Genome Instability Induced by Low Levels of Replicative DNA Polymerases in Yeast


Abstract
1. Introduction
2. Analysis of Genome Instability in Yeast
2.1. Commonly Used Assays of Genome Instability
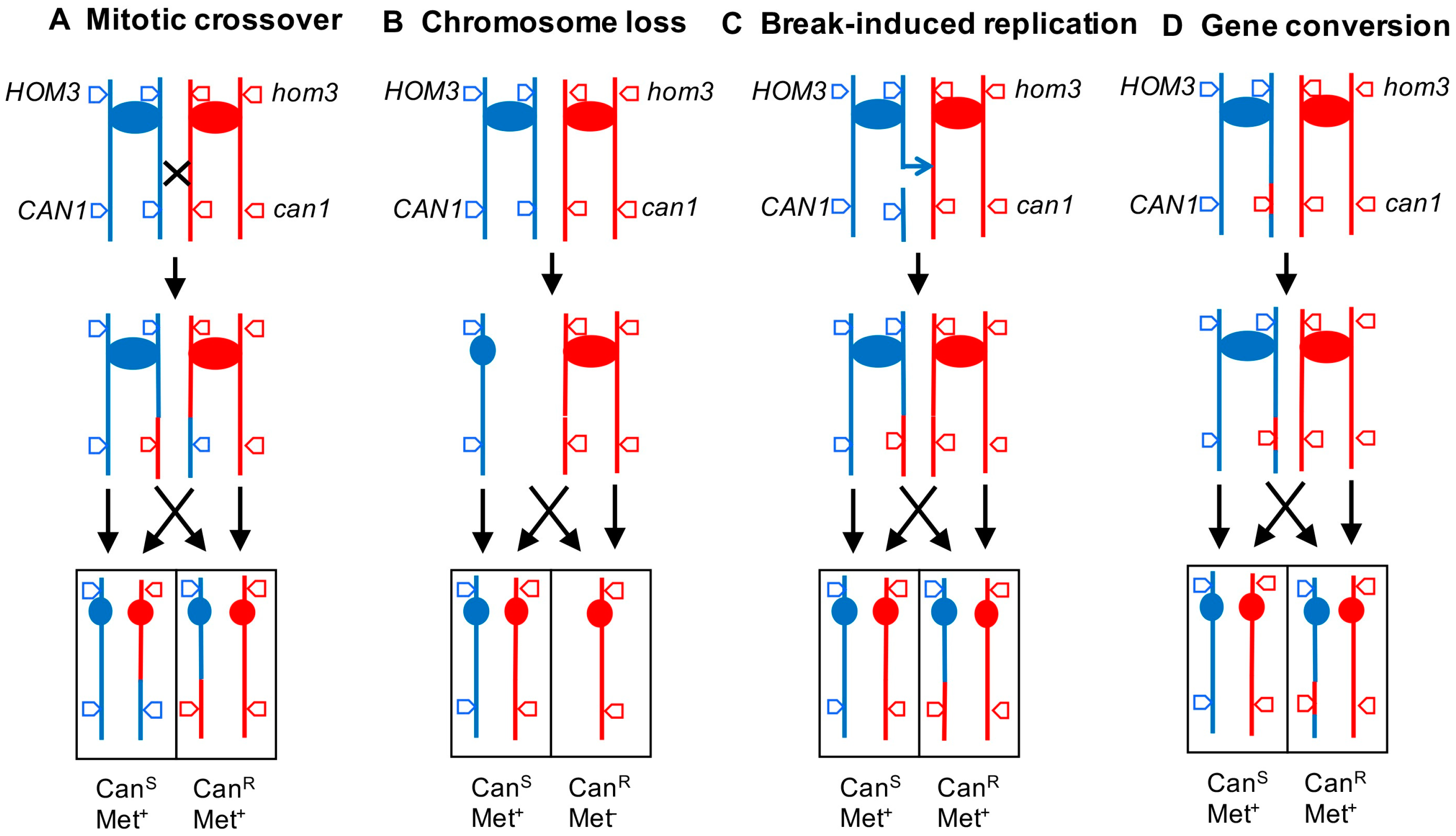
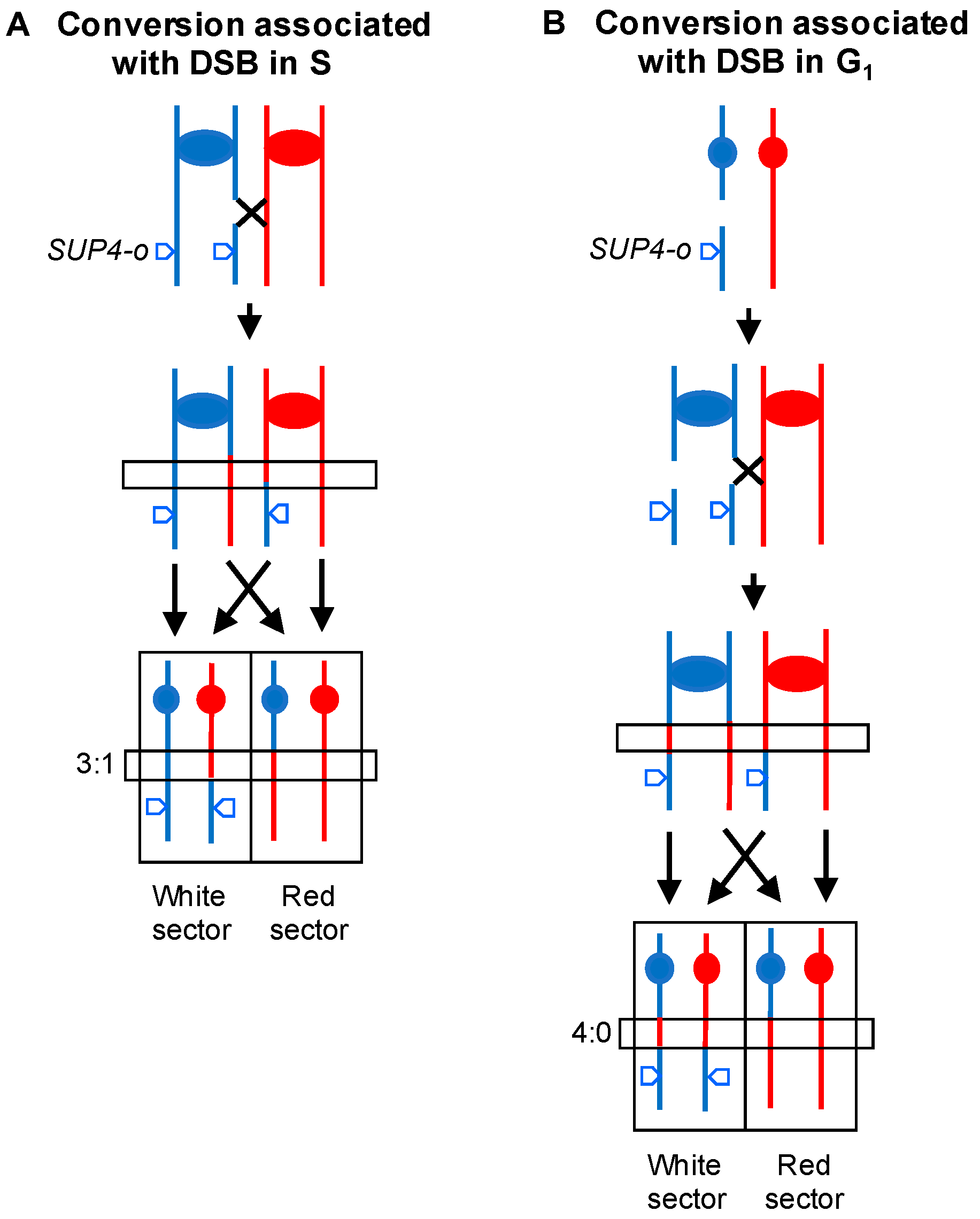
2.2. Analysis of Mitotic Crossovers in Yeast Using Microarrays
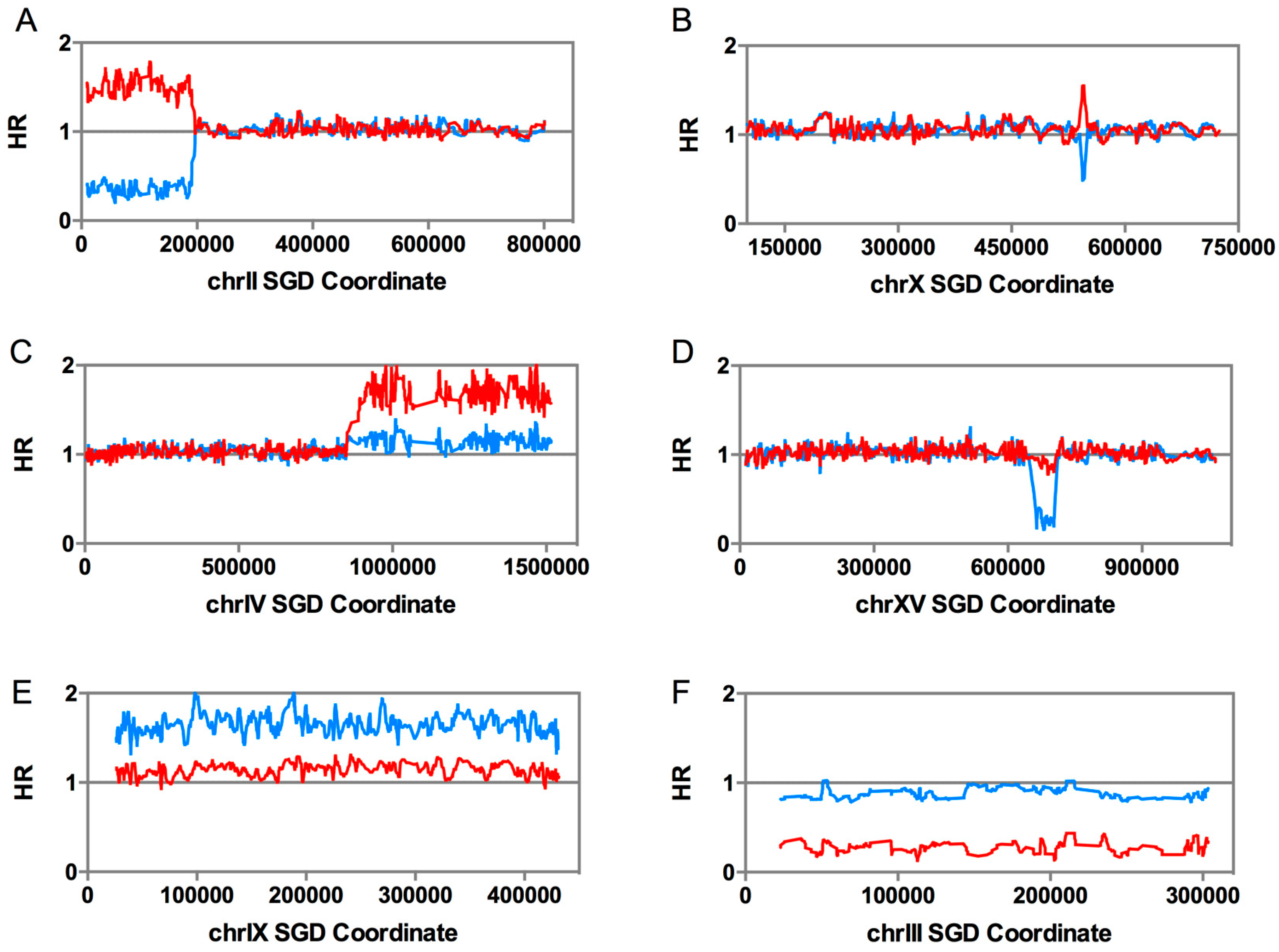
2.3. Diagnosis of Other Genomic Alterations Using Microarrays
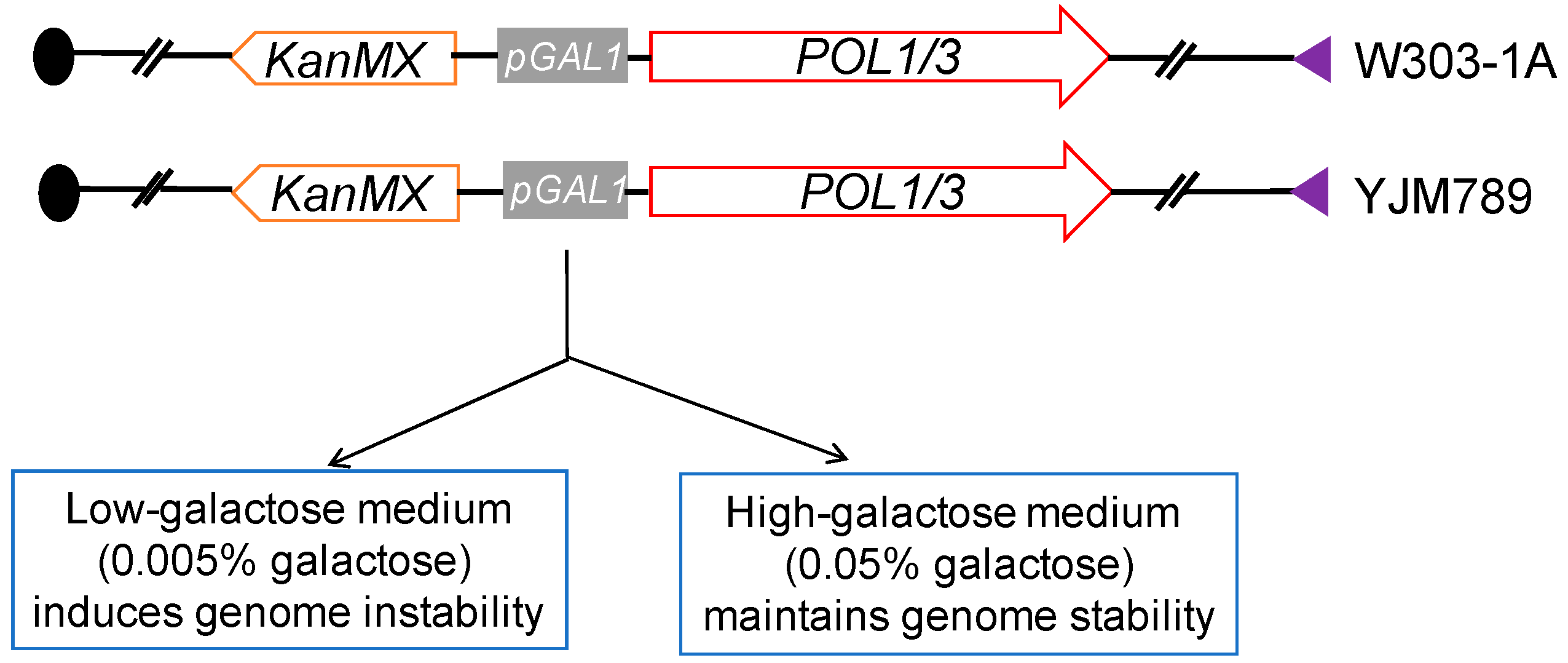
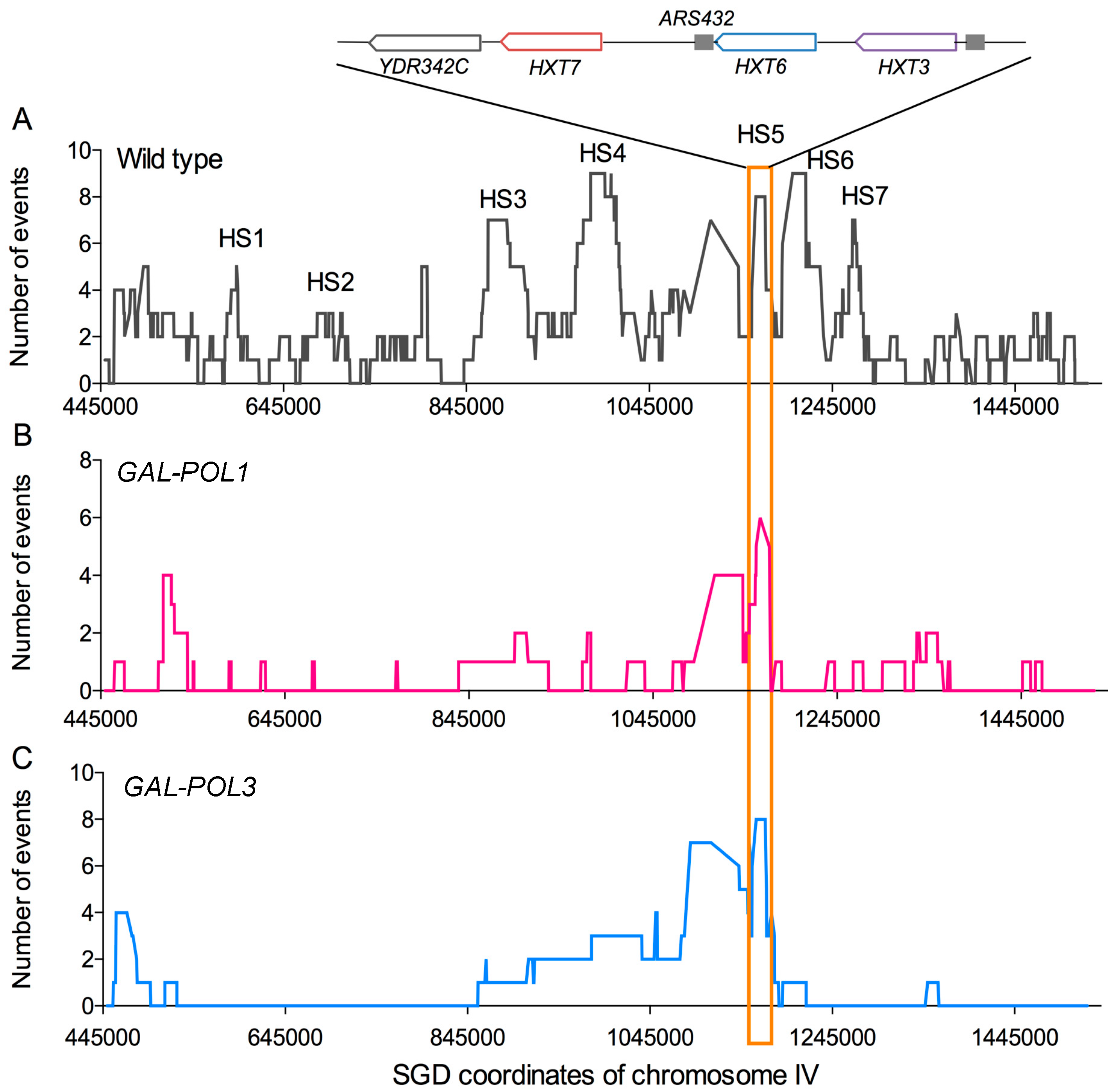
2.4. Systems for Inducing DNA Replication Stress by Depleting DNA Polymerases
3. Genomic Instability in Wild-Type Strains and in Strains with Low Levels of Replicative DNA Polymerases
3.1. Genetic Instability in Wild-Type Diploids
3.2. Genetic Instability in Strains with Low Levels of Polα
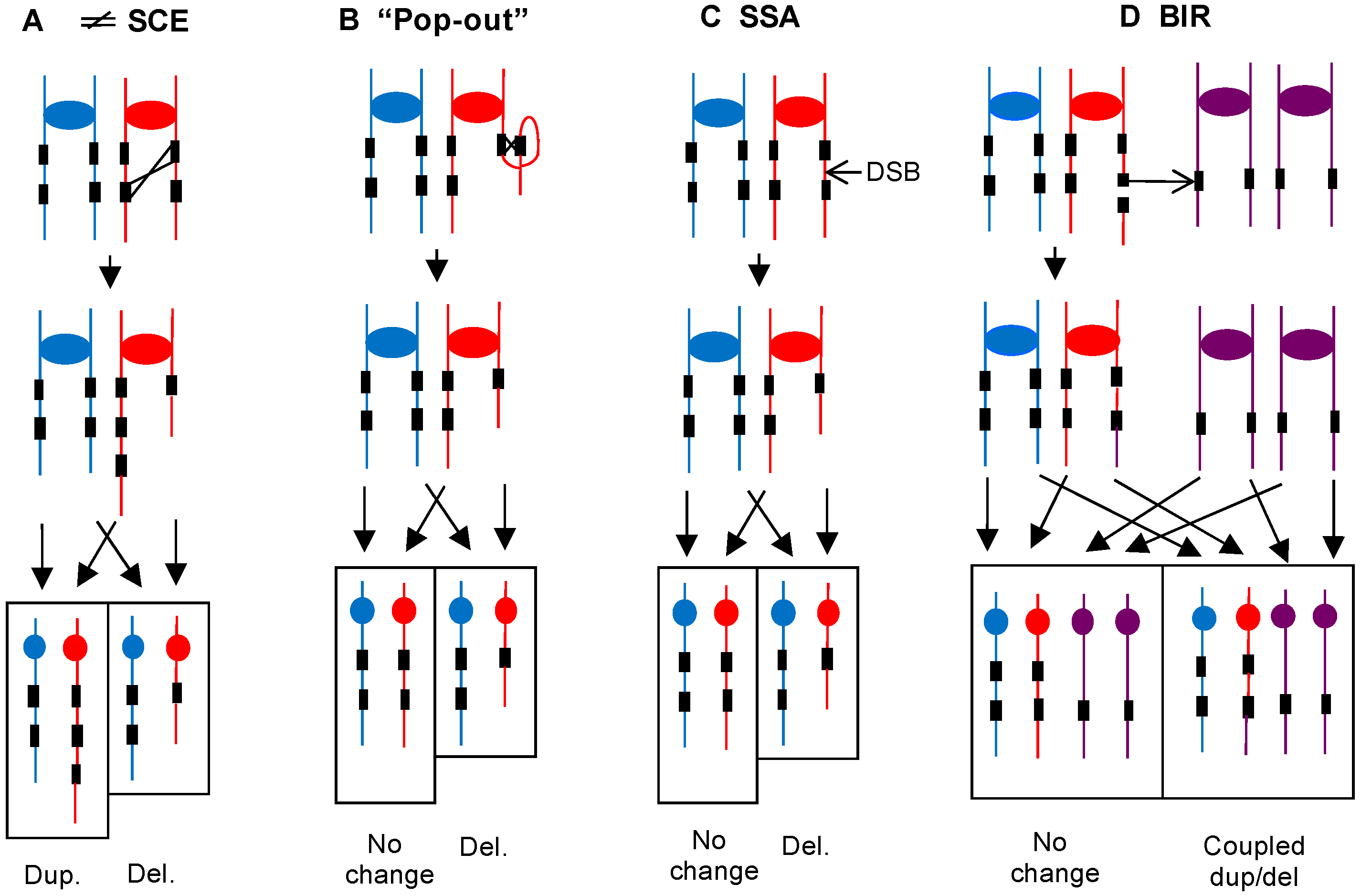
3.2.1. Loss of Heterozygosity Events Resulting from Mitotic Recombination between Homologs
3.2.2. Large (>5 kb) Deletions and Duplications
3.2.3. Rate of Aneuploidy
3.2.4. Rate of Base Substitutions and Small Insertions/Deletions
3.3. Genetic Instability in Strains with Low Levels of Polδ
3.3.1. Loss of Heterozygosity Events Resulting from Mitotic Recombination between Homologs
3.3.2. Large (>5 kb) Deletions and Duplications
3.3.3. Rate of Aneuploidy
3.3.4. Rate of Base Substitutions and Small Insertions/Deletions
3.3.5. Evolution of the Genome under Conditions of Low Polδ
3.4. Comparison of Genomic Instability Induced by Low Levels of Polα or Polδ
4. Relevance of Yeast Studies to Fragile Sites and Chromosome Alterations in Tumors
5. Summary
Funding
Acknowledgments
Conflicts of Interest
References
- Burgers, P.M.; Kunkel, T.A. Eukaryotic DNA replication fork. Annu. Rev. Biochem. 2017, 86, 417–438. [Google Scholar] [CrossRef] [PubMed]
- Lujan, S.A.; Williams, J.S.; Kunkel, T.A. DNA polymerases divide the labor of genome replication. Trends Cell Biol. 2016, 26, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Mazouzi, A.; Velimezi, G.; Loizou, J.I. DNA replication stress: Causes, resolution and disease. Exp. Cell Res. 2014, 329, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, S.; Méndez, J. DNA replication stress: From molecular mechanisms to human disease. Chromosoma 2017, 126, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Macheret, M.; Halazonetis, T.D. DNA replication stress as a hallmark of cancer. Annu. Rev. Pathol. 2015, 10, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.M.; Lindsay, H.D. DNA replication stress and cancer: Cause or cure? Future Oncol. 2016, 12, 221–237. [Google Scholar] [CrossRef] [PubMed]
- Glover, T.W.; Wilson, T.E.; Arlt, M.F. Fragile sites in cancer: More than meets the eye. Nat. Rev. Cancer 2017, 17, 489. [Google Scholar] [CrossRef] [PubMed]
- Neil, A.J.; Kim, J.C.; Mirkin, S.M. Precarious maintenance of simple DNA repeats in eukaryotes. Bioessays 2017, 39, 1700077. [Google Scholar] [CrossRef] [PubMed]
- Freudenreich, C.H.; Kantrow, S.M.; Zakian, V.A. Expansion and length-dependent fragility of CTG repeats in yeast. Science 1998, 279, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Narayanan, V.; Mieczkowski, P.A.; Petes, T.D.; Krasilnikova, M.M.; Mirkin, S.M.; Lobachev, K.S. Chromosome fragility at GAA tracts in yeast depends on repeat orientation and requires mismatch repair. EMBO J. 2008, 27, 2896–2906. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Dominska, M.; Greenwell, P.W.; Harvanek, Z.; Lobachev, K.S.; Kim, H.-M.; Narayanan, V.; Mirkin, S.M.; Petes, T.D. Friedreich’s Ataxia (GAA)n•(TTC)n repeats strongly stimulate mitotic crossovers in Saccharomyces cerevisiae. PLoS Genet. 2011, 7, e1001270. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Dominska, M.; Greenwell, P.W.; Petes, T.D. Genome-wide high-resolution mapping of chromosome fragile sites in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2014, 111, E2210–E2218. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.Q.; Zhang, K.; Wu, X.C.; Mieczkowski, P.A.; Petes, T.D. Global analysis of genomic instability caused by DNA replication stress in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2016, 113, E8114–E8121. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.-E.; Rio, A.-G.; Nicolas, A.; Kolodner, R.D. A genomewide screen in Saccharomyces cerevisiae for genes that suppress the accumulation of mutations. Proc. Natl. Acad. Sci. USA 2003, 100, 11529–11534. [Google Scholar] [CrossRef] [PubMed]
- Lea, D.E.; Coulson, C.A. The distribution of the numbers of mutants in bacterial populations. J. Genet. 1949, 49, 264–285. [Google Scholar] [CrossRef] [PubMed]
- Boeke, J.D.; La Croute, F.; Fink, G.R. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol. Gen. Genet. 1984, 197, 345–346. [Google Scholar] [CrossRef] [PubMed]
- Kunz, B.A.; Ramachandran, K.; Vonarx, E.J. DNA sequence analysis of spontaneous mutagenesis in Saccharomyces cerevisiae. Genetics 1998, 148, 1491–1505. [Google Scholar] [PubMed]
- Kiktev, D.A.; Sheng, Z.; Lobachev, K.S.; Petes, T.D. GC content elevates mutation and recombination rates in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2018, 115, E7109–E7118. [Google Scholar] [CrossRef] [PubMed]
- Nishant, K.; Wei, W.; Mancera, E.; Argueso, J.L.; Schlattl, A.; Delhomme, N.; Ma, X.; Bustamante, C.D.; Korbel, J.O.; Gu, Z. The baker’s yeast diploid genome is remarkably stable in vegetative growth and meiosis. PLoS Genet. 2010, 6, e1001109. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.O.; Siegal, M.L.; Hall, D.W.; Petrov, D.A. Precise estimates of mutation rate and spectrum in yeast. Proc. Natl. Acad. Sci. USA 2014, 111, E2310–E2318. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H.; Smith, D. Altered fidelity of mitotic chromosome transmission in cell cycle mutants of S. cerevisiae. Genetics 1985, 110, 381–395. [Google Scholar] [PubMed]
- Symington, L.S.; Rothstein, R.; Lisby, M. Mechanisms and regulation of mitotic recombination in Saccharomyces cerevisiae. Genetics 2014, 198, 795–835. [Google Scholar] [CrossRef] [PubMed]
- St Charles, J.; Petes, T.D. High-resolution mapping of spontaneous mitotic recombination hotspots on the 1.1 Mb arm of yeast chromosome IV. PLoS Genet. 2013, 9, e1003434. [Google Scholar] [CrossRef]
- Yim, E.; O’Connell, K.E.; Charles, J.S.; Petes, T.D. High-resolution mapping of two types of spontaneous mitotic gene conversion events in Saccharomyces cerevisiae. Genetics 2014, 198, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Kolodner, R.D. Gross chromosomal rearrangements in Saccharomyces cerevisiae replication and recombination defective mutants. Nat. Genet. 1999, 23, 81. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.S.; Hazkani-Covo, E.; Yin, Y.; Andersen, S.L.; Dietrich, F.S.; Greenwell, P.W.; Malc, E.; Mieczkowski, P.; Petes, T.D. High-resolution genome-wide analysis of irradiated (UV and γ rays) diploid yeast cells reveals a high frequency of genomic loss of heterozygosity (LOH) events. Genetics 2012, 190, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Gresham, D.; Curry, B.; Ward, A.; Gordon, D.B.; Brizuela, L.; Kruglyak, L.; Botstein, D. Optimized detection of sequence variation in heterozygous genomes using DNA microarrays with isothermal-melting probes. Proc. Natl. Acad. Sci. USA 2010, 107, 1482–1487. [Google Scholar] [CrossRef] [PubMed]
- Mancera, E.; Bourgon, R.; Brozzi, A.; Huber, W.; Steinmetz, L.M. High-resolution mapping of meiotic crossovers and non-crossovers in yeast. Nature 2008, 454, 479. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.; Greenwell, P.W.; Dominska, M.; Gawel, M.; Hamilton, M.; Petes, T.D. A fine-structure map of spontaneous mitotic crossovers in the yeast Saccharomyces cerevisiae. PLoS Genet. 2009, 5, e1000410. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.J.; Degtyareva, N.P.; Lobachev, K.; Petes, T.D. Chromosomal translocations in yeast induced by low levels of DNA polymerase a model for chromosome fragile sites. Cell 2005, 120, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Kokoska, R.J.; Stefanovic, L.; DeMai, J.; Petes, T.D. Increased rates of genomic deletions generated by mutations in the yeast gene encoding DNA polymerase δ or by decreases in the cellular levels of DNA polymerase δ. Mol. Cell Biol. 2000, 20, 7490–7504. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.J.; Degtyareva, N.P.; Kokoska, R.J.; Petes, T.D. Reduced levels of DNA polymerase δ induce chromosome fragile site instability in yeast. Mol Cell Biol. 2008, 28, 5359–5368. [Google Scholar] [CrossRef] [PubMed]
- Strathern, J.; Hicks, J.; Herskowitz, I. Control of cell type in yeast by the mating type locus: The α1-α2 hypothesis. J. Mol. Biol. 1981, 147, 357–372. [Google Scholar] [CrossRef]
- O’Connell, K.; Jinks-Robertson, S.; Petes, T.D. Elevated genome-wide instability in yeast mutants lacking RNase H activity. Genetics 2015, 201, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Azvolinsky, A.; Giresi, P.G.; Lieb, J.D.; Zakian, V.A. Highly transcribed RNA polymerase II genes are impediments to replication fork progression in Saccharomyces cerevisiae. Mol. Cell 2009, 34, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.M.; Sokolsky, T.; Tucker, C.M.; Chan, L.Y.; Boselli, M.; Dunham, M.J.; Amon, A. Effects of aneuploidy on cellular physiology and cell division in haploid yeast. Science 2007, 317, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L.; Petes, T.D. Reciprocal uniparental disomy in yeast. Proc. Natl. Acad. Sci. USA 2012, 109, 9947–9952. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M. Ribosomal RNA genes, RNA polymerases, nucleolar structures, and synthesis of rRNA in the yeast Saccharomyces cerevisiae. Cold Spring Harb. Symp. Quant. Biol. 2001, 66, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Strope, P.K.; Kozmin, S.G.; McCusker, J.H.; Dietrich, F.S.; Kokoska, R.J.; Petes, T.D. Structures of naturally evolved CUP1 tandem arrays in yeast indicate that these arrays are generated by unequal nonhomologous recombination. G3 2014, 4, 2259–2269. [Google Scholar] [CrossRef] [PubMed]
- Casper, A.M.; Mieczkowski, P.A.; Gawel, M.; Petes, T.D. Low levels of DNA polymerase alpha induce mitotic and meiotic instability in the ribosomal DNA gene cluster of Saccharomyces cerevisiae. PLoS Genet. 2008, 4, e1000105. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Dominska, M.; Petrova, A.; Bagshaw, H.; Kokoska, R.J.; Petes, T.D. Properties of mitotic and meiotic recombination in the tandemly-repeated CUP1 gene cluster in the yeast Saccharomyces cerevisiae. Genetics 2017, 206, 785–800. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.L.; Sloan, R.S.; Petes, T.D.; Jinks-Robertson, S. Genome-destabilizing effects associated with top1 loss or accumulation of top1 cleavage complexes in yeast. PLoS Genet. 2015, 11, e1005098. [Google Scholar] [CrossRef] [PubMed]
- Casper, A.M.; Greenwell, P.W.; Tang, W.; Petes, T.D. Chromosome aberrations resulting from double-strand DNA breaks at a naturally-occurring yeast fragile site composed of inverted Ty elements are independent of Mre11p and Sae2p. Genetics 2009, 183, 423. [Google Scholar] [CrossRef] [PubMed]
- Rosen, D.M.; Younkin, E.M.; Miller, S.D.; Casper, A.M. Fragile site instability in Saccharomyces cerevisiae causes loss of heterozygosity by mitotic crossovers and break-induced replication. PLoS Genet. 2013, 9, e1003817. [Google Scholar] [CrossRef] [PubMed]
- Mieczkowski, P.A.; Lemoine, F.J.; Petes, T.D. Recombination between retrotransposons as a source of chromosome rearrangements in the yeast Saccharomyces cerevisiae. DNA Repair. 2006, 5, 1010–1020. [Google Scholar] [CrossRef] [PubMed]
- Salim, D.; Bradford, W.D.; Freeland, A.; Cady, G.; Wang, J.; Pruitt, S.C.; Gerton, J.L. DNA replication stress restricts ribosomal DNA copy number. PLoS Genet. 2017, 13, e1007006. [Google Scholar] [CrossRef] [PubMed]
- Kwan, E.X.; Foss, E.J.; Tsuchiyama, S.; Alvino, G.M.; Kruglyak, L.; Kaeberlein, M.; Raghuraman, M.; Brewer, B.J.; Kennedy, B.K.; Bedalov, A. A natural polymorphism in rDNA replication origins links origin activation with calorie restriction and lifespan. PLoS Genet. 2013, 9, e1003329. [Google Scholar] [CrossRef] [PubMed]
- Foss, E.J.; Lao, U.; Dalrymple, E.; Adrianse, R.L.; Loe, T.; Bedalov, A. SIR2 suppresses replication gaps and genome instability by balancing replication between repetitive and unique sequences. Proc. Natl. Acad. Sci. USA 2017, 114, 552–557. [Google Scholar] [CrossRef] [PubMed]
- McCulley, J.L.; Petes, T.D. Chromosome rearrangements and aneuploidy in yeast strains lacking both Tel1p and Mec1p reflect deficiencies in two different mechanisms. Proc. Natl. Acad. Sci. USA 2010, 107, 11465–11470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zeidler, A.F.; Song, W.; Puccia, C.M.; Malc, E.; Greenwell, P.W.; Mieczkowski, P.A.; Petes, T.D.; Argueso, J.L. Gene copy number variation (CNV) in haploid and diploid strains of the yeast Saccharomyces cerevisiae. Genetics 2013, 193, 785. [Google Scholar] [CrossRef] [PubMed]
- Her, J.; Bunting, S.F. How cells ensure correct repair of DNA double-strand breaks. J. Biol. Chem. 2018, 293. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zhou, W.; Fu, W.; Du, R.; Jin, L.; Zhang, F. Correlation between frequency of non-allelic homologous recombination and homology properties: Evidence from homology-mediated CNV mutations in the human genome. Hum. Mol. Genet. 2014, 24, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Szilard, R.K.; Jacques, P.-É.; Laramée, L.; Cheng, B.; Galicia, S.; Bataille, A.R.; Yeung, M.; Mendez, M.; Bergeron, M.; Robert, F. Systematic identification of fragile sites via genome-wide location analysis of γ-H2AX. Nat. Struct. Mol. Biol. 2010, 17, 299. [Google Scholar] [CrossRef] [PubMed]
- Makarova, A.V.; Burgers, P.M. Eukaryotic DNA polymerase ζ. DNA Repair. 2015, 29, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Moller, H.D.; Parsons, L.; Jorgensen, T.S.; Botstein, D.; Regenberg, B. Extrachromosomal circular DNA is common in yeast. Proc. Natl. Acad. Sci. USA 2015, 112, E3114–E3122. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Degtyareva, N.P.; Koloteva, N.N.; Sugino, A.; Masumoto, H.; Gordenin, D.A.; Resnick, M.A. Replication slippage between distant short repeats in Saccharomyces cerevisiae depends on the direction of replication and the RAD50 and RAD52 genes. Mol. Cell Biol. 1995, 15, 5607–5617. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sterling, J.; Storici, F.; Resnick, M.A.; Gordenin, D.A. Hypermutability of damaged single-strand DNA formed at double-strand breaks and uncapped telomeres in yeast Saccharomyces cerevisiae. PLoS Genet. 2008, 4, e1000264. [Google Scholar] [CrossRef] [PubMed]
- Northam, M.R.; Robinson, H.A.; Kochenova, O.V.; Shcherbakova, P.V. Participation of DNA polymerase zeta in replication of undamaged DNA in Saccharomyces cerevisiae. Genetics 2010, 184, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, Y.; Frahm, C.; McElhinny, S.N.; Niimi, A.; Suzuki, M.; Kunkel, T. Evidence that errors made by DNA polymerase α are corrected by DNA polymerase δ. Curr. Biol. 2006, 16, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Nethanel, T.; Kaufmann, G. Two DNA polymerases may be required for synthesis of the lagging DNA strand of simian virus 40. J. Virol. 1990, 64, 5912–5918. [Google Scholar] [PubMed]
- Wang, X.; Ira, G.; Tercero, J.A.; Holmes, A.M.; Diffley, J.F.; Haber, J.E. Role of DNA replication proteins in double-strand break-induced recombination in Saccharomyces cerevisiae. Mol. Cell Biol. 2004, 24, 6891–6899. [Google Scholar] [CrossRef] [PubMed]
- Maloisel, L.; Fabre, F.; Gangloff, S. DNA polymerase δ is preferentially recruited during homologous recombination to promote heteroduplex DNA extension. Mol. Cell Biol. 2008, 28, 1373–1382. [Google Scholar] [CrossRef] [PubMed]
- Lengauer, C.; Kinzler, K.W.; Vogelstein, B. Genetic instabilities in human cancers. Nature 1998, 396, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Loeb, L.A. Mutator phenotype may be required for multistage carcinogenesis. Cancer Res. 1991, 51, 3075–3079. [Google Scholar] [PubMed]
- Ehrlich, M. DNA Alterations in Cancer: Genetic and Epigenetic Changes; Eaton Pub Co.: Natick, MA, USA, 2000. [Google Scholar]
- Walker, B.A.; Leone, P.E.; Jenner, M.W.; Li, C.; Gonzalez, D.; Johnson, D.C.; Ross, F.M.; Davies, F.E.; Morgan, G.J. Integration of global SNP-based mapping and expression arrays reveals key regions, mechanisms, and genes important in the pathogenesis of multiple myeloma. Blood 2006, 108, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Symington, L.S.; Gautier, J. Double-strand break end resection and repair pathway choice. Annu. Rev. Genet. 2011, 45, 247–271. [Google Scholar] [CrossRef] [PubMed]
- Startek, M.; Szafranski, P.; Gambin, T.; Campbell, I.M.; Hixson, P.; Shaw, C.A.; Stankiewicz, P.; Gambin, A. Genome-wide analyses of LINE–LINE-mediated nonallelic homologous recombination. Nucleic Acids Res. 2015, 43, 2188–2198. [Google Scholar] [CrossRef] [PubMed]
- Briggs, S.; Tomlinson, I. Germline and somatic polymerase ϵ and δ mutations define a new class of hypermutated colorectal and endometrial cancers. J. Pathol. 2013, 230, 148–153. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relevant Phenotype | Rate of Mitotic Crossovers/BIR (Terminal LOH) | Rate of Gene Conversions (Interstitial LOH) | Large (>5 kb) Interstitial Duplications Plus Deletions | Large (>5 kb) Terminal Duplications Plus Deletions | Ploidy Changes and UPD (Monosomy, Trisomy, UPD) | Base Substitutions Plus Small (≤700 bp) in/dels |
|---|---|---|---|---|---|---|
| Low Polα | 450 | 90 | 350 2 | 250 2,3 | (13,000; 190; 1200) 4 | 2 |
| Low Polδ | 200 | 15 | 240 2 | 90 2,3 | (16,000; 95; 800) 4 | 55 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, D.-Q.; Petes, T.D. Genome Instability Induced by Low Levels of Replicative DNA Polymerases in Yeast. Genes 2018, 9, 539. https://doi.org/10.3390/genes9110539
Zheng D-Q, Petes TD. Genome Instability Induced by Low Levels of Replicative DNA Polymerases in Yeast. Genes. 2018; 9(11):539. https://doi.org/10.3390/genes9110539
Chicago/Turabian StyleZheng, Dao-Qiong, and Thomas D. Petes. 2018. "Genome Instability Induced by Low Levels of Replicative DNA Polymerases in Yeast" Genes 9, no. 11: 539. https://doi.org/10.3390/genes9110539
APA StyleZheng, D.-Q., & Petes, T. D. (2018). Genome Instability Induced by Low Levels of Replicative DNA Polymerases in Yeast. Genes, 9(11), 539. https://doi.org/10.3390/genes9110539