Advances in Non-Viral DNA Vectors for Gene Therapy

Abstract
:1. Introduction to Gene Therapy
2. Challenges of Using Plasmid Vectors for Gene Therapy
3. Improvements to Plasmid Vectors
4. Development of Minicircles and Minivectors
4.1. Minicircles
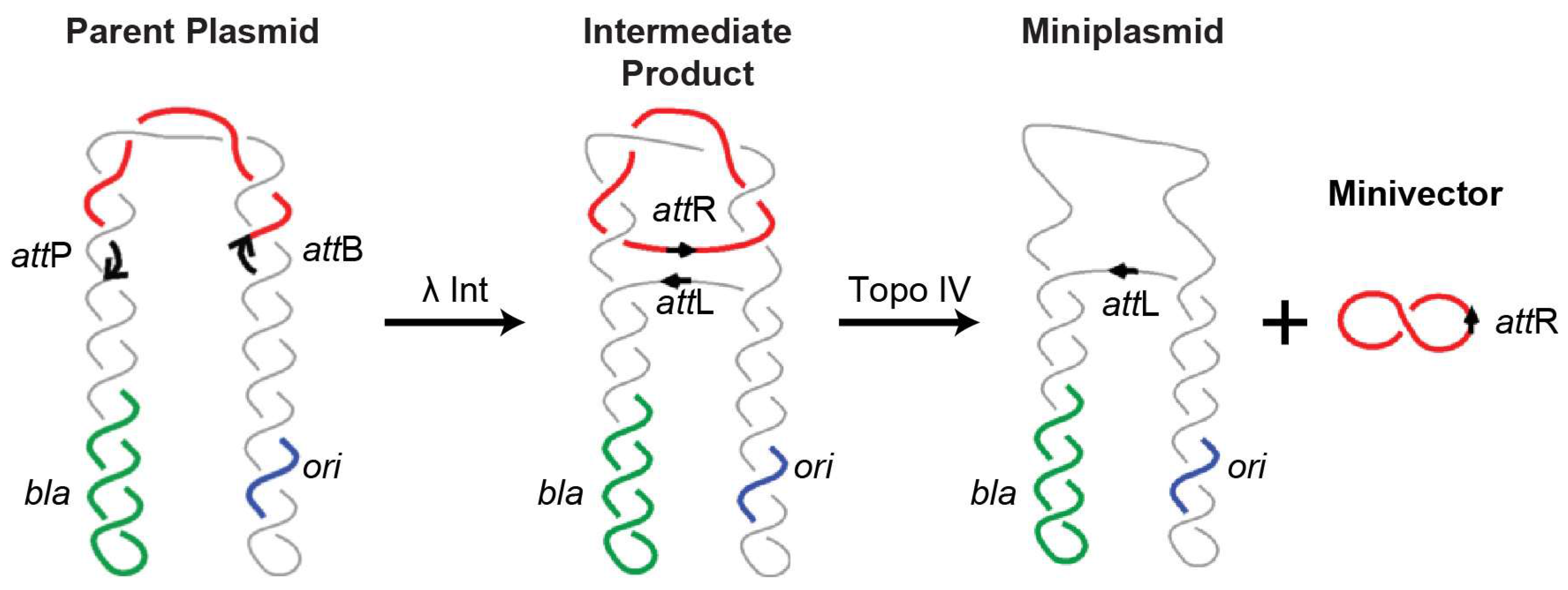
4.2. Minivectors
4.3. Increasing the Functionality of Minimized DNA Vectors
4.3.1. Nuclear Localization Signals
4.3.2. Cytosine-Phosphate-Guanine Dinucleotides
4.3.3. Scaffold/Matrix Attachment Regions
4.3.4. Viral Replication Genes
5. Therapeutic Needs Best Addressed by Minimized Vectors
5.1. DNA Vaccines
5.2. Cancer and Immunotherapy
5.3. Stem Cell Reprogramming
5.4. Therapy for the Lungs
5.5. Cardiovascular Uses
5.6. Dermal Uses
6. Concluding Remarks: Moving Minimized Non-Viral DNA Vectors into the Clinic
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rao, R.C.; Zacks, D.N. Cell and gene therapy. Dev. Ophthalmol. 2014, 53, 167–177. [Google Scholar] [PubMed]
- Cooney, A.L.; Abou Alaiwa, M.H.; Shah, V.S.; Bouzek, D.C.; Stroik, M.R.; Powers, L.S.; Gansemer, N.D.; Meyerholz, D.K.; Welsh, M.J.; Stoltz, D.A.; et al. Lentiviral-mediated phenotypic correction of cystic fibrosis pigs. JCI Insight 2016, 1, e88730. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, R.; Berges, B.K.; Solis-Leal, A.; Igbinedion, O.; Strong, C.L.; Schiller, M.R. TALEN gene editing takes aim on HIV. Hum. Genet. 2016, 135, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Samson, S.L.; Gonzalez, E.V.; Yechoor, V.; Bajaj, M.; Oka, K.; Chan, L. Gene therapy for diabetes: metabolic effects of helper-dependent adenoviral exendin 4 expression in a diet-induced obesity mouse model. Mol. Ther. J. Am. Soc. Gene Ther. 2008, 16, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- Bicho, D.; Queiroz, J.A.; Tomaz, C.T. Influenza plasmid DNA vaccines: Progress and prospects. Curr. Gene Ther. 2015, 15, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Al-Dosari, M.S.; Gao, X. Nonviral gene delivery: Principle, limitations, and recent progress. AAPS J. 2009, 11, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.H.; Hill, A.; Chen, M.; Pfeifer, B.A. Contemporary approaches for nonviral gene therapy. Discov. Med. 2015, 19, 447–454. [Google Scholar] [PubMed]
- Keles, E.; Song, Y.; Du, D.; Dong, W.-J.; Lin, Y. Recent progress in nanomaterials for gene delivery applications. Biomater. Sci. 2016, 4, 1291–1309. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, V.; de Melo-Diogo, D.; Costa, E.; Moreira, A.; Queiroz, J.; Pichon, C.; Correia, I.; Sousa, F. Minicircle DNA vectors for gene therapy: Advances and applications. Expert Opin. Biol. Ther. 2015, 15, 353–379. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey-Abina, S.; de Saint Basile, G.; Cavazzana-Calvo, M. Gene therapy of X-linked severe combined immunodeficiency. Methods Mol. Biol. Clifton NJ 2003, 215, 247–259. [Google Scholar]
- Hacein-Bey-Abina, S.; von Kalle, C.; Schmidt, M.; Le Deist, F.; Wulffraat, N.; McIntyre, E.; Radford, I.; Villeval, J.-L.; Fraser, C.C.; Cavazzana-Calvo, M.; Fischer, A. A serious adverse event after successful gene therapy for X-linked severe combined immunodeficiency. N. Engl. J. Med. 2003, 348, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Loring, H.S.; ElMallah, M.K.; Flotte, T.R. Development of rAAV2-CFTR: History of the First rAAV Vector Product to be Used in Humans. Hum. Gene Ther. Methods 2016, 27, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Wold, W.S.M.; Toth, K. Adenovirus vectors for gene therapy, vaccination and cancer gene therapy. Curr. Gene Ther. 2013, 13, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.E.; Ehrhardt, A.; Kay, M. A Progress and problems with the use of viral vectors for gene therapy. Nat. Rev. Genet. 2003, 4, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Kreppel, F.; Kochanek, S. Modification of adenovirus gene transfer vectors with synthetic polymers: A scientific review and technical guide. Mol. Ther. J. Am. Soc. Gene Ther. 2008, 16, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Sinn, P.L.; Burnight, E.R.; McCray, P.B. Progress and Prospects: Prospects of repeated pulmonary administration of viral vectors. Gene Ther. 2009, 16, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Lesch, H.P.; Pikkarainen, J.T.; Kaikkonen, M.U.; Taavitsainen, M.; Samaranayake, H.; Lehtolainen-Dalkilic, P.; Vuorio, T.; Määttä, A.-M.; Wirth, T.; Airenne, K.J.; et al. Avidin fusion protein-expressing lentiviral vector for targeted drug delivery. Hum. Gene Ther. 2009, 20, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Schleef, M. Minicircle and Miniplasmid DNA Vectors: The Future of Non-Viral and Viral Gene Transfer; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Kormann, M.S.D.; Hasenpusch, G.; Aneja, M.K.; Nica, G.; Flemmer, A.W.; Herber-Jonat, S.; Huppmann, M.; Mays, L.E.; Illenyi, M.; Schams, A. Expression of therapeutic proteins after delivery of chemically modified mRNA in mice. Nat. Biotechnol. 2011, 29, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Izsvák, Z.; Chuah, M.K.L.; Vandendriessche, T.; Ivics, Z. Efficient stable gene transfer into human cells by the Sleeping Beauty transposon vectors. Methods San Diego Calif. 2009, 49, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Home - ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/home (accessed on 6 February 2017).
- New and Updated Clinical Gene Therapy Trials. Available online: http://www.genetherapynet.com/clinicaltrialsgov.html (accessed on 6 February 2017).
- Rodriguez, E. Nonviral DNA vectors for immunization and therapy: Design and methods for their obtention. J. Mol. Med. 2004, 82, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Mairhofer, J.; Grabherr, R. Rational vector design for efficient non-viral gene delivery: Challenges facing the use of plasmid DNA. Mol. Biotechnol. 2008, 39, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Stenler, S.; Blomberg, P.; Smith, C.I.E. Safety and efficacy of DNA vaccines: Plasmids vs. minicircles. Hum. Vaccines Immunother. 2014, 10, 1306–1308. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, F.; Xu, S.; Fire, A.Z.; Kay, M.A. The extragenic spacer length between the 5’ and 3’ ends of the transgene expression cassette affects transgene silencing from plasmid-based vectors. Mol. Ther. J. Am. Soc. Gene Ther. 2012, 20, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Faurez, F.; Dory, D.; Le Moigne, V.; Gravier, R.; Jestin, A. Biosafety of DNA vaccines: New generation of DNA vectors and current knowledge on the fate of plasmids after injection. Vaccine 2010, 28, 3888–3895. [Google Scholar] [CrossRef] [PubMed]
- Ahmad-Nejad, P.; Häcker, H.; Rutz, M.; Bauer, S.; Vabulas, R.M.; Wagner, H. Bacterial CpG-DNA and lipopolysaccharides activate Toll-like receptors at distinct cellular compartments. Eur. J. Immunol. 2002, 32, 1958–1968. [Google Scholar] [CrossRef]
- Kreiss, P.; Cameron, B.; Rangara, R.; Mailhe, P.; Aguerre-Charriol, O.; Airiau, M.; Scherman, D.; Crouzet, J.; Pitard, B. Plasmid DNA size does not affect the physicochemical properties of lipoplexes but modulates gene transfer efficiency. Nucleic Acids Res. 1999, 27, 3792–3798. [Google Scholar] [CrossRef] [PubMed]
- Stenler, S.; Wiklander, O.P.; Badal-Tejedor, M.; Turunen, J.; Nordin, J.Z.; Hallengärd, D.; Wahren, B.; El Andaloussi, S.; Rutland, M.W.; Smith, C.E. Micro-minicircle gene therapy: Implications of size on fermentation, complexation, shearing resistance, and expression. Mol. Ther. Acids 2014, 2, e140. [Google Scholar] [CrossRef] [PubMed]
- Hornstein, B.D.; Roman, D.; Arévalo-Soliz, L.M.; Engevik, M.A.; Zechiedrich, L. Effects of circular DNA length on transfection efficiency by electroporation into HeLa cells. PLoS ONE 2016, 11, e0167537. [Google Scholar] [CrossRef] [PubMed]
- Cranenburgh, R.M.; Hanak, J.A.; Williams, S.G.; Sherratt, D.J. Escherichia coli strains that allow antibiotic-free plasmid selection and maintenance by repressor titration. Nucleic Acids Res. 2001, 29, E26. [Google Scholar] [CrossRef] [PubMed]
- Soubrier, F.; Cameron, B.; Manse, B.; Somarriba, S.; Dubertret, C.; Jaslin, G.; Jung, G.; Caer, C.L.; Dang, D.; Mouvault, J.M.; et al. pCOR: A new design of plasmid vectors for nonviral gene therapy. Gene Ther. 1999, 6, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Marie, C.; Vandermeulen, G.; Quiviger, M.; Richard, M.; Préat, V.; Scherman, D. pFARs, plasmids free of antibiotic resistance markers, display high-level transgene expression in muscle, skin and tumour cells. J. Gene Med. 2010, 12, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Mwau, M.; Cebere, I.; Sutton, J.; Chikoti, P.; Winstone, N.; Wee, E.G.-T.; Beattie, T.; Chen, Y.-H.; Dorrell, L.; McShane, H.; et al. A human immunodeficiency virus 1 (HIV-1) clade A vaccine in clinical trials: Stimulation of HIV-specific T-cell responses by DNA and recombinant modified vaccinia virus Ankara (MVA) vaccines in humans. J. Gen. Virol. 2004, 85, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Ramos, I.; Alonso, A.; Peris, A.; Marcen, J.M.; Abengozar, M.A.; Alcolea, P.J.; Castillo, J.A.; Larraga, V. Antibiotic resistance free plasmid DNA expressing LACK protein leads towards a protective Th1 response against Leishmania infantum infection. Vaccine 2009, 27, 6695–6703. [Google Scholar] [CrossRef] [PubMed]
- Saubi, N.; Mbewe-Mvula, A.; Gea-Mallorqui, E.; Rosario, M.; Gatell, J.M.; Hanke, T.; Joseph, J. Pre-clinical development of BCG.HIVA(CAT), an antibiotic-free selection strain, for HIV-TB pediatric vaccine vectored by lysine auxotroph of BCG. PLoS ONE 2012, 7, e42559. [Google Scholar] [CrossRef] [PubMed]
- Garmory, H.S.; Leckenby, M.W.; Griffin, K.F.; Elvin, S.J.; Taylor, R.R.; Hartley, M.G.; Hanak, J.A.J.; Williamson, E.D.; Cranenburgh, R.M. Antibiotic-free plasmid stabilization by operator-repressor titration for vaccine delivery by using live Salmonella enterica Serovar typhimurium. Infect. Immun. 2005, 73, 2005–2011. [Google Scholar] [CrossRef] [PubMed]
- Leckenby, M.W.; Spear, A.M.; Neeson, B.N.; Williamson, E.D.; Cranenburgh, R.M.; Atkins, H.S. Enhanced vaccine antigen delivery by Salmonella using antibiotic-free operator-repressor titration-based plasmid stabilisation compared to chromosomal integration. Microb. Pathog. 2009, 46, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-M.; Sali, M.; Leckenby, M.W.; Radford, D.S.; Huynh, H.A.; Delogu, G.; Cranenburgh, R.M.; Cutting, S.M. Oral delivery of a DNA vaccine against tuberculosis using operator-repressor titration in a Salmonella enterica vector. Vaccine 2010, 28, 7523–7528. [Google Scholar] [CrossRef] [PubMed]
- Lilly, J.; Camps, M. Mechanisms of theta plasmid replication. Microbiol. Spectr. 2015, 3, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Nikol, S.; Baumgartner, I.; Van Belle, E.; Diehm, C.; Visoná, A.; Capogrossi, M.C.; Ferreira-Maldent, N.; Gallino, A.; Wyatt, M.G.; Wijesinghe, L.D. TALISMAN 201 investigators Therapeutic angiogenesis with intramuscular NV1FGF improves amputation-free survival in patients with critical limb ischemia. Mol. Ther. J. Am. Soc. Gene Ther. 2008, 16, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Vandermeulen, G.; Marie, C.; Scherman, D.; Préat, V. New generation of plasmid backbones devoid of antibiotic resistance marker for gene therapy trials. Mol. Ther. 2011, 19, 1942–1949. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.H.; Mairhofer, J. Marker-free plasmids for biotechnological applications—Implications and perspectives. Trends Biotechnol. 2013, 31, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Bathula, S.R.; Huang, L.; Abraham, D.J. Gene Therapy with Plasmid DNA. In Burger’s Medicinal Chemistry and Drug Discovery; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003. [Google Scholar]
- Williams, S.G.; Cranenburgh, R.M.; Weiss, A.M.; Wrighton, C.J.; Sherratt, D.J.; Hanak, J.A. Repressor titration: A novel system for selection and stable maintenance of recombinant plasmids. Nucleic Acids Res. 1998, 26, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Cranenburgh, R.M.; Lewis, K.S.; Hanak, J.A.J. Effect of plasmid copy number and lac operator sequence on antibiotic-free plasmid selection by operator-repressor titration in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2004, 7, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Durany, O.; Bassett, P.; Weiss, A.M.E.; Cranenburgh, R.M.; Ferrer, P.; López-Santín, J.; de Mas, C.; Hanak, J.A.J. Production of fuculose-1-phosphate aldolase using operator-repressor titration for plasmid maintenance in high cell density Escherichia coli fermentations. Biotechnol. Bioeng. 2005, 91, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Kornowski, R.; Fuchs, S.; Epstein, S.E.; Branellec, D.; Schwartz, B. Catheter-based plasmid-mediated transfer of genes into ischemic myocardium using the pCOR plasmid. Coron. Artery Dis. 2000, 11, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Soubrier, F.; Laborderie, B.; Cameron, B. Improvement of pCOR plasmid copy number for pharmaceutical applications. Appl. Microbiol. Biotechnol. 2005, 66, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Witzenbichler, B.; Mahfoudi, A.; Soubrier, F.; Le Roux, A.; Branellec, D.; Schultheiss, H.-P.; Isner, J.M. Intramuscular gene transfer of fibroblast growth factor-1 using improved pCOR plasmid design stimulates collateral formation in a rabbit ischemic hindlimb model. J. Mol. Med. Berl. Ger. 2006, 84, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Maulik, N. NV1FGF, a pCOR plasmid-based angiogenic gene therapy for the treatment of intermittent claudication and critical limb ischemia. Curr. Opin. Investig. Drugs Lond. Engl. 2000 2009, 10, 259–268. [Google Scholar]
- Van Melderen, L. Molecular interactions of the CcdB poison with its bacterial target, the DNA gyrase. Int. J. Med. Microbiol. IJMM 2002, 291, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Szpirer, C.Y.; Milinkovitch, M.C. Separate-component-stabilization system for protein and DNA production without the use of antibiotics. BioTechniques 2005, 38, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Peubez, I.; Chaudet, N.; Mignon, C.; Hild, G.; Husson, S.; Courtois, V.; De Luca, K.; Speck, D.; Sodoyer, R. Antibiotic-free selection in E. coli: New considerations for optimal design and improved production. Microb. Cell Factories 2010, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Luke, J.; Carnes, A.E.; Hodgson, C.P.; Williams, J.A. Improved antibiotic-free DNA vaccine vectors utilizing a novel RNA based plasmid selection system. Vaccine 2009, 27, 6454–6459. [Google Scholar] [CrossRef] [PubMed]
- Luke, J.M.; Carnes, A.E.; Williams, J.A. Development of antibiotic-free selection system for safer DNA vaccination. Methods Mol. Biol. Clifton NJ 2014, 1143, 91–111. [Google Scholar]
- Pfaffenzeller, I.; Mairhofer, J.; Striedner, G.; Bayer, K.; Grabherr, R. Using ColE1-derived RNA I for suppression of a bacterially encoded gene: Implication for a novel plasmid addiction system. Biotechnol. J. 2006, 1, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Mairhofer, J.; Cserjan-Puschmann, M.; Striedner, G.; Nöbauer, K.; Razzazi-Fazeli, E.; Grabherr, R. Marker-free plasmids for gene therapeutic applications—Lack of antibiotic resistance gene substantially improves the manufacturing process. J. Biotechnol. 2010, 146, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.H.; Real, L.; Bailey, M.W. An Array of Escherichia coli clones over-expressing essential proteins: A new strategy of identifying cellular targets of potent antibacterial compounds. Biochem. Biophys. Res. Commun. 2006, 349, 1250. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.; Good, L. Plasmid selection in Escherichia coli using an endogenous essential gene marker. BMC Biotechnol. 2008, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Šimčíková, M.; Prather, K.L.J.; Prazeres, D.M.F.; Monteiro, G.A. Towards effective non-viral gene delivery vector. Biotechnol. Genet. Eng. Rev. 2015, 31, 82–107. [Google Scholar] [CrossRef] [PubMed]
- Darquet, A.M.; Cameron, B.; Wils, P.; Scherman, D.; Crouzet, J. A new DNA vehicle for nonviral gene delivery: Supercoiled minicircle. Gene Ther. 1997, 4, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Kreiss, P.; Cameron, B.; Darquet, A.M.; Scherman, D.; Crouzet, J. Production of a new DNA vehicle for gene transfer using site-specific recombination. Appl. Microbiol. Biotechnol. 1998, 49, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Darquet, A.M.; Rangara, R.; Kreiss, P.; Schwartz, B.; Naimi, S.; Delaere, P.; Crouzet, J.; Scherman, D. Minicircle: An improved DNA molecule for in vitro and in vivo gene transfer. Gene Ther. 1999, 6, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Bigger, B.W.; Tolmachov, O.; Collombet, J.M.; Fragkos, M.; Palaszewski, I.; Coutelle, C. An araC-controlled bacterial cre expression system to produce DNA minicircle vectors for nuclear and mitochondrial gene therapy. J. Biol. Chem. 2001, 276, 23018–23027. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, V.M.; Maia, C.J.; Queiroz, J.A.; Pichon, C.; Correia, I.J.; Sousa, F. Improved minicircle DNA biosynthesis for gene therapy applications. Hum. Gene Ther. Methods 2014, 25, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Schleef, M.; Schirmbeck, R.; Reiser, M.; Michel, M.-L.; Schmeer, M. Minicircle: Next generation DNA vectors for vaccination. Methods Mol. Biol. Clifton NJ 2015, 1317, 327–339. [Google Scholar]
- Catanese, D.J.; Fogg, J.M.; Schrock, D.E.; Gilbert, B.E.; Zechiedrich, L. Supercoiled minivector DNA resists shear forces associated with gene therapy delivery. Gene Ther. 2012, 19, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Fogg, J.M.; Zechiedrich, L.; Zu, Y. Transfection of shRNA-encoding Minivector DNA of a few hundred base pairs to regulate gene expression in lymphoma cells. Gene Ther. 2011, 18, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Fogg, J.M.; Kolmakova, N.; Rees, I.; Magonov, S.; Hansma, H.; Perona, J.J.; Zechiedrich, E.L. Exploring writhe in supercoiled minicircle DNA. J. Phys. Condens. Matter Inst. Phys. J. 2006, 18, S145–S159. [Google Scholar] [CrossRef] [PubMed]
- Tolmachov, O.E. Tightly-wound miniknot vectors for gene therapy: A potential improvement over supercoiled minicircle DNA. Med. Hypotheses 2010, 74, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Schirmbeck, R.; König-Merediz, S.A.; Riedl, P.; Kwissa, M.; Sack, F.; Schroff, M.; Junghans, C.; Reimann, J.; Wittig, B. Priming of immune responses to hepatitis B surface antigen with minimal DNA expression constructs modified with a nuclear localization signal peptide. J. Mol. Med. Berl. Ger. 2001, 79, 343–350. [Google Scholar] [CrossRef]
- Wittig, B.; Märten, A.; Dorbic, T.; Weineck, S.; Min, H.; Niemitz, S.; Trojaneck, B.; Flieger, D.; Kruopis, S.; Albers, A.; et al. Therapeutic vaccination against metastatic carcinoma by expression-modulated and immunomodified autologous tumor cells: A first clinical phase I/II trial. Hum. Gene Ther. 2001, 12, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Schakowski, F.; Gorschlüter, M.; Junghans, C.; Schroff, M.; Buttgereit, P.; Ziske, C.; Schöttker, B.; König-Merediz, S.A.; Sauerbruch, T.; Wittig, B.; et al. A novel minimal-size vector (MIDGE) improves transgene expression in colon carcinoma cells and avoids transfection of undesired DNA. Mol. Ther. J. Am. Soc. Gene Ther. 2001, 3, 793–800. [Google Scholar] [CrossRef] [PubMed]
- López-Fuertes, L.; Pérez-Jiménez, E.; Vila-Coro, A.J.; Sack, F.; Moreno, S.; Konig, S.A.; Junghans, C.; Wittig, B.; Timón, M.; Esteban, M. DNA vaccination with linear minimalistic (MIDGE) vectors confers protection against Leishmania major infection in mice. Vaccine 2002, 21, 247–257. [Google Scholar] [CrossRef]
- Moreno, S.; López-Fuertes, L.; Vila-Coro, A.J.; Sack, F.; Smith, C.A.; Konig, S.A.; Wittig, B.; Schroff, M.; Juhls, C.; Junghans, C.; Timón, M. DNA immunisation with minimalistic expression constructs. Vaccine 2004, 22, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Schakowski, F.; Gorschlüter, M.; Buttgereit, P.; Märten, A.; Lilienfeld-Toal, M.V.; Junghans, C.; Schroff, M.; König-Merediz, S.A.; Ziske, C.; Strehl, J.; et al. Minimal size MIDGE vectors improve transgene expression in vivo. Vivo Athens Greece 2007, 21, 17–23. [Google Scholar]
- Endmann, A.; Baden, M.; Weisermann, E.; Kapp, K.; Schroff, M.; Kleuss, C.; Wittig, B.; Juhls, C. Immune response induced by a linear DNA vector: Influence of dose, formulation and route of injection. Vaccine 2010, 28, 3642–3649. [Google Scholar] [CrossRef] [PubMed]
- Galling, N.; Kobelt, D.; Aumann, J.; Schmidt, M.; Wittig, B.; Schlag, P.M.; Walther, W. Intratumoral dispersion, retention, systemic biodistribution, and clearance of a small-size tumor necrosis factor-α-expressing MIDGE vector after nonviral in vivo jet-injection gene transfer. Hum. Gene Ther. Methods 2012, 23, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, X.; Peng, X.; Jiang, W.; Yao, L. Construction of PPENK-MIDGE-NLS gene vector and the expression in rat. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2015, 31, 258–268. [Google Scholar]
- Jiang, X.; Yu, H.; Teo, C.R.; Tan, G.S.X.; Goh, S.C.; Patel, P.; Chua, Y.K.; Hameed, N.B.S.; Bertoletti, A.; Patzel, V. Advanced design of dumbbell-shaped genetic minimal vectors improves non-coding and coding RNA expression. Mol. Ther. 2016, 24, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-S.; Chen, Z.-J.; Zhang, G.; Ou, X.-L.; Yang, X.-L.; Wong, C.K.C.; Giesy, J.P.; Du, J.; Chen, S.-Y. A novel micro-linear vector for in vitro and in vivo gene delivery and its application for EBV positive tumors. PLoS ONE 2012, 7, e47159. [Google Scholar] [CrossRef] [PubMed]
- Nafissi, N.; Slavcev, R. Construction and characterization of an in vivo linear covalently closed DNA vector production system. Microb. Cell Factories 2012, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Nafissi, N.; Alqawlaq, S.; Lee, E.A.; Foldvari, M.; Spagnuolo, P.A.; Slavcev, R.A. DNA ministrings: Highly safe and effective gene delivery vectors. Mol. Ther. Nucleic Acids 2014, 3, e165. [Google Scholar] [CrossRef] [PubMed]
- Sum, C.H.; Nafissi, N.; Slavcev, R.A.; Wettig, S. Physical characterization of gemini surfactant-based synthetic vectors for the delivery of linear covalently closed (LCC) DNA ministrings. PLoS ONE 2015, 10, e0142875. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Lam, P.; Nafissi, N.; Denniss, S.; Slavcev, R. Production of double-stranded DNA ministrings. J. Vis. Exp. JoVE 2016, 108, 53177. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, F.; Kay, M.A. A Mini-intronic Plasmid (MIP): A novel robust transgene expression vector in vivo and in vitro. Mol. Ther. 2013, 21, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Riu, E.; He, C.-Y.; Xu, H.; Kay, M.A. Silencing of episomal transgene expression in liver by plasmid bacterial backbone DNA is independent of CpG methylation. Mol. Ther. 2008, 16, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.H.; Guo, X.Y.; Chen, Y.; He, C.-Y.; Chen, Z.-Y. Increasing the minicircle DNA purity using an enhanced triplex DNA technology to eliminate DNA contaminants. Mol. Ther. Methods Clin. Dev. 2015, 1, 14062. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.P.A.; Šimčíková, M.; Brito, L.; Monteiro, G.A.; Prazeres, D.M.F. Development of a nicking endonuclease-assisted method for the purification of minicircles. J. Chromatogr. A 2016, 1443, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Diamantino, T.; Pereira, P.; Queiroz, J.A.; Sousa, Â.; Sousa, F. Minicircle DNA purification using a CIM® DEAE-1 monolithic support: Liquid chromatography. J. Sep. Sci. 2016, 39, 3544–3549. [Google Scholar] [CrossRef] [PubMed]
- Šimčíková, M.; Alves, C.P.A.; Brito, L.; Prather, K.L.J.; Prazeres, D.M.F.; Monteiro, G.A. Improvement of DNA minicircle production by optimization of the secondary structure of the 5′-UTR of ParA resolvase. Appl. Microbiol. Biotechnol. 2016, 100, 6725–6737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, H.; Gao, S.; Huang, W.; Wang, Z. Polyethylenimine and minicircle DNA based gene transfer. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2010, 26, 772–779. [Google Scholar]
- Dietz, W.M.; Skinner, N.E.B.; Hamilton, S.E.; Jund, M.D.; Heitfeld, S.M.; Litterman, A.J.; Hwu, P.; Chen, Z.-Y.; Salazar, A.M.; Ohlfest, J.R.; et al. Minicircle DNA is superior to plasmid DNA in eliciting antigen-specific CD8+ T-cell responses. Mol. Ther. 2013, 21, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Remaut, K.; Sanders, N.N.; Fayazpour, F.; Demeester, J.; De Smedt, S.C. Influence of plasmid DNA topology on the transfection properties of DOTAP/DOPE lipoplexes. J. Control. Release Off. J. Control. Release Soc. 2006, 115, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Bodine, T.; Catanese D.J., Jr.; Arevalo-Soliz, L.M.; Fogg, J.M.; Zechiedrich, L. Effect of size and topology on DNA vector survival in human serum. Unpublished.
- Munkonge, F.M.; Dean, D.A.; Hillery, E.; Griesenbach, U.; Alton, E.W.F.W. Emerging significance of plasmid DNA nuclear import in gene therapy. Adv. Drug Deliv. Rev. 2003, 55, 749–760. [Google Scholar] [CrossRef]
- Brandén, L.J.; Mohamed, A.J.; Smith, C.I. A peptide nucleic acid-nuclear localization signal fusion that mediates nuclear transport of DNA. Nat. Biotechnol. 1999, 17, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Zanta, M.A.; Belguise-Valladier, P.; Behr, J.P. Gene delivery: A single nuclear localization signal peptide is sufficient to carry DNA to the cell nucleus. Proc. Natl. Acad. Sci. USA 1999, 96, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Vaysse, L.; Gregory, L.G.; Harbottle, R.P.; Perouzel, E.; Tolmachov, O.; Coutelle, C. Nuclear-targeted minicircle to enhance gene transfer with non-viral vectors in vitro and in vivo. J. Gene Med. 2006, 8, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.K.; Liu, F.; Liu, D. Enhanced gene expression in mouse lung by prolonging the retention time of intravenously injected plasmid DNA. Gene Ther. 1998, 5, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M. Cpg motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, J.; Krieg, A.M. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv. Drug Deliv. Rev. 2009, 61, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M.; Yi, A.K.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995, 374, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Alonso, S.; Martinez-Lopez, A.; Estepa, A.; Cuesta, A.; Tafalla, C. The introduction of multi-copy CpG motifs into an antiviral DNA vaccine strongly up-regulates its immunogenicity in fish. Vaccine 2011, 29, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shi, J.-L.; Wu, X.-Y.; Fu, F.; Yu, J.; Yuan, X.-Y.; Peng, Z.; Cong, X.-Y.; Xu, S.-J.; Sun, W.-B.; et al. Improvement of the immunogenicity of porcine circovirus type 2 DNA vaccine by recombinant ORF2 gene and CpG motifs. Viral Immunol. 2015, 28, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, Q.; Hou, S.; Zhai, G.; Zhu, H.; Sánchez-Vizcaíno, J.M. Plasmid containing CpG motifs enhances the efficacy of porcine reproductive and respiratory syndrome live attenuated vaccine. Vet. Immunol. Immunopathol. 2011, 144, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, J.; Klinman, D.M. Clinical evaluation of CpG oligonucleotides as adjuvants for vaccines targeting infectious diseases and cancer. Vaccine 2014, 32, 6377. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Jiang, W.; Chen, Y.; Liu, P.; Sheng, C.; Chen, S.; Zhang, H.; Pan, C.; Gao, S.; Huang, W. In vivo electroporation of minicircle DNA as a novel method of vaccine delivery to enhance HIV-1-specific immune responses. J. Virol. 2014, 88, 1924–1934. [Google Scholar] [CrossRef] [PubMed]
- Mirkovitch, J.; Mirault, M.-E.; Laemmli, U.K. Organization of the higher-order chromatin loop: Specific DNA attachment sites on nuclear scaffold. Cell 1984, 39, 223–232. [Google Scholar] [CrossRef]
- Wong, S.P.; Argyros, O.; Harbottle, R.P. Sustained expression from DNA vectors. Adv. Genet. 2015, 89, 113–152. [Google Scholar] [PubMed]
- Patrushev, L.I.; Kovalenko, T.F. Functions of noncoding sequences in mammalian genomes. Biochem. Biokhimiia 2014, 79, 1442–1469. [Google Scholar] [CrossRef] [PubMed]
- Argyros, O.; Wong, S.P.; Fedonidis, C.; Tolmachov, O.; Waddington, S.N.; Howe, S.J.; Niceta, M.; Coutelle, C.; Harbottle, R.P. Development of S/MAR minicircles for enhanced and persistent transgene expression in the mouse liver. J. Mol. Med. Berl. Ger. 2011, 89, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Keaton, M.A.; Taylor, C.M.; Layer, R.M.; Dutta, A. Nuclear scaffold attachment sites within ENCODE regions associate with actively transcribed genes. PLoS ONE 2011, 6, e17912. [Google Scholar] [CrossRef] [PubMed]
- Verghese, S.C.; Goloviznina, N.A.; Skinner, A.M.; Lipps, H.J.; Kurre, P. S/MAR sequence confers long-term mitotic stability on non-integrating lentiviral vector episomes without selection. Nucleic Acids Res. 2014, 42, e53. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, F.; Zhang, L.; Lu, J.; Xu, P.; Liu, G.; Xie, X.; Mu, W.; Wang, Y.; Liu, D. Non-integrating lentiviral vectors based on the minimal S/MAR sequence retain transgene expression in dividing cells. Sci. China Life Sci. 2016, 59, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Fotaki, G.; Ramachandran, M.; Nilsson, B.; Essand, M.; Yu, D. Safe engineering of CAR T cells for adoptive cell therapy of cancer using long-term episomal gene transfer. EMBO Mol. Med. 2016, 8, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Koirala, A.; Conley, S.M.; Naash, M.I. Episomal maintenance of S/MAR-containing non-viral vectors for RPE-based diseases. Adv. Exp. Med. Biol. 2014, 801, 703–709. [Google Scholar] [PubMed]
- Nehlsen, K.; Broll, S.; Bode, J. Replicating minicircles: Generation of nonviral episomes for the efficient modification of dividing cells. Gene Ther Mol Biol 2006, 10, 233–244. [Google Scholar]
- Van Craenenbroeck, K.; Vanhoenacker, P.; Haegeman, G. Episomal vectors for gene expression in mammalian cells. Eur. J. Biochem. 2000, 267, 5665–5678. [Google Scholar] [CrossRef] [PubMed]
- Leight, E.R.; Sugden, B. EBNA-1: A protein pivotal to latent infection by Epstein-Barr virus. Rev. Med. Virol. 2000, 10, 83–100. [Google Scholar] [CrossRef]
- Ali Hosseini Rad, S.M.; Bamdad, T.; Arefian, E.; Mossahebi-Mohammadi, M.; Sadeghizadeh, M. An EBV-based plasmid can replicate and maintain in stem cells. Biotechnol. Prog. 2015, 31, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Drozd, A.M.; Walczak, M.P.; Piaskowski, S.; Stoczynska-Fidelus, E.; Rieske, P.; Grzela, D.P. Generation of human iPSCs from cells of fibroblastic and epithelial origin by means of the oriP/EBNA-1 episomal reprogramming system. Stem Cell Res. Ther. 2015, 6, 122. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.A.; Juranek, S.; Lipps, H.J. Designing nonviral vectors for efficient gene transfer and long-term gene expression. Mol. Ther. 2006, 14, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Geiben-Lynn, R.; Greenland, J.R.; Frimpong-Boateng, K.; van Rooijen, N.; Hovav, A.-H.; Letvin, N.L. CD4+ T lymphocytes mediate in vivo clearance of plasmid DNA vaccine antigen expression and potentiate CD8+ T-cell immune responses. Blood 2008, 112, 4585–4590. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.A. Improving DNA vaccine performance through vector design. Curr. Gene Ther. 2014, 14, 170–189. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, F.; Rostami, S.; Ghayour-Mobarhan, M.; Meshkat, Z. Current progress in the development of therapeutic vaccines for chronic hepatitis B virus infection. Iran. J. Basic Med. Sci. 2016, 19, 692–704. [Google Scholar] [PubMed]
- Halstead, S.B.; Thomas, S.J. New Japanese encephalitis vaccines: Alternatives to production in mouse brain. Expert Rev. Vaccines 2011, 10, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Borggren, M.; Nielsen, J.; Karlsson, I.; Dalgaard, T.S.; Trebbien, R.; Williams, J.A.; Fomsgaard, A. A polyvalent influenza DNA vaccine applied by needle-free intradermal delivery induces cross-reactive humoral and cellular immune responses in pigs. Vaccine 2016, 34, 3634–3640. [Google Scholar] [CrossRef] [PubMed]
- Borggren, M.; Nielsen, J.; Bragstad, K.; Karlsson, I.; Krog, J.S.; Williams, J.A.; Fomsgaard, A. Vector optimization and needle-free intradermal application of a broadly protective polyvalent influenza A DNA vaccine for pigs and humans. Hum. Vaccines Immunother. 2015, 11, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- The Influenza Virus: Structure and Replication—Article in Motion. Available online: http://www.rapidreferenceinfluenza.com/chapter/B978-0-7234-3433-7.50009-8/aim/influenza-virus-structure (accessed on 28 October 2016).
- Sousa, F.; Passarinha, L.; Queiroz, J.A. Biomedical application of plasmid DNA in gene therapy: A new challenge for chromatography. Biotechnol. Genet. Eng. Rev. 2009, 26, 83–116. [Google Scholar] [CrossRef]
- Gorecki, D.C. “Dressed-up” naked plasmids: Emerging vectors for non-viral gene therapy. Discov. Med. 2009, 6, 191–197. [Google Scholar]
- Williams, P.D.; Kingston, P.A. Plasmid-mediated gene therapy for cardiovascular disease. Cardiovasc. Res. 2011, 91, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xiao, X.; Zhao, P.; Xue, G.; Zhu, Y.; Zhu, X.; Zheng, L.; Zeng, Y.; Huang, W. Minicircle-IFNgamma induces antiproliferative and antitumoral effects in human nasopharyngeal carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 4702–4713. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Volz, B.; Wittig, B. MIDGE Vectors and dSLIM Immunomodulators: DNA-based Molecules for Gene Therapeutic Strategies. In Modern Biopharmaceuticals; Knäblein, J., Ed.; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2005; pp. 183–211. [Google Scholar]
- Köchling, J.; Prada, J.; Bahrami, M.; Stripecke, R.; Seeger, K.; Henze, G.; Wittig, B.; Schmidt, M. Anti-tumor effect of DNA-based vaccination and dSLIM immunomodulatory molecules in mice with Ph+ acute lymphoblastic leukaemia. Vaccine 2008, 26, 4669–4675. [Google Scholar] [CrossRef] [PubMed]
- Kobelt, D.; Aumann, J.; Schmidt, M.; Wittig, B.; Fichtner, I.; Behrens, D.; Lemm, M.; Freundt, G.; Schlag, P.M.; Walther, W. Preclinical study on combined chemo- and nonviral gene therapy for sensitization of melanoma using a human TNF-alpha expressing MIDGE DNA vector. Mol. Oncol. 2014, 8, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Monjezi, R.; Miskey, C.; Gogishvili, T.; Schleef, M.; Schmeer, M.; Einsele, H.; Ivics, Z.; Hudecek, M. Enhanced CAR T-cell engineering using non-viral Sleeping Beauty transposition from minicircle vectors. Leukemia 2016. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Ma, F.; Zhang, P.; Zhong, Y.; Zhang, J.; Wang, T.; Zheng, G.; Hou, X.; Zhao, J.; He, C.-Y.; Chen, Z.-Y. Treatment of human B-cell lymphomas using minicircle DNA vector expressing anti-CD3/CD20 in a mouse model. Hum. Gene Ther. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ronald, J.A.; Chuang, H.-Y.; Dragulescu-Andrasi, A.; Hori, S.S.; Gambhir, S.S. Detecting cancers through tumor-activatable minicircles that lead to a detectable blood biomarker. Proc. Natl. Acad. Sci. USA 2015, 112, 3068–3073. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Wilson, K.D.; Sun, N.; Gupta, D.M.; Huang, M.; Li, Z.; Panetta, N.J.; Chen, Z.Y.; Robbins, R.C.; Kay, M.A.; et al. A nonviral minicircle vector for deriving human iPS cells. Nat. Methods 2010, 7, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Daneshvar, N.; Rasedee, A.; Shamsabadi, F.T.; Moeini, H.; Mehrboud, P.; Rahman, H.S.; Boroojerdi, M.H.; Vellasamy, S. Induction of pluripotency in human umbilical cord mesenchymal stem cells in feeder layer-free condition. Tissue Cell 2015, 47, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.R.; Chari, D.M. Part II: Functional delivery of a neurotherapeutic gene to neural stem cells using minicircle DNA and nanoparticles: Translational advantages for regenerative neurology. J. Control. Release Off. J. Control. Release Soc. 2016, 238, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Griesenbach, U.; Geddes, D.M.; Alton, E.W.F.W. Gene therapy for cystic fibrosis: An example for lung gene therapy. Gene Ther. 2004, 11, S43–S50. [Google Scholar] [CrossRef] [PubMed]
- Munye, M.M.; Tagalakis, A.D.; Barnes, J.L.; Brown, R.E.; McAnulty, R.J.; Howe, S.J.; Hart, S.L. Minicircle DNA provides enhanced and prolonged transgene expression following airway gene transfer. Sci. Rep. 2016, 6, 23125. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Engelhardt, J.F. Current status of gene therapy for inherited lung diseases. Annu. Rev. Physiol. 2003, 65, 585–612. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.; Munegowda, M.A.; Cao, H.; Hu, J. Lung gene therapy—How to capture illumination from the light already present in the tunnel. Genes Dis. 2014, 1, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Duncan, G.A.; Jung, J.; Hanes, J.; Suk, J.S. The mucus barrier to inhaled gene therapy. Mol. Ther. 2016, 24, 2043–2053. [Google Scholar] [CrossRef] [PubMed]
- Alton, E.W.F.W.; Boyd, A.C.; Cheng, S.H.; Davies, J.C.; Davies, L.A.; Dayan, A.; Gill, D.R.; Griesenbach, U.; Higgins, T.; Hyde, S.C.; et al. Toxicology study assessing efficacy and safety of repeated administration of lipid/DNA complexes to mouse lung. Gene Ther. 2014, 21, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, S.-P.; Whitmore, M.; Loeffert, E.J.; Wang, L.; Watkins, S.C.; Pitt, B.R.; Huang, L. Effect of immune response on gene transfer to the lung via systemic administration of cationic lipidic vectors. Am. J. Physiol.-Lung Cell. Mol. Physiol. 1999, 276, L796–L804. [Google Scholar]
- Bazzani, R.P.; Pringle, I.A.; Connolly, M.M.; Davies, L.A.; Sumner-Jones, S.G.; Schleef, M.; Hyde, S.C.; Gill, D.R. Transgene sequences free of CG dinucleotides lead to high level, long-term expression in the lung independent of plasmid backbone design. Biomaterials 2016, 93, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Matkar, P.N.; Leong-Poi, H.; Singh, K.K. Cardiac gene therapy: Are we there yet? Gene Ther. 2016, 23, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Stenler, S.; Andersson, A.; Simonson, O.E.; Lundin, K.E.; Chen, Z.-Y.; Kay, M.A.; Smith, C.E.; Sylvén, C.; Blomberg, P. Gene transfer to mouse heart and skeletal muscles using a minicircle expressing human vascular endothelial growth factor. J. Cardiovasc. Pharmacol. 2009, 53, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Jiao, R.; Guo, X.; Wang, T.; Chen, P.; Wang, D.; Chen, Y.; He, C.-Y.; Chen, Z.-Y. Construction of minicircle DNA vectors capable of correcting familial hypercholesterolemia phenotype in a LDLR-deficient mouse model. Gene Ther. 2016, 23, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Huang, M.; Li, Z.; Jia, F.; Ghosh, Z.; Lijkwan, M.A.; Fasanaro, P.; Sun, N.; Wang, X.; Martelli, F.; et al. MicroRNA-210 as a novel therapy for treatment of ischemic heart disease. Circulation 2010, 122, S124–S131. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Nguyen, P.; Jia, F.; Hu, S.; Gong, Y.; de Almeida, P.E.; Wang, L.; Nag, D.; Kay, M.A.; Giaccia, A.J.; et al. Double knockdown of prolyl hydroxylase and factor-inhibiting hypoxia-inducible factor with nonviral minicircle gene therapy enhances stem cell mobilization and angiogenesis after myocardial infarction. Circulation 2011, 124, S46–S54. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.S.; Jung, H.S.; Kwon, M.J.; Lee, S.H.; Kim, C.W.; Kim, M.K.; Lee, M.; Park, J.H. Sonoporation of the minicircle-VEGF(165) for wound healing of diabetic mice. Pharm. Res. 2009, 26, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Zibert, J.R.; Wallbrecht, K.; Schön, M.; Mir, L.M.; Jacobsen, G.K.; Trochon-Joseph, V.; Bouquet, C.; Villadsen, L.S.; Cadossi, R.; Skov, L.; et al. Halting angiogenesis by non-viral somatic gene therapy alleviates psoriasis and murine psoriasiform skin lesions. J. Clin. Invest. 2011, 121, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.R.; Tovey, J.C.K.; Sharma, A.; Tandon, A. Gene therapy in the Cornea: 2005–Present. Prog. Retin. Eye Res. 2012, 31, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Gene Therapy Clinical Trials Worldwide. Available online: http://www.abedia.com/wiley/years.php (accessed on 6 February 2017).
- Lijkwan, M.A.; Hellingman, A.A.; Bos, E.J.; van der Bogt, K.E.A.; Huang, M.; Kooreman, N.G.; de Vries, M.R.; Peters, H.A.B.; Robbins, R.C.; Hamming, J.F. Short hairpin RNA gene silencing of prolyl hydroxylase-2 with a minicircle vector improves neovascularization of hindlimb ischemia. Hum. Gene Ther. 2014, 25, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Rim, Y.A.; Yi, H.; Kim, Y.; Park, N.; Jung, H.; Kim, J.; Jung, S.M.; Park, S.-H.; Ju, J.H. Self in vivo production of a synthetic biological drug CTLA4Ig using a minicircle vector. Sci. Rep. 2014, 4, 6935. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Kim, Y.; Kim, J.; Jung, H.; Rim, Y.A.; Jung, S.M.; Park, S.-H.; Ju, J.H. A new strategy to deliver synthetic protein drugs: Self-reproducible biologics using minicircles. Sci. Rep. 2014, 4, 5961. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, V.M.; Gonçalves, C.; de Melo-Diogo, D.; Costa, E.C.; Queiroz, J.A.; Pichon, C.; Sousa, F.; Correia, I.J. Poly(2-ethyl-2-oxazoline)-PLA-g-PEI amphiphilic triblock micelles for co-delivery of minicircle DNA and chemotherapeutics. J. Control. Release Off. J. Control. Release Soc. 2014, 189, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, V.M.; Baril, P.; Costa, E.C.; de Melo-Diogo, D.; Foucher, F.; Queiroz, J.A.; Sousa, F.; Pichon, C.; Correia, I.J. Bioreducible poly(2-ethyl-2-oxazoline)-PLA-PEI-SS triblock copolymer micelles for co-delivery of DNA minicircles and doxorubicin. J. Control. Release Off. J. Control. Release Soc. 2015, 213, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, V.M.; Moreira, A.F.; Costa, E.C.; Queiroz, J.A.; Sousa, F.; Pichon, C.; Correia, I.J. Gas-generating TPGS-PLGA microspheres loaded with nanoparticles (NIMPS) for co-delivery of minicircle DNA and anti-tumoral drugs. Colloids Surf. B Biointerfaces 2015, 134, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Nobles, C.L.; Zechiedrich, L.; Maresso, A.W.; Silberg, J.J. Combining random gene fission and rational gene fusion to discover near-infrared fluorescent protein fragments that report on protein-protein interactions. ACS Synth. Biol. 2015, 4, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, A.; Segall-Shapiro, T.H.; Kou, S.; Jindal, G.A.; Hoff, K.G.; Liu, S.; Chitsaz, M.; Ismagilov, R.F.; Silberg, J.J.; Tirrell, D.A. A genetically encoded AND gate for cell-targeted metabolic labeling of proteins. J. Am. Chem. Soc. 2013, 135, 2979–2982. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Irobalieva, R.N.; Chiu, W.; Schmid, M.F.; Fogg, J.M.; Zechiedrich, L.; Pettitt, B.M. Influence of DNA sequence and structure of minicircles under torsional stress. (submitted for publication).
- Michnick, S.W.; Ear, P.H.; Manderson, E.N.; Remy, I.; Stefan, E. Universal strategies in research and drug discovery based on protein-fragment complementation assays. Nat. Rev. Drug Discov. 2007, 6, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, A.; Perrin, D.; Jacob, F.; Monod, J. Identification, by in vitro complementation and purification, of a peptide fraction of Escherichia coli beta-galactosidase. J. Mol. Biol. 1965, 12, 918–923. [Google Scholar] [CrossRef]


{kind=link}
| Non-Viral DNA Vector | Disease1 (n) | Phase | ||
|---|---|---|---|---|
| Plasmid (190 studies) | Monogenic | Cystic fibrosis (1) | 1/2 | |
| Polygenic | Cancer | T-cell immunotherapy (32) | 1,2 | |
| Therapeutic DNA vaccines (31) | 1,2 | |||
| Other cancer treatments (32) | 1,2 | |||
| Diabetes (1) | 1,2 | |||
| Heart (13) | 1,2 | |||
| Other2 (10) | 1,2 | |||
| Infectious | Hepatitis B (5) | 1,2 | ||
| Hepatitis C (3) | 1,2 | |||
| HIV (52) | 1,2 | |||
| HPV (2) | 1,2 | |||
| Influenza (14) | 1 | |||
| Malaria (2) | 1 | |||
| Other3 (22) | 1,2 | |||
| Naked DNA (9 studies) | Monogenic | Von Willebrand disease (1) | - | |
| Polygenic | Cancer (2) | 1/2 | ||
| Cancer vaccine (2) | 1 | |||
| Infectious | Hepatitis B (2) | 1,2 | ||
| HIV (3) | 1 | |||
| Oligonucleotide (141 studies) | Monogenic | Cystic fibrosis (2) | 1,2 | |
| Muscular dystrophy (4) | 1,2 | |||
| Von Willebrand disease (2) | 2 | |||
| Polygenic | Asthma (6) | 1,2 | ||
| Cancer (79) | 1–4 | |||
| Diabetes (3) | 1,2 | |||
| Heart (5) | 1,2 | |||
| Other4 (26) | 1-3 | |||
| Infectious | Hepatitis B (3) | 1,2 | ||
| Hepatitis C (1) | 2 | |||
| HIV (3) | 1,2 | |||
| Other5 (7) | 1-3 | |||
| pCOR6 (1 study) | Polygenic | Heart (1) | 2 | |
| pORT7 (1 study) | Infectious | HIV (1) | 1 | |
| MIDGE8 (1 study) | Polygenic | Cancer | 1/2 |
| Type of DNA vector | ori | AR | Advantages | Disadvantages | Reference |
|---|---|---|---|---|---|
| Plasmid | Yes | Yes | Safer than viruses and can be delivered more than once 1 Low risk of integration1 Can accommodate a long genetic payload Cheap, and easy to construct, produce, and store | DNA carrier/vehicle introduces additional complexity 1 Poor transfection efficiency naked CpG motifs can lead to inflammation and/or gene silencing Difficulty surrounding residual antibiotic and/or endotoxin | [47] |
| AR-free miniplasmids | Yes | No | Shorter than plasmids Lower metabolic burden on host strain | Still contains bacterial sequence which can lead to immune response and transgene silencing | [18,45,46] |
| pORT | Sequence addition to miniplasmid is short and selection does not involve gene expression Used in clinical trials as a DNA vaccine Easy to generate and produce | Mutant host strain required for propagation | [34,37,38,39,40,41,42,48,49,50] | ||
| pCOR | Narrow host range/low risk of spread No requirement for complex growth medium Used in clinical trials | Mutant host strain required for propagation tRNA expression required for selection | [35,44,51,52,53,54] | ||
| pFAR | Vector is only 1.1 kb The amber mutation used is very efficiently suppressed Growth medium does not contain animal-derived components Production process yields mostly monomeric closed circular DNA Both the plasmid and host strain are well-defined and described | Mutant host strain required for propagation tRNA expression required for selection | [36] | ||
| Post-segregational killing (PSK) systems | Sequence addition to miniplasmid is short Toxin is highly efficient for selection | PSK genes can cause mild toxicity | [55,56,57] | ||
| RNA IN/RNA OUT | RNA sequence is only transcribed in prokaryotes | Mutant host strain required for propagation | [58,59] | ||
| RNA I/RNA II | No sequence addition to plasmid necessary Can be produced in gram quantities more easily than standard plasmids | Mutant host strain required for propagation | [60,61] | ||
| Overexpression systems | No mutant host strain needed | Not much shorter than plasmids Potential metabolic burden on host strain Possible antibiotic contamination of final product | [62,63] | ||
| Circular Covalently Closed Vectors | No | No | Enhanced transfection and persistence in vitro and in vivo Little to no bacterial sequence | High production costs relative to plasmids Potential issues with scaling for clinical use | [9,18,32,64] |
| Minicircle | Vectors have been designed that are appropriate for mammalian mitochondrial gene therapy | Some plasmid and other DNA contaminants can remain in the final product | [27,65,66,67,68,69,70] | ||
| Minivector | Smallest circular DNA vector Most supercoiled DNA vector Greatest purity Naked minivector <1200 bp resists nebulization shear forces | Not well-known in the field | [33,71,72,73] | ||
| Miniknot | Potentially superior compactness Potentially resistant to nicking Potentially valuable for forceful administration procedures (e.g., aerosolization, jet-injection, etc.) | In hypothesis stage | [74] | ||
| Linear Covalently Closed Vectors (“dumbbell-shaped”) | No | No | Decreased risk of negative genome insertion events and nuclease attack because of protected ends Some systems are just the expression cassette alone | Vectors do not exhibit normal supercoiling, possessing only the qualities of linear DNA | |
| MIDGE | Smallest expression vectors reported Used in phase 1−2 clinical trials as a DNA vaccine Can be chemically modified to allow targeting | Potential rapid clearance Costly and time-consuming production | [75,76,77,78,79,80,81,82,83,84] | ||
| MiLV | Production process avoids bacterial contaminants such as endotoxin | PCR amplification of product is potentially error prone | [85] | ||
| Ministring | Chromosomal integration causes apoptotic cell death One-step in vivo production system This system can also create ccc vector topology | Uses ampicillin resistance as a selection marker Residual ampicillin or endotoxin possible in final product | [86,87,88,89] | ||
| Mini-intronic plasmid | Yes2 | No | Production process and yield is the same as standard plasmids, but subsequent splicing removes problematic sequences Inclusion of introns is reported to enhance transgene expression Uses RNA IN/RNA OUT as a selectable marker system Reported to have higher transgene expression levels than plasmids or minicircles | Vector length is not as reduced as some minimal systems, requiring the use of more transfection reagent Small chance of unspliced or mis-spliced mRNA Possible immune recognition of ori or other bacterial sequences before splicing occurs | [90] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardee, C.L.; Arévalo-Soliz, L.M.; Hornstein, B.D.; Zechiedrich, L. Advances in Non-Viral DNA Vectors for Gene Therapy. Genes 2017, 8, 65. https://doi.org/10.3390/genes8020065
Hardee CL, Arévalo-Soliz LM, Hornstein BD, Zechiedrich L. Advances in Non-Viral DNA Vectors for Gene Therapy. Genes. 2017; 8(2):65. https://doi.org/10.3390/genes8020065
Chicago/Turabian StyleHardee, Cinnamon L., Lirio Milenka Arévalo-Soliz, Benjamin D. Hornstein, and Lynn Zechiedrich. 2017. "Advances in Non-Viral DNA Vectors for Gene Therapy" Genes 8, no. 2: 65. https://doi.org/10.3390/genes8020065
APA StyleHardee, C. L., Arévalo-Soliz, L. M., Hornstein, B. D., & Zechiedrich, L. (2017). Advances in Non-Viral DNA Vectors for Gene Therapy. Genes, 8(2), 65. https://doi.org/10.3390/genes8020065