
Centromere Stability: The Replication Connection


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. DNA Replication
2.1. Assembly and Activation of the Replisome
2.2. DNA Replication Stress Causes Genome Instability
3. Centromere Dynamics
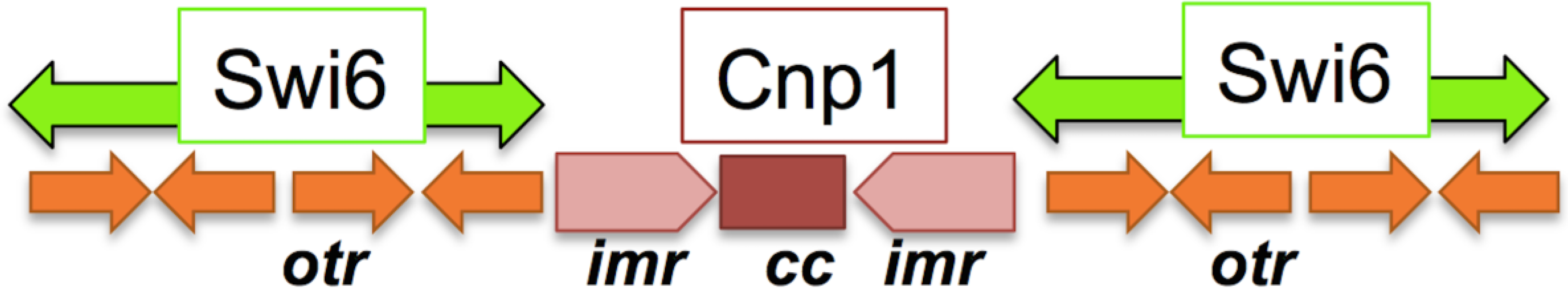
3.1. Centromere Structure
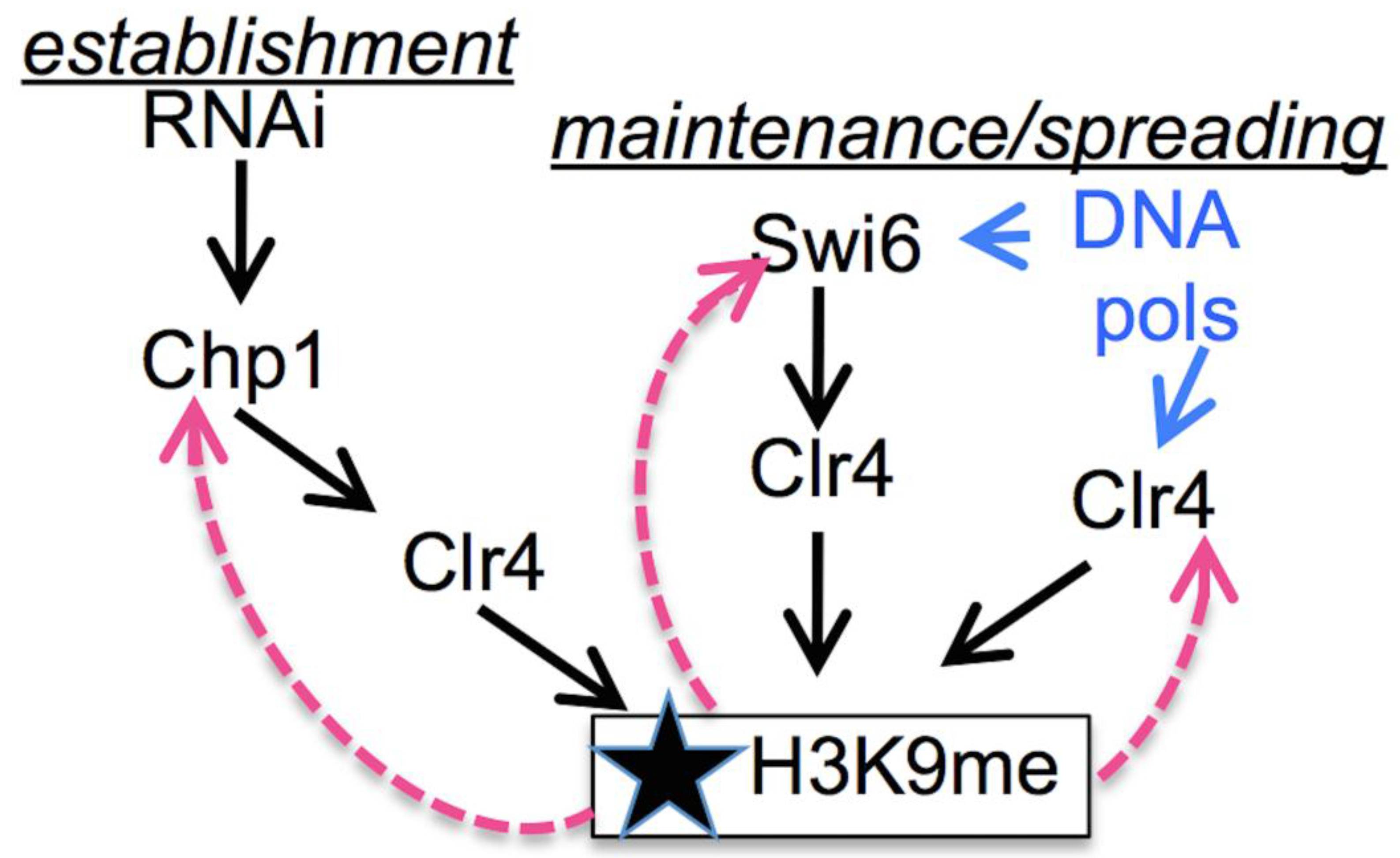
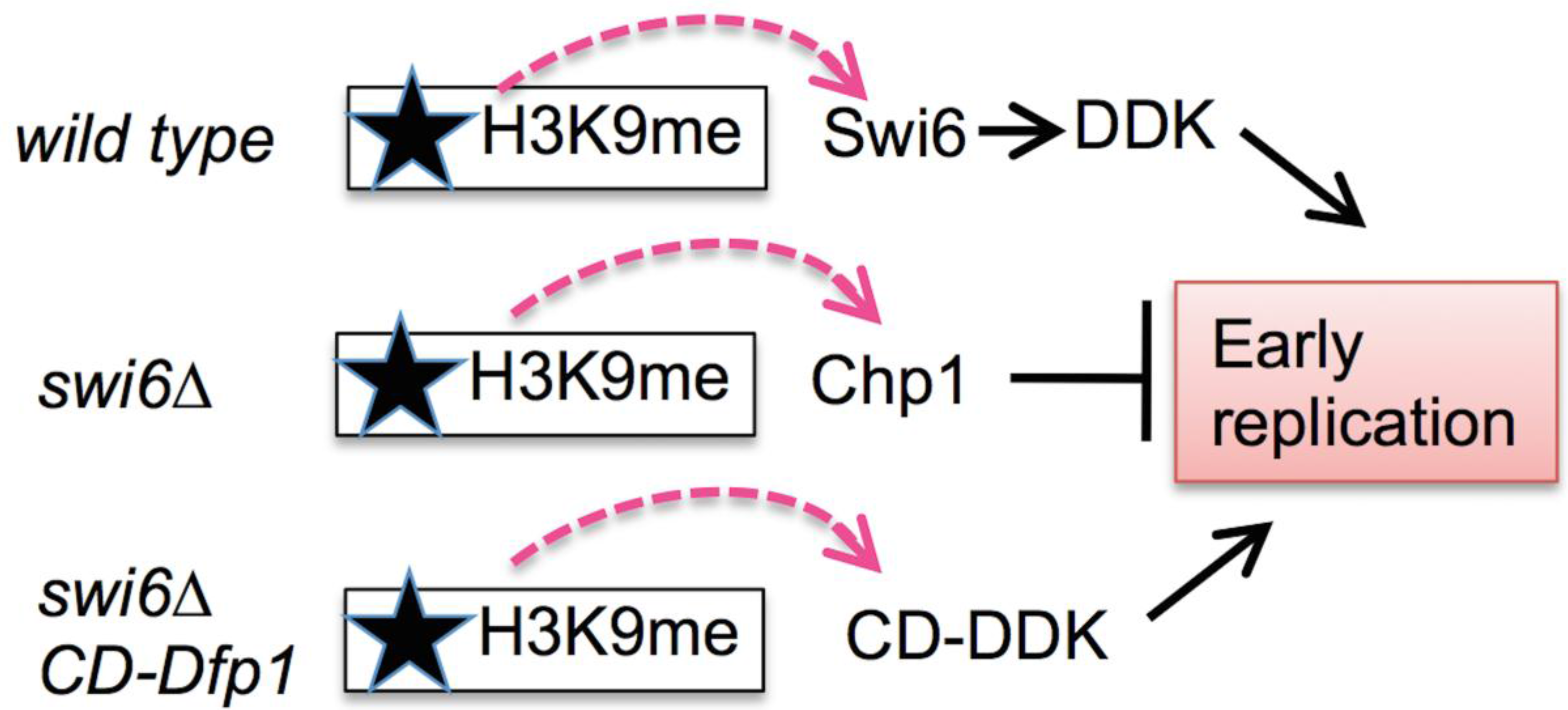
3.2. Early Replication in the Centromere
3.3. Genome Stress in the Centromere
4. Cohesion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Verdaasdonk, J.S.; Bloom, K. Centromeres: Unique chromatin structures that drive chromosome segregation. Nat. Rev. Mol. Cell Biol. 2011, 12, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Slee, R.B.; Steiner, C.M.; Herbert, B.S.; Vance, G.H.; Hickey, R.J.; Schwarz, T.; Christan, S.; Radovich, M.; Schneider, B.P.; Schindelhauer, D.; et al. Cancer-associated alteration of pericentromeric heterochromatin may contribute to chromosome instability. Oncogene 2012, 31, 3244–3253. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.C.; van Wely, K.H. Centromere fission, not telomere erosion, triggers chromosomal instability in human carcinomas. Carcinogenesis 2011, 32, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Beeharry, N.; Rattner, J.B.; Caviston, J.P.; Yen, T. Centromere fragmentation is a common mitotic defect of S and G 2 checkpoint override. Cell Cycle 2013, 12, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.; van der Burg, M.; Szuhai, K.; Kops, G.J.; Medema, R.H. Chromosome segregation errors as a cause of DNA damage and structural chromosome aberrations. Science 2011, 333, 1895–1898. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.A.; Moazed, D. Centromere assembly and propagation. Cell 2007, 128, 647–650. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.U.; Clayton, L.; Natsume, T. Three wise centromere functions: See no error, hear no break, speak no delay. EMBO Rep. 2013, 14, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Voineagu, I.; Surka, C.F.; Shishkin, A.A.; Krasilnikova, M.M.; Mirkin, S.M. Replisome stalling and stabilization at CGG repeats, which are responsible for chromosomal fragility. Nat. Struct. Mol. Biol. 2009, 16, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Voineagu, I.; Narayanan, V.; Lobachev, K.S.; Mirkin, S.M. Replication stalling at unstable inverted repeats: Interplay between DNA hairpins and fork stabilizing proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 9936–9941. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Lambert, S.; Baldacci, G.; Murray, J.M.; Carr, A.M. Nearby inverted repeats fuse to generate acentric and dicentric palindromic chromosomes by a replication template exchange mechanism. Genes Dev. 2009, 23, 2876–2886. [Google Scholar] [CrossRef] [PubMed]
- Sofueva, S.; Du, L.L.; Limbo, O.; Williams, J.S.; Russell, P. Brct domain interactions with phospho-histone H2A target Crb2 to chromatin at double-strand breaks and maintain the DNA damage checkpoint. Mol. Cell. Biol. 2010, 30, 4732–4743. [Google Scholar] [CrossRef] [PubMed]
- Arlt, M.F.; Durkin, S.G.; Ragland, R.L.; Glover, T.W. Common fragile sites as targets for chromosome rearrangements. DNA Repair (Amst) 2006, 5, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Alper, B.J.; Lowe, B.R.; Partridge, J.F. Centromeric heterochromatin assembly in fission yeast—Balancing transcription, RNA interference and chromatin modification. Chromosome Res. 2012, 20, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Li, P.C.; Petreaca, R.C.; Jensen, A.; Yuan, J.P.; Green, M.D.; Forsburg, S.L. Replication fork stability is essential for the maintenance of centromere integrity in the absence of heterochromatin. Cell Rep. 2013, 3, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Dialynas, G.K.; Vitalini, M.W.; Wallrath, L.L. Linking heterochromatin protein 1 (HP1) to cancer progression. Mutat. Res. 2008, 647, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.A.; Shilatifard, A. Chromatin signatures of cancer. Genes Dev. 2015, 29, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Kloc, A.; Zaratiegui, M.; Nora, E.; Martienssen, R. RNA interference guides histone modification during the S phase of chromosomal replication. Curr. Biol. 2008, 18, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.S.; Zhang, K.; Nicolas, E.; Cam, H.P.; Zofall, M.; Grewal, S.I. Cell cycle control of centromeric repeat transcription and heterochromatin assembly. Nature 2008, 451, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.T.; Takahashi, T.S.; Nakagawa, T.; Nakayama, J.; Masukata, H. The heterochromatin protein Swi6/HP1 activates replication origins at the pericentromeric region and silent mating-type locus. Nat. Cell Biol. 2009, 11, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Li, P.C.; Chretien, L.; Cote, J.; Kelly, T.J.; Forsburg, S.L.S. Pombe replication protein Cdc18 (Cdc6) interacts with Swi6 (HP1) heterochromatin protein: Region specific effects and replication timing in the centromere. Cell Cycle 2011, 10, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Dubey, D.D.; Huberman, J.A. Early-replicating heterochromatin. Genes Dev. 2003, 17, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Bernard, P.; Maure, J.F.; Partridge, J.F.; Genier, S.; Javerzat, J.P.; Allshire, R.C. Requirement of heterochromatin for cohesion at centromeres. Science 2001, 294, 2539–2542. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, N.; Kitajima, T.; Shihori, Y.; Xiao, G.; Yamamoto, M.; Grewal, S.I.; Watanabe, Y. Recruitment of cohesin to heterochromatic regions by Swi6/HP1 in fission yeast. Nat. Cell Biol. 2002, 4, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Bailis, J.M.; Bernard, P.; Antonelli, R.; Allshire, R.; Forsburg, S.L. Hsk1/Dfp1 is required for heterochromatin-mediated cohesion at centromeres. Nat. Cell Biol. 2003, 5, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; O’Donnell, M. The eukaryotic replication machine. Enzymes 2016, 39, 191–229. [Google Scholar] [PubMed]
- Hills, S.A.; Diffley, J.F. DNA replication and oncogene-induced replicative stress. Curr. Biol. 2014, 24, R435–R444. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Araki, H. Regulation of the initiation step of DNA replication by cyclin-dependent kinases. Chromosoma 2010, 119, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Masai, H. Regulation of chromosome dynamics by hsk1/cdc7 kinase. Biochem. Soc. Trans. 2013, 41, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Labib, K.; Gambus, A. A key role for the GINS complex at DNA replication forks. Trends Cell Biol. 2007, 17, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Shi, Y.; Georgescu, R.E.; Yuan, Z.; Chait, B.T.; Li, H.; O’Donnell, M.E. The architecture of a eukaryotic replisome. Nat. Struct. Mol. Biol. 2015, 22, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Langston, L.D.; Zhang, D.; Yurieva, O.; Georgescu, R.E.; Finkelstein, J.; Yao, N.Y.; Indiani, C.; O’Donnell, M.E. CMG helicase and DNA polymerase epsilon form a functional 15-subunit holoenzyme for eukaryotic leading-strand DNA replication. Proc. Natl. Acad. Sci. USA 2014, 111, 15390–15395. [Google Scholar] [CrossRef] [PubMed]
- Sabatinos, S.A.; Forsburg, S.L. Managing single-stranded DNA during replication stress in fission yeast. Biomolecules 2015, 5, 2123–2139. [Google Scholar] [CrossRef] [PubMed]
- Alcasabas, A.A.; Osborn, A.J.; Bachant, J.; Hu, F.; Werler, P.J.; Bousset, K.; Furuya, K.; Diffley, J.F.; Carr, A.M.; Elledge, S.J. MRC1 transduces signals of DNA replication stress to activate rad53. Nat. Cell Biol. 2001, 3, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Russell, P. MRC1 channels the DNA replication arrest signal to checkpoint kinase Cds1. Nat. Cell Biol. 2001, 3, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Leman, A.R.; Noguchi, E. Local and global functions of Timeless and Tipin in replication fork protection. Cell Cycle 2012, 11, 3945–3955. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.C.; Zhou, J.C.; Perera, R.L.; van Deursen, F.; Evrin, C.; Ivanova, M.E.; Kilkenny, M.L.; Renault, L.; Kjaer, S.; Matak-Vinkovic, D.; et al. A ctf4 trimer couples the CMG helicase to DNA polymerase alpha in the eukaryotic replisome. Nature 2014, 510, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Villa, F.; Simon, A.C.; Ortiz Bazan, M.A.; Kilkenny, M.L.; Wirthensohn, D.; Wightman, M.; Matak-Vinkovic, D.; Pellegrini, L.; Labib, K. Ctf4 is a hub in the eukaryotic replisome that links multiple CIP-Box proteins to the cmg helicase. Mol. Cell 2016, 63, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Zeman, M.K.; Cimprich, K.A. Causes and consequences of replication stress. Nat. Cell Biol. 2014, 16, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Debatisse, M.; le Tallec, B.; Letessier, A.; Dutrillaux, B.; Brison, O. Common fragile sites: Mechanisms of instability revisited. Trends Genet. 2012, 28, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.M.; Lambert, S. Replication stress-induced genome instability: The dark side of replication maintenance by homologous recombination. J. Mol. Biol. 2013, 425, 4733–4744. [Google Scholar] [CrossRef] [PubMed]
- Macheret, M.; Halazonetis, T.D. DNA replication stress as a hallmark of cancer. Annu. Rev. Pathol. 2015, 10, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Marechal, A.; Zou, L. Rpa-coated single-stranded DNA as a platform for post-translational modifications in the DNA damage response. Cell Res. 2015, 25, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Byun, T.S.; Pacek, M.; Yee, M.C.; Walter, J.C.; Cimprich, K.A. Functional uncoupling of MCM helicase and DNA polymerase activities activates the ATR-dependent checkpoint. Genes Dev. 2005, 19, 1040–1052. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.; Foiani, M.; Sogo, J.M. Multiple mechanisms control chromosome integrity after replication fork uncoupling and restart at irreparable UV lesions. Mol. Cell 2006, 21, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Sun, L.; Shen, F.; Chen, Y.; Hua, Y.; Liu, Y.; Zhang, M.; Hu, Y.; Wang, Q.; Xu, W.; et al. The intra-S phase checkpoint targets DNA2 to prevent stalled replication forks from reversing. Cell 2012, 149, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Ira, G.; Pellicioli, A.; Balijja, A.; Wang, X.; Fiorani, S.; Carotenuto, W.; Liberi, G.; Bressan, D.; Wan, L.; Hollingsworth, N.M.; et al. DNA end resection, homologous recombination and DNA damage checkpoint activation require cdk1. Nature 2004, 431, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Sofueva, S.; Osman, F.; Lorenz, A.; Steinacher, R.; Castagnetti, S.; Ledesma, J.; Whitby, M.C. Ultrafine anaphase bridges, broken DNA and illegitimate recombination induced by a replication fork barrier. Nucleic Acids Res. 2011, 39, 6568–6584. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.L.; Palmai-Pallag, T.; Ying, S.; Hickson, I.D. Replication stress induces sister-chromatid bridging at fragile site loci in mitosis. Nat. Cell Biol 2009, 11, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Sterling, J.; Thompson, C.; Harris, S.; Mav, D.; Shah, R.; Klimczak, L.J.; Kryukov, G.V.; Malc, E.; Mieczkowski, P.A.; et al. Clustered mutations in yeast and in human cancers can arise from damaged long single-strand DNA regions. Mol. Cell 2012, 46, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Fanning, E.; Klimovich, V.; Nager, A.R. A dynamic model for replication protein a (RPA) function in DNA processing pathways. Nucleic Acids Res. 2006, 34, 4126–4137. [Google Scholar] [CrossRef] [PubMed]
- Toledo, L.I.; Altmeyer, M.; Rask, M.B.; Lukas, C.; Larsen, D.H.; Povlsen, L.K.; Bekker-Jensen, S.; Mailand, N.; Bartek, J.; Lukas, J. ATR prohibits replication catastrophe by preventing global exhaustion of RPA. Cell 2013, 155, 1088–1103. [Google Scholar] [CrossRef] [PubMed]
- Branzei, D.; Foiani, M. The checkpoint response to replication stress. DNA Repair (Amst) 2009, 8, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Branzei, D.; Foiani, M. Maintaining genome stability at the replication fork. Nat. Rev. Mol. Cell Biol. 2010, 11, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Petermann, E.; Helleday, T. Pathways of mammalian replication fork restart. Nat. Rev. Mol. Cell Biol. 2010, 11, 683–687. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, D.; Blow, J.J. Dormant origins, the licensing checkpoint, and the response to replicative stresses. Cold Spring Harb. Perspect. Biol. 2012, 4, a012955. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Miyabe, I.; Schalbetter, S.A.; Carr, A.M.; Murray, J.M. Recombination-restarted replication makes inverted chromosome fusions at inverted repeats. Nature 2013, 493, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Minocherhomji, S.; Ying, S.; Bjerregaard, V.A.; Bursomanno, S.; Aleliunaite, A.; Wu, W.; Mankouri, H.W.; Shen, H.; Liu, Y.; Hickson, I.D. Replication stress activates DNA repair synthesis in mitosis. Nature 2015, 528, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Bachant, J.; Collingwood, D.; Raghuraman, M.K.; Brewer, B.J. Centromere replication timing determines different forms of genomic instability in saccharomyces cerevisiae checkpoint mutants during replication stress. Genetics 2009, 183, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Sabatinos, S.A.; Ranatunga, N.S.; Yuan, J.P.; Green, M.D.; Forsburg, S.L. Replication stress in early S phase generates apparent micronuclei and chromosome rearrangement in fission yeast. Mol. Biol. Cell 2015, 26, 3439–3450. [Google Scholar] [CrossRef] [PubMed]
- Allshire, R.C.; Karpen, G.H. Epigenetic regulation of centromeric chromatin: Old dogs, new tricks? Nat. Rev. Genet. 2008, 9, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Wood, V.; Gwilliam, R.; Rajandream, M.A.; Lyne, M.; Lyne, R.; Stewart, A.; Sgouros, J.; Peat, N.; Hayles, J.; Baker, S.; et al. The genome sequence of schizosaccharomyces pombe. Nature 2002, 415, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, J.; Rice, J.C.; Strahl, B.D.; Allis, C.D.; Grewal, S.I. Role of histone H3 lysine 9 methylation in epigenetic control of heterochromatin assembly. Science 2001, 292, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Rea, S.; Eisenhaber, F.; O’Carroll, D.; Strahl, B.D.; Sun, Z.W.; Schmid, M.; Opravil, S.; Mechtler, K.; Ponting, C.P.; Allis, C.D.; et al. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature 2000, 406, 593–599. [Google Scholar] [PubMed]
- Noma, K.; Sugiyama, T.; Cam, H.; Verdel, A.; Zofall, M.; Jia, S.; Moazed, D.; Grewal, S.I. Rits acts in Cis to promote RNA interference-mediated transcriptional and post-transcriptional silencing. Nat. Genet. 2004, 36, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Verdel, A.; Jia, S.; Gerber, S.; Sugiyama, T.; Gygi, S.; Grewal, S.I.; Moazed, D. RNAi-mediated targeting of heterochromatin by the RITS complex. Science 2004, 303, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Schalch, T.; Job, G.; Shanker, S.; Partridge, J.F.; Joshua-Tor, L. The Chp1-Tas3 core is a multifunctional platform critical for gene silencing by RITS. Nat. Struct. Mol. Biol. 2011, 18, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Schalch, T.; Job, G.; Noffsinger, V.J.; Shanker, S.; Kuscu, C.; Joshua-Tor, L.; Partridge, J.F. High-affinity binding of Chp1 chromodomain to K9 methylated histone H3 is required to establish centromeric heterochromatin. Mol. Cell 2009, 34, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.; Ishida, M.; Kawaguchi, R.; Urano, T.; Murakami, Y.; Nakayama, J. Heterochromatin protein 1 homologue swi6 acts in concert with Ers1 to regulate rnai-directed heterochromatin assembly. Proc. Natl. Acad. Sci. USA 2012, 109, 6159–6164. [Google Scholar] [CrossRef] [PubMed]
- Sadaie, M.; Iida, T.; Urano, T.; Nakayama, J. A chromodomain protein, Chp1, is required for the establishment of heterochromatin in fission yeast. EMBO J. 2004, 23, 3825–3835. [Google Scholar] [CrossRef] [PubMed]
- Debeauchamp, J.L.; Moses, A.; Noffsinger, V.J.; Ulrich, D.L.; Job, G.; Kosinski, A.M.; Partridge, J.F. Chp1-tas3 interaction is required to recruit RITS to fission yeast centromeres and for maintenance of centromeric heterochromatin. Mol. Cell. Biol. 2008, 28, 2154–2166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Mosch, K.; Fischle, W.; Grewal, S.I. Roles of the clr4 methyltransferase complex in nucleation, spreading and maintenance of heterochromatin. Nat. Struct. Mol. Biol. 2008, 15, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Hall, I.M.; Shankaranarayana, G.D.; Noma, K.; Ayoub, N.; Cohen, A.; Grewal, S.I. Establishment and maintenance of a heterochromatin domain. Science 2002, 297, 2232–2237. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Sonoda, M. Self-interaction of heterochromatin protein 1 is required for direct binding to histone methyltransferase, SUV39H1. Biochem. Biophys. Res. Commun. 2003, 301, 287–292. [Google Scholar] [CrossRef]
- Li, F.; Martienssen, R.; Cande, W.Z. Coordination of DNA replication and histone modification by the rik1-dos2 complex. Nature 2011, 475, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, E.; Bayne, E.H.; Allshire, R.C. On the connection between RNAi and heterochromatin at centromeres. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Doe, C.L.; Wang, G.; Chow, C.-M.; Fricker, M.D.; Singh, P.B.; Mellor, E.J. The fission yeast chromo domain encoding gene chp1+ is required for chromosome segregation and shows a genetic interaction with alpha-tubulin. Nucleic Acids Res. 1998, 26, 4222–4229. [Google Scholar] [CrossRef] [PubMed]
- Ekwall, K.; Javerzat, J.P.; Lorentz, A.; Schmidt, H.; Cranston, G.; Allshire, R. The chromodomain protein Swi6: A key component at fission yeast centromeres. Science 1995, 269, 1429–1431. [Google Scholar] [CrossRef] [PubMed]
- Ekwall, K.; Nimmo, E.R.; Javerzat, J.P.; Borgstrom, B.; Egel, R.; Cranston, G.; Allshire, R. Mutations in the fission yeast silencing factors Clr4+ and Rik1+ disrupt the localisation of the chromo domain protein Swi6p and impair centromere function. J. Cell Sci. 1996, 109, 2637–2648. [Google Scholar] [PubMed]
- Thon, G.; Verhein-Hansen, J. Four chromo-domain proteins of schizosaccharmyces pombe differentially repress transcription at various chromosomal locations. Genetics 2000, 155, 551–568. [Google Scholar] [PubMed]
- Li, P.C.; Green, M.D.; Forsburg, S.L. Mutations disrupting histone methylation have different effects on replication timing in S. Pombe centromere. PLoS ONE 2013, 8, e61464. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, J.G.; Bulboaca, G.H.; Moghadam, M.; Caddle, M.S.; Calos, M.P. Physical mapping of origins of replication in the fission yeast schizosaccharomyces pombe. Mol. Biol. Cell 1994, 5, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Zaratiegui, M.; Castel, S.E.; Irvine, D.V.; Kloc, A.; Ren, J.; Li, F.; de Castro, E.; Marin, L.; Chang, A.Y.; Goto, D.; et al. RNAi promotes heterochromatic silencing through replication-coupled release of RNA pol ii. Nature 2011, 479, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, J.; Allshire, R.C.; Klar, A.J.; Grewal, S.I. A role for DNA polymerase alpha in epigenetic control of transcriptional silencing in fission yeast. EMBO J. 2001, 20, 2857–2866. [Google Scholar] [CrossRef] [PubMed]
- Dohke, K.; Miyazaki, S.; Tanaka, K.; Urano, T.; Grewal, S.I.; Murakami, Y. Fission yeast chromatin assembly factor 1 assists in the replication-coupled maintenance of heterochromatin. Genes Cells 2008, 13, 1027–1043. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, M.R.; Hong, E.J.; Li, X.; Gerber, S.; Denison, C.; Gygi, S.; Moazed, D. Hp1 proteins form distinct complexes and mediate heterochromatic gene silencing by nonoverlapping mechanisms. Mol. Cell 2008, 32, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Cam, H.; Verdel, A.; Moazed, D.; Grewal, S.I. RNA-dependent RNA polymerase is an essential component of a self-enforcing loop coupling heterochromatin assembly to sirna production. Proc. Natl. Acad. Sci. USA 2005, 102, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Forsburg, S.L. The CINS of the centromere. Biochem. Soc. Trans. 2013, 41, 1706–1711. [Google Scholar] [CrossRef] [PubMed]
- Natsume, T.; Tsutsui, Y.; Sutani, T.; Dunleavy, E.M.; Pidoux, A.L.; Iwasaki, H.; Shirahige, K.; Allshire, R.C.; Yamao, F. A DNA polymerase alpha accessory protein, Mcl1, is required for propagation of centromere structures in fission yeast. PLoS ONE 2008, 3, e2221. [Google Scholar] [CrossRef] [PubMed]
- Bachant, J.; Jessen, S.R.; Kavanaugh, S.E.; Fielding, C.S. The yeast s phase checkpoint enables replicating chromosomes to bi-orient and restrain spindle extension during s phase distress. J. Cell Biol. 2005, 168, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Natsume, T.; Muller, C.A.; Katou, Y.; Retkute, R.; Gierlinski, M.; Araki, H.; Blow, J.J.; Shirahige, K.; Nieduszynski, C.A.; Tanaka, T.U. Kinetochores coordinate pericentromeric cohesion and early DNA replication by Cdc7-Dbf4 kinase recruitment. Mol. Cell 2013, 50, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Tsao, S.W.; Guan, X.Y.; Cheung, A.L. Pericentromeric regions are refractory to prompt repair after replication stress-induced breakage in HPV16 E6E7-expressing epithelial cells. PLoS ONE 2012, 7, e48576. [Google Scholar] [CrossRef] [PubMed]
- Simi, S.; Simili, M.; Bonatti, S.; Campagna, M.; Abbondandolo, A. Fragile sites at the centromere of chinese hamster chromosomes: A possible mechanism of chromosome loss. Mutat. Res. 1998, 397, 239–246. [Google Scholar] [CrossRef]
- Rozenzhak, S.; Mejia-Ramirez, E.; Williams, J.S.; Schaffer, L.; Hammond, J.A.; Head, S.R.; Russell, P. Rad3 decorates critical chromosomal domains with gammah2a to protect genome integrity during S-phase in fission yeast. PLoS Genet. 2010, 6, e1001032. [Google Scholar] [CrossRef] [PubMed]
- Burma, S.; Chen, B.P.; Murphy, M.; Kurimasa, A.; Chen, D.J. Atm phosphorylates histone H2AX in response to DNA double-strand breaks. J. Biol. Chem. 2001, 276, 42462–42467. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.M.; Du, L.L.; Redon, C.; Russell, P. Histone H2A phosphorylation controls Crb2 recruitment at DNA breaks, maintains checkpoint arrest, and influences DNA repair in fission yeast. Mol. Cell. Biol. 2004, 24, 6215–6230. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, A.; Ampatzidou, E.; Mizuno, K.; O’Connell, M.J.; Murray, J.M. Smc5/6 maintains stalled replication forks in a recombination-competent conformation. EMBO J. 2009, 28, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Pebernard, S.; Schaffer, L.; Campbell, D.; Head, S.R.; Boddy, M.N. Localization of Smc5/6 to centromeres and telomeres requires heterochromatin and sumo, respectively. Embo J. 2008, 27, 3011–3023. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Alveal, C.; O’Connell, M.J. Nse1-dependent recruitment of Smc5/6 to lesion-containing loci contributes to the repair defects of mutant complexes. Mol. Biol. Cell 2011, 22, 4669–4682. [Google Scholar] [CrossRef] [PubMed]
- Ampatzidou, E.; Irmisch, A.; O’Connell, M.J.; Murray, J.M. Smc5/6 is required for repair at collapsed replication forks. Mol. Cell. Biol. 2006, 26, 9387–9401. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Rozenzhak, S.; Russell, P. Gammah2a-binding protein Brc1 affects centromere function in fission yeast. Mol. Cell. Biol. 2013, 33, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, R.M.; Clarke, L.; Carbon, J. Clustered tRNA genes in schizosaccharomyces pombe centromeric DNA sequence repeats. Proc. Natl. Acad. Sci. USA 1991, 88, 1306–1310. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, O.; Tanaka, A.; Tanizawa, H.; Grewal, S.I.; Noma, K. Centromeric localization of dispersed pol III genes in fission yeast. Mol. Biol. Cell 2010, 21, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.C.; Merrett, S.L.; Willard, H.F. A heterochromatin barrier partitions the fission yeast centromere into discrete chromatin domains. Curr. Biol. 2006, 16, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.M.; Newlon, C.S. Dna replication fork pause sites dependent on transcription. Science 1996, 272, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, I.; Taddei, A. Linking replication stress with heterochromatin formation. Chromosoma 2016, 125, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Greenfeder, S.A.; Newlon, C.S. Replication forks pause at yeast centromeres. Mol. Cell. Biol. 1992, 12, 4056–4066. [Google Scholar] [CrossRef] [PubMed]
- Baum, M.; Clarke, L. Fission yeast homologs of human Cenp-B have redundant functions affecting cell growth and chromosome segregation. Mol. Cell. Biol. 2000, 20, 2852–2864. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Lee, J.K.; Hurwitz, J.; Allshire, R.C.; Nakayama, J.; Grewal, S.I.; Tanaka, K.; Murakami, Y. Fission yeast Cenp-B homologs nucleate centromeric heterochromatin by promoting heterochromatin-specific histone tail modifications. Genes Dev. 2002, 16, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Casola, C.; Hucks, D.; Feschotte, C. Convergent domestication of pogo-like transposases into centromere-binding proteins in fission yeast and mammals. Mol. Biol. Evol. 2008, 25, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Huberman, J.A.; Hurwitz, J. Identification, purification, and molecular cloning of autonomously replicating sequence-binding protein 1 from fission yeast schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1996, 93, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Irelan, J.T.; Gutkin, G.I.; Clarke, L. Functional redundancies, distinct localizations and interactions among three fission yeast homologs of centromere protein-B. Genetics 2001, 157, 1191–1203. [Google Scholar] [PubMed]
- Lee, J.K.; Huberman, J.A.; Hurwitz, J. Purification and characterization of a Cenp-B homologue protein that binds to the centromeric k-type repeat DNA of schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1997, 94, 8427–8432. [Google Scholar] [CrossRef] [PubMed]
- Zaratiegui, M.; Vaughn, M.W.; Irvine, D.V.; Goto, D.; Watt, S.; Bahler, J.; Arcangioli, B.; Martienssen, R.A. Cenp-B preserves genome integrity at replication forks paused by retrotransposon ltr. Nature 2011, 469, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Johansen, P.; Cam, H.P. Suppression of meiotic recombination by Cenp-B homologs in schizosaccharomyces pombe. Genetics 2015, 201, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Krings, G.; Bastia, D. Sap1p binds to TER1 at the ribosomal DNA of schizosaccharomyces pombe and causes polar replication fork arrest. J. Biol. Chem. 2005, 280, 39135–39142. [Google Scholar] [CrossRef] [PubMed]
- Mejia-Ramirez, E.; Sanchez-Gorostiaga, A.; Krimer, D.B.; Schvartzman, J.B.; Hernandez, P. The mating type switch-activating protein Sap1 is required for replication fork arrest at the rRNA genes of fission yeast. Mol. Cell. Biol. 2005, 25, 8755–8761. [Google Scholar] [CrossRef] [PubMed]
- Arcangioli, B.; Copeland, T.D.; Klar, A.J.S. Sap1, a protein that binds to sequences required for mating-type switching, is essential for viability in schizosaccharomyces pombe. Mol. Cell. Biol. 1994, 14, 2058–2065. [Google Scholar] [CrossRef] [PubMed]
- De Lahondes, R.; Ribes, V.; Arcangioli, B. Fission yeast Sap1 protein is essential for chromosome stability. Eukaryot Cell. 2003, 2, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, C.; Noguchi, E. Sap1 promotes the association of the replication fork protection complex with chromatin and is involved in the replication checkpoint in schizosaccharomyces pombe. Genetics 2007, 175, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, M.C.; Das, M.M.; Tanizawa, H.; Chang, Y.T.; Noma, K.; Nakamura, T.M.; Noguchi, E. Swi1timeless prevents repeat instability at fission yeast telomeres. PLoS Genet. 2016, 12, e1005943. [Google Scholar] [CrossRef] [PubMed]
- Nakaseko, Y.; Kinoshita, N.; Yanagida, M. A novel sequence common to the centromere regions of schizosaccharomyces pombe chromosomes. Nucleic Acids Res. 1987, 15, 4705–4715. [Google Scholar] [CrossRef] [PubMed]
- Ellermeier, C.; Higuchi, E.C.; Phadnis, N.; Holm, L.; Geelhood, J.L.; Thon, G.; Smith, G.R. RNAi and heterochromatin repress centromeric meiotic recombination. Proc. Natl. Acad. Sci. USA 2010, 107, 8701–8705. [Google Scholar] [CrossRef] [PubMed]
- Vincenten, N.; Kuhl, L.M.; Lam, I.; Oke, A.; Kerr, A.R.; Hochwagen, A.; Fung, J.; Keeney, S.; Vader, G.; Marston, A.L. The kinetochore prevents centromere-proximal crossover recombination during meiosis. Elife 2015, 4, e10850. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Murakami, S.; Chikashige, Y.; Funabiki, H.; Niwa, O.; Yanagida, M. A low copy number central sequence with strict symmetry and unusual chromatin structure in fission yeast centromere. Mol. Biol. Cell 1992, 3, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Pidoux, A.L.; Allshire, R.C. The role of heterochromatin in centromere function. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 569–579. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, R.J.; Humphrey, T.C. A role for recombination in centromere function. Trends Genet. 2010, 26, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Tinline-Purvis, H.; Savory, A.P.; Cullen, J.K.; Dave, A.; Moss, J.; Bridge, W.L.; Marguerat, S.; Bahler, J.; Ragoussis, J.; Mott, R.; et al. Failed gene conversion leads to extensive end processing and chromosomal rearrangements in fission yeast. EMBO J. 2009, 28, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Okamoto, A.; Katou, Y.; Yadani, C.; Shitanda, T.; Kaweeteerawat, C.; Takahashi, T.S.; Itoh, T.; Shirahige, K.; Masukata, H.; et al. Rad51 suppresses gross chromosomal rearrangement at centromere in schizosaccharomyces pombe. EMBO J. 2008, 27, 3036–3046. [Google Scholar] [CrossRef] [PubMed]
- Onaka, A.T.; Toyofuku, N.; Inoue, T.; Okita, A.K.; Sagawa, M.; Su, J.; Shitanda, T.; Matsuyama, R.; Zafar, F.; Takahashi, T.S.; et al. Rad51 and Rad54 promote noncrossover recombination between centromere repeats on the same chromatid to prevent isochromosome formation. Nucleic Acids Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Naim, V.; Wilhelm, T.; Debatisse, M.; Rosselli, F. Ercc1 and Mus81-Eme1 promote sister chromatid separation by processing late replication intermediates at common fragile sites during mitosis. Nat. Cell Biol. 2013, 15, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Minocherhomji, S.; Chan, K.L.; Palmai-Pallag, T.; Chu, W.K.; Wass, T.; Mankouri, H.W.; Liu, Y.; Hickson, I.D. Mus81 promotes common fragile site expression. Nat. Cell Biol. 2013, 15, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Roguev, A.; Bandyopadhyay, S.; Zofall, M.; Zhang, K.; Fischer, T.; Collins, S.R.; Qu, H.; Shales, M.; Park, H.O.; Hayles, J.; et al. Conservation and rewiring of functional modules revealed by an epistasis map in fission yeast. Science 2008, 322, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. Chromosome dynamics during mitosis. Cold Spring Harb. Perspect. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Haarhuis, J.H.; Elbatsh, A.M.; Rowland, B.D. Cohesin and its regulation: On the logic of X-shaped chromosomes. Dev. Cell 2014, 31, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K.; Haering, C.H. Cohesin: Its roles and mechanisms. Annu. Rev. Genet. 2009, 43, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Ansbach, A.B.; Noguchi, C.; Klansek, I.W.; Heidlebaugh, M.; Nakamura, T.M.; Noguchi, E. Rfcctf18 and the swi1-swi3 complex function in separate and redundant pathways required for the stabilization of replication forks to facilitate sister chromatid cohesion in schizosaccharomyces pombe. Mol. Biol. Cell 2008, 19, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.R.; McIntosh, J.R. Mcl1+, the schizosaccharomyces pombe homologue of Ctf4, is important for chromosome replication, cohesion, and segregation. Eukaryot. Cell 2002, 1, 758–773. [Google Scholar] [CrossRef] [PubMed]
- Guillou, E.; Ibarra, A.; Coulon, V.; Casado-Vela, J.; Rico, D.; Casal, I.; Schwob, E.; Losada, A.; Mendez, J. Cohesin organizes chromatin loops at DNA replication factories. Genes Dev. 2010, 24, 2812–2822. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, A.K.; Zhang, N.; Otta, S.K.; Pati, D. A cohesin-rad21 interactome. Biochem. J. 2012, 442, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Unal, E.; Heidinger-Pauli, J.M.; Koshland, D. DNA double-strand breaks trigger genome-wide sister-chromatid cohesion through eco1 (ctf7). Science 2007, 317, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Unal, E.; Arbel-Eden, A.; Sattler, U.; Shroff, R.; Lichten, M.; Haber, J.E.; Koshland, D. DNA damage response pathway uses histone modification to assemble a double-strand break-specific cohesin domain. Mol. Cell 2004, 16, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.J.; Kim, B.J.; Lee, J.W.; Lee, M.W.; Choi, H.K.; Kim, S.T. Direct interaction between cohesin complex and DNA replication machinery. Biochem. Biophys. Res. Commun. 2006, 341, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, Y.; Sakuno, T.; Shimura, M.; Watanabe, Y. Heterochromatin links to centromeric protection by recruiting shugoshin. Nature 2008, 455, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Baxter, J. “Breaking up is hard to do”: The formation and resolution of sister chromatid intertwines. J. Mol. Biol. 2015, 427, 590–607. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Nielsen, C.F.; Yao, Q.; Hickson, I.D. The origins and processing of ultra fine anaphase DNA bridges. Curr. Opin. Genet. Dev. 2014, 26, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Burrell, R.A.; McClelland, S.E.; Endesfelder, D.; Groth, P.; Weller, M.C.; Shaikh, N.; Domingo, E.; Kanu, N.; Dewhurst, S.M.; Gronroos, E.; et al. Replication stress links structural and numerical cancer chromosomal instability. Nature 2013, 494, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Farcas, A.M.; Uluocak, P.; Helmhart, W.; Nasmyth, K. Cohesin’s concatenation of sister dnas maintains their intertwining. Mol. Cell 2011, 44, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Mayer, B.; Stemmann, O.; Nigg, E.A. Centromere DNA decatenation depends on cohesin removal and is required for mammalian cell division. J. Cell Sci. 2010, 123, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.A.; Hamilton, R.S.; Pauli, A.; Davis, I.; Nasmyth, K. Cohesin cleavage and cdk inhibition trigger formation of daughter nuclei. Nat. Cell Biol. 2010, 12, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Haarhuis, J.H.; Elbatsh, A.M.; van den Broek, B.; Camps, D.; Erkan, H.; Jalink, K.; Medema, R.H.; Rowland, B.D. Wapl-mediated removal of cohesin protects against segregation errors and aneuploidy. Curr. Biol. 2013, 23, 2071–2077. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Leonard, J.; Torres, R.; Garcia-Luis, J.; Palou-Marin, G.; Aragon, L. Physical proximity of sister chromatids promotes top2-dependent intertwining. Mol. Cell 2016, 64, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Charbin, A.; Bouchoux, C.; Uhlmann, F. Condensin aids sister chromatid decatenation by topoisomerase ii. Nucleic Acids Res. 2014, 42, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Coelho, P.A.; Queiroz-Machado, J.; Sunkel, C.E. Condensin-dependent localisation of topoisomerase ii to an axial chromosomal structure is required for sister chromatid resolution during mitosis. J. Cell Sci. 2003, 116, 4763–4776. [Google Scholar] [CrossRef] [PubMed]
- Tittel-Elmer, M.; Lengronne, A.; Davidson, M.B.; Bacal, J.; Francois, P.; Hohl, M.; Petrini, J.H.; Pasero, P.; Cobb, J.A. Cohesin association to replication sites depends on rad50 and promotes fork restart. Mol. Cell 2012, 48, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, C.; Nasmyth, K. Sister chromatid cohesion is required for postreplicative double-strand break repair in saccharomyces cerevisiae. Curr. Biol. 2001, 11, 991–995. [Google Scholar] [CrossRef]
- Tatebayashi, K.; Kato, J.; Ikeda, H. Isolation of a schizosaccharomyces pombe rad21ts mutant that is aberrant in chromosome segregation, microtubule function, DNA repair and sensitive to hydroxyurea: Possible involvement of rad21 in ubiquitin-mediated proteolysis. Genetics 1998, 148, 49–57. [Google Scholar] [PubMed]
- Birkenbihl, R.P.; Subramani, S. Cloning and characterization of rad21 an essential gene of schizosaccharomyces pombe involved in DNA double-strand-break repair. Nucliec Acids Res. 1992, 20, 6605–6611. [Google Scholar] [CrossRef]
- Cortes-Ledesma, F.; Aguilera, A. Double-strand breaks arising by replication through a nick are repaired by cohesin-dependent sister-chromatid exchange. EMBO Rep. 2006, 7, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Gelot, C.; Guirouilh-Barbat, J.; Lopez, B.S. The cohesin complex prevents the end-joining of distant DNA double-strand ends in s phase: Consequences on genome stability maintenance. Nucleus 2016, 7, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Hamperl, S.; Cimprich, K.A. Conflict resolution in the genome: How transcription and replication make it work. Cell 2016, 167, 1455–1467. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Muse, T.; Aguilera, A. Transcription-replication conflicts: How they occur and how they are resolved. Nat. Rev. Mol. Cell Biol. 2016, 17, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Furuya, K.; Takahashi, K.; Yanagida, M. Faithful anaphase is endured by Mis4, a sister chromatid cohesion molecule required in sphase and not destroyed N G1 phase. Genes Dev. 1998, 12, 3408–3418. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, J.Z.; Klar, A.J.S. Swi1 and swi3 perform imprinting, pausing and termination of DNA replication in S. Pombe. Cell 2000, 102, 745–751. [Google Scholar] [CrossRef]
- Krings, G.; Bastia, D. Swi1- and Swi3-dependent and independent replication fork arrest at the ribosomal DNA of schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 2004, 101, 14085–14090. [Google Scholar] [CrossRef] [PubMed]
- Erliandri, I.; Fu, H.; Nakano, M.; Kim, J.H.; Miga, K.H.; Liskovykh, M.; Earnshaw, W.C.; Masumoto, H.; Kouprina, N.; Aladjem, M.I.; et al. Replication of alpha-satellite DNA arrays in endogenous human centromeric regions and in human artificial chromosome. Nucleic Acids Res. 2014, 42, 11502–11516. [Google Scholar] [CrossRef] [PubMed]
- Aze, A.; Sannino, V.; Soffientini, P.; Bachi, A.; Costanzo, V. Centromeric DNA replication reconstitution reveals DNA loops and ATR checkpoint suppression. Nat. Cell Biol. 2016, 18, 684–691. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forsburg, S.L.; Shen, K.-F. Centromere Stability: The Replication Connection. Genes 2017, 8, 37. https://doi.org/10.3390/genes8010037
Forsburg SL, Shen K-F. Centromere Stability: The Replication Connection. Genes. 2017; 8(1):37. https://doi.org/10.3390/genes8010037
Chicago/Turabian StyleForsburg, Susan L., and Kuo-Fang Shen. 2017. "Centromere Stability: The Replication Connection" Genes 8, no. 1: 37. https://doi.org/10.3390/genes8010037
APA StyleForsburg, S. L., & Shen, K.-F. (2017). Centromere Stability: The Replication Connection. Genes, 8(1), 37. https://doi.org/10.3390/genes8010037