Potential Respiratory Deposition and Species Composition of Airborne Culturable, Viable, and Non-Viable Fungi during Occupancy in a Pig Farm


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Location
2.2. Temperature and Humidity and Gravimetric Measurements
2.3. Sampling of Culturable Fungi
2.4. Species Identification by MALDI-TOF MS
2.5. Sampling of Airborne Fungi with ACI-8 and Treatment of Filters
2.6. DNA Extraction and Next Generation Sequencing
2.7. Bioinformatics
2.8. Statistical Analyses
- Dg
- is the geometrical mean diameter of fungal aerosols,
- D1
- is the geometrical midpoint of the first interval,
- n1
- is the measured number of particles in the interval, and
- N
- is the total number of particles summed over each interval.
3. Results
3.1. Humidity, Temperature, and Concentration of Airborne Dust
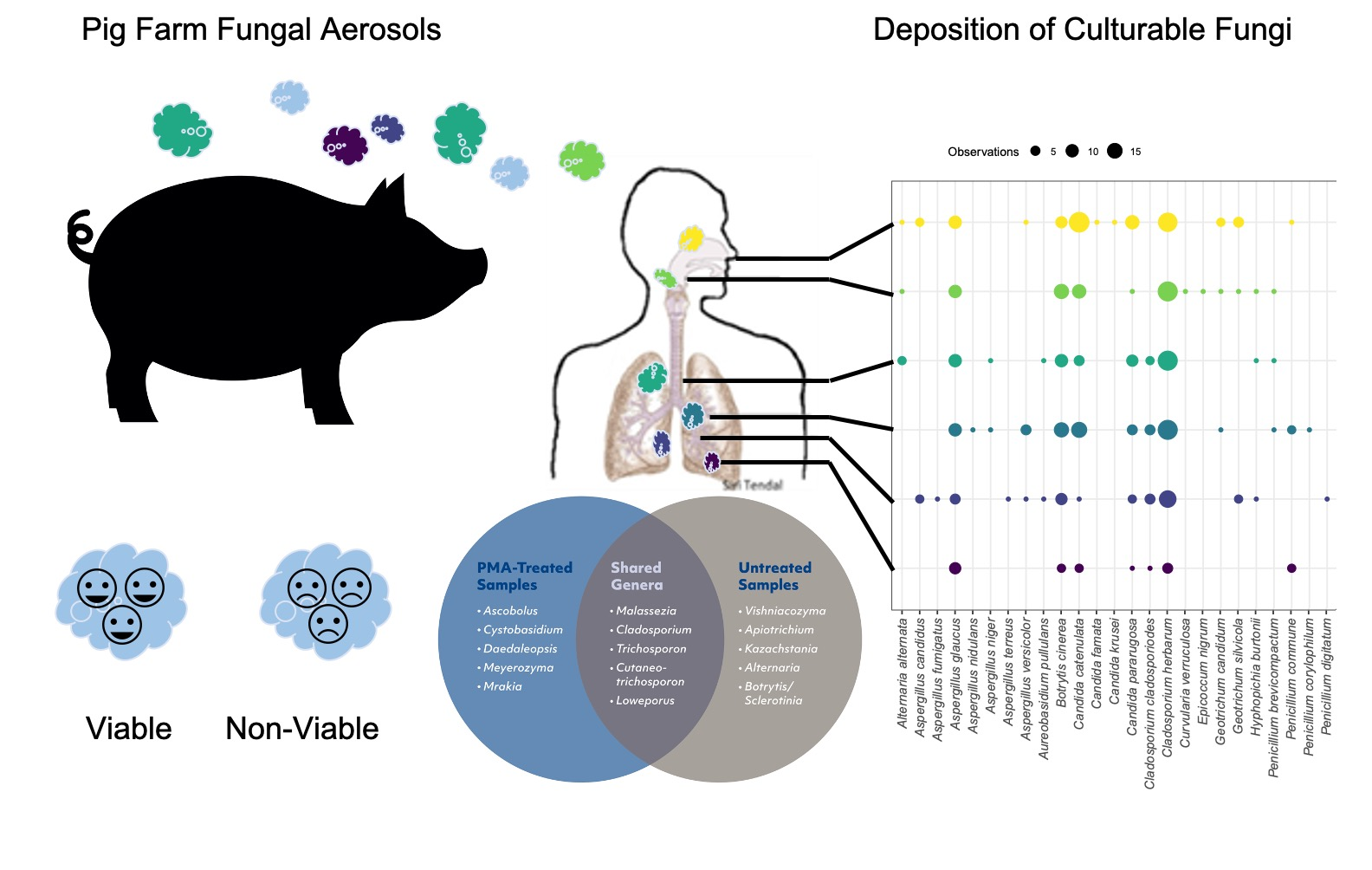
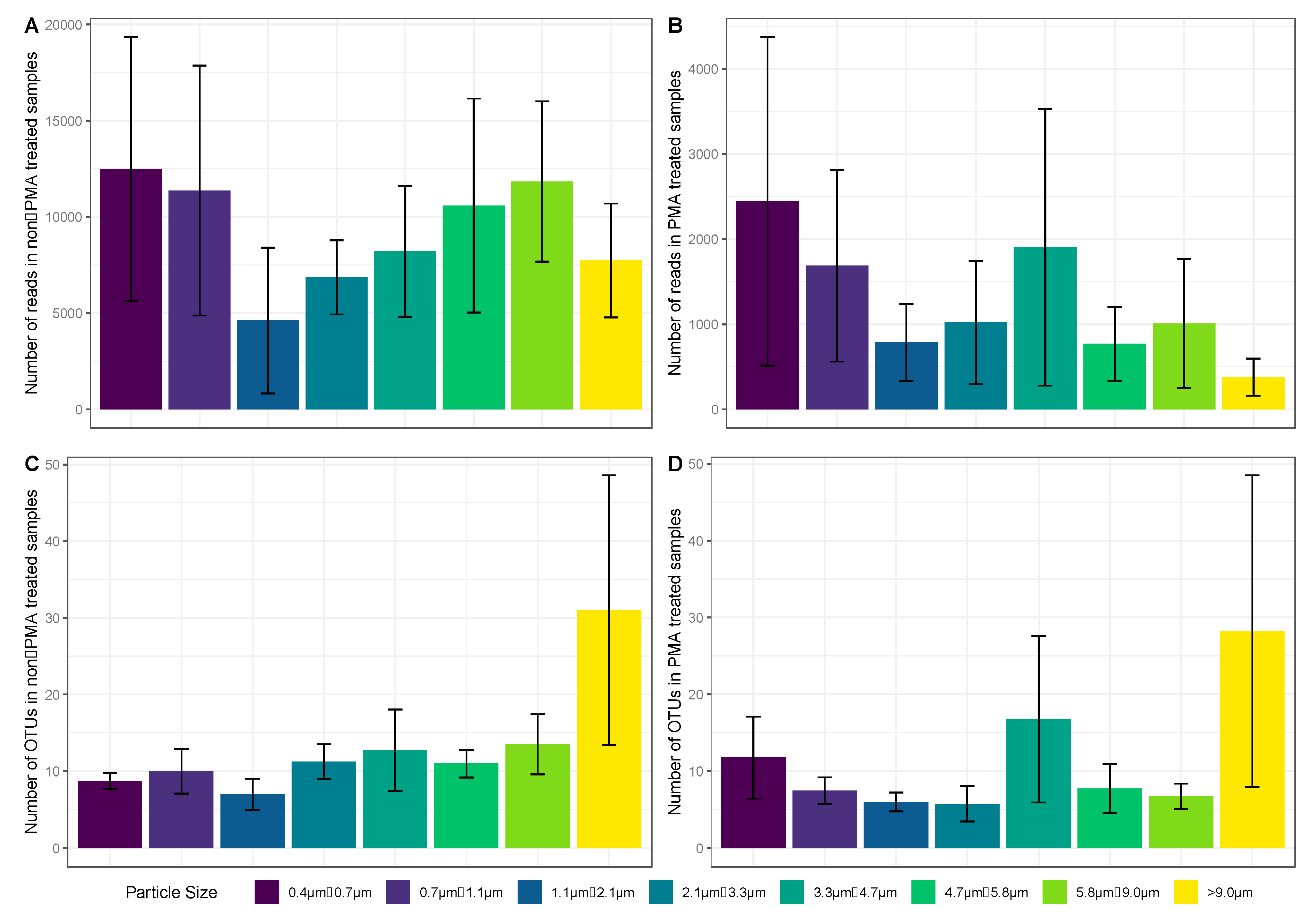
3.2. Concentration of Culturable Fungi and Aerodynamic Diameter
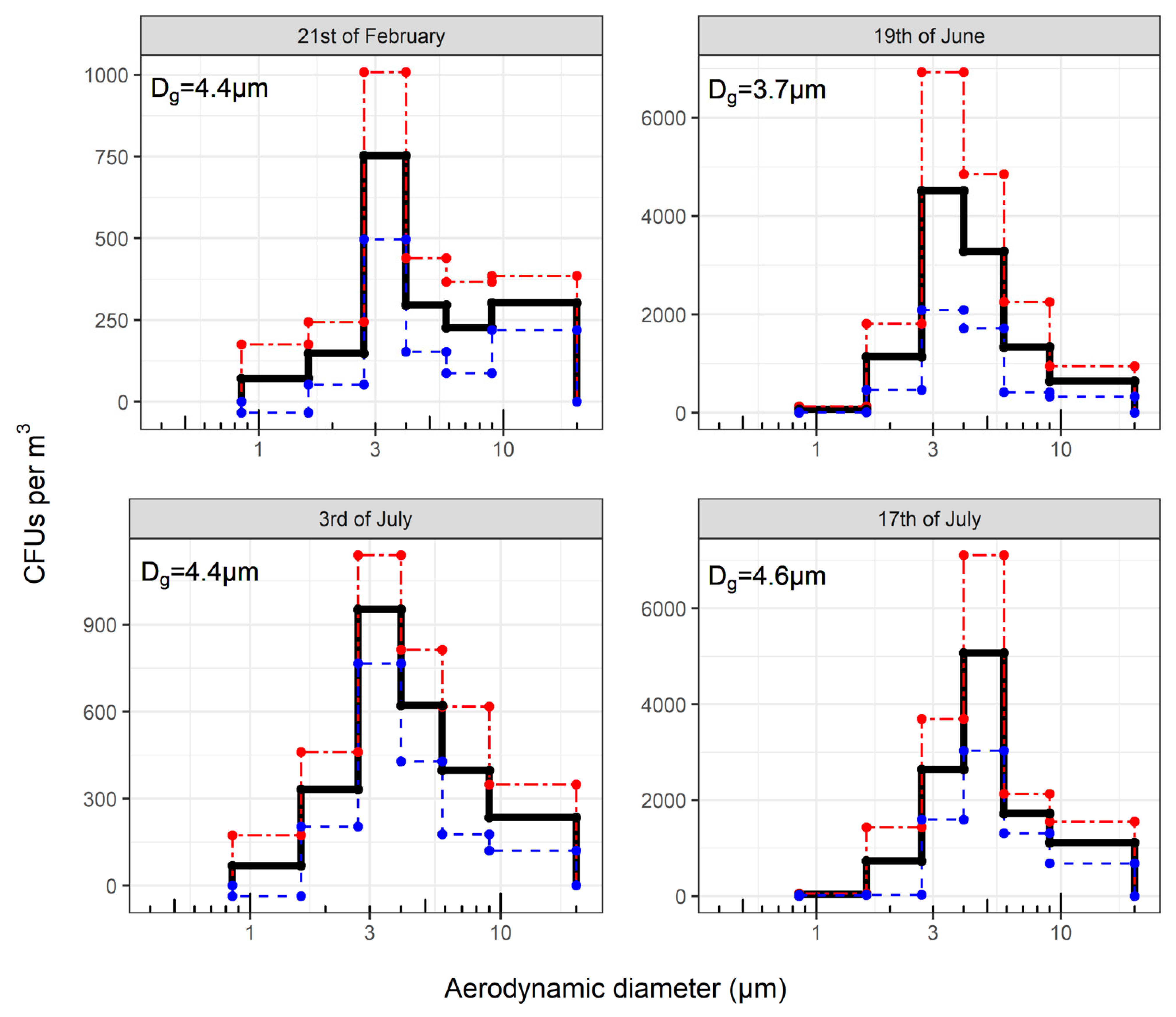
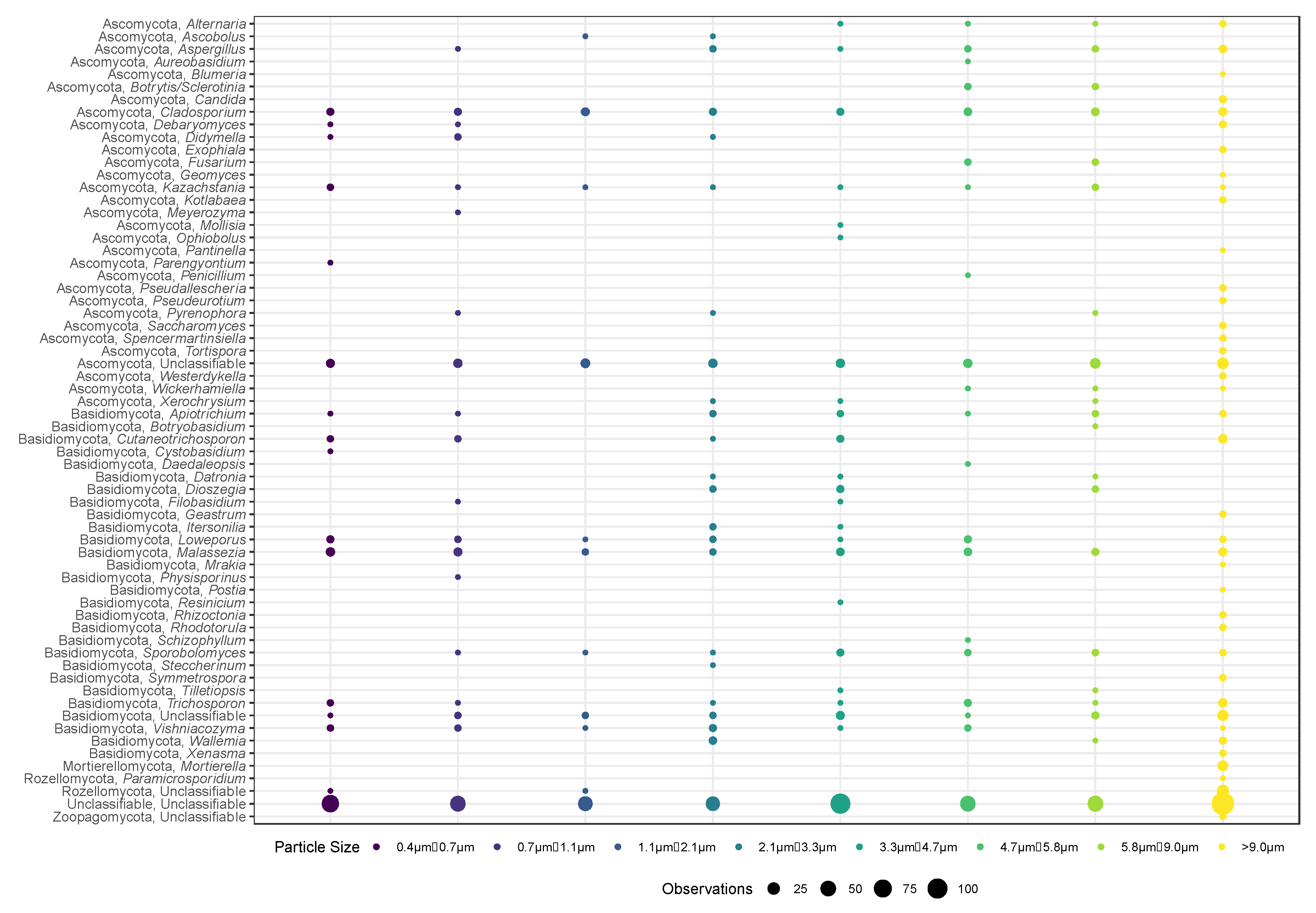
3.3. Species Identified in Samples Taken with ACI-6
3.4. Sequencing after Sampling with ACI-8
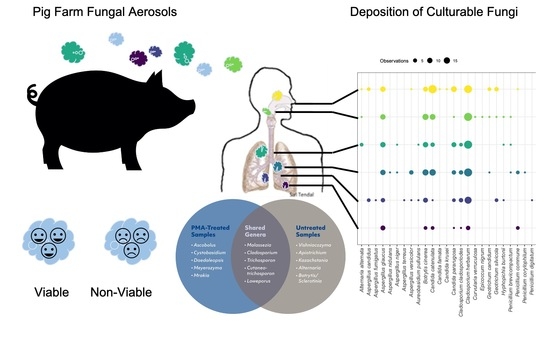
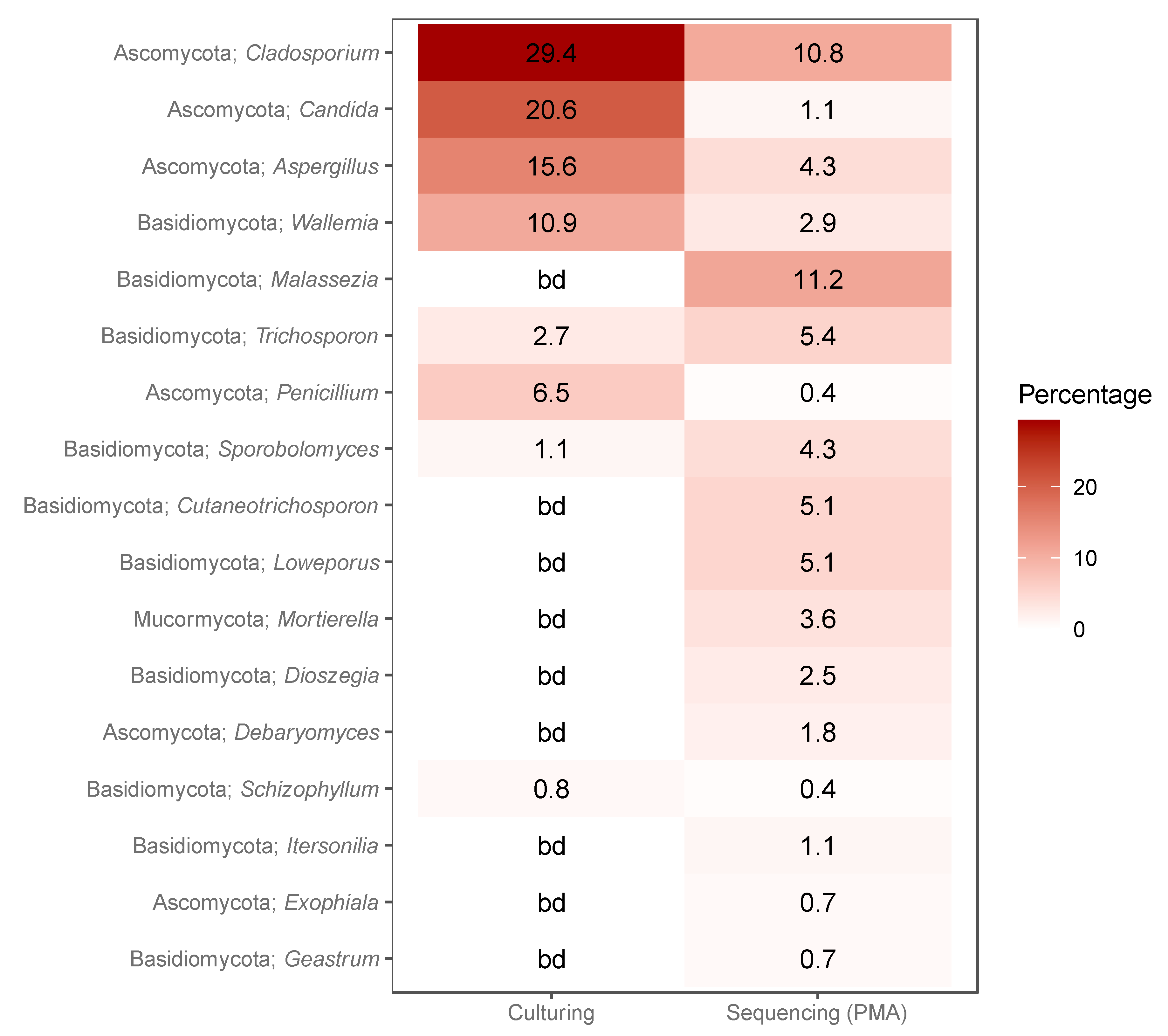
3.5. Concordance between Cultured and Sequenced Fungi
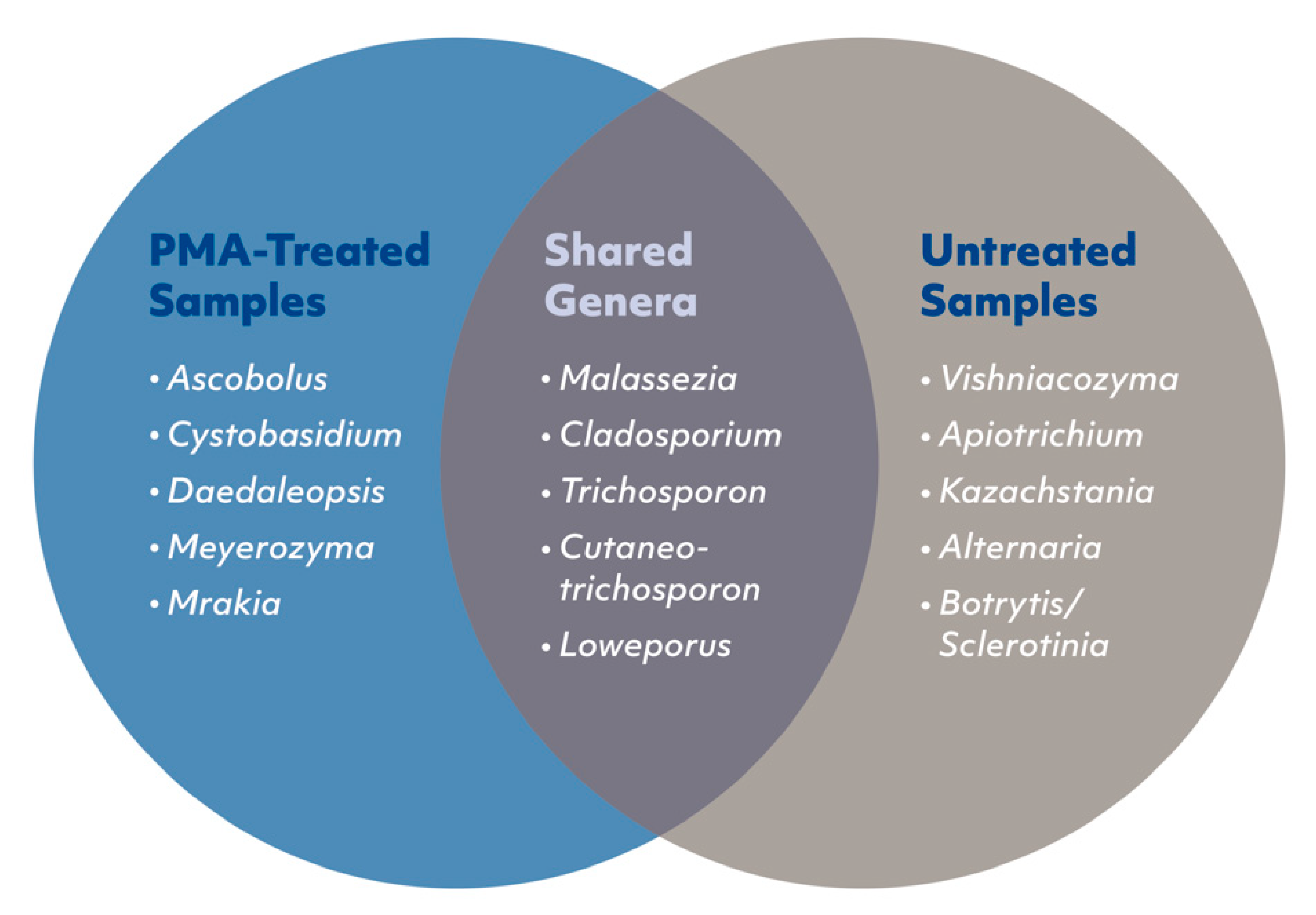
3.6. Viable and Non-Viable Fungi
3.7. Statistical Comparisons between Diversity Observed on Different Sampling Days and Amongst Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vogelzang, P.F.J.; Van Der Gulden, J.W.J.; Schayck, C.P.; va Folgering, H.; Heederik, D.; Tielen, M.J.M. Longitudinal changes in bronchial responsiveness associated with swine confinement dust exposure. Chest 2000, 117, 1488–1495. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cambra-López, M.; Aarnink, A.J.A.; Zhao, Y.; Calvet, S.; Torres, A.G. Airborne particulate matter from livestock production systems: A review of an air pollution problem. Environ. Pollut. 2010, 158, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Baxi, S.N.; Portnoy, J.M.; Larenas-Linnemann, D.; Phipatanakul, W.; Barnes, C.; Grimes, C.; Horner, W.E.; Kennedy, K.; Levetin, E.; Miller, J.D.; et al. Exposure and Health Effects of Fungi on Humans. J. Allergy Clin. Immunol. Pract. 2016, 4, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Barac, A.; Ong, D.S.Y.; Jovancevic, L.; Peric, A.; Surda, P.; Spiric, V.T.; Rubino, S. Fungi-induced upper and lower respiratory tract allergic diseases: One entity. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Panjabi, C. Allergic bronchopulmonary aspergillosis: A perplexing clinical entity. Allergy Asthma Immunol. Res. 2016, 8, 282–297. [Google Scholar] [CrossRef]
- Stevens, D.A.; Kan, V.L.; Judson, M.A.; Morrison, V.A.; Dummer, S.; Denning, D.W.; Bennett, J.E.; Walsh, T.J.; Patterson, T.F.; Pankey, G.A. Practice Guidelines for Diseases Caused by Aspergillus. Clin. Infect. Dis. 2000, 30, 696–709. [Google Scholar] [CrossRef]
- Xu, Z.; Wei, K.; Wu, Y.; Shen, F.; Chen, Q.; Li, M.; Yao, M. Enhancing Bioaerosol Sampling by Andersen Impactors Using Mineral-Oil-Spread Agar Plate. PLoS ONE 2013, 8, e56896. [Google Scholar] [CrossRef]
- Uhrbrand, K.; Schultz, A.C.; Koivisto, A.J.; Nielsen, U.; Madsen, A.M. Assessment of airborne bacteria and noroviruses in air emission from a new highly-advanced hospital wastewater treatment plant. Water Res. 2017, 112, 110–119. [Google Scholar] [CrossRef]
- Yamamoto, N.; Nazaroff, W.W.; Peccia, J. Assessing the aerodynamic diameters of taxon-specific fungal bioaerosols by quantitative PCR and next-generation DNA sequencing. J. Aerosol Sci. 2014, 78, 1–10. [Google Scholar] [CrossRef]
- Yamamoto, N.; Bibby, K.; Qian, J.; Hospodsky, D.; Rismani-Yazdi, H.; Nazaroff, W.W.; Peccia, J. Particle-size distributions and seasonal diversity of allergenic and pathogenic fungi in outdoor air. ISME J. 2012, 6, 1801–1811. [Google Scholar] [CrossRef]
- Emerson, J.B.; Adams, R.I.; Román, C.M.B.; Brooks, B.; Coil, D.A.; Dahlhausen, K.; Ganz, H.H.; Hartmann, E.M.; Hsu, T.; Justice, N.B.; et al. Schrödinger’s microbes: Tools for distinguishing the living from the dead in microbial ecosystems. Microbiome 2017, 5, 86. [Google Scholar] [CrossRef]
- Nguyen, L.D.N.; Deschaght, P.; Merlin, S.; Loywick, A.; Audebert, C.; Van Daele, S.; Viscogliosi, E.; Vaneechoutte, M.; Delhaes, L. Effects of propidium monoazide (PMA) treatment on mycobiome and bacteriome analysis of cystic fibrosis airways during exacerbation. PLoS ONE 2016, 11, e0168860. [Google Scholar] [CrossRef]
- Templeton, S.P.; Buskirk, A.D.; Law, B.; Green, B.J.; Beezhold, D.H. Role of germination in Murine airway CD8+ T-cell responses to Aspergillus conidia. PLoS ONE 2011, 6, e18777. [Google Scholar] [CrossRef] [PubMed]
- Croston, T.L.; Nayak, A.P.; Lemons, A.R.; Goldsmith, W.T.; Gu, J.K.; Germolec, D.R.; Beezhold, D.H.; Green, B.J. Influence of Aspergillus fumigatus conidia viability on murine pulmonary microRNA and mRNA expression following subchronic inhalation exposure. Clin. Exp. Allergy 2016, 46, 1315–1327. [Google Scholar] [CrossRef] [PubMed]
- Simon-Nobbe, B.; Denk, U.; Pöll, V.; Rid, R.; Breitenbach, M. The Spectrum of Fungal Allergy. Int. Arch. Allergy Immunol. 2008, 145, 58–86. [Google Scholar] [CrossRef]
- Horner, W.E.; Helbling, A.; Salvaggio, J.E.; Lehrer, S.B. Fungal allergens. Clin. Microbiol. Rev. 1995, 8, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Mitakakis, T.Z.; O’Meara, T.J.; Tovey, E.R. The effect of sunlight on allergen release from spores of the fungus Alternaria. Grana 2003, 42, 43–46. [Google Scholar] [CrossRef]
- White, J.K.; Nielsen, J.L.; Madsen, A.M. Microbial species and biodiversity in settling dust within and between pig farms. Environ. Res. 2019, 171, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.M.; Kurdi, I.; Feld, L.; Tendal, K. Airborne MRSA and Total Staphylococcus aureus as Associated With Particles of Different Sizes on Pig Farms. Ann. Work Exp. Health 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.M.; Zervas, A.; Tendal, K.; Nielsen, J.L. Microbial diversity in bioaerosol samples causing ODTS compared to reference bioaerosol samples as measured using Illumina sequencing and MALDI-TOF. Environ. Res. 2015, 140, 255–267. [Google Scholar] [CrossRef]
- Checinska Sielaff, A.; Urbaniak, C.; Mohan, G.B.M.; Stepanov, V.G.; Tran, Q.; Wood, J.M.; Minich, J.; McDonald, D.; Mayer, T.; Knight, R.; et al. Characterization of the total and viable bacterial and fungal communities associated with the International Space Station surfaces. Microbiome 2019, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2014, 22, 5271–5277. [Google Scholar] [CrossRef]
- RSudio Team. RStudio: Integrated Development Environment for R; RSudio Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Okasanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Packgge; R Package Version 2.0-10; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Ausschuss für Biologische Arbeitsstoffe. Technische Regeln für Biologische Arbeitsstoffe: Einstufung von Pilzen in Risikogruppen; BAuA: Dortmund, Germany, 2016. [Google Scholar]
- Sowiak, M.; Bródka, K.; Buczyńska, A.; Cyprowski, M.; Kozajda, A.; Sobala, W.; Szadkowska-Stańczyk, I. An assessment of potential exposure to bioaerosols among swine farm workers with particular reference to airborne microorganisms in the respirable fraction under various breeding conditions. Aerobiologia (Bologna) 2011, 28, 121–133. [Google Scholar] [CrossRef]
- Chang, C.W.; Chung, H.; Huang, C.F.; Su, H.J.J. Exposure of workers to airborne microorganisms in open-air swine houses. Appl. Environ. Microbiol. 2001, 67, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.M.; Markouch, A.; Frederiksen, M.W.; Tendal, K. Measurement of dust-borne MRSA in pig farms using different approaches. J. Appl. Microbiol. 2019, 126, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Heederik, D.; Brouwer, R.; Biersteker, K.; Boleij, J.S.M. Relationship of airborne endotoxin and bacteria levels in pig farms with the lung function and respiratory symptoms of farmers. Int. Arch. Occup. Environ. Health 1991, 62, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Fødevarestyrelsen. Bekendtgørelse af Lov om Indendørs Hold af Smågrise, Avls-og Slagtesvin; Retsinformation: Copenhagen, Denmark, 2017. [Google Scholar]
- Prostański, D. Use of Air-and-Water Spraying Systems for Improving Dust Control in Mines. J. Sustain. Min. 2013, 12, 29–34. [Google Scholar] [CrossRef]
- Pedersen, S.; Nonnenmann, M.; Rautiainen, R.; Demmers, T.G.M.; Banhazi, T.; Lyngbye, M. Dust in pig buildings. J. Agric. Saf. Health 2000, 6, 261–274. [Google Scholar] [CrossRef]
- Gustafsson, G. Factors affecting the release and concentration of dust in pig houses. J. Agric. Eng. Res. 1999, 74, 379–390. [Google Scholar] [CrossRef]
- Kousha, M.; Tadi, R.; Soubani, A.O. Pulmonary aspergillosis: A clinical review. Eur. Respir. Rev. 2011, 20, 156–174. [Google Scholar] [CrossRef] [PubMed]
- Arfken, A.M.; Frey, J.F.; Ramsay, T.G.; Summers, K.L. Yeasts of Burden: Exploring the Mycobiome–Bacteriome of the Piglet GI Tract. Front. Microbiol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Porter, T.M.; Golding, G.B. Factors that affect large subunit ribosomal DNA amplicon sequencing studies of fungal communities: Classification method, primer choice, and error. PLoS ONE 2012, 7, e35749. [Google Scholar] [CrossRef]
- Kaneko, T.; Makimura, K.; Abe, M.; Shiota, R.; Nakamura, Y.; Kano, R.; Hasegawa, A.; Sugita, T.; Shibuya, S.; Watanabe, S.; et al. Revised culture-based system for identification of Malassezia species. J. Clin. Microbiol. 2007, 45, 3737–3742. [Google Scholar] [CrossRef]
- Todd, R.T.; Forche, A.; Selmecki, A. Ploidy Variation in Fungi: Polyploidy, Aneuploidy, and Genome Evolution. Microbiol. Spectr. 2017, 5, 139–148. [Google Scholar] [CrossRef]
- Lofgren, L.A.; Uehling, J.K.; Branco, S.; Bruns, T.D.; Martin, F.; Kennedy, P.G. Genome-based estimates of fungal rDNA copy number variation across phylogenetic scales and ecological lifestyles. Mol. Ecol. 2019, 28, 721–730. [Google Scholar] [CrossRef]
- Yan, D.; Zhang, T.; Su, J.; Zhao, L.L.; Wang, H.; Fang, X.M.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and composition of airborne fungal community associated with particulate matters in Beijing during haze and non-haze days. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Karlsson, E.; Johansson, A.M.; Ahlinder, J.; Lundkvist, M.J.; Singh, N.J.; Brodin, T.; Forsman, M.; Stenberg, P. Airborne microbial biodiversity and seasonality in Northern and Southern Sweden. PeerJ 2020, 2020, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Lin, Y.C. Sampling Performance of Impactors for Bacterial Bioaerosols. Aerosol Sci. Technol. 1999, 30, 280–287. [Google Scholar] [CrossRef]
- Grinshpun, S.A.; Willeke, K.; Ulevicius, V.; Juozaitis, A.; Terzieva, S.; Donnelly, J.; Stelma, G.N.; Brenner, K.P. Effect of impaction, bounce and reaerosolization on the collection efficiency of impingers. Aerosol Sci. Technol. 1997, 26, 326–342. [Google Scholar] [CrossRef]
- Mainelis, G.; Tabayoyong, M. The effect of sampling time on the overall performance of portable microbial impactors. Aerosol Sci. Technol. 2010, 44, 75–82. [Google Scholar] [CrossRef]
- Urubschurov, V.; Janczyk, P.; Souffrant, W.B.; Freyer, G.; Zeyner, A. Establishment of intestinal microbiota with focus on yeasts of unweaned and weaned piglets kept under different farm conditions. FEMS Microbiol. Ecol. 2011, 77, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Ramayo-Caldas, Y.; Prenafeta, F.; Zingaretti, L.M.; Gonzales, O.; Dalmau, A.; Quintanilla, R.; Ballester, M. Gut eukaryotic communities in pigs: Diversity, composition and host genetics contribution. bioRxiv 2020, 2. [Google Scholar] [CrossRef]
- Nocker, A.; Richter-Heitmann, T.; Montijn, R.; Schuren, F.; Kort, R. Discrimination between live and dead cells in bacterial communities from environmental water samples analyzed by 454 pyrosequencing. Int. Microbiol. 2010, 13, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Desneux, J.; Chemaly, M.; Pourcher, A.M. Experimental design for the optimization of propidium monoazide treatment to quantify viable and non-viable bacteria in piggery effluents. BMC Microbiol. 2015, 15, 164. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Season | Relative Humidity | Temperature |
|---|---|---|---|
| 21st of February 2019 | Winter | 38.3% ± 0.2 | 12.3 °C ± 0.7 |
| 19th of June 2019 | Summer | 62.9% ± 8.5 * | 24.2 °C ± 0.8 |
| 3rd of July 2019 | Summer | 53.9% ± 3.5 | 15.6 °C ± 0.5 |
| 17th of July 2019 | Summer | 69.5% ± 9.8 ** | 19.9 °C ± 1.4 |
| Test | p-Value | R Statistic | ||
|---|---|---|---|---|
| Culturing | Day | ANOSIM | 0.001 | 0.4005 |
| Size Fraction | ANOSIM | 0.001 | 0.06021 | |
| Sampling Round | ANOSIM | 0.546 | −0.003511 | |
| Sequencing | Day | ANOSIM | 0.001 | 0.2449 |
| Size Fraction | ANOSIM | 0.953 | −0.03387 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, J.K.; Nielsen, J.L.; Madsen, A.M. Potential Respiratory Deposition and Species Composition of Airborne Culturable, Viable, and Non-Viable Fungi during Occupancy in a Pig Farm. Atmosphere 2020, 11, 639. https://doi.org/10.3390/atmos11060639
White JK, Nielsen JL, Madsen AM. Potential Respiratory Deposition and Species Composition of Airborne Culturable, Viable, and Non-Viable Fungi during Occupancy in a Pig Farm. Atmosphere. 2020; 11(6):639. https://doi.org/10.3390/atmos11060639
Chicago/Turabian StyleWhite, John Kerr, Jeppe Lund Nielsen, and Anne Mette Madsen. 2020. "Potential Respiratory Deposition and Species Composition of Airborne Culturable, Viable, and Non-Viable Fungi during Occupancy in a Pig Farm" Atmosphere 11, no. 6: 639. https://doi.org/10.3390/atmos11060639
APA StyleWhite, J. K., Nielsen, J. L., & Madsen, A. M. (2020). Potential Respiratory Deposition and Species Composition of Airborne Culturable, Viable, and Non-Viable Fungi during Occupancy in a Pig Farm. Atmosphere, 11(6), 639. https://doi.org/10.3390/atmos11060639