6:00, and then placed in an indoor water storage barrel for 1 h to maintain the natural state of the rice plant. Each tiller was photographed in a natural state. We adjusted the leaves to both sides of the background plate according to the direction of the leaves of each stem, and occasionally, when the non-half leaf sequence occurred, the rotation of the plant was adjusted to the appropriate position and a picture was added. After photographing, the leaf tilt angle was measured by Image-Pro Plus 5 software (Media Cybernetics, Inc., Rockville, MD, USA). For a straight blade, the angle between the base and the stem was measured, and the angle of the leaf was calculated. For a curved blade, the blade was divided into 2–5 segments according to the blade bending degree, and the average leaf angle was calculated from the leaf area as the weight, allowing the average leaf inclination to be obtained.
- (4)
- At the end of the growing season in 2013, the grain yield was measured in each plot, the spike length was measured by a ruler, and the effective spikes were calculated. Spikes were collected and first dried at 105 °C for 1 h, and then dried at 80 °C to a constant weight for 48 h to determine the yield component, including spike weight, filled grain, unfilled grain, setting rate, and 1000-grain weight.
2.5. Statistical Analysis
3. Results
3.1. Plant Height and Tiller
3.2. Leaf Angle
3.3. Canopy Interception
3.4. Yield Characters
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Enríquez, S.; Pantoja-Reyes, N.I. Form-function analysis of the effect of canopy morphology on leaf self-shading in the seagrass Thalassia testudinum. Oecologia 2005, 145, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.M.; Romero, J. Effects of in situ experimental shading on the Mediterranean seagrass Posidonia oceanica. Mar. Ecol. Prog. Ser. 2001, 215, 107–120. [Google Scholar] [CrossRef]
- Gordon, D.M.; Grey, K.A.; Chase, S.C.; Simpson, C.J. Changes to the structure and productivity of a Posidonia sinuosa, meadow during and after imposed shading. Aquat. Bot. 1994, 47, 265–275. [Google Scholar] [CrossRef]
- Lee, K.S.; Dunton, K.H. Effect of in situ light reduction on the maintenance, growth and partitioning of carbon resources in Thalassia testudinum Banks ex Konig. J. Exp. Mar. Biol. Ecol. 1997, 210, 53–73. [Google Scholar] [CrossRef]
- Campbell, S.J.; Miller, C.J. Shoot and abundance characteristics of the seagrass Heterozostera tasmanica in Westernport estuary (South-Eastern Australia). Aquat. Bot. 2002, 3, 33–46. [Google Scholar] [CrossRef]
- Casanova, D.; Epema, G.F.; Goudriaan, J. Monitoring rice reflectance at field level for estimating biomass and LAI. Field Crops Res. 1998, 55, 83–92. [Google Scholar] [CrossRef]
- Xie, L.; Xu, Z.; Liu, H. Distribution of Photosynthetic Active Radiation in Different Panicle Type Rice Communities during Later Growth Stage. J. Jilin Agric. Univ. 2004, 26, 476–479. [Google Scholar]
- Zhang, X.; Lu, C.; Hu, N.; Yao, K.; Zhang, Q.; Dai, Q. Simulation of Leaf Inclination Angle Distribution for Rice with Different Plant Types. Chin. J. Rice Sci. 2012, 26, 205–210. [Google Scholar]
- Lemeur, R.; Blad, B.L. A critical review of light models for estimating the short-wave radiation regime of plant canopies. Agric. For. Meteorol. 1974, 14, 255–286. [Google Scholar] [CrossRef]
- Myneni, R.B.; Ross, J.; Asrar, G. A review on the theory of photon transport in leaf canopies. Agric. For. Meteorol. 1989, 45, 1–153. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, L.; Liu, X.; Liu, L.; Cao, W.; Yan, Z. Modeling the leaf angle dynamics in rice plant. PLoS ONE 2017, 12, e0171890. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Chen, Y.; Zhang, H.; LI, Z.; Zhou, Q.; Yu, C.; Kong, X.; Liu, L.; Wang, Z.; Yang, J. Canopy light and nitrogen distributions are related to grain yield and nitrogen use efficiency in rice. Field Crops Res. 2017, 206, 74–85. [Google Scholar] [CrossRef]
- Sakamoto, T.; Morinaka, Y.; Ohnishi, T.; Sunohara, H.; Fujioka, S.; Ueguchi-Tanaka, M.; Mizutani, M.; Sakata, K.; Takatsuto, S.; Yoshida, S.; et al. Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice. Nat. Biotechnol. 2006, 24, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Van Camp, W. Yield enhancement genes: Seeds for growth. Curr. Opin. Biotechnol. 2005, 6, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Sharma-Natu, P.; Ghildiyal, M.C. Potential targets for improving photosynthesis and crop yield. Curr. Sci. 2005, 88, 1918–1928. [Google Scholar]
- Katsura, K.; Maeda, S.; Lubis, I.; Horie, T.; Cao, W.X.; Shiraiwa, T. The high yield of irrigated rice in Yunnan, China: A cross-location analysis. Field Crops Res. 2008, 107, 1–11. [Google Scholar] [CrossRef]
- Hua, S.; Cao, B.; Zheng, B.; Lia, B.; Sun, C. Quantitative evaluation of influence of Prostrate Growth 1 gene on rice canopy structure based on three-dimensional structure model. Field Crops Res. 2016, 194, 65–74. [Google Scholar] [CrossRef]
- Kumagai, E.; Hamaoka, N.; Araki, T.; Ueno, O. Dorsoventral asymmetry of photosynthesis and photoinhibition in flag leaves of two rice cultivars that differ in nitrogen response and leaf angle. Physiol. Plant. 2014, 151, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Goto, A.; Sasahara, H.; Shigemune, A.; Miura, K. Hokuriku 193: A new high-yielding indica rice cultivar bred in Japan. Jpn. Agric. Res. Q. 2012, 43, 13–18. [Google Scholar] [CrossRef]
- Jang, S.; An, G.; Li, H.Y. Rice leaf angle and grain size are affected by the OsBUL1 transcriptional activator complex. Plant Physiol. 2017, 173, 688–702. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Liu, F. Study on canopy characteristics of rice under different water supplies. Water Sav. Irrig. 2004, 6, 1–3. [Google Scholar]
- Hao, S.; Guo, X.; Zhang, Z. Compensation Effects of Water Stress and Rewatering on the Structure of Rice Canopy. Trans. Chin. Soc. Agric. Mach. 2010, 41, 52–55. [Google Scholar]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zhang, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christiea, P.; Zhu, Z.L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.P.; Cui, Z.L.; Fan, M.S.; Vitousek, P.; Zhao, M.; Ma, W.Q.; Wang, Z.L.; Zhang, W.J.; Yan, X.Y.; Yang, J.C.; et al. Producing more grain with lower environmental costs. Nature 2014, 514, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.Y.F.; Tan, C.S.; Drury, C.F.; Gaynor, J.D. Controlled drainage and subirrigation influences tile nitrate loss and corn yields in a sandy loam soil in Southwestern Ontario. Agric. Ecosyst. Environ. 2002, 90, 81–88. [Google Scholar] [CrossRef]
- Woli, K.P.; David, M.B.; Cooke, R.A.; McIsaac, G.F.; Mitchell, C.A. Nitrogen balance in and export from agricultural fields associated with controlled drainage systems and denitrifying bioreactors. Ecol. Eng. 2010, 36, 1558–1566. [Google Scholar] [CrossRef]
- Skaggs, R.W.; Fausey, N.R.; Evans, R.O. Drainage water management. J. Soil Water Conserv. 2012, 67, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Bouman, B.A.M. A conceptual framework for the improvement of crop water productivity at different spatial scales. Agric. Syst. 2007, 93, 43–60. [Google Scholar] [CrossRef]
- Ye, Y.; Liang, X.Q.; Chen, Y.X.; Liu, J.; Gu, J.T.; Guo, R.; Liang, L. Alternate wetting and drying irrigation and controlled-release nitrogen fertilizer in late-season rice. Effects on dry matter accumulation, yield, water and nitrogen use. Field Crops Res. 2013, 144, 212–224. [Google Scholar] [CrossRef]
- Lampayan, R.M.; Samoy-Pascual, K.C.; Sibayan, E.B.; Ella, V.B.; Jayag, O.P.; Cabangon, R.J.; Bouman, B.A.M. Effects of alternate wetting and drying (AWD) threshold level and plant seedling age on crop performance, water input, and water productivity of transplanted rice in Central Luzon, Philippines. Paddy Water Environ. 2015, 13, 215–227. [Google Scholar] [CrossRef]
- Tan, X.; Shao, D.; Liu, H.; Yang, F.S.; Xiao, C.; Yang, H.D. Effects of alternate wetting and drying irrigation on percolation and nitrogen leaching in paddy fields. Paddy Water Environ. 2013, 11, 381–395. [Google Scholar] [CrossRef]
- Peng, S.; Luo, Y.; Xu, J.; Shahbaz, K.; Jiao, X.; Wang, W. Integrated Irrigation and Drainage Practices to Enhance Water Productivity and Reduce Pollution in a Rice Production System. Irrig. Drain. 2012, 61, 285–293. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Q.; Zeng, X.; Bian, J.; Feng, Y. Effect of water-saving irrigation on growth and yield of rice in Heilongjiang Province. Heilongjiang Agric. Sci. 2010, 12, 14–16. [Google Scholar]
- Yang, S.; Peng, S.; Xu, J.; He, Y.; Wang, Y. Effects of water saving irrigation and controlled release nitrogen fertilizer managements on nitrogen losses from paddy fields. Paddy Water Environ. 2015, 13, 71–80. [Google Scholar] [CrossRef]
- Luo, Y.; Fu, H.; Xiong, Y.; Xiang, Z.; Wang, F.; Bugingo, Y.C.; Khan, S.; Cui, Y. Effects of water-saving irrigation on weed infestation and diversity in paddy fields in East China. Paddy Water Environ. 2016, 15, 1–12. [Google Scholar] [CrossRef]
- Parry, M.L.; Canziani, O.F.; Palutikof, J.P. (Eds.) Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2007. [Google Scholar]
- Emanuel, K. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 2005, 436, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Ghadikolaei, M.B.; Vaghefi, N.; Shamsudin, M.N.; Makmom, A. The economic impact of climate change on the rice production in Malaysia. Int. J. Agric. Res. 2011, 6, 67–74. [Google Scholar]
- International Food Policy Research Institute (IFPRI). Food Security, Farming and Climate Change to 2050, Scenarios, Results and Policy Options; IFPRI: Washington, DC, USA, 2010. [Google Scholar]
- Guo, X.P.; Zhen, B.; Lu, H. Research advances in pile-up effects of drought and waterlogging alternative stress on rice. Adv. Sci. Technol. Water Resour. 2013, 33, 83–86. [Google Scholar]
- Feng, G.; Yang, H.; Zhang, S.; Wang, K.; Shen, B. A preliminary research on the reason of a sharp turn from drought to flood in the middle and lower reaches of the Yangtze River in late spring and early summer of 2011. Chin. J. Atmos. Sci. 2012, 36, 1009–1026. [Google Scholar]
- He, H.; Lu, H. Characteristics of the Sharp Turn from Drought to Flood over Guangxi in Summer of 2013. Trop. Geogr. 2013, 4, 767–775. [Google Scholar]
- Linquist, B.A.; Phengsouvanna, V.; Sengxue, P. Benefits of organic residues and chemical fertilizer to productivity of rain-fed lowland rice and to soil nutrient balances. Nutr. Cycling Agroecosyst. 2007, 79, 59–72. [Google Scholar] [CrossRef]
- Hasegawa, T.; Sawano, S.; Goto, S.; Konghakote, P.; Polthanee, A.; Ishigooka, Y.; Kuwagata, T.; Toritani, H.; Furuya, J. A model driven by crop water use and nitrogen supply for simulating changes in the regional yield of rain-fed lowland rice in Northeast Thailand. Paddy Water Environ. 2008, 6, 73–82. [Google Scholar] [CrossRef]
- Ly, P.; Jensen, L.S.; Bruun, T.B.; Neergaard, A.D. Methane (CH4) and nitrous oxide (N2O) emissions from the system of rice intensification (SRI) under a rain-fed lowland rice ecosystem in Cambodia. Nutr. Cycling Agroecosyst. 2013, 97, 13–27. [Google Scholar] [CrossRef]
- Nagasawa, N.; Miyoshi, M.; Sano, Y.; Nagato, Y. DL regulates both leaf and pistil development in rice. Rice Genet. Newsl. 1996, 13, 102–105. [Google Scholar]
- Guo, X.P.; Yuan, J.; Guo, F. Preliminary study on water-catching and controlled irrigation technology of rice. Trans. CSAE 2009, 25, 70–73. [Google Scholar]
- Guo, X.; Wang, F.; Wang, Z.; Huang, S.; Liu, W. Effects of Irrigation Modes on Yield and Chlorophyll Fluorescence Characteristics after Heading Stage of Rice. J. Irrig. Drain. 2017, 36, 1–6. [Google Scholar]
- De Datta, S.K. Principles and Practices of Rice Production; John Wiley & Sons: New York, NY, USA, 1981; p. 618. [Google Scholar]
- He, J.; Shao, G.; Wei, X.; Huang, F.; Sheng, Z.; Tang, S.; Hu, P. Fine mapping and candidate gene analysis of qTAC8, a major quantitative trait locus controlling tiller angle in rice (Oryza sativa L.). PLoS ONE 2017, 12, 0178177. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Sun, A.; Zhang, Z.; Wang, Z.; Du, P. Effects of Different Water-saving Irrigation Modes on Rice Tillering, Height and Yield in Cold Area. Water Sav. Irrig. 2013, 12, 16–19. [Google Scholar]
- Biswas, A.K.; Choudhuri, M.A. Mechanism of Monocarpic Senescence in Rice. Plant Physiol. 1980, 65, 340–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohapatra, P.K.; Masamoto, Y.; Morita, S.; Takanashi, J.; Kato, T.; Itani, T.; Adu-Gyamfi, J.J.; Shunmugasundaram, M.; Nguyen, N.T.; Fujita, K. Partitioning of C-La-belled Photosynthate Varies with Growth Stage and Panicle Size in High Yielding Rice. Funct. Plant Biol. 2004, 31, 131–139. [Google Scholar] [CrossRef]
- Zhou, L.Q.; Shi, Z.; Tian, Y.F. Rapid estimation of rice canopy LAI using multi-source proximal sensors. In Precision Agriculture 13; Stafford, J.V., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2013; pp. 87–93. [Google Scholar]
- Zheng, Z.; Hiroyoshi, I.; Yutaka, H.; Yoshiyasu, T. Quantitative evaluation of the degree of sprout leaf bending of rice cultivars using P-type Fourier descriptors and rincipal component analysis. Euphytica 2008, 163, 259–266. [Google Scholar] [CrossRef]
- Dong, Y.; Kamiunten, H.; Ogawa, T.; Tsuzuki, E.; Terao, H.; Lin, D.; Matsuo, M. Mapping of QTLs for leaf developmental behavior in rice (Oryza sativa L.). Euphytica. 2004, 138, 169–175. [Google Scholar] [CrossRef]
- Li, Z.; Paterson, A.H.; Pinson, S.R.M.; Stanse, J.W. RFLP facilitated analysis of tiller and leaf angles in rice (Oryza sativa L.). Euphytica 1999, 109, 79–84. [Google Scholar] [CrossRef]
- Luo, X.; Zheng, J.; Huang, R.; Huang, Y.; Wang, H.; Jiang, L.; Fang, X. Phytohormones signaling and crosstalk regulating leaf angle in rice. Plant Cell Rep. 2016, 35, 2423–2433. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Navasero, S.A.; Ramirez, E.A. Effects of silica and nitrogen supply on some leaf characters of the rice plant. Plant Soil 1969, 31, 48–56. [Google Scholar] [CrossRef]
- Venkateswarlu, B.; Rao, P.K.; Rao, A.V. Canopy analysis on the relationships between leaf area index and productivity in lowland rice, Oryza sativa L. Plant Soil 1976, 45, 49–56. [Google Scholar] [CrossRef]
- Mdemu, M.V.; Magayane, M.D.; Lankford, B.; Hatibu, N.; Kadigi, R.M.J. Conjoining rainfall and irrigation seasonality to enhance productivity of water in rice irrigated farms in the Upper Ruaha River Basin, Tanzania. Phys. Chem. Earth Parts A/B/C 2004, 29, 1119–1124. [Google Scholar] [CrossRef]
- Suralta, R.R.; Yamauchi, A. Root growth, aerenchyma development, and oxygen transport in rice genotypes subjected to drought and waterlogging. Environ. Exp. Bot. 2008, 64, 75–82. [Google Scholar] [CrossRef]
- Li, S.; Li, Y.; Wu, H.; Liao, J.; Li, D. Optimum fertilization and its corresponding mechanism under complete submergence at booting stage in rice. Acta Agron. Sin. 2002, 8, 115–120. [Google Scholar]
- Ning, J.; Huo, Z.; Long, Z.; Shen, J.; Liu, B. The Preliminary Study in Rice Morphology under Waterlogging Stress. Chin. Agric. Sci. Bull. 2013, 29, 24–29. [Google Scholar]
- Chen, J.; Duan, R.; Peng, B.; Dong, S.; Lu, Z.; Zhang, Y.; Feng, D.; Xu, H.; Chen, S.; Zhang, Q.; et al. Study on the Intensified Cultivation Techniques of Hybrid Rice for High-yield and High Quality. Chin. Agric. Sci. Bull. 2009, 25, 138–142. [Google Scholar]
- Wei, M.; Liao, X.; Li, D.; Duan, H. Comparison of tillering productivity among nodes along the mian stem of rice. Chin. J. Plant Ecol. 2012, 36, 324–332. [Google Scholar] [CrossRef]
- Zhou, H.; Lu, X.; Zheng, Q. Studies on Tiller Regularity of Middle tillering Part and Productive Forces of Rice. Acta Agric. Boreal. Sin. 2000, 15, 112–117. [Google Scholar]
- Shao, G.; Cui, J.; Yu, S.; Lu, B.; Brian, B.J.; Ding, J.; She, D. Impacts of controlled irrigation and drainage on the yield and physiological attributes of rice. Agric. Water Manag. 2015, 149, 156–165. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Soil Depth/cm | Soil Texture | Organic Matter/(mg∙kg−1) | pH | Total Phosphorus/(mg∙kg−1) | Available P/(mg∙kg−1) | Total N/% | Available Nitrogen/(mg∙kg−1) |
|---|---|---|---|---|---|---|---|
| 0~20 | Clay | 8.06 | 8.06 | 330.9 | 10.13 | 0.1 | 65 |
| Meteorological Index | 2012 | 2013 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Apr | May | Jun | Jul | Aug | Sep | Oct | Apr | May | Jun | Jul | Aug | Sep | Oct | |
| Rain (mm) | 73 | 102 | 193 | 186 | 129 | 72 | 65 | 24.2 | 101.2 | 154.6 | 245.2 | 63.4 | 63.2 | 27.4 |
| Temperature (°C) | 15.5 | 21 | 25 | 28.5 | 28 | 23 | 17.5 | 13.4 | 18.6 | 21.9 | 26.9 | 29.1 | 24.8 | 20.7 |
| Humidity (%) | 73 | 74 | 78 | 81 | 81 | 79 | 77 | 62.6 | 78.7 | 82.6 | 72.2 | 72.8 | 79.6 | 76.7 |
| Treatment | Seedling | Early Tillering | Later Tillering | Elongation | Heading | Milky | Ripening |
|---|---|---|---|---|---|---|---|
| CK | 10~30~70 | 0~30~70 | 0~30~90 | 0~30~120 | 0~30~100 | 0~30~60 | 70~80% |
| T1 | 10~30~70 | 80%~100%~80 | 70%~100%~100 | 70%~100%~150 | 80%~100%~150 | 80%~100%~80 | 70~80% |
| T2 | 10~30~70 | 80%~100%~100 | 70%~100%~120 | 70%~100%~200 | 80%~100%~200 | 80%~100%~80 | 70~80% |
| Treatment | Days after Transplanting | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | 17 | 24 | 31 | 38 | 45 | 52 | 59 | 66 | 73 | 80 | |
| CK | 2.25 a | 4.25 a | 6.50 a | 7.50 a | 9.00 a | 9.50 b | 9.50 a | 8.75 a | 8.25 a | 8.25 a | 8.00 a |
| T1 | 2.50 a | 4.50 a | 6.75 a | 8.00 a | 9.25 a | 10.25 a | 9.75 a | 8.75 a | 8.50 a | 8.25 a | 8.25 a |
| T2 | 2.75 a | 4.00 a | 6.25 a | 7.75 a | 8.75 a | 9.75 b | 9.50 a | 8.75 a | 8.00 a | 7.75 a | 8.00 a |
| Treatment | Days after Transplanting | |||||||
|---|---|---|---|---|---|---|---|---|
| 25 | 31 | 38 | 49 | 57 | 73 | 94 | 119 | |
| CK | 7.33 a | 7.33 a | 8.33 ab | 8.00 a | 7.67 a | 7.00 ab | 8.00 a | 8.00 a |
| T1 | 7.67 a | 8.00 a | 9.33 a | 8.33 a | 8.00 a | 8.33 a | 8.33 a | 8.33 a |
| T2 | 5.33 a | 6.67 a | 7.33 b | 7.00 a | 6.33 a | 6.67 b | 6.67 a | 6.67 a |
| Year | Treatment | Days after Transplanting | ||
|---|---|---|---|---|
| 31 | 38 | 73 | ||
| 2012 | CK | 7.50 a | 9.00 a | 8.25 a |
| T1 | 8.00 a | 9.25 a | 8.25 a | |
| T2 | 7.75 a | 8.75 ab | 7.75 ab | |
| 2013 | CK | 7.33 a | 8.33 ab | 7.00 ab |
| T1 | 8.00 a | 9.33 a | 8.33 a | |
| T2 | 6.67 a | 7.33 b | 6.67 b | |
| DAT | Treatment | t | LAI | Canopy Interception | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 20 | 40 | 60 | 80 | ||||
| 52 | CK | 0.71 ± 0.07 a | 0.99 ± 0.36 a | 0.29 ± 0.07 a | ||||
| T1 | 0.60 ± 0.12 a | 1.48 ± 0.45 a | 0.39 ± 0.12 a | |||||
| T2 | 0.73 ± 0.19 a | 0.95 ± 0.77 a | 0.28 ± 0.19 a | |||||
| 106 | CK | 0.34 ± 0.11 a | 2.67 ± 0.92 a | 0.66 ± 0.12 a | 0.52 ± 0.09 a | 0.35 ± 0.09 a | 0.16 ± 0.05 ab | 0.03 ± 0.02 a |
| T1 | 0.29 ± 0.07 a | 2.93 ± 0.70 a | 0.71 ± 0.07 a | 0.51 ± 0.03 a | 0.38 ± 0.05 a | 0.10 ± 0.03 b | 0.04 ± 0.04 a | |
| T2 | 0.43 ± 0.10 a | 2.00 ± 0.56 a | 0.57 ± 0.11 a | 0.51 ± 0.11 a | 0.35 ± 0.16 a | 0.18 ± 0.01 a | 0.04 ± 0.02 a | |
| DAT | Treatment | t | LAI | Canopy Interception at Different Height (cm) | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 20 | 40 | 60 | ||||
| 64 | CK | 0.39 ± 0.09 a | 2.15 ± 0.49 b | 0.61 ± 0.08 b | 0.52 ± 0.03 b | 0.29 ± 0.08 b | 0.05 ± 0.02 b | |
| T1 | 0.24 ± 0.05 b | 3.38 ± 0.38 a | 0.76 ± 0.05 a | 0.73 ± 0.05 a | 0.60 ± 0.05 a | 0.13 ± 0.08 a | ||
| T2 | 0.40 ± 0.04 a | 2.11 ± 0.23 b | 0.60 ± 0.04 b | 0.54 ± 0.03 b | 0.30 ± 0.05 b | 0.07 ± 0.04 b | ||
| 79 | CK | 0.20 ± 0.05 b | 3.30 ± 0.58 ab | 0.80 ± 0.03 a | 0.76 ± 0.05 ab | 0.64 ± 0.07 a | 0.50 ± 0.10 ab | 0.14 ± 0.04 a |
| T1 | 0.18 ± 0.03 b | 3.90 ± 0.17 a | 0.82 ± 0.03 a | 0.82 ± 0.03 a | 0.76 ± 0.05 a | 0.65 ± 0.07 a | 0.17 ± 0.11 a | |
| T2 | 0.28 ± 0.02 a | 2.69 ± 0.27 b | 0.72 ± 0.02 b | 0.69 ± 0.03 b | 0.63 ± 0.07 a | 0.47 ± 0.09 b | 0.13 ± 0.03 a | |
| 93 | CK | 0.23 ± 0.09 a | 2.83 ± 0.52 a | 0.77 ± 0.06 a | 0.74 ± 0.09 a | 0.67 ± 0.09 a | 0.23 ± 0.18 a | 0.10 ± 0.07 a |
| T1 | 0.22 ± 0.04 a | 3.18 ± 0.33 a | 0.78 ± 0.04 a | 0.78 ± 0.04 a | 0.69 ± 0.08 a | 0.45 ± 0.17 a | 0.17 ± 0.09 a | |
| T2 | 0.25 ± 0.09 a | 2.95 ± 0.75 a | 0.75 ± 0.09 a | 0.74 ± 0.09 a | 0.61 ± 0.15 a | 0.41 ± 0.15 a | 0.10 ± 0.06 a | |
| Year | DAT | Interception Rate at Different Height (cm) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 20 | 40 | 60 | 80 | |||
| 2012 | 52 | LAI | 0.993 ** | |||||
| 106 | 0.977 ** | 0.582 | 0.509 | 0.004 | 0.019 | |||
| 2013 | 64 | 0.992 ** | 0.850 ** | 0.826 ** | 0.412 | |||
| 79 | 0.915 ** | 0.979 ** | 0.760 * | 0.828 ** | 0.593 | |||
| 93 | 0.954 ** | 0.973 ** | 0.886 ** | 0.538 | 0.454 | |||
| Year | Treatments | Actual Yield (t/hm2) | Theoretical Yield (t/hm2) | Ear Length (cm) |
|---|---|---|---|---|
| 2012 | CK | 4.79 ab | 5.12 ab | 17.80 c |
| T1 | 5.16 a | 5.35 a | 18.95 b | |
| T2 | 4.68 b | 5.05 b | 19.54 a | |
| 2013 | CK | 3.11 c | 3.56 b | 17.23 b |
| T1 | 4.39 a | 4.54 a | 18.61 a | |
| T2 | 3.73 b | 4.33 ab | 18.54 a |
| Treatment | Tiller Number | Stem Mass/g | Leaf Dry Quality/g | Spike Quality/g | Filled Grain per Spike | Shriveled Grain per Spike | Seed Setting Rate | 1000 Grain Quality/g |
|---|---|---|---|---|---|---|---|---|
| CK | 120.75 ± 11.62 a | 285.00 ± 42.00 a | 52.28 ± 5.62 a | 355.75 ± 23.62 b | 122.33 ± 24.14 ab | 28.71 ± 18.77 a | 0.82 ± 0.11 a | 26.51 ± 2.15 a |
| T1 | 143.67 ± 19.76 a | 373.25 ± 57.28 a | 76.70 ± 21.46 a | 453.87 ± 73.71 a | 107.70 ± 25.89 b | 23.26 ± 18.37 a | 0.83 ± 0.12 a | 27.44 ± 3.45 a |
| T2 | 126.67 ± 2.31 a | 306.18 ± 43.37 a | 70.60 ± 14.01 a | 432.63 ± 42.99 ab | 130.96 ± 29.61 a | 29.46 ± 23.58 a | 0.83 ± 0.12 a | 26.66 ± 0.79 a |
| Treatment | High Tiller | Without High Tiller | ||||||
|---|---|---|---|---|---|---|---|---|
| High Tiller Number | Stem Mass | Stem Length | Leaf Quality | Spike Quality | Stem Mass | Leaf Quality | Spike Quality | |
| CK | 66.75 ± 15.95 a | 34.66 ± 10.35 a | 35.55 ± 2.49 a | 8.92 ± 2.65 a | 32.98 ± 10.74 a (9.27%) | 250.34 ± 45.65 b | 43.36 ± 5.51 b | 322.78 ± 22.97 b (90.73%) |
| T1 | 74.33 ± 31.88 a | 32.26 ± 15.89 a | 33.17 ± 4.80 a | 8.94 ± 4.42 a | 29.30 ± 16.08 a (6.46%) | 340.99 ± 41.45 a | 67.75 ± 17.30 a | 424.57 ± 60.97 a (93.54%) |
| T2 | 72.00 ± 15.87 a | 34.65 ± 8.67 a | 34.17 ± 1.26 a | 9.34 ± 0.99 a | 36.93 ± 5.21 a (8.54%) | 271.53 ± 41.70 ab | 61.26 ± 13.39 ab | 395.70 ± 47.78 ab (91.46%) |
| Index | Total Tiller Number | High Tiller | Total Stem Mass | Total Leaf Quality | Total Spike Quality | Irrigation Amount | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Tiller Number | Stem Mass | Stem Length | Leaf Quality | Spike Quality | |||||||
| Total tiller number | 1 | 0.614 | 0.373 | 0.227 | 0.415 | 0.248 | 0.908 ** | 0.849 ** | 0.821 ** | 0.466 | |
| High tiller | Tiller number | 1 | 0.951 ** | 0.692 * | 0.931 ** | 0.884 ** | 0.502 | 0.698 * | 0.454 | 0.054 | |
| Stem mass | 1 | 0.808 ** | 0.953 ** | 0.956 ** | 0.243 | 0.469 | 0.226 | −0.095 | |||
| Stem length | 1 | 0.828 ** | 0.825 ** | 0.073 | 0.170 | 0.142 | −0.149 | ||||
| Leaf quality | 0.964 ** | 0.295 | 0.548 | 0.391 | −0.059 | ||||||
| Spike quality | 1 | 0.116 | 0.433 | 0.264 | −0.294 | ||||||
| Total stem mass | 1 | 0.855 ** | 0.734 * | 0.493 | |||||||
| Total leaf quality | 1 | 0.864 ** | 0.167 | ||||||||
| Total leaf quality | 1 | 0.160 | |||||||||
| Irrigation amount | 1 | ||||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, H.; Qi, X.; Guo, X.; Towa, J.J.; Zhen, B.; Qiao, D.; Wang, Z.; Yang, B.; Han, Y. Canopy Light Utilization and Yield of Rice under Rain-Catching and Controlled Irrigation. Water 2018, 10, 1340. https://doi.org/10.3390/w10101340
Lu H, Qi X, Guo X, Towa JJ, Zhen B, Qiao D, Wang Z, Yang B, Han Y. Canopy Light Utilization and Yield of Rice under Rain-Catching and Controlled Irrigation. Water. 2018; 10(10):1340. https://doi.org/10.3390/w10101340
Chicago/Turabian StyleLu, Hongfei, Xuebin Qi, Xiangping Guo, Jacob Jeremiah Towa, Bo Zhen, Dongmei Qiao, Zhenchang Wang, Bo Yang, and Yang Han. 2018. "Canopy Light Utilization and Yield of Rice under Rain-Catching and Controlled Irrigation" Water 10, no. 10: 1340. https://doi.org/10.3390/w10101340
APA StyleLu, H., Qi, X., Guo, X., Towa, J. J., Zhen, B., Qiao, D., Wang, Z., Yang, B., & Han, Y. (2018). Canopy Light Utilization and Yield of Rice under Rain-Catching and Controlled Irrigation. Water, 10(10), 1340. https://doi.org/10.3390/w10101340