Detection and Monitoring of Insecticide Resistance Mutations in Anopheles gambiae: Individual vs. Pooled Specimens

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Strains and Field Populations
2.2. Preparation of Pooled Samples
2.3. Nucleic Acids Extraction from Mosquito Pools
2.4. Multiplex Quantitative Polymerase Chain Reaction for Assessing kdr Mutations
2.5. Linear Regression Models for Determining Allelic Frequency in Mosquito Pools
3. Results
3.1. Quality Control of quantitative Polymerase Chain Reaction Assays and Genotyping of Mosquito Control Strains
3.2. Development of Regression Models with the Training Set
3.3. Validation of Regression Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Edi, C.V.; Koudou, B.G.; Jones, C.M.; Weetman, D.; Ranson, H. Multiple-insecticide resistance in Anopheles gambiae mosquitoes, Southern Côte d’Ivoire. Emerg. Infect. Dis. 2012, 18, 1508–1511. [Google Scholar] [CrossRef] [PubMed]
- Ranson, H.; N’Guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in African anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2011, 27, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Knox, T.B.; Juma, E.O.; Ochomo, E.O.; Pates Jamet, H.; Ndungo, L.; Chege, P.; Bayoh, N.M.; N’Guessan, R.; Christian, R.N.; Hunt, R.H.; et al. An online tool for mapping insecticide resistance in major Anopheles vectors of human malaria parasites and review of resistance status for the Afrotropical region. Parasit. Vectors 2014, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect. Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juarez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Nikou, D.; Donnelly, M.J.; Williamson, M.S.; Ranson, H.; Ball, A.; Vontas, J.; Field, L.M. Detection of knockdown resistance (kdr) mutations in Anopheles gambiae: A comparison of two new high-throughput assays with existing methods. Malar. J. 2007, 6, 111. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.P.; Santos, J.M.; Martins, A.J. Mutations in the voltage-gated sodium channel gene of anophelines and their association with resistance to Pyrethroids—A review. Parasit. Vectors 2014, 7, 450. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Liyanapathirana, M.; Agossa, F.R.; Weetman, D.; Ranson, H.; Donnelly, M.J.; Wilding, C.S. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2012, 109, 6614–6619. [Google Scholar] [CrossRef] [PubMed]
- David, J.P.; Ismail, H.M.; Chandor-Proust, A.; Paine, M.J. Role of cytochrome P450s in insecticide resistance: Impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120429. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.; Warr, E.; Stevenson, B.J.; Pignatelli, P.M.; Morgan, J.C.; Steven, A.; Yawson, A.E.; Mitchell, S.N.; Ranson, H.; Hemingway, J.; et al. Field-caught permethrin-resistant Anopheles gambiae overexpress CYP6P3, a P450 that metabolises pyrethroids. PLoS Genet. 2008, 4, e1000286. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.N.; Stevenson, B.J.; Muller, P.; Wilding, C.S.; Egyir-Yawson, A.; Field, S.G.; Hemingway, J.; Paine, M.J.; Ranson, H.; Donnelly, M.J. Identification and validation of a gene causing cross-resistance between insecticide classes in Anopheles gambiae from Ghana. Proc. Natl. Acad. Sci. USA 2012, 109, 6147–6152. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.J.; Isaacs, A.T.; Weetman, D. Identification, validation, and application of molecular diagnostics for insecticide resistance in malaria vectors. Trends Parasitol. 2016, 32, 197–206. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Techniques to Detect Insecticide Resistance Mechanisms (Field and Laboratory Manual); World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Bass, C.; Nikou, D.; Vontas, J.; Donnelly, M.J.; Williamson, M.S.; Field, L.M. The vector population monitoring tool (VPMT): High-throughput DNA-based diagnostics for the monitoring of mosquito vector populations. Malar. Res. Treat. 2010, 2010, 190434. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Vontas, J.; Poupardin, R.; Raman, J.; Lines, J.; Schwabe, C.; Matias, A.; Kleinschmidt, I. Country-level operational implementation of the Global Plan for Insecticide Resistance Management. Proc. Natl. Acad. Sci. USA 2013, 110, 9397–9402. [Google Scholar] [CrossRef] [PubMed]
- Vontas, J.; Mitsakakis, K.; Zengerle, R.; Yewhalaw, D.; Sikaala, C.H.; Etang, J.; Fallani, M.; Carman, B.; Muller, P.; Chouaibou, M.; et al. Automated innovative diagnostic, data management and communication tool, for improving malaria vector control in endemic settings. Stud. Health Technol. Inform. 2016, 224, 54–60. [Google Scholar] [PubMed]
- Chen, H.H.; Jou, Y.S.; Lee, W.J.; Pan, W.H. Applying polynomial standard curve method to correct bias encountered in estimating allele frequencies using DNA pooling strategy. Genomics 2008, 92, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Lavebratt, C.; Sengul, S.; Jansson, M.; Schalling, M. Pyrosequencing-based SNP allele frequency estimation in DNA pools. Hum. Mutat. 2004, 23, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Nikou, D.; Vontas, J.; Williamson, M.S.; Field, L.M. Development of high-throughput real-time PCR assays for the identification of insensitive acetylcholinesterase (ace-1R) in Anopheles gambiae. Pest. Biochem. Physiol. 2010, 96, 80–85. [Google Scholar] [CrossRef]
- Bass, C.; Williamson, M.S.; Field, L.M. Development of a multiplex real-time PCR assay for identification of members of the Anopheles gambiae species complex. Acta Trop. 2008, 107, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.E.; Barth, J.F.; Turell, M.J.; Gordon, S.W.; Sattabongkot, J.; Copeland, R.; Wirtz, R.A. Development and evaluation of a dipstick assay for detection of Plasmodium falciparum and P. vivax sporozoites in mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2000, 37, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Kefi, M.; Mavridis, K.; Simoes, M.L.; Dimopoulos, G.; Siden-Kiamos, I.; Vontas, J. New rapid one-step PCR diagnostic assay for Plasmodium falciparum infective mosquitoes. Sci. Rep. 2018, 8, 1462. [Google Scholar] [CrossRef] [PubMed]
- Mitsakakis, K.; Hin, S.; Muller, P.; Wipf, N.; Thomsen, E.; Coleman, M.; Zengerle, R.; Vontas, J.; Mavridis, K. Converging human and malaria vector diagnostics with data management towards an integrated holistic one health approach. Int. J. Environ. Res. Public Health 2018, 15, 259. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, F.; Schwemmer, F.; Hutzenlaub, T.; Baumann, D.; Strohmeier, O.; Dingemanns, G.; Simons, G.; Sager, C.; Plobner, L.; von Stetten, F.; et al. LabDisk with complete reagent prestorage for sample-to-answer nucleic acid based detection of respiratory pathogens verified with influenza A H3N2 virus. Lab Chip 2016, 16, 199–207. [Google Scholar] [CrossRef] [PubMed]



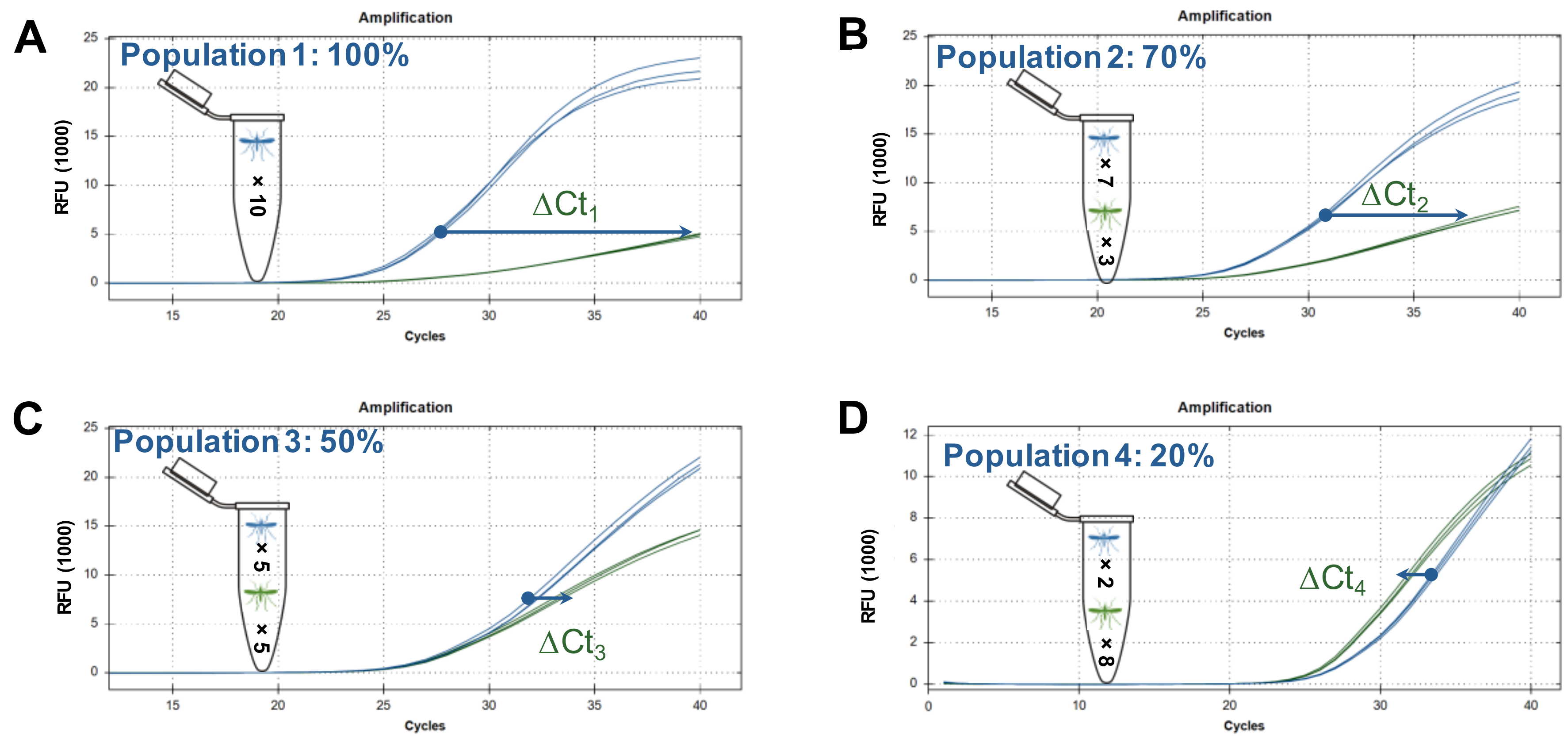{kind=link}
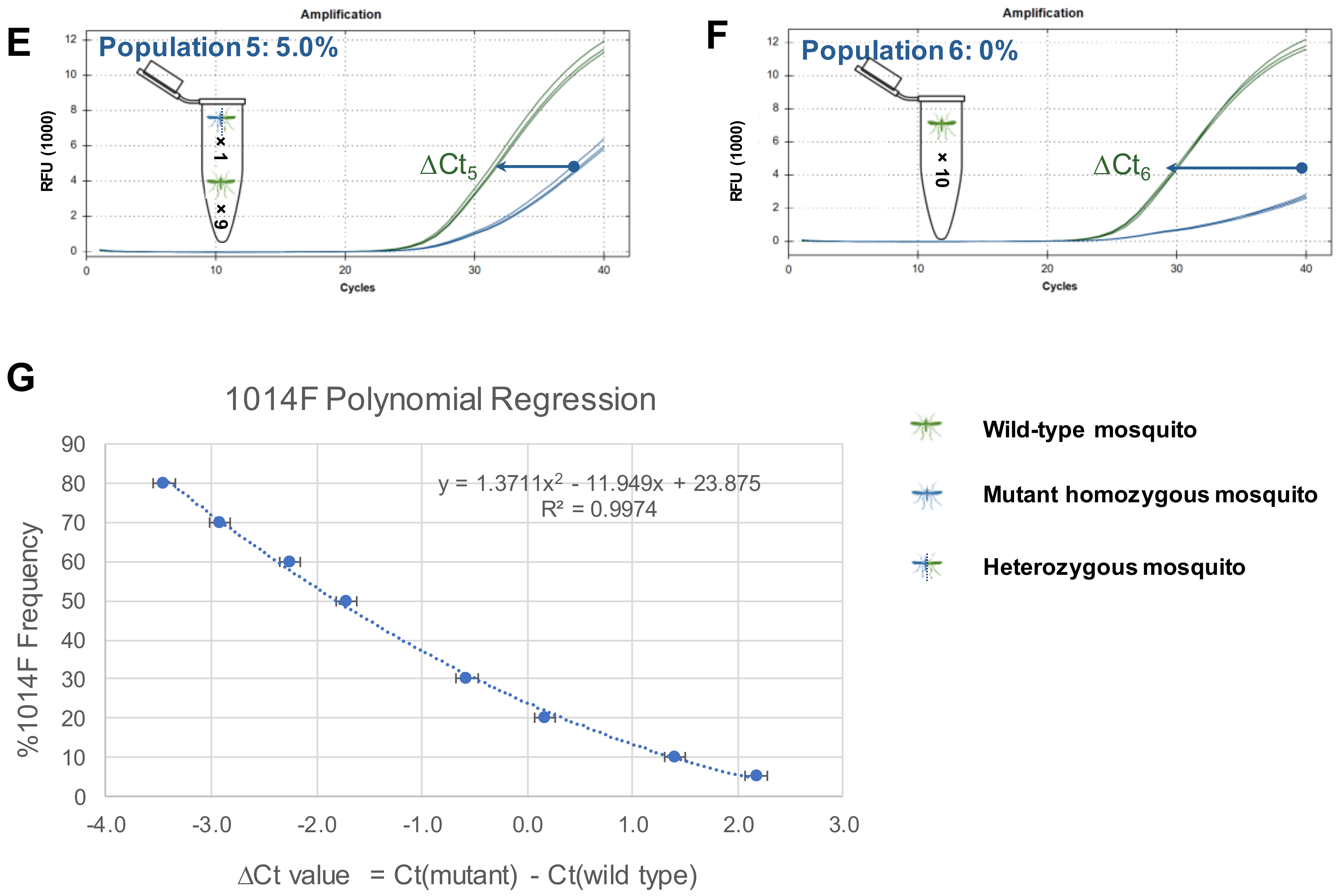{kind=link}
| Kdr Mutation | Equation | R Square (Adjusted) | SE of the Estimate | p Value | σ2pf Mean (Range) | Detection Limit |
|---|---|---|---|---|---|---|
| 1014F | %MAF = 1.37 × (ΔCt)2 − 11.9 × (ΔCt) + 23.9 | 0.996 | 1.72 | 3.55 × 10−7 | 4.9 (0.72–10.6) | 5.0% |
| 1014S | %MAF = 1.11 × (ΔCt)2 − 10.6 × (ΔCt) + 21.7 | 0.996 | 1.53 | 1.07 × 10−4 | 2.9 (1.6–12.1) | 5.0% |
| 1575Y | %MAF = 5.68 × (ΔCt)2 + 35.4 × (ΔCt) + 60.0 | 0.997 | 1.28 | 5.88 × 10−5 | 2.2 (0.13–16.6) | 5.0% |
| kdr | Accuracy ± SE | Precision (Range) | rs | p Value |
|---|---|---|---|---|
| 1014F | 3.58 ± 0.84 | 2.99 (1.73–3.66) | 0.978 | 5.20 × 10−6 |
| 1014S | 5.9 ± 1.5 | 2.32 (0.69–4.1) | 0.989 | 2.50 × 10−5 |
| 1575Y | 3.26 ± 0.62 | 1.66 (0.517–3.86) | 0.959 | 8.37 × 10−7 |
| kdr | Species | Population | Individuals Genotyped | Actual MAF | Measured MAF ± SE | Absolute Difference |
|---|---|---|---|---|---|---|
| 1014F | An. gambiae s.s. S-form and An. gambiae s.s. M-form | P1a, P1b | RR =6; RS = 0; SS = 4 | 60% | 55.54% ± 2.25 | 4.46% |
| P2a, P2b | RR =3; RS = 0; SS = 7 | 30% | 33.14% ± 1.97 | 3.14% | ||
| P3a, P3b | RR = 2; RS = 0; SS = 6 | 20% | 25.35% ± 2.59 | 5.35% | ||
| P4a, P4b | RR = 1; RS = 0; SS = 9 | 10% | 11.60% ± 1.22 | 1.60% | ||
| P5a | RR = 1; RS= 0; SS = 18 | 5% | 1.90% | 3.1% | ||
| 1014S | An. gambiae s.s. S-form | P6a, P6b | RR = 8; RS = 0; SS = 2 | 80% | 90.96% ± 1.64 | 10.96% |
| P7a | RR = 5; RS = 0; SS = 5 | 50% | 45.73% | 4.27% | ||
| P8a, P8b | RR = 3; RS = 0; SS = 7 | 30% | 33.54% ± 0.49 | 3.54% | ||
| P9a, P9b | RR = 1; RS = 0; SS = 9 | 10% | 14.00% ± 2.89 | 4.00% | ||
| 1575Y | An. gambiae s.s. S-form and An. gambiae s.s. M-form | P10a | RR = 4; RS = 2; SS = 4 | 50% | 47.94% | 2.06% |
| P11a, P11b | RR = 1; RS = 4; SS = 5 | 30% | 33.97% ± 1.05 | 3.97% | ||
| P12a, P12b | RR = 1; RS = 1; SS = 8 | 15% | 13.83% ± 2.19 | 1.17% | ||
| P13a, P13b | RR = 1; RS = 0; SS = 9 | 10% | 9.63% ± 3.56 | 0.37% | ||
| P14a, P14b | RR = 0; RS = 1; SS = 9 | 5% | 7.59% ± 1.85 | 2.59% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavridis, K.; Wipf, N.; Müller, P.; Traoré, M.M.; Muller, G.; Vontas, J. Detection and Monitoring of Insecticide Resistance Mutations in Anopheles gambiae: Individual vs. Pooled Specimens. Genes 2018, 9, 479. https://doi.org/10.3390/genes9100479
Mavridis K, Wipf N, Müller P, Traoré MM, Muller G, Vontas J. Detection and Monitoring of Insecticide Resistance Mutations in Anopheles gambiae: Individual vs. Pooled Specimens. Genes. 2018; 9(10):479. https://doi.org/10.3390/genes9100479
Chicago/Turabian StyleMavridis, Konstantinos, Nadja Wipf, Pie Müller, Mohamed M. Traoré, Gunter Muller, and John Vontas. 2018. "Detection and Monitoring of Insecticide Resistance Mutations in Anopheles gambiae: Individual vs. Pooled Specimens" Genes 9, no. 10: 479. https://doi.org/10.3390/genes9100479
APA StyleMavridis, K., Wipf, N., Müller, P., Traoré, M. M., Muller, G., & Vontas, J. (2018). Detection and Monitoring of Insecticide Resistance Mutations in Anopheles gambiae: Individual vs. Pooled Specimens. Genes, 9(10), 479. https://doi.org/10.3390/genes9100479