
Composition and Concentration of the Biogenic Components of the Aerosols Collected over Vasyugan Marshes and Karakan Pine Forest at Altitudes from 500 to 7000 m

 ,
,  , , and
, , and
Abstract
1. Introduction
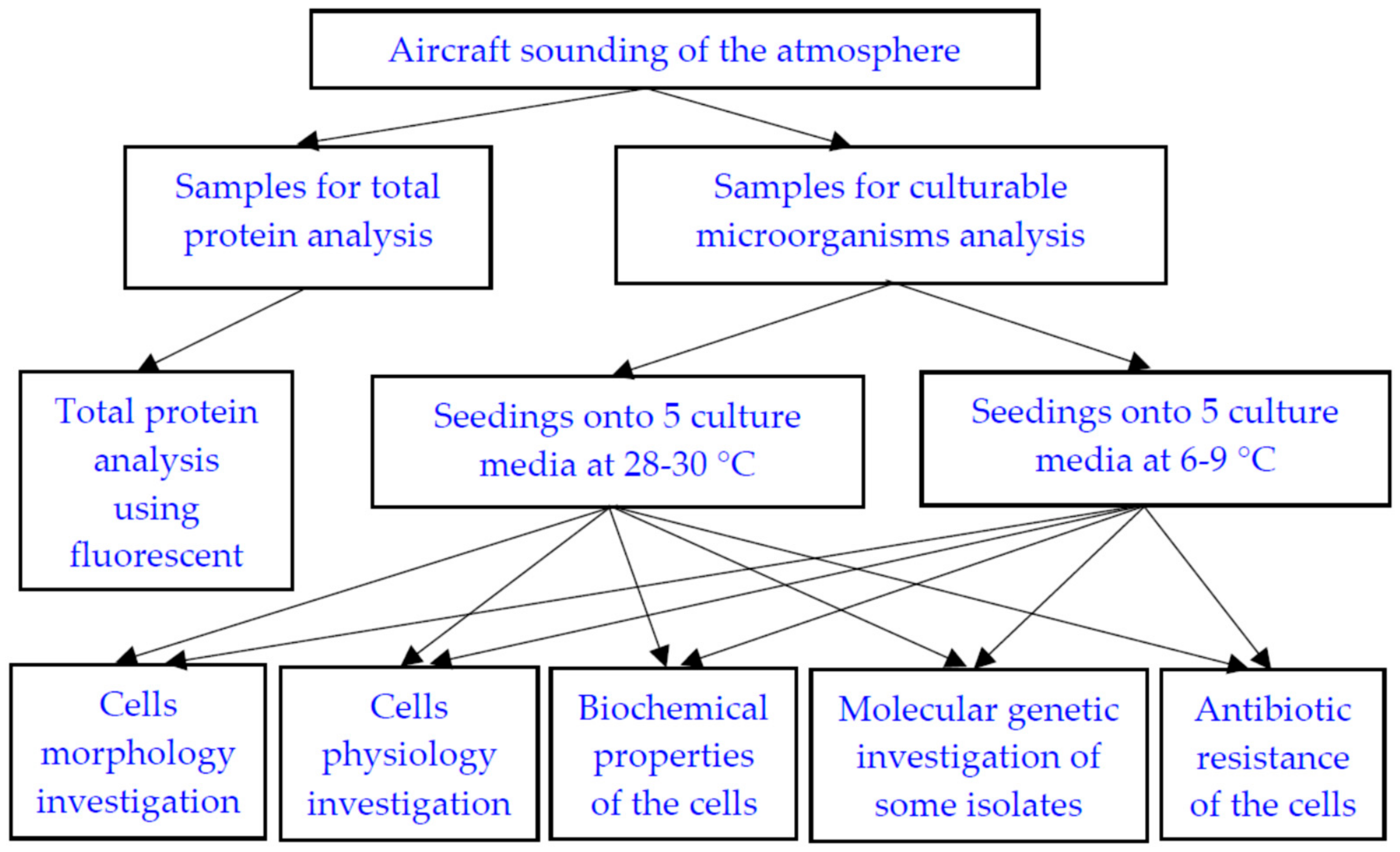
2. Materials and Methods
3. Results and Discussion
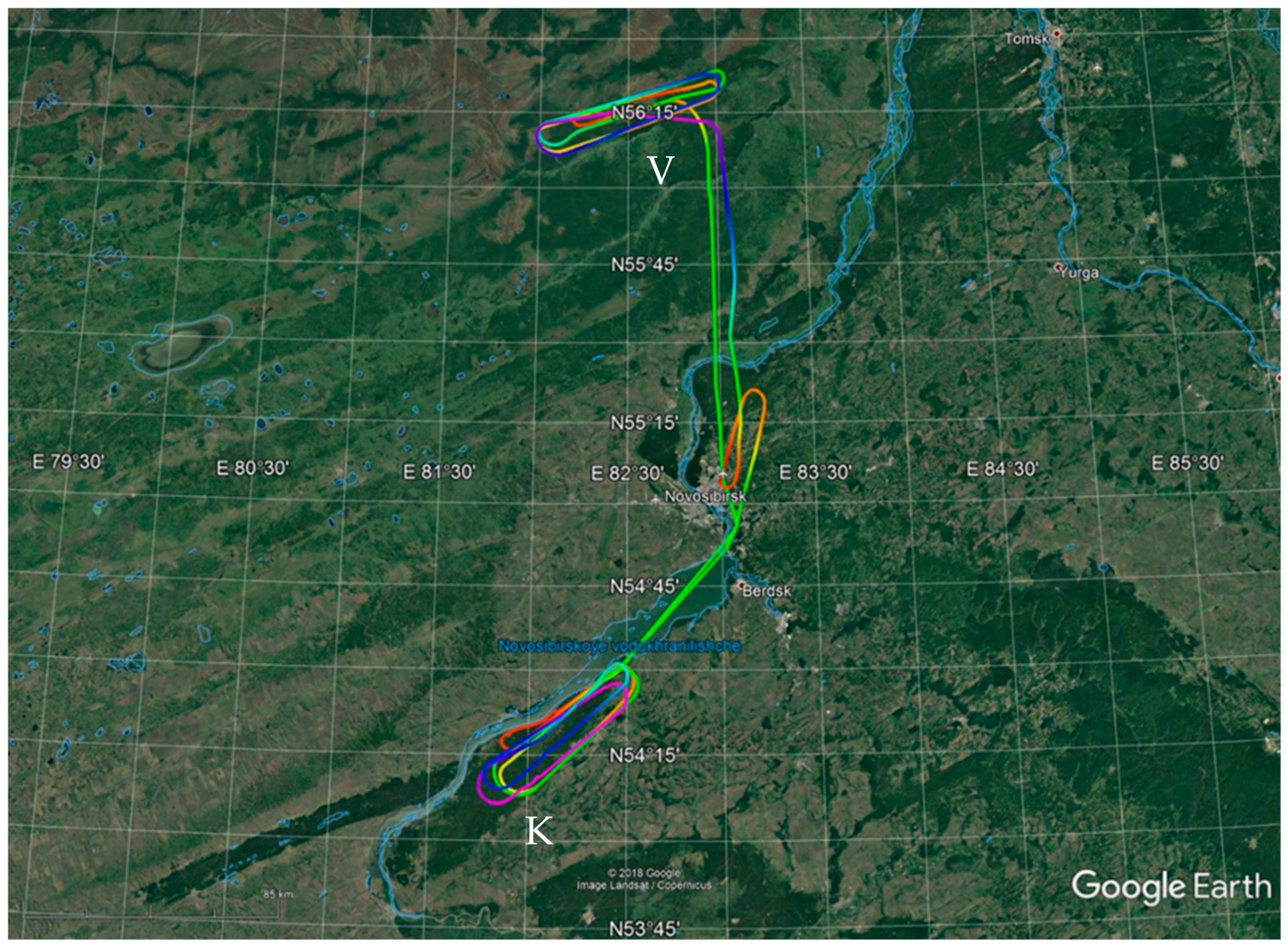
3.1. Backward Trajectories Analysis
3.2. Total Potein Cocentrations
3.3. Culturable Microorganisms Cocentrations
3.4. Culturable Microorganisms Diversity
3.5. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Després, V.R.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.S.; Buryak, G.; Fröhlich-Nowoisky, J.; Elbert, W.; Andreae, M.O.; Pöschl, U.; et al. Primary biological aerosols in the atmosphere: A review. Tellus. B 2012, 64, 15598. [Google Scholar] [CrossRef]
- Dommergue, A.; Amato, P.; Tignat-Perrier, R.; Magand, O.; Thollot, A.; Joly, M.; Bouvier, L.; Sellegri, K.; Vogel, T.; Sonke, J.E.; et al. Methods to investigate the global atmospheric microbiome. Front. Microbiol. 2019, 10, 243. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Burrows, S.M.; Xie, Z.; Engling, G.; Solomon, P.A.; Fraser, M.P.; Mayol-Bracero, O.L.; Artaxo, P.; Begerow, D.; Conrad, R.; et al. Biogeography in the air: Fungal diversity over land and oceans. Biogeosciences 2012, 9, 1125–1136. [Google Scholar] [CrossRef]
- Hyde, P.; Mahalov, A. Contribution of bioaerosols to airborne particulate matter. J. Air Waste Manag. Assoc. 2020, 70, 71–77. [Google Scholar] [CrossRef]
- Pearce, D.A.; Alekhina, I.A.; Terauds, A.; Wilmotte, A.; Quesada, A.; Edwards, A.; Dommergue, A.; Sattler, B.; Adams, B.J.; Magalhães, C.; et al. Aerobiology over Antarctica—A new initiative for atmospheric ecology. Front. Microbiol. 2016, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Safatov, A.; Andreeva, I.; Buryak, G.; Olkin, S.; Reznikova, I.; Belan, B.; Panchenko, M.; Simonenkov, D. Long-term studies of biological components of atmospheric aerosol: Trends and variability. Atmosphere 2022, 13, 651. [Google Scholar] [CrossRef]
- Andreeva, I.S.; Safatov, A.S.; Puchkova, L.I.; Emelyanova, E.K.; Buryak, G.A.; Olkin, S.E.; Reznikova, I.K.; Ohlopkova, O.V. Culturable microorganisms in high-altitude atmospheric aerosol samples collected above northern Siberia by aircraft sounding. Bull. Nizhnevartovsk. State Univer. 2019, 2, 3–11. (In Russian) [Google Scholar] [CrossRef]
- Andreeva, I.S.; Safatov, A.S.; Morozova, V.V.; Tikunova, N.V.; Emelyanova, E.K.; Solovyanova, N.A.; Babkin, I.V.; Buryak, G.A.; Puchkova, L.I. Saprophytic and pathogenic yeasts in atmospheric aerosols of Southwestern Siberia. Atmos. Ocean. Opt. 2020, 33, 505–511. [Google Scholar] [CrossRef]
- Inisheva, L.I. Vasyugan Bog Nature Conditions, Structure and Functioning; TsNTI: Tomsk, Russia, 2003. (In Russian) [Google Scholar]
- Bodelier, P.L.E.; Dedysh, S.N. Microbiology of wetlands. Front. Microbiol. 2013, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Tfaily, M.M.; Green, S.J.; Steinweg, J.M.; Chanton, P.; Imvittaya, A.; Chanton, J.P.; Cooper, W.; Schadt, C.; Kostka, J.E. Microbial metabolic potential for carbon degradation and nutrient (nitrogen and phosphorus) acquisition in an ombrotrophic peatland. Appl. Environ. Microbiol. 2014, 80, 3531–3540. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaya, T.G.; Kukharenko, O.S.; Golovchenko, A.V. Features of the taxonomic composition of bacterial complexes of peatlands of different genesis. In Proceedings of the Wetlands and Biosphere: Collection of Materials of the Fourth Scientific School, Tomsk, Russia, 12–15 September 2005. (In Russian). [Google Scholar]
- Grodnitskaya, I.D.; Trusova, M.Y. Microbial communities and transformation of carbon compounds in bog soils of the taiga zone (Tomsk oblast). Eurasian Soil Sci. 2009, 42, 1021–1028. (In Russian) [Google Scholar] [CrossRef]
- Dobrovolskaya, T.G.; Golovchenko, A.V.; Kukharenko, O.S.; Yakushev, A.V.; Semenova, T.A.; Inisheva, L.I. The structure of the microbial communities in low-moor and high-moor peat bogs of Tomsk oblast. Eurasian Soil Sci. 2012, 45, 273–281. (In Russian) [Google Scholar] [CrossRef]
- Sergeeva, M.A.; Khokhlova, A.M. Microbial biomass and its activity in peat bogs of Siberia. TSPU Bull. 2015, 2, 143–149. (In Russian) [Google Scholar]
- Dedysh, S.N. Cultivating uncultured bacteria from Northern wetlands: Knowledge gained and remaining gaps. Front. Microbiol. 2011, 2, 184. [Google Scholar] [CrossRef]
- Alekseeva, M.N.; Preis, Y.I.; Dyukarev, E.A. Spatial structure of vegetation cover and top layer of peat of northeastern spurs of the great Vasyugan mire by remote and ground data. Bull. Tomsk. Polytech. Univ. 2015, 326, 81–90. (In Russian) [Google Scholar]
- Sergeeva, M.A.; Inisheva, L.I. Biochemical processes in oligotrophic peat deposits of the Vasyugan bog. TSPU Bull. 2008, 4, 57–63. (In Russian) [Google Scholar]
- Andreeva, I.S.; Safatov, A.S.; Puchkova, L.I.; Emelyanova, E.K.; Buryak, G.A.; Ternovoi, V.A. Spore forming bacteria isolated from atmospheric aerosols in Southwestern Siberia during atmospheric transfer of air masses. Bull. Nizhnevartovsk State Univer. 2018, 3, 123–130. (In Russian) [Google Scholar]
- Shevchenko, V.P.; Vorob′ev, S.N.; Kirpotin, S.N.; Kritskov, I.V.; Manasypov, R.M.; Pokrovsky, O.S.; Politova, N.V. Investigations of insoluble particles in the snow cover of the Western Siberia from Tomsk to the Ob estuary. Atmos. Ocean. Opt. 2015, 28, 499–504. (In Russian) [Google Scholar]
- Moskovchenko, D.V.; Babushkin, A.G. Peculiarities of formation of chemical composition of snow waters (on example of Khanty-Mansi autonomous district). Earth′s Cryosphere 2012, XVI, 71–81. (In Russian) [Google Scholar]
- Korica, A.; Polis, O.; Spalvis, K.; Bartkevics, V. Quantitative and qualitative seasonal changes of Scots pine and Norway spruce foliage essential oils in Latvia, and the extraction dynamics thereof. Balt. For. 2015, 21, 51–58. [Google Scholar]
- Apetrei, L.C.; Tuchiluş, C.; Aprotosoaie, A.C.; Oprea, A.; Malterud, K.E.; Miron, A. Chemical, antioxidant and antimicrobial investigations of Pinus cembra L. bark and needles. Molecules 2011, 16, 7773–7788. [Google Scholar] [CrossRef] [PubMed]
- Mimoune, N.A.; Mimoune, D.A.; Yataghene, A. Chemical composition and antimicrobial activity of the essential oils of Pinus pinaster. J. Coast. Life Med. 2013, 1, 55–59. [Google Scholar] [CrossRef]
- Vainio-Kaila, T.; Kyyhkynen, A.; Rautkari, L.; Siitonen, A. Antibacterial effects of extracts of Pinus sylvestris and Picea abies against Staphylococcus aureus, Enterococcus faecalis, Escherichia coli, and Streptococcus pneumoniae. BioResources 2015, 10, 7763–7771. [Google Scholar] [CrossRef]
- Efremov, A.A.; Zykova, I.D.; Senashova, V.A.; Grodnickaya, I.D.; Pashenova, N.V. Antimicrobial and antiradical activity of individual fractions of pinus sibirica du tour and abies sibirica ledeb., native to the Siberian region. Chem. Plant Mater. 2020, 4, 203–210. [Google Scholar] [CrossRef]
- Tarakanov, V.V.; Chindyaeva, L.N.; Tsybulya, N.V.; Tikhonova, I.V. Variability of needle antimicrobic activity in clone plantation of Scots pine Pinus sylvestris L. Sib. J. For. Sci. 2017, 1, 95–104. (In Russian) [Google Scholar] [CrossRef]
- Draxler, R.R.; Hess, G.D. An overview of the HYSPLIT_4 modelling system for trajectories, dispersion and deposition. Aust. Meteorol. Mag. 1998, 47, 295–308. [Google Scholar]
- Belan, B.D.; Rasskazchikova, T.M.; Simonenkov, D.V.; Tolmachev, G.N.; Fofonov, A.V. Chemical composition of atmospheric aerosols over background areas of the southern part of Western Siberia observed during the IAO complex atmospheric radiation experiment carried out in December 2015. Proc. SPIE 2016, 10035, 1003549. [Google Scholar] [CrossRef]
- Freitag, S.; Clarke, A.D.; Howell, S.G.; Kapustin, V.N.; Campos, T.; Brekhovskikh, V.L.; Zhou, J. Combining airborne gas and aerosol measurements with HYSPLIT: A visualization tool for simultaneous evaluation of air mass history and back trajectory consistency. Atmos. Meas. Tech. 2014, 7, 107–128. [Google Scholar] [CrossRef]
- Antokhin, P.N.; Arshinov, M.Y.; Belan, B.D.; Davydov, D.K.; Zhidovkin, E.V.; Ivlev, G.A.; Kozlov, A.V.; Kozlov, V.S.; Panchenko, M.V.; Penner, I.E.; et al. Optik-É AN-30 Aircraft Laboratory for Studies of the Atmospheric Composition. J. Atmos. Ocean. Technol. 2012, 29, 64–75. [Google Scholar] [CrossRef]
- APA. Filters. Catalogue-Guide; Atomizdat: Moscow, Russia, 1970. (In Russian) [Google Scholar]
- You, W.W.; Haugland, R.P.; Ryan, D.K.; Haugland, R.P. 3-(4-Carboxybenzoyl)quinoline-2-carboxaldehyde, a reagent with broad dynamic range for the assay of proteins and lipoproteins in solution. Ann. Biochem. 1997, 244, 277–282. [Google Scholar] [CrossRef]
- Ashmarin, I.P.; Vorobyov, A.A. Statistical Methods in Microbiological Studies; Medgiz: Leningrad, Russia, 1962. (In Russian) [Google Scholar]
- Babieva, I.P.; Chernov, I.Y. Biology of Yeast; Association of Scientific Publications KMK: Moscow, Russia, 2004. (In Russian) [Google Scholar]
- Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G. Guide to Clinically Significant Fungi; Williams & Wilkins: Philadelphia, PA, USA; A Waverly Company: Baltimore, MD, USA, 1998. [Google Scholar]
- Gerhardt, F.; Murray, R.G.E.; Wood, W.A.; Krieg, N.R. Methods of General Bacteriology, 2nd ed.; American Society for Microbiology: Washington, DC, USA, 1994.
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology; Williams & Wilkins: Philadelphia, PA, USA, 1994. [Google Scholar]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef]
- Hara, K.; Zhang, D. Bacterial abundance and viability in long-range transported dust. Atmos. Environ. 2012, 47, 20–25. [Google Scholar] [CrossRef]
- Jeon, E.M.; Kim, H.J.; Jung, K.; Kim, J.H.; Kim, M.Y.; Kim, Y.P.; Ka, J.-O. Impact of Asian dust events on airborne bacterial community assessed by molecular analyses. Atmos. Environ. 2011, 45, 4313–4321. [Google Scholar] [CrossRef]
- Visca, P.; Seifert, H.; Towner, K.J. Acinetobacter infection—An emerging threat to human health. IUBMB Life 2011, 63, 1048–1054. [Google Scholar] [CrossRef]
- Golovchenko, A.V.; Zvyagintsev, D.G.; Tikhonova, E.Y. Abundance, biomass, structure, and activity of the microbial complexes of minerotrophic and ombrotrophic peatlands. Microbiology 2007, 76, 630–637. [Google Scholar] [CrossRef]
- Golovchenko, A.V.; Semenova, T.A.; Polyakova, A.V.; Inisheva, L.I. The structure of the micromycete complexes of oligotrophic peat deposits in the southern taiga subzone of West Siberia. Microbiology 2002, 71, 667–674. (In Russian) [Google Scholar] [CrossRef]
- Polyakova, A.V.; Chernov, I.Y.; Panikov, N.S. Yeast diversity in hydromorphic soils with reference to a grass-sphagnum wetland in Western Siberia and a hummocky tundra region at cape Barrow (Alaska). Microbiology 2001, 70, 714–720. (In Russian) [Google Scholar] [CrossRef]
- Polyakova, A.V. Yeast in Peat-Bog Soils. Ph.D. Thesis, Institute of Microbiology of the Russian Academy of Sciences, Moscow, Russia, 2002. (In Russian). [Google Scholar]
- Yakushev, A.V.; Matyshak, G.V.; Tarkhov, M.O.; Kachalkin, A.V.; Sefilyan, A.R.; Petrov, D.G. Microbiological characteristics of bare peat circles on flat-topped peat mounds in the north of Western Siberia. Eurasian Soil Sci. 2019, 52, 1081–1090. [Google Scholar] [CrossRef]
- Bagirova, N.S. Aureobasidium pullulans place among human pathogens opportunistic infections (literature review). Lab. Serv. 2018, 7, 12–18. (In Russian) [Google Scholar] [CrossRef]
- Soeria-Atmadja, D.; Önell, A.; Borga, A. IgE sensitization to fungi mirrors fungal phylogenetic systematics. J. Allergy Clin. Immunol. 2010, 125, 1379–1386. [Google Scholar] [CrossRef]
- Javad Najafzadeh, M.; Sutton, D.A.; Saradeghi Keisari, M.; Zarrinfar, H.; Sybren de Hoog, G.; Chowdhary, A.; Meise, J.F. In vitro activities of eight antifungal drugs against 104 environmental and clinical isolates of Aureobasidium pullulans. Antimicrob. Agents Chemother. 2014, 58, 5629–5631. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Mendez, M.; Kibbler, C.; Erzsebet, P.; Chang, S.-C.; Gibbs, D.L.; Newell, V.A. Candida guilliermondii, an opportunistic fungal pathogen with decreased susceptibility to fluconazole: Geographic and temporal trends from the ARTEMIS DISK antifungal surveillance program. J. Clin. Microbiol. 2006, 44, 3551–3556. [Google Scholar] [CrossRef]
- Gamaletsou, M.N.; Kontoyiannis, D.P.; Sipsas, N.V.; Moriyama, B.; Alexander, E.; Roilides, E.; Brause, B.; Walsh, T.J. Candida osteomyelitis: Analysis of 207 pediatric and adult cases (1970–2011). Clin. Infect. Dis. 2012, 55, 1338–1351. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sampling Altitude (m) | Total Protein Concentration, μg/m3 | |
|---|---|---|
| Vasyugan Marshes | Karakan Pine Forest | |
| 7000 | <0.001 | 0.037 |
| 5500 | <0.001 | <0.001 |
| 4000 | <0.001 | 0.219 |
| 3000 | <0.001 | <0.001 |
| 2000 | 0.198 | 0.130 |
| 1500 | 0.113 | <0.001 |
| 1000 | 0.020 | 0.290 |
| 500 | 0.136 | 0.358 |
| Region of Sampling | Altitude, m | Total Number of Isolated Microorganisms, CFU/m3 | Titer of Isolated Microorganisms on of Different Culture Media, CFU/m3 | ||||
|---|---|---|---|---|---|---|---|
| GFM * | LB 1:10 | SAA | SA | Sabouraud | |||
| Cultivation temperature 28–30 °C | |||||||
| Vasyugan marshes | 5500 | 6.1 × 106 | 4.2 × 106 | 1.5 × 105 | 2.0 × 106 | 3.3 × 106 | 2.3 × 105 |
| 3000 | 4.7 × 105 | 6.9 × 104 | 1.5 × 106 | 1.4 × 106 | 1.6 × 106 | 1.2 × 105 | |
| 1000 | 2.3 × 105 | 0 | 1.1 × 105 | 0 | 8.4 × 104 | 3.3 × 104 | |
| 500 | 4.8 × 106 | 5.7 × 104 | 2.4 × 105 | 2.6 × 106 | 1.7 × 107 | 2.7 × 106 | |
| Karakan pine forest | 5500 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4000 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3000 | 5.0 × 103 | 0 | 0 | 0 | 5.0 × 103 | 0 | |
| 1000 | 3.7 × 105 | 4.0 × 103 | 3.0 × 105 | 2.0 × 103 | 3.3 × 104 | 3.2 × 104 | |
| 500 | 8.3 × 105 | 9.7 × 104 | 2.9 × 104 | 3.0 × 104 | 8.5 × 104 | 3.3 × 105 | |
| Cultivation temperature 6–9 °C | |||||||
| Vasyugan marshes | 5500 | 5.4 × 106 | 5.5 × 105 | 3.1 × 105 | 2.0 × 107 | 2.2 × 107 | 3.2 × 106 |
| 3000 | 2.1 × 106 | 0 | 1.1 × 106 | 1.0 × 106 | 0 | 7.4 × 103 | |
| 1000 | 3.4 × 105 | 2.0 × 103 | 1.5 × 105 | 0 | 2.4 × 104 | 1.6 × 105 | |
| 500 | 1.9 × 106 | 2.5 × 104 | 8.8 × 104 | 0 | 1.5 × 106 | 2.9 × 105 | |
| Karakan pine forest | 5500 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4000 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3000 | 1.7 × 103 | 0 | 0 | 0 | 1.7 × 103 | 0 | |
| 1000 | 1.9 × 106 | 2.8 × 104 | 8.3 × 105 | 0 | 6.3 × 105 | 3.8 × 105 | |
| 500 | 2.0 × 106 | 5.2 × 104 | 7.0 × 105 | 1.5 × 105 | 8.4 × 105 | 2.7 × 106 | |
| Region of Sampling | Altitude, m | Titer, CFU/m3, 6–9 °C | Titer, CFU/m3, 28–30 °C | ||
|---|---|---|---|---|---|
| Bacteria | Fungi | Bacteria | Fungi | ||
| Vasyugan marshes | 5500 | 3.6 × 106 | 1.8 × 106 | 5.8 × 106 | 3.0 × 105 |
| 3000 | 2.1 × 106 | 4.6 × 103 | 4.5 × 106 | 2.0 × 105 | |
| 1000 | 9.6 × 104 | 2.4 × 105 | 8.8 × 104 | 1.4 × 105 | |
| 500 | 1.5 × 106 | 4.1 × 105 | 4.4 × 106 | 4.3 × 105 | |
| Karakan pine forest | 5500 | 0 | 0 | 0 | 0 |
| 4000 | 0 | 0 | 0 | 0 | |
| 3000 | 0 | 1.7 × 103 | 3.3 × 103 | 1.7 × 103 | |
| 1000 | 2.5 × 105 | 1.6 × 106 | 2.8 × 105 | 9.1 × 104 | |
| 500 | 1.3 × 106 | 7.7 × 105 | 3.0 × 105 | 5.3 × 105 | |
| Strain | Class | Family | Genus | Species |
|---|---|---|---|---|
| Dr 9-1 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. leucospermi |
| Dr 9-2 | Tremellomycetes | Rhynchogastrema-taceae | Papiliotrema | P. flavescens/P. aurea |
| Dr 9-5 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. leucospermi |
| Dr 9-6 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. leucospermi |
| Dr 9-26 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. leucospermi |
| Dr 9-30 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. pullulans |
| Dr 10-13 | Tremellomycetes | Bulleribasidiaceae | Vishniacozyma | V. (Cryptococcus) heimaeyensis |
| Dr 10-15 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. pullulans |
| Dr 10-16M | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. pullulans |
| Dr 11-7 | Tremellomycetes | Bulleraceae | Bullera | B. alba |
| Dr 11-8 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. pullulans |
| Dr 11-12 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. pullulans |
| Dr 11-13 | Tremellomycetes | Bulleribasidiaceae | Vishniacozyma | V. (Cryptococcus) heimaeyensis |
| MR 12 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. pullulans |
| MR-14 | Dothideomycetes | Saccotheciaceae | Auerobasidium | A. leucospermi |
| MR 58 | Saccharomycetes | Debaryomycetaceae | Meyerozyma | M. (Pichia) guilliermondii |
| MR 135 | Tremellomycetes | Bulleribasidiaceae | Vishniacozyma | V. victoriae |
| MR 160 | Tremellomycetes | Cystofilobasidiaceae | Cystofilobasidium | C. macerans |
| MR 166 | Tremellomycetes | Cystofilobasidiaceae | Cystofilobasidium | C. macerans |
| MR 189 | Tremellomycetes | Filobasidiaceae | Filobasidium | F. (Cryptococcus) magnum |
| Strain | Antibiotic, Amount on Disk, μg/Diameter of Zone of Growth Inhibition, mm | |||||
|---|---|---|---|---|---|---|
| Derivatives of Triazole | Derivatives of Imidazole | Polyene Antibiotics | ||||
| Fluconazole, 40 | Itraconazole, 10 | Ketoconazole, 20 | Clotrimazole, 10 | Nystatin, 80 | Amphotericin B, 40 | |
| Dr 9-1 | 0 | 0 | 35 | 10 | 28 | 8 |
| Dr 9-5 | 0 | 10 | 12 | 12 | 12 | 12 |
| Dr 9-6 | 0 | 0 | 25 | 11 | 26 | 10 |
| Dr 9-2 | 0 | 0 | 30 | 18 | 25 | 0 |
| Dr 9-26 | 0 | 0 | 28 | 12 | 30 | 12 |
| Dr 9-30 | 0 | 9 | 28 | 13 | 24 | 0 |
| Dr 10-13 | 0 | 0 | 40 | 18 | 40 | 23 |
| Dr 10-15 | 0 | 0 | 24 | 0 | 30 | 9 |
| Dr 10-16 | 0 | 0 | 18 | 15 | 28 | 9 |
| Dr 11-7 | 13 | 13 | 40 | 22 | 25 | 9 |
| Dr 11-8 | 0 | 14 | 20 | 12 | 25 | 0 |
| Dr 11-12 | 0 | 0 | 22 | 12 | 26 | 8 |
| Dr 11-13 | 0 | 0 | 42 | 18 | 39 | 22 |
| MR 12 | 0 | 9 | 26 | 12 | 25 | 0 |
| MR 14 | 0 | 10 | 25 | 15 | 25 | 9 |
| MR 58 | 20 | 0 | 26 | 17 | 16 | 11 |
| MR 135 | 20 | 17 | 35 | 25 | 30 | 11 |
| MR 160 | 0 | 8 | 26 | 12 | 28 | 10 |
| MR 166 | 0 | 12 | 28 | 12 | 30 | 10 |
| MR 189 | 0 | 0 | 20 | 10 | 20 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreeva, I.S.; Safatov, A.S.; Morozova, V.V.; Solovyanova, N.A.; Puchkova, L.I.; Buryak, G.A.; Olkin, S.E.; Reznikova, I.K.; Emelyanova, E.K.; Okhlopkova, O.V.; et al. Composition and Concentration of the Biogenic Components of the Aerosols Collected over Vasyugan Marshes and Karakan Pine Forest at Altitudes from 500 to 7000 m. Atmosphere 2023, 14, 301. https://doi.org/10.3390/atmos14020301
Andreeva IS, Safatov AS, Morozova VV, Solovyanova NA, Puchkova LI, Buryak GA, Olkin SE, Reznikova IK, Emelyanova EK, Okhlopkova OV, et al. Composition and Concentration of the Biogenic Components of the Aerosols Collected over Vasyugan Marshes and Karakan Pine Forest at Altitudes from 500 to 7000 m. Atmosphere. 2023; 14(2):301. https://doi.org/10.3390/atmos14020301
Chicago/Turabian StyleAndreeva, Irina S., Aleksandr S. Safatov, Vera V. Morozova, Nadezhda A. Solovyanova, Larisa I. Puchkova, Galina A. Buryak, Sergei E. Olkin, Irina K. Reznikova, Elena K. Emelyanova, Olesya V. Okhlopkova, and et al. 2023. "Composition and Concentration of the Biogenic Components of the Aerosols Collected over Vasyugan Marshes and Karakan Pine Forest at Altitudes from 500 to 7000 m" Atmosphere 14, no. 2: 301. https://doi.org/10.3390/atmos14020301
APA StyleAndreeva, I. S., Safatov, A. S., Morozova, V. V., Solovyanova, N. A., Puchkova, L. I., Buryak, G. A., Olkin, S. E., Reznikova, I. K., Emelyanova, E. K., Okhlopkova, O. V., Simonenkov, D. V., & Belan, B. D. (2023). Composition and Concentration of the Biogenic Components of the Aerosols Collected over Vasyugan Marshes and Karakan Pine Forest at Altitudes from 500 to 7000 m. Atmosphere, 14(2), 301. https://doi.org/10.3390/atmos14020301