
Transcription Regulation of the Human Telomerase Reverse Transcriptase (hTERT) Gene


Abstract
1. Introduction
2. Trans-Acting Regulators of hTERT Transcription
2.1. Transcription Activators of hTERT
2.1.1. c-Myc
2.1.2. NF-κB
2.1.3. STAT Proteins
2.1.4. Paired Box Proteins (Pax)
2.1.5. Estrogen Receptor
2.2. Transcription Repressors of hTERT
2.2.1. Mad1
2.2.2. Sp3
2.2.3. CTCF
2.2.4. E2F1
2.2.5. Hormone Nuclear Receptors
2.3. Transcription Regulators with Dual Roles
2.3.1. Specificity Protein 1 (Sp1)
2.3.2. Activator Protein 1 (AP-1)
2.3.3. Early growth response-1 (EGR-1)
2.3.4. Hypoxia-inducible factor 2-alpha (HIF-2α)
2.3.5. Kruppel-like family of transcription factors (KLF) Proteins
2.3.6. Nuclear Transcription Factor X Box-Binding Protein 1 (NFX1)
2.3.7. Upstream stimulatory factors (USF) Proteins
2.4. Epigenetic Modifiers Regulating hTERT Transcription
2.4.1. Histone Modifiers
2.4.2. Regulators of hTERT Promoter Methylation
2.5. Monoallelic Expression of hTERT Gene
2.6. Complexity of Trans-Regulation of hTERT Gene Transcription
3. Genetic Alterations Regulating hTERT Transcription
3.1. hTERT Promoter Mutations
3.1.1. hTERT Promoter Mutations in Different Types of Human Cancers
3.1.2. Mechanism by Which hTERT Promoter Mutations Leads to Enhanced Telomerase Levels
3.2. Chromosomal Rearrangements and hTERT Expression
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Blackburn, E.H. Telomere states and cell fates. Nature 2000, 408, 53–56. [Google Scholar] [CrossRef] [PubMed]
- De Lange, T. How shelterin solves the telomere end-protection problem. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Makarov, V.L.; Hirose, Y.; Langmore, J.P. Long G tails at both ends of human chromosomes suggest a C strand degradation mechanism for telomere shortening. Cell 1997, 88, 657–666. [Google Scholar] [CrossRef]
- Wright, W.E.; Tesmer, V.M.; Huffman, K.E.; Levene, S.D.; Shay, J.W. Normal human chromosomes have long G-rich telomeric overhangs at one end. Genes Dev. 1997, 11, 2801–2809. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Funk, W.D.; Wang, S.S.; Weinrich, S.L.; Avilion, A.A.; Chiu, C.P.; Adams, R.R.; Chang, E.; Allsopp, R.C.; Yu, J.; et al. The RNA component of human telomerase. Science 1995, 269, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Masutomi, K.; Yu, E.Y.; Khurts, S.; Ben-Porath, I.; Currier, J.L.; Metz, G.B.; Brooks, M.W.; Kaneko, S.; Murakami, S.; DeCaprio, J.A.; et al. Telomerase maintains telomere structure in normal human cells. Cell 2003, 114, 241–253. [Google Scholar] [CrossRef]
- Wright, W.E.; Piatyszek, M.A.; Rainey, W.E.; Byrd, W.; Shay, J.W. Telomerase activity in human germline and embryonic tissues and cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Levy, M.Z.; Allsopp, R.C.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere end-replication problem and cell aging. J. Mol. Biol. 1992, 225, 951–960. [Google Scholar] [CrossRef]
- De Lange, T.; Lundblad, V.; Blackburn, E.H. Telomeres, 2nd ed.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 2005; p. 576. [Google Scholar]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Takai, H.; Smogorzewska, A.; de Lange, T. DNA damage foci at dysfunctional telomeres. Curr. Biol. 2003, 13, 1549–1556. [Google Scholar] [CrossRef]
- d’Adda di Fagagna, F.; Reaper, P.M.; Clay-Farrace, L.; Fiegler, H.; Carr, P.; Von Zglinicki, T.; Saretzki, G.; Carter, N.P.; Jackson, S.P. A DNA damage checkpoint response in telomere-initiated senescence. Nature 2003, 426, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Kaul, Z.; Cesare, A.J.; Huschtscha, L.I.; Neumann, A.A.; Reddel, R.R. Five dysfunctional telomeres predict onset of senescence in human cells. EMBO Rep. 2012, 13, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Flores, I.; Canela, A.; Vera, E.; Tejera, A.; Cotsarelis, G.; Blasco, M.A. The longest telomeres: A general signature of adult stem cell compartments. Genes Dev. 2008, 22, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Artandi, S.E.; DePinho, R.A. Telomeres and telomerase in cancer. Carcinogenesis 2010, 31, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Schaetzlein, S.; Lucas-Hahn, A.; Lemme, E.; Kues, W.A.; Dorsch, M.; Manns, M.P.; Niemann, H.; Rudolph, K.L. Telomere length is reset during early mammalian embryogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 8034–8038. [Google Scholar] [CrossRef] [PubMed]
- Holohan, B.; Wright, W.E.; Shay, J.W. Cell biology of disease: Telomeropathies: An emerging spectrum disorder. J. Cell Biol. 2014, 205, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Armanios, M.; Blackburn, E.H. The telomere syndromes. Nat. Rev. Genet. 2012, 13, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Lansdorp, P.M. Telomeres and disease. EMBO J. 2009, 28, 2532–2540. [Google Scholar] [CrossRef] [PubMed]
- Blasco, M.A.; Lee, H.W.; Hande, M.P.; Samper, E.; Lansdorp, P.M.; DePinho, R.A.; Greider, C.W. Telomere shortening and tumor formation by mouse cells lacking telomerase rna. Cell 1997, 91, 25–34. [Google Scholar] [CrossRef]
- Lee, H.W.; Blasco, M.A.; Gottlieb, G.J.; Horner, J.W., II; Greider, C.W.; DePinho, R.A. Essential role of mouse telomerase in highly proliferative organs. Nature 1998, 392, 569–574. [Google Scholar] [PubMed]
- Chou, W.C.; Chen, H.Y.; Yu, S.L.; Cheng, L.; Yang, P.C.; Dang, C.V. Arsenic suppresses gene expression in promyelocytic leukemia cells partly through Sp1 oxidation. Blood 2005, 106, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Smith, L.L.; Coller, H.A.; Roberts, J.M. Telomerase modulates expression of growth-controlling genes and enhances cell proliferation. Nat. Cell Biol. 2003, 5, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.S.; Wen, J.; Bacchetti, S. The human telomerase catalytic subunit hTERT: Organization of the gene and characterization of the promoter. Hum. Mol. Genet. 1999, 8, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, I.; Cable, P.L.; Afshari, C.; Barrett, J.C. Cloning and characterization of the promoter region of human telomerase reverse transcriptase gene. Cancer Res. 1999, 59, 826–830. [Google Scholar] [PubMed]
- Takakura, M.; Kyo, S.; Kanaya, T.; Hirano, H.; Takeda, J.; Yutsudo, M.; Inoue, M. Cloning of human telomerase catalytic subunit (hTERT) gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res. 1999, 59, 551–557. [Google Scholar] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef] [PubMed]
- Moehren, U.; Papaioannou, M.; Reeb, C.A.; Grasselli, A.; Nanni, S.; Asim, M.; Roell, D.; Prade, I.; Farsetti, A.; Baniahmad, A. Wild-type but not mutant androgen receptor inhibits expression of the hTERT telomerase subunit: A novel role of AR mutation for prostate cancer development. FASEB J. 2008, 22, 1258–1267. [Google Scholar] [CrossRef] [PubMed]
- Takakura, M.; Kyo, S.; Inoue, M.; Wright, W.E.; Shay, J.W. Function of AP-1 in transcription of the telomerase reverse transcriptase gene (TERT) in human and mouse cells. Mol. Cell. Biol. 2005, 25, 8037–8043. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, R.T.; Degerman, S.; Ljungberg, B.; Andersson, E.; Oji, Y.; Sugiyama, H.; Roos, G.; Li, A. Wilms’ tumour 1 can suppress hTERT gene expression and telomerase activity in clear cell renal cell carcinoma via multiple pathways. Br. J. Cancer 2010, 103, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, A.S.; Villaudy, J.; Gazzolo, L.; Castellazzi, M.; Mesnard, J.M.; Duc Dodon, M. HTLV-1 HBZ cooperates with jund to enhance transcription of the human telomerase reverse transcriptase gene (hTERT). Retrovirology 2007. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.G.; Jayachandran, G.; Wu, G.; Xu, K.; Roth, J.A.; Ji, L. Tumor-specific activation of human telomerase reverses transcriptase promoter activity by activating enhancer-binding Protein-2β in human lung cancer cells. J. Biol. Chem. 2007, 282, 26460–26470. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Chen, W.; Guo, W.; Wang, J.; Tian, Y.; Shi, D.; Zhang, X.; Qiu, H.; Xiao, X.; Kang, T.; et al. Berberine targets AP-2/hTERT, NF-κB/COX-2, HIF-1α/VEGF and cytochrome-c/caspase signaling to suppress human cancer cell growth. PLoS ONE 2013, 8, e69240. [Google Scholar] [CrossRef] [PubMed]
- Suryo Rahmanto, Y.; Jung, J.G.; Wu, R.C.; Kobayashi, Y.; Heaphy, C.M.; Meeker, A.K.; Wang, T.L.; Shih Ie, M. Inactivating arid1a tumor suppressor enhances TERT transcription and maintains telomere length in cancer cells. J. Biol. Chem. 2016, 291, 9690–9699. [Google Scholar] [CrossRef] [PubMed]
- Hoffmeyer, K.; Raggioli, A.; Rudloff, S.; Anton, R.; Hierholzer, A.; Del Valle, I.; Hein, K.; Vogt, R.; Kemler, R. Wnt/β-catenin signaling regulates telomerase in stem cells and cancer cells. Science 2012, 336, 1549–1554. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Toh, L.; Lau, P.; Wang, X. Human telomerase reverse transcriptase (hTERT) is a novel target of the Wnt/β-catenin pathway in human cancer. J. Biol. Chem. 2012, 287, 32494–32511. [Google Scholar] [CrossRef] [PubMed]
- Isenmann, S.; Cakouros, D.; Zannettino, A.; Shi, S.; Gronthos, S. hTERT transcription is repressed by Cbfa1 in human mesenchymal stem cell populations. J. Bone Miner. Res. 2007, 22, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.J.; Grandori, C.; Amacker, M.; Simon-Vermot, N.; Polack, A.; Lingner, J.; Dalla-Favera, R. Direct activation of TERT transcription by c-Myc. Nat. Genet. 1999, 21, 220–224. [Google Scholar] [PubMed]
- Xu, D.; Popov, N.; Hou, M.; Wang, Q.; Bjorkholm, M.; Gruber, A.; Menkel, A.R.; Henriksson, M. Switch from Myc/Max to Mad1/Max binding and decrease in histone acetylation at the telomerase reverse transcriptase promoter during differentiation of HL60 cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3826–3831. [Google Scholar] [CrossRef] [PubMed]
- Ogretmen, B.; Kraveka, J.M.; Schady, D.; Usta, J.; Hannun, Y.A.; Obeid, L.M. Molecular mechanisms of ceramide-mediated telomerase inhibition in the a549 human lung adenocarcinoma cell line. J. Biol. Chem. 2001, 276, 32506–32514. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Taira, T.; Kanaya, T.; Itoh, H.; Yutsudo, M.; Ariga, H.; Inoue, M. Sp1 cooperates with c-Myc to activate transcription of the human telomerase reverse transcriptase gene (hTERT). Nucleic Acids Res. 2000, 28, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Veldman, T.; Liu, X.; Yuan, H.; Schlegel, R. Human papillomavirus E6 and Myc proteins associate in vivo and bind to and cooperatively activate the telomerase reverse transcriptase promoter. Proc. Natl. Acad. Sci. USA 2003, 100, 8211–8216. [Google Scholar] [CrossRef] [PubMed]
- Gewin, L.; Myers, H.; Kiyono, T.; Galloway, D.A. Identification of a novel telomerase repressor that interacts with the human papillomavirus type-16 E6/E6-AP complex. Genes Dev. 2004, 18, 2269–2282. [Google Scholar] [CrossRef] [PubMed]
- Faiola, F.; Liu, X.; Lo, S.; Pan, S.; Zhang, K.; Lymar, E.; Farina, A.; Martinez, E. Dual regulation of c-Myc by p300 via acetylation-dependent control of Myc protein turnover and coactivation of Myc-induced transcription. Mol. Cell. Biol. 2005, 25, 10220–10234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.W.; Chang, H.S.; Lin, C.H.; Yu, W.C. HPV-18 E7 conjugates to c-Myc and mediates its transcriptional activity. Int. J. Biochem. Cell Biol. 2007, 39, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Kanaya, T.; Zhuo, W.; Fujimoto, K.; Nishio, Y.; Orimo, A.; Inoue, M. Estrogen activates telomerase. Cancer Res. 1999, 59, 5917–5921. [Google Scholar] [PubMed]
- Xu, D.; Dwyer, J.; Li, H.; Duan, W.; Liu, J.P. Ets2 maintains hTERT gene expression and breast cancer cell proliferation by interacting with c-Myc. J. Biol. Chem. 2008, 283, 23567–23580. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.T.; Kyo, S.; Laimins, L.A. Telomerase activation by human papillomavirus type 16 E6 protein: Induction of human telomerase reverse transcriptase expression through Myc and GC-rich Sp1 binding sites. J. Virol. 2001, 75, 5559–5566. [Google Scholar] [CrossRef] [PubMed]
- McMurray, H.R.; McCance, D.J. Human papillomavirus type 16 E6 activates TERT gene transcription through induction of c-Myc and release of USF-mediated repression. J. Virol. 2003, 77, 9852–9861. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Dakic, A.; Zhang, Y.; Dai, Y.; Chen, R.; Schlegel, R. HPV E6 protein interacts physically and functionally with the cellular telomerase complex. Proc. Natl. Acad. Sci. USA 2009, 106, 18780–18785. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lee, T.H.; Avraham, H. A novel tricomplex of BRCA1, Nmi, and c-Myc inhibits c-Myc-induced human telomerase reverse transcriptase gene (hTERT) promoter activity in breast cancer. J. Biol. Chem. 2002, 277, 20965–20973. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.X.; Johansson, M.E.; Ren, J.; Xu, D.W.; Johnson, F.B.; Edfeldt, K.; Yan, Z.Q. Nuclear factor {kappa}B-mediated transactivation of telomerase prevents intimal smooth muscle cell from replicative senescence during vascular repair. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2604–2610. [Google Scholar] [CrossRef] [PubMed]
- Sinha-Datta, U.; Horikawa, I.; Michishita, E.; Datta, A.; Sigler-Nicot, J.C.; Brown, M.; Kazanji, M.; Barrett, J.C.; Nicot, C. Transcriptional activation of hTERT through the NF-κB pathway in HTLV-I-transformed cells. Blood 2004, 104, 2523–2531. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Ogawa, K.; Ikei, T.; Fujiki, T.; Katakura, Y. FOXO3a potentiates hTERT gene expression by activating c-Myc and extends the replicative life-span of human fibroblast. PLoS ONE 2014, 9, e101864. [Google Scholar] [CrossRef] [PubMed]
- Ponnala, S.; Chetty, C.; Veeravalli, K.K.; Dinh, D.H.; Klopfenstein, J.D.; Rao, J.S. MMP-9 silencing regulates hTERT expression via β1 integrin-mediated FAK signaling and induces senescence in glioma xenograft cells. Cell Signal 2011, 23, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ou, C.C.; Feldman, R.I.; Nicosia, S.V.; Kruk, P.A.; Cheng, J.Q. Aurora-a kinase regulates telomerase activity through c-Myc in human ovarian and breast epithelial cells. Cancer Res. 2004, 64, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Deng, X.; Deng, L.; Gu, H.; Fan, W.; Cao, Y. Telomerase activation by Epstein-Barr virus latent membrane protein 1 is associated with c-Myc expression in human nasopharyngeal epithelial cells. J. Exp. Clin. Cancer Res. 2004, 23, 495–506. [Google Scholar] [PubMed]
- Endoh, T.; Tsuji, N.; Asanuma, K.; Yagihashi, A.; Watanabe, N. Survivin enhances telomerase activity via up-regulation of specificity protein 1- and c-Myc-mediated human telomerase reverse transcriptase gene transcription. Exp. Cell Res. 2005, 305, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.Y.; Chen, Y.R.; Wang, T.C. A major role of PKC θ and NFκB in the regulation of hTERT in human T lymphocytes. FEBS Lett. 2006, 580, 6819–6824. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fang, X.; Ge, Z.; Jalink, M.; Kyo, S.; Bjorkholm, M.; Gruber, A.; Sjoberg, J.; Xu, D. The telomerase reverse transcriptase (hTERT) gene is a direct target of the histone methyltransferase SMYD3. Cancer Res. 2007, 67, 2626–2631. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Vorontchikhina, M.; Wang, Y.L.; Faiola, F.; Martinez, E. Staga recruits mediator to the Myc oncoprotein to stimulate transcription and cell proliferation. Mol. Cell. Biol. 2008, 28, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, Y.; Yang, H.; Cheng, J.Q.; Kruk, P.A. Pyk2/Erk 1/2 mediate Sp1- and c-Myc -dependent induction of telomerase activity by epidermal growth factor. Growth Factors 2008, 26, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, N.F.; Kocher, H.M.; Salako, M.A.; Obermueller, E.; Sandle, J.; Balkwill, F. A novel function of colony-stimulating factor 1 receptor in hTERT immortalization of human epithelial cells. Oncogene 2009, 28, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Bilsland, A.E.; Hoare, S.; Stevenson, K.; Plumb, J.; Gomez-Roman, N.; Cairney, C.; Burns, S.; Lafferty-Whyte, K.; Roffey, J.; Hammonds, T.; et al. Dynamic telomerase gene suppression via network effects of GSK3 inhibition. PLoS ONE 2009, 4, e6459. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, R.T.; Cairney, C.J.; Grabowski, P.; Keith, W.N.; Hallberg, B.; Ljungberg, B.; Roos, G. The PTEN regulator DJ-1 is associated with hTERT expression in clear cell renal cell carcinoma. Int. J. Cancer 2009, 125, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Stefanou, N.; Papanikolaou, V.; Furukawa, Y.; Nakamura, Y.; Tsezou, A. Leptin as a critical regulator of hepatocellular carcinoma development through modulation of human telomerase reverse transcriptase. BMC Cancer 2010. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Yip, H.K. Neuroprotective signaling mechanisms of telomerase are regulated by brain-derived neurotrophic factor in rat spinal cord motor neurons. J. Neuropathol. Exp. Neurol. 2011, 70, 634–652. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Ogawa, K.; Ikei, T.; Udono, M.; Fujiki, T.; Katakura, Y. SIRT1 prevents replicative senescence of normal human umbilical cord fibroblast through potentiating the transcription of human telomerase reverse transcriptase gene. Biochem. Biophys. Res. Commun. 2012, 417, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Comandini, A.; Aquino, A.; Bonmassar, L.; Guglielmi, L.; Bonmassar, E.; Franzese, O. The antiretroviral agent saquinavir enhances hTERT expression and telomerase activity in human T leukaemia cells in vitro. J. Exp. Clin. Cancer Res. 2013. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, P.; Sun, X.; Zheng, J.; Wei, G.; Zhang, L.; Wang, H.; Yao, J.; Lu, S.; Jia, P. Acidified bile acids increase hTERT expression via c-Myc activation in human gastric cancer cells. Oncol. Rep. 2015, 33, 3038–3044. [Google Scholar] [CrossRef] [PubMed]
- Hrdlickova, R.; Nehyba, J.; Bose, H.R., Jr. Regulation of telomerase activity by interferon regulatory factors 4 and 8 in immune cells. Mol. Cell. Biol. 2009, 29, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Koshiji, M.; Kageyama, Y.; Pete, E.A.; Horikawa, I.; Barrett, J.C.; Huang, L.E. Hif-1alpha induces cell cycle arrest by functionally counteracting Myc. EMBO J. 2004, 23, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Cerezo, A.; Kalthoff, H.; Schuermann, M.; Schafer, B.; Boukamp, P. Dual regulation of telomerase activity through c-Myc -dependent inhibition and alternative splicing of hTERT. J. Cell Sci. 2002, 115, 1305–1312. [Google Scholar]
- Li, H.; Xu, D.; Li, J.; Berndt, M.C.; Liu, J.P. Transforming growth factor beta suppresses human telomerase reverse transcriptase (hTERT) by Smad3 interactions with c-Myc and the hTERT gene. J. Biol. Chem. 2006, 281, 25588–25600. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.S.; Park, S.; Gwak, J.; Ju, B.G.; Oh, S. Involvement of transcription repressor snail in the regulation of human telomerase reverse transcriptase (hTERT) by transforming growth factor-beta. Biochem. Biophys. Res. Commun. 2015, 465, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Fan, S.; Meng, Q.; Schramm, L.; Wang, C.; Bouzahza, B.; Zhou, J.; Zafonte, B.; Goldberg, I.D.; Haddad, B.R.; et al. Brca1 inhibition of telomerase activity in cultured cells. Mol. Cell. Biol. 2003, 23, 8668–8690. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, J. Inhibition of human telomerase reverse transcriptase gene expression by BRCA1 in human ovarian cancer cells. Biochem. Biophys. Res. Commun. 2003, 303, 130–136. [Google Scholar] [CrossRef]
- Jang, K.J.; Kwon, G.S.; Jeong, J.W.; Kim, C.H.; Yoon, H.M.; Kim, G.Y.; Shim, J.H.; Moon, S.K.; Kim, W.J.; Choi, Y.H. Cordyceptin induces apoptosis through repressing hTERT expression and inducing extranuclear export of hTERT. J. Biosci. Bioeng. 2015, 119, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Moriai, M.; Tsuji, N.; Kobayashi, D.; Kuribayashi, K.; Watanabe, N. Down-regulation of hTERT expression plays an important role in 15-deoxy-delta12,14-prostaglandin J2-induced apoptosis in cancer cells. Int. J. Oncol. 2009, 34, 1363–1372. [Google Scholar] [PubMed]
- Toaldo, C.; Pizzimenti, S.; Cerbone, A.; Pettazzoni, P.; Menegatti, E.; Daniela, B.; Minelli, R.; Giglioni, B.; Dianzani, M.U.; Ferretti, C.; et al. Ppargamma ligands inhibit telomerase activity and hTERT expression through modulation of the Myc /Mad/Max network in colon cancer cells. J. Cell. Mol. Med. 2010, 14, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Song, Y.H.; Yim, J.; Kim, T.K. Identification of Mad as a repressor of the human telomerase (hTERT) gene. Oncogene 2000, 19, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Wahlstrom, T.; Belikov, S.; Arsenian Henriksson, M. Chromatin dynamics at the hTERT promoter during transcriptional activation and repression by c-Myc and Mnt in xenopus leavis oocytes. Exp. Cell Res. 2013, 319, 3160–3169. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Bellon, M.; Shelton, S.N.; Nicot, C. Tumor suppressors p53, p63TAα, p63TAy, p73α, and p73β use distinct pathways to repress telomerase expression. J. Biol. Chem. 2012, 287, 20737–20747. [Google Scholar] [CrossRef] [PubMed]
- Gabet, A.S.; Mortreux, F.; Charneau, P.; Riou, P.; Duc-Dodon, M.; Wu, Y.; Jeang, K.T.; Wattel, E. Inactivation of hTERT transcription by tax. Oncogene 2003, 22, 3734–3741. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, Y.; Kyo, S.; Takakura, M.; Kanaya, T.; Koshida, K.; Namiki, M.; Inoue, M. Demethylating reagent 5-azacytidine inhibits telomerase activity in human prostate cancer cells through transcriptional repression of hTERT. Clin. Cancer Res. 2000, 6, 2868–2875. [Google Scholar] [PubMed]
- Grandjenette, C.; Schnekenburger, M.; Karius, T.; Ghelfi, J.; Gaigneaux, A.; Henry, E.; Dicato, M.; Diederich, M. 5-aza-2’-deoxycytidine-mediated c-Myc down-regulation triggers telomere-dependent senescence by regulating human telomerase reverse transcriptase in chronic myeloid leukemia. Neoplasia 2014, 16, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Grand, C.L.; Han, H.; Munoz, R.M.; Weitman, S.; Von Hoff, D.D.; Hurley, L.H.; Bearss, D.J. The cationic porphyrin Tmpyp4 down-regulates c-Myc and human telomerase reverse transcriptase expression and inhibits tumor growth in vivo. Mol. Cancer Ther. 2002, 1, 565–573. [Google Scholar] [PubMed]
- Kanzawa, T.; Germano, I.M.; Kondo, Y.; Ito, H.; Kyo, S.; Kondo, S. Inhibition of telomerase activity in malignant glioma cells correlates with their sensitivity to temozolomide. Br. J. Cancer 2003, 89, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Jagadeesh, S.; Kyo, S.; Banerjee, P.P. Genistein represses telomerase activity via both transcriptional and posttranslational mechanisms in human prostate cancer cells. Cancer Res. 2006, 66, 2107–2115. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Hsiao, Y.M.; Hsu, C.P.; Lin, M.Y.; Wang, J.C.; Huang, Y.L.; Ko, J.L. Transcriptionally mediated inhibition of telomerase of fungal immunomodulatory protein from ganoderma tsugae in A549 human lung adenocarcinoma cell line. Mol. Carcinog. 2006, 45, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, S.; Briatore, F.; Laurora, S.; Toaldo, C.; Maggio, M.; De Grandi, M.; Meaglia, L.; Menegatti, E.; Giglioni, B.; Dianzani, M.U.; et al. 4-hydroxynonenal inhibits telomerase activity and hTERT expression in human leukemic cell lines. Free Radic. Biol. Med. 2006, 40, 1578–1591. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Guo, Q.L.; You, Q.D.; Lin, S.S.; Li, Z.; Gu, H.Y.; Zhang, H.W.; Tan, Z.; Wang, X. Repression of telomerase reverse transcriptase mrna and hTERT promoter by gambogic acid in human gastric carcinoma cells. Cancer Chemother. Pharmacol. 2006, 58, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Kim, M.O.; Lee, J.D.; Choi, Y.H.; Kim, G.Y. Butein suppresses c-Myc -dependent transcription and Akt-dependent phosphorylation of hTERT in human leukemia cells. Cancer Lett. 2009, 286, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.T.; Wang, C.Y.; Yang, R.C.; Chu, C.J.; Wu, H.T.; Pang, J.H. Wogonin, an active compound in Scutellaria baicalensis, induces apoptosis and reduces telomerase activity in the HL-60 leukemia cells. Phytomedicine 2010, 17, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, A.; Shen, C.; Zhang, B.; Rao, Z.; Wang, R.; Yang, S.; Ning, S.; Mao, G.; Fang, D. E2F1 acts as a negative feedback regulator of c-Myc induced hTERT transcription during tumorigenesis. Oncol. Rep. 2014, 32, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, M.; Shi, W.; Yang, Q.; Chen, C.; Wang, Z.; Zhou, X. Arsenic trioxide suppresses transcription of hTERT through down-regulation of multiple transcription factors in HL-60 leukemia cells. Toxicol. Lett. 2015, 232, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Sengupta, S.B. Role of diallyl disulfide-mediated cleavage of c-Myc and Sp-1 in the regulation of telomerase activity in human lymphoma cell line U937. Nutrition 2015, 31, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Bai, Z.; Li, X.; Hou, L.; Zhang, B. The evidences of human orphan receptor COUP-TFII inhibiting telomerase activity through decreasing hTERT transcription. Cancer Lett. 2004, 214, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Qin, L.; Wang, S.; Li, M.; Shi, D.; Tian, Y.; Wang, J.; Fu, L.; Li, Z.; Guo, W.; et al. CPSF4 activates telomerase reverse transcriptase and predicts poor prognosis in human lung adenocarcinomas. Mol. Oncol. 2014, 8, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; Loukinov, D.; Abdullaev, Z.; Guilleret, I.; Bosman, F.T.; Lobanenkov, V.; Benhattar, J. Dual role of DNA methylation inside and outside of CTCF-binding regions in the transcriptional regulation of the telomerase hTERT gene. Nucleic Acids Res. 2007, 35, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Min, N.Y.; Kim, J.H.; Choi, J.H.; Liang, W.; Ko, Y.J.; Rhee, S.; Bang, H.; Ham, S.W.; Park, A.J.; Lee, K.H. Selective death of cancer cells by preferential induction of reactive oxygen species in response to (−)-epigallocatechin-3-gallate. Biochem. Biophys. Res. Commun. 2012, 421, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yao, Z.X.; Chen, J.S.; Gi, Y.J.; Munoz, N.M.; Kundra, S.; Herlong, H.F.; Jeong, Y.S.; Goltsov, A.; Ohshiro, K.; et al. TGF-β/β2-spectrin/CTCF-regulated tumor suppression in human stem cell disorder beckwith-wiedemann syndrome. J. Clin. Investig. 2016, 126, 527–542. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Patel, S.N.; Tollefsbol, T.O. Sulforaphane causes epigenetic repression of hTERT expression in human breast cancer cell lines. PLoS ONE 2010, 5, e11457. [Google Scholar] [CrossRef] [PubMed]
- Karam, M.; Thenoz, M.; Capraro, V.; Robin, J.-P.; Pinatel, C.; Lançon, A.; Galia, P.; Sibon, D.; Thomas, X.; Ducastelle-Lepretre, S.; et al. Chromatin redistribution of the DEK oncoprotein represses hTERT transcription in leukemias. Neoplasia 2014, 16, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Crowe, D.L.; Nguyen, D.C.; Tsang, K.J.; Kyo, S. E2F-1 represses transcription of the human telomerase reverse transcriptase gene. Nucleic Acids Res. 2001, 29, 2789–2794. [Google Scholar] [CrossRef] [PubMed]
- Lacerte, A.; Korah, J.; Roy, M.; Yang, X.J.; Lemay, S.; Lebrun, J.J. Transforming growth factor-beta inhibits telomerase through SMAD3 and E2F transcription factors. Cell Signal 2008, 20, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Gizard, F.; Nomiyama, T.; Zhao, Y.; Findeisen, H.M.; Heywood, E.B.; Jones, K.L.; Staels, B.; Bruemmer, D. The PPARalpha/p16INK4a pathway inhibits vascular smooth muscle cell proliferation by repressing cell cycle-dependent telomerase activation. Circ. Res. 2008, 103, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Sekaric, P.; Cherry, J.J.; Androphy, E.J. Binding of human papillomavirus type 16 E6 to E6ap is not required for activation of hTERT. J. Virol. 2008, 82, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Akutagawa, O.; Nishi, H.; Kyo, S.; Higuma, C.; Inoue, M.; Isaka, K. Early growth response-1 mediates up-regulation of telomerase in placenta. Placenta 2007, 28, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Akutagawa, O.; Nishi, H.; Kyo, S.; Terauchi, F.; Yamazawa, K.; Higuma, C.; Inoue, M.; Isaka, K. Early growth response-1 mediates downregulation of telomerase in cervical cancer. Cancer Sci. 2008, 99, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, A.; Nanni, S.; Colussi, C.; Aiello, A.; Benvenuti, V.; Ragone, G.; Moretti, F.; Sacchi, A.; Bacchetti, S.; Gaetano, C.; et al. Estrogen receptor-alpha and endothelial nitric oxide synthase nuclear complex regulates transcription of human telomerase. Circ. Res. 2008, 103, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Goueli, B.S.; Janknecht, R. Upregulation of the catalytic telomerase subunit by the transcription factor ER81 and oncogenic HER2/Neu, Ras, or Raf. Mol. Cell. Biol. 2003, 24, 25–35. [Google Scholar] [CrossRef]
- Misiti, S.; Nanni, S.; Fontemaggi, G.; Cong, Y.S.; Wen, J.; Hirte, H.W.; Piaggio, G.; Sacchi, A.; Pontecorvi, A.; Bacchetti, S.; et al. Induction of hTERT expression and telomerase activity by estrogens in human ovary epithelium cells. Mol. Cell. Biol. 2000, 20, 3764–3771. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, K.; Tsuji, N.; Asanuma, K.; Kobayashi, D.; Watanabe, N. Inhibition of estrogen receptor beta-mediated human telomerase reverse transcriptase gene transcription via the suppression of mitogen-activated protein kinase signaling plays an important role in 15-deoxy-delta(12,14)-prostaglandin J2-induced apoptosis in cancer cells. Exp. Cell Res. 2007, 313, 3486–3496. [Google Scholar] [PubMed]
- Kasiappan, R.; Sun, Y.; Lungchukiet, P.; Quarni, W.; Zhang, X.; Bai, W. Vitamin d suppresses leptin stimulation of cancer growth through microrna. Cancer Res. 2014, 74, 6194–6204. [Google Scholar] [CrossRef] [PubMed]
- Marconett, C.N.; Sundar, S.N.; Tseng, M.; Tin, A.S.; Tran, K.Q.; Mahuron, K.M.; Bjeldanes, L.F.; Firestone, G.L. Indole-3-carbinol downregulation of telomerase gene expression requires the inhibition of estrogen receptor-alpha and Sp1 transcription factor interactions within the hTERT promoter and mediates the g1 cell cycle arrest of human breast cancer cells. Carcinogenesis 2011, 32, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Kyo, S.; Takakura, M.; Tanaka, M.; Yatabe, N.; Maida, Y.; Fujiwara, M.; Hayakawa, J.; Ohmichi, M.; Koike, K.; et al. Progesterone regulates human telomerase reverse transcriptase gene expression via activation of mitogen-activated protein kinase signaling pathway. Cancer Res. 2000, 60, 5376–5381. [Google Scholar] [PubMed]
- Hsu, C.P.; Lee, L.W.; Tang, S.C.; Hsin, I.L.; Lin, Y.W.; Ko, J.L. Epidermal growth factor activates telomerase activity by direct binding of ETS-2 to hTERT promoter in lung cancer cells. Tumour Biol. 2015, 36, 5389–5398. [Google Scholar] [CrossRef] [PubMed]
- Maida, Y.; Kyo, S.; Kanaya, T.; Wang, Z.; Yatabe, N.; Tanaka, M.; Nakamura, M.; Ohmichi, M.; Gotoh, N.; Murakami, S.; et al. Direct activation of telomerase by EGF through Ets-mediated transactivation of TERT via map kinase signaling pathway. Oncogene 2002, 21, 4071–4079. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Athanasiou, M.; Sidorov, I.A.; Horikawa, I.; Cremona, G.; Blair, D.; Barret, J.C.; Dimitrov, D.S. Role of Ets/Id proteins for telomerase regulation in human cancer cells. Exp. Mol. Pathol. 2003, 75, 238–247. [Google Scholar] [CrossRef]
- Takahashi, A.; Higashino, F.; Aoyagi, M.; Yoshida, K.; Itoh, M.; Kyo, S.; Ohno, T.; Taira, T.; Ariga, H.; Nakajima, K.; et al. EWS/EST fusions activate telomerase in ewing’s tumors. Cancer Res. 2003, 63, 8338–8344. [Google Scholar] [PubMed]
- Ogawa, D.; Nomiyama, T.; Nakamachi, T.; Heywood, E.B.; Stone, J.F.; Berger, J.P.; Law, R.E.; Bruemmer, D. Activation of peroxisome proliferator-activated receptor gamma suppresses telomerase activity in vascular smooth muscle cells. Circ. Res. 2006, 98, e50–e59. [Google Scholar] [CrossRef] [PubMed]
- Mazumdar, T.; Sandhu, R.; Qadan, M.; DeVecchio, J.; Magloire, V.; Agyeman, A.; Li, B.; Houghton, J.A. Hedgehog signaling regulates telomerase reverse transcriptase in human cancer cells. PLoS ONE 2013, 8, e75253. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Dong, Q.; Shin, K.H.; Kim, R.H.; Oh, J.E.; Park, N.H.; Kang, M.K. Grainyhead-like 2 enhances the human telomerase reverse transcriptase gene expression by inhibiting DNA methylation at the 5’-CPG island in normal human keratinocytes. J. Biol. Chem. 2010, 285, 40852–40863. [Google Scholar] [CrossRef]
- Kang, X.; Chen, W.; Kim, R.H.; Kang, M.K.; Park, N.H. Regulation of the hTERT promoter activity by MSH2, the hnRNPs K and D, and GRHL2 in human oral squamous cell carcinoma cells. Oncogene 2009, 28, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Yatabe, N.; Kyo, S.; Maida, Y.; Nishi, H.; Nakamura, M.; Kanaya, T.; Tanaka, M.; Isaka, K.; Ogawa, S.; Inoue, M. HIF-1-mediated activation of telomerase in cervical cancer cells. Oncogene 2004, 23, 3708–3715. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Nakada, T.; Kyo, S.; Inoue, M.; Shay, J.W.; Isaka, K. Hypoxia-inducible factor 1 mediates upregulation of telomerase (hTERT). Mol. Cell. Biol. 2004, 24, 6076–6083. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Zheng, D.; Deng, X.; Bai, L.; Xu, Y.; Cong, Y.S. Lysophosphatidic acid activates telomerase in ovarian cancer cells through hypoxia-inducible factor-1α and the PI3K pathway. J. Cell. Biochem. 2008, 105, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Lou, F.; Chen, X.; Jalink, M.; Zhu, Q.; Ge, N.; Zhao, S.; Fang, X.; Fan, Y.; Bjorkholm, M.; Liu, Z.; et al. The opposing effect of hypoxia-inducible factor-2α on expression of telomerase reverse transcriptase. Mol. Cancer Res. 2007, 5, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.H.; Kim, R.; Chen, W.; Hu, S.; Shin, K.H.; Park, N.H.; Kang, M.K. Association of HSP90 to the hTERT promoter is necessary for hTERT expression in human oral cancer cells. Carcinogenesis 2008, 29, 2425–2431. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Mizuguchi, M.; Fujii, M.; Nakamura, M. Kruppel-like factor 2 represses transcription of the telomerase catalytic subunit human telomerase reverse transcriptase (hTERT) in human t cells. J. Biol. Chem. 2015, 290, 8758–8763. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.W.; Hou, P.S.; Tseng, S.F.; Chien, C.L.; Wu, K.J.; Chen, H.F.; Ho, H.N.; Kyo, S.; Teng, S.C. Kruppel-like transcription factor 4 contributes to maintenance of telomerase activity in stem cells. Stem Cells 2010, 28, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Liu, C.; Ge, Z.; Fang, X.; Zhang, X.; Straat, K.; Bjorkholm, M.; Xu, D. Lysine-specific demethylase 1 (LSD1) is required for the transcriptional repression of the telomerase reverse transcriptase (hTERT) gene. PLoS ONE 2008, 3, e1446. [Google Scholar] [CrossRef] [PubMed]
- Gunes, C.; Lichtsteiner, S.; Vasserot, A.P.; Englert, C. Expression of the hTERT gene is regulated at the level of transcriptional initiation and repressed by mad1. Cancer Res. 2000, 60, 2116–2121. [Google Scholar] [PubMed]
- Farhana, L.; Dawson, M.I.; Fontana, J.A. Down regulation of miR-202 modulates Mxd1 and Sin3A repressor complexes to induce apoptosis of pancreatic cancer cells. Cancer Biol. Ther. 2015, 16, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Katzenellenbogen, R.A.; Grandori, C.; Galloway, D.A. An unbiased in vivo screen reveals multiple transcription factors that control HPV E6-regulated hTERT in keratinocytes. Virology 2013, 446, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Su, J.M.; Lai, X.M.; Lan, K.H.; Li, C.P.; Chao, Y.; Yen, S.H.; Chang, F.Y.; Lee, S.D.; Lee, W.P. X protein of hepatitis B virus functions as a transcriptional corepressor on the human telomerase promoter. Hepatology 2007, 46, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Li, M.; Su, B. MCPH1/BRIT1 represses transcription of the human telomerase reverse transcriptase gene. Gene 2012, 495, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Kyo, S.; Hua, X.; Tahara, H.; Nakajima, M.; Takakura, M.; Sakaguchi, J.; Maida, Y.; Nakamura, M.; Ikoma, T.; et al. Role of menin in the regulation of telomerase activity in normal and cancer cells. Int. J. Oncol. 2008, 33, 333–340. [Google Scholar] [PubMed]
- Borowiak, M.; Kuhlmann, A.S.; Girard, S.; Gazzolo, L.; Mesnard, J.M.; Jalinot, P.; Dodon, M.D. HTLV-1 BZIP factor impedes the menin tumor suppressor and upregulates jund-mediated transcription of the hTERT gene. Carcinogenesis 2013, 34, 2664–2672. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Kyo, S.; Takakura, M.; Kanaya, T.; Kitagawa, Y.; Itoh, H.; Takahashi, M.; Inoue, M. Identification and characterization of negative regulatory elements of the human telomerase catalytic subunit (hTERT) gene promoter: Possible role of MZF-2 in transcriptional repression of hTERT. Nucleic Acids Res. 2000, 28, 2557–2562. [Google Scholar] [CrossRef] [PubMed]
- Gizard, F.; Heywood, E.B.; Findeisen, H.M.; Zhao, Y.; Jones, K.L.; Cudejko, C.; Post, G.R.; Staels, B.; Bruemmer, D. Telomerase activation in atherosclerosis and induction of telomerase reverse transcriptase expression by inflammatory stimuli in macrophages. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, N.; Veeraraghavan, J.; Madhusoodhanan, R.; Herman, T.S.; Natarajan, M. Curcumin regulates low-linear energy transfer gamma-radiation-induced NFκB-dependent telomerase activity in human neuroblastoma cells. Int. J. Radiat. Oncol. Biol. Phys. 2011, 79, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.S.; Aroh, C.; Vadgama, J.V. Constitutive activation of STAT3 signaling regulates hTERT and promotes stem cell-like traits in human breast cancer cells. PLoS ONE 2013, 8, e83971. [Google Scholar] [CrossRef] [PubMed]
- Bellon, M.; Nicot, C. Central role of PI3K in transcriptional activation of hTERT in HTLV-I-Infected cells. Blood 2008, 112, 2946–2955. [Google Scholar] [CrossRef] [PubMed]
- Kawagoe, J.; Ohmichi, M.; Takahashi, T.; Ohshima, C.; Mabuchi, S.; Takahashi, K.; Igarashi, H.; Mori-Abe, A.; Saitoh, M.; Du, B.; et al. Raloxifene inhibits estrogen-induced up-regulation of telomerase activity in a human breast cancer cell line. J. Biol. Chem. 2003, 278, 43363–43372. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, M.; Mohan, S.; Konopinski, R.; Otto, R.A.; Herman, T.S. Induced telomerase activity in primary aortic endothelial cells by low-let gamma-radiation is mediated through NF-κb activation. Br. J. Radiol. 2008, 81, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, N.; Aravindan, S.; Herman, T.S.; Natarajan, M. EGFR tyrosine kinase inhibitor pelitinib regulates radiation-induced p65-dependent telomerase activation in squamous cell carcinoma. Radiat. Res. 2013, 179, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Chebel, A.; Rouault, J.P.; Urbanowicz, I.; Baseggio, L.; Chien, W.W.; Salles, G.; Ffrench, M. Transcriptional activation of hTERT, the human telomerase reverse transcriptase, by nuclear factor of activated T cells. J. Biol. Chem. 2009, 284, 35725–35734. [Google Scholar] [CrossRef] [PubMed]
- Galloway, D.A.; Gewin, L.C.; Myers, H.; Luo, W.; Grandori, C.; Katzenellenbogen, R.A.; McDougall, J.K. Regulation of telomerase by human papillomaviruses. Cold Spring Harb. Symp. Quant. Biol. 2005, 70, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Bougel, S.; Renaud, S.; Braunschweig, R.; Loukinov, D.; Morse, H.C., III; Bosman, F.T.; Lobanenkov, V.; Benhattar, J. PAX5 activates the transcription of the human telomerase reverse transcriptase gene in B cells. J. Pathol. 2010, 220, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Campbell, H.G.; Wiles, A.K.; Eccles, M.R.; Reddel, R.R.; Braithwaite, A.W.; Royds, J.A. PAX8 regulates telomerase reverse transcriptase and telomerase RNA component in glioma. Cancer Res. 2008, 68, 5724–5732. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.L.; Ohhira, T.; Fujisaki, C.; Inoue, T.; Ohta, T.; Osaki, M.; Ohshiro, E.; Seko, T.; Aoki, S.; Oshimura, M.; et al. Identification of PITX1 as a TERT suppressor gene located on human chromosome 5. Mol. Cell. Biol. 2011, 31, 1624–1636. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Naohiro, S.; Nakayama, Y.; Osaki, M.; Okada, F.; Oshimura, M.; Kugoh, H. miR-19b regulates hTERT mRNA expression through targeting PITX1 mRNA in melanoma cells. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; Liu, H.; Gao, L.; Liu, J.; Sun, Z.; Ju, Y.; Hou, N.; Guo, C.; Liang, X.; Zhang, L.; et al. Hepatitis B virus protein preS2 potentially promotes HCC development via its transcriptional activation of hTERT. Gut 2009, 58, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zeng, J.; Li, Q.; Zhao, L.; Liu, T.; Bjorkholm, M.; Jia, J.; Xu, D. Reptin is required for the transcription of telomerase reverse transcriptase and over-expressed in gastric cancer. Mol. Cancer 2010. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Lu, J.; Qin, Y.; Wang, J.; Tian, Y.; Shi, D.; Wang, S.; Xiao, Y.; Dai, M.; Liu, L.; et al. Ret finger protein-like 3 promotes tumor cell growth by activating telomerase reverse transcriptase expression in human lung cancer cells. Oncotarget 2014, 5, 11909–11923. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Chen, W.; Xiao, Y.; Yu, W.; Cai, X.; Dai, M.; Xu, T.; Huang, W.; Guo, W.; Deng, W.; et al. RFPL3 and CBP synergistically upregulate hTERT activity and promote lung cancer growth. Oncotarget 2015, 6, 27130–27145. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Lu, J.; Dai, M.; Wu, T.; Yu, Z.; Wang, J.; Chen, W.; Shi, D.; Yu, W.; Xiao, Y.; et al. Transcriptional coactivator cbp upregulates hTERT expression and tumor growth and predicts poor prognosis in human lung cancers. Oncotarget 2014, 5, 9349–9361. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.S.; Cotter, M.A., II; Robertson, E.S. The latency-associated nuclear antigen of kaposi’s sarcoma-associated herpesvirus transactivates the telomerase reverse transcriptase promoter. J. Biol. Chem. 2001, 276, 22971–22978. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.C.; Borah, S.; Robertson, E.S. Latency-associated nuclear antigen of kaposi’s sarcoma-associated herpesvirus up-regulates transcription of human telomerase reverse transcriptase promoter through interaction with transcription factor Sp1. J. Virol. 2004, 78, 10348–10359. [Google Scholar] [CrossRef] [PubMed]
- Chau, M.N.; El Touny, L.H.; Jagadeesh, S.; Banerjee, P.P. Physiologically achievable concentrations of genistein enhance telomerase activity in prostate cancer cells via the activation of STAT3. Carcinogenesis 2007, 28, 2282–2290. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ishihara, K.; Ichimura, T.; Fujita, N.; Hino, S.; Tomita, S.; Watanabe, S.; Saitoh, N.; Ito, T.; Nakao, M. MCAF1/AM is involved in Sp1-mediated maintenance of cancer-associated telomerase activity. J. Biol. Chem. 2009, 284, 5165–5174. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shi, W.; Luan, F.; Xu, S.; Yang, F.; Sun, W.; Liu, J.; Ma, C. Hepatitis B virus X protein upregulates transcriptional activation of human telomerase reverse transcriptase. Virus Genes 2010, 40, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Li, A.Y.; Lin, H.H.; Kuo, C.Y.; Shih, H.M.; Wang, C.C.; Yen, Y.; Ann, D.K. High-mobility group a2 protein modulates hTERT transcription to promote tumorigenesis. Mol. Cell. Biol. 2011, 31, 2605–2617. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wang, Q.; Gruber, A.; Bjorkholm, M.; Chen, Z.; Zaid, A.; Selivanova, G.; Peterson, C.; Wiman, K.G.; Pisa, P. Downregulation of telomerase reverse transcriptase mrna expression by wild type p53 in human tumor cells. Oncogene 2000, 19, 5123–5133. [Google Scholar] [CrossRef] [PubMed]
- Beitzinger, M.; Oswald, C.; Beinoraviciute-Kellner, R.; Stiewe, T. Regulation of telomerase activity by the p53 family member p73. Oncogene 2006, 25, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Racek, T.; Mise, N.; Li, Z.; Stoll, A.; Putzer, B.M. C-terminal p73 isoforms repress transcriptional activity of the human telomerase reverse transcriptase (hTERT) promoter. J. Biol. Chem. 2005, 280, 40402–40405. [Google Scholar] [CrossRef] [PubMed]
- Wooten, L.G.; Ogretmen, B. Sp1/Sp3-dependent regulation of human telomerase reverse transcriptase promoter activity by the bioactive sphingolipid ceramide. J. Biol. Chem. 2005, 280, 28867–28876. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, H.Z.; Jeong, K.W.; Shim, Y.S.; Horikawa, I.; Barrett, J.C.; Choe, J. Human papillomavirus E2 down-regulates the human telomerase reverse transcriptase promoter. J. Biol. Chem. 2002, 277, 27748–27756. [Google Scholar] [CrossRef] [PubMed]
- Won, J.; Yim, J.; Kim, T.K. Opposing regulatory roles of E2F in human telomerase reverse transcriptase (hTERT) gene expression in human tumor and normal somatic cells. FASEB J. 2002, 16, 1943–1945. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Nakagawa, K.; Hamada, K.; Hirose, S.; Harada, H.; Kohno, S.; Nagato, S.; Ohnishi, T. Introduction of p16INK4a inhibits telomerase activity through transcriptional suppression of human telomerase reverse transcriptase expression in human gliomas. Int. J. Oncol. 2004, 24, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.C.; Lin, S.Z.; Chen, Y.L.; Chang, J.S.; Ho, L.I.; Liu, P.Y.; Chang, L.F.; Harn, Y.C.; Chen, S.P.; Sun, L.Y.; et al. Butylidenephthalide suppresses human telomerase reverse transcriptase (TERT) in human glioblastomas. Ann. Surg. Oncol. 2011, 18, 3514–3527. [Google Scholar] [CrossRef] [PubMed]
- Shapira, S.; Granot, G.; Mor-Tzuntz, R.; Raanani, P.; Uziel, O.; Lahav, M.; Shpilberg, O. Second-generation tyrosine kinase inhibitors reduce telomerase activity in K562 cells. Cancer Lett. 2012, 323, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Deeb, D.; Brigolin, C.; Gao, X.; Liu, Y.; Pindolia, K.R.; Gautam, S.C. Induction of apoptosis in pancreatic cancer cells by CDDO-Me involves repression of telomerase through epigenetic pathways. J. Carcinog. Mutagen. 2014. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, X.; Liu, J.; Xie, X.; Cui, W.; Zhu, Y. Homocysteine accelerates senescence of endothelial cells via DNA hypomethylation of human telomerase reverse transcriptase. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Wang, J.; Guo, W.; Wang, H.; Wang, C.; Liu, Y.; Sun, X. Triptolide inhibits transcription of hTERT through down-regulation of transcription factor specificity protein 1 in primary effusion lymphoma cells. Biochem. Biophys. Res. Commun. 2016, 469, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Won, J.; Yim, J.; Kim, T.K. Sp1 and Sp3 recruit histone deacetylase to repress transcription of human telomerase reverse transcriptase (hTERT) promoter in normal human somatic cells. J. Biol. Chem. 2002, 277, 38230–38238. [Google Scholar] [CrossRef] [PubMed]
- Glasspool, R.M.; Burns, S.; Hoare, S.F.; Svensson, C.; Keith, W.N. The hTERT and hTERC telomerase gene promoters are activated by the second exon of the adenoviral protein, E1 A, identifying the transcriptional corepressor CtBP as a potential rearessor of both genes. Neoplasia 2005, 7, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, T.; Miura, T.; Maura, M.; Shiraishi, H.; Nishimura, S.; Imada, Y.; Uehara, N.; Tashiro, K.; Shirahata, S.; Katakura, Y. TAK1 represses transcription of the human telomerase reverse transcriptase gene. Oncogene 2007, 26, 5258–5266. [Google Scholar] [CrossRef] [PubMed]
- Wooten-Blanks, L.G.; Song, P.; Senkal, C.E.; Ogretmen, B. Mechanisms of ceramide-mediated repression of the human telomerase reverse transcriptase promoter via deacetylation of Sp3 by histone deacetylase 1. FASEB J. 2007, 21, 3386–3397. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhu, J.; Dong, Y.; He, C.; Hu, X. Suppressor of Ty homolog-5, a novel tumor-specific human telomerase reverse transcriptase promoter-binding protein and activator in colon cancer cells. Oncotarget 2015, 6, 32841–32855. [Google Scholar] [PubMed]
- Ren, H.; Zhao, T.; Wang, X.; Gao, C.; Wang, J.; Yu, M.; Hao, J. Leptin upregulates telomerase activity and transcription of human telomerase reverse transcriptase in MCF-7 breast cancer cells. Biochem. Biophys. Res. Commun. 2010, 394, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Nakatake, M.; Kakiuchi, Y.; Sasaki, N.; Murakami-Murofushi, K.; Yamada, O. STAT3 and PKC differentially regulate telomerase activity during megakaryocytic differentiation of K562 cells. Cell Cycle 2007, 6, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Konnikova, L.; Simeone, M.C.; Kruger, M.M.; Kotecki, M.; Cochran, B.H. Signal transducer and activator of transcription 3 (STAT3) regulates human telomerase reverse transcriptase (hTERT) expression in human cancer and primary cells. Cancer Res. 2005, 65, 6516–6520. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Sun, G.; Luo, H.; Wang, X.F.; Lan, F.M.; Yue, X.; Fu, L.S.; Pu, P.Y.; Kang, C.S.; Liu, N.; et al. MiR-21 modulates hTERT through a STAT3-dependent manner on glioblastoma cell growth. CNS Neurosci. Ther. 2012, 18, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Cheng, D.; Li, Z.H.; Zhou, Q.B.; Zhou, J.J.; Lin, Q.; Zeng, B.; Liao, Q.; Chen, R.F. Transfection of HCVc improves hTERT expression through STAT3 pathway by epigenetic regulation in Huh7 cells. J. Cell. Biochem. 2012, 113, 3419–3426. [Google Scholar] [CrossRef] [PubMed]
- Yamada, O.; Ozaki, K.; Akiyama, M.; Kawauchi, K. JAK-STAT and JAK-PI3K-mTORC1 pathways regulate telomerase transcriptionally and posttranslationally in atl cells. Mol. Cancer Ther. 2012, 11, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Yamada, O.; Ozaki, K.; Furukawa, T.; Machida, M.; Wang, Y.H.; Motoji, T.; Mitsuishi, T.; Akiyama, M.; Yamada, H.; Kawauchi, K.; et al. Activation of STAT5 confers imatinib resistance on leukemic cells through the transcription of TERT and mdr1. Cell Signal 2011, 23, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Kawano, T.; Mikami-Terao, Y.; Agawa-Ohta, M.; Yamada, O.; Ida, H.; Yamada, H. Erythropoietin activates telomerase through transcriptional and posttranscriptional regulation in human erythroleukemic JAS-REN-A cells. Leuk. Res. 2011, 35, 416–418. [Google Scholar] [CrossRef] [PubMed]
- Prade-Houdellier, N.; Frebet, E.; Demur, C.; Gautier, E.F.; Delhommeau, F.; Bennaceur-Griscelli, A.L.; Gaudin, C.; Martinel, V.; Laurent, G.; Mansat-De Mas, V.; et al. Human telomerase is regulated by erythropoietin and transforming growth factor-β in human erythroid progenitor cells. Leukemia 2007, 21, 2304–2310. [Google Scholar] [CrossRef] [PubMed]
- Terme, J.M.; Mocquet, V.; Kuhlmann, A.S.; Zane, L.; Mortreux, F.; Wattel, E.; Duc Dodon, M.; Jalinot, P. Inhibition of the hTERT promoter by the proto-oncogenic protein TAL1. Leukemia 2009, 23, 2081–2089. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Zhao, Y.; Mei, F.; Yang, S.; Li, X.; Lv, J.; Hou, L.; Zhang, B. Molecular cloning and characterization of a human gene involved in transcriptional regulation of hTERT. Biochem. Biophys. Res. Commun. 2004, 324, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Chen, C.H.; Hsu, T.I.; Hung, J.J.; Ko, J.L.; Zhang, B.; Lee, Y.C.; Chen, H.K.; Chang, W.C.; Lin, D.Y. The 58-kDa microspherule protein (MSP58) represses human telomerase reverse transcriptase (hTERT) gene expression and cell proliferation by interacting with telomerase transcriptional element-interacting factor (TEIF). Biochim. Biophys. Acta 2014, 1843, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Goueli, B.S.; Janknecht, R. Regulation of telomerase reverse transcriptase gene activity by upstream stimulatory factor. Oncogene 2003, 22, 8042–8047. [Google Scholar] [CrossRef] [PubMed]
- Yago, M.; Ohki, R.; Hatakeyama, S.; Fujita, T.; Ishikawa, F. Variant forms of upstream stimulatory factors (USFs) control the promoter activity of hTERT, the human gene encoding the catalytic subunit of telomerase. FEBS Lett. 2002, 520, 40–46. [Google Scholar] [CrossRef]
- Chang, J.T.; Yang, H.T.; Wang, T.C.; Cheng, A.J. Upstream stimulatory factor (USF) as a transcriptional suppressor of human telomerase reverse transcriptase (hTERT) in oral cancer cells. Mol. Carcinog. 2005, 44, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, N.; Uemura, H.; Ishiguro, H.; Hori, M.; Hosaka, M.; Kyo, S.; Miyamoto, K.; Takeda, E.; Kubota, Y. Combination treatment with 1α,25-dihydroxyvitamin D3 and 9-Cis-Retinoic acid directly inhibits human telomerase reverse transcriptase transcription in prostate cancer cells. Mol. Cancer Ther. 2003, 2, 739–746. [Google Scholar] [PubMed]
- Oh, S.; Song, Y.; Yim, J.; Kim, T.K. The wilms’ tumor 1 tumor suppressor gene represses transcription of the human telomerase reverse transcriptase gene. J. Biol. Chem. 1999, 274, 37473–37478. [Google Scholar] [CrossRef] [PubMed]
- Veldman, T.; Horikawa, I.; Barrett, J.C.; Schlegel, R. Transcriptional activation of the telomerase hTERT gene by human papillomavirus type 16 E6 oncoprotein. J. Virol. 2001, 75, 4467–4472. [Google Scholar] [CrossRef] [PubMed]
- Kanzawa, T.; Komata, T.; Kyo, S.; Germano, I.M.; Kondo, Y.; Kondo, S. Down-regulation of telomerase activity in malignant glioma cells by p27KIP1. Int. J. Oncol. 2003, 23, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Seo, H.S.; Choi, W.S.; Kim, O.; Nam, S.W.; Lee, J.Y.; Park, W.S. Gastrokine 1 induces senescence and apoptosis through regulating telomere length in gastric cancer. Oncotarget 2014, 5, 11695–11708. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, K.L.; Newbold, R.F.; Mokbel, K. There is no correlation between c-Myc mRNA expression and telomerase activity in human breast cancer. Int. Semin. Surg. Oncol. 2004. [Google Scholar] [CrossRef] [PubMed]
- Elkak, A.E.; Meligonis, G.; Salhab, M.; Mitchell, B.; Blake, J.R.; Newbold, R.F.; Mokbel, K. hTERT protein expression is independent of clinicopathological parameters and c-Myc protein expression in human breast cancer. J. Carcinog. 2005. [Google Scholar] [CrossRef]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Saginc, G.; Leow, S.C.; Khattar, E.; Shin, E.M.; Yan, T.D.; Wong, M.; Zhang, Z.; Li, G.; Sung, W.K.; et al. Telomerase directly regulates NF-κB-dependent transcription. Nat. Cell Biol. 2012, 14, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Xi, P.; Zhou, J.; Wang, M.; Cong, Y.S. Human telomerase reverse transcriptase regulates mmp expression independently of telomerase activity via NF-κB-dependent transcription. FASEB J. 2013, 27, 4375–4383. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.L.; Sun, W.; Chandrasekharan, P.; Cheng, H.S.; Ying, Z.; Lakshmanan, M.; Raju, A.; Tenen, D.G.; Cheng, S.Y.; et al. Non-canonical NF-κB signalling and ETS1/2 cooperatively drive C250T mutant TERT promoter activation. Nat. Cell Biol. 2015, 17, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Liu, B.R.; Qu, B.; Xing, H.; Gao, S.L.; Yin, J.M.; Wang, X.F.; Cheng, Y.Q. Silencing STAT3 may inhibit cell growth through regulating signaling pathway, telomerase, cell cycle, apoptosis and angiogenesis in hepatocellular carcinoma: Potential uses for gene therapy. Neoplasma 2011, 58, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Boggess, J.F.; Zhou, C.; Bae-Jump, V.L.; Gehrig, P.A.; Whang, Y.E. Estrogen-receptor-dependent regulation of telomerase activity in human endometrial cancer cell lines. Gynecol. Oncol. 2006, 103, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Briatore, F.; Barrera, G.; Pizzimenti, S.; Toaldo, C.; Casa, C.D.; Laurora, S.; Pettazzoni, P.; Dianzani, M.U.; Ferrero, D. Increase of telomerase activity and hTERT expression in myelodysplastic syndromes. Cancer Biol. Ther. 2009, 8, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Meeran, S.M.; Patel, S.N.; Chan, T.H.; Tollefsbol, T.O. A novel prodrug of epigallocatechin-3-gallate: Differential epigenetic hTERT repression in human breast cancer cells. Cancer Prev. Res. 2011, 4, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Yu, R.A.; He, L.F.; An, S.J.; Wu, Z.G.; Yang, K.D.; Chen, X.M. Inhibitory effects of selenium on telomerase activity and hTERT expression in cadmium-transformed 16HBE cells. Biomed. Environ. Sci. 2007, 20, 307–312. [Google Scholar] [PubMed]
- Cheng, D.; Zhao, Y.; Wang, S.; Jia, W.; Kang, J.; Zhu, J. Human telomerase reverse transcriptase (hTERT) transcription requires Sp1/Sp3 binding to the promoter and a permissive chromatin environment. J. Biol. Chem. 2015, 290, 30193–30203. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.X.; Chen, X.Y.; Wu, S.G. Regulation of telomerase activity in Jurkat T cells by transcription factors Sp1 and Sp3. Di Yi Jun Yi Da Xue Xue Bao 2002, 22, 481–485. [Google Scholar] [PubMed]
- Eldholm, V.; Haugen, A.; Zienolddiny, S. Ctcf mediates the TERT enhancer-promoter interactions in lung cancer cells: Identification of a novel enhancer region involved in the regulation of TERT gene. Int. J. Cancer 2014, 134, 2305–2313. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Bao, J.; Li, P.; Nicosia, S.V.; Bai, W. Induction of ovarian cancer cell apoptosis by 1,25-dihydroxyvitamin D3 through the down-regulation of telomerase. J. Biol. Chem. 2004, 279, 53213–53221. [Google Scholar] [CrossRef] [PubMed]
- Kasiappan, R.; Shen, Z.; Tse, A.K.; Jinwal, U.; Tang, J.; Lungchukiet, P.; Sun, Y.; Kruk, P.; Nicosia, S.V.; Zhang, X.; et al. 1,25-Dihydroxyvitamin d3 suppresses telomerase expression and human cancer growth through microrna-498. J. Biol. Chem. 2012, 287, 41297–41309. [Google Scholar] [CrossRef] [PubMed]
- Kanaya, T.; Kyo, S.; Hamada, K.; Takakura, M.; Kitagawa, Y.; Harada, H.; Inoue, M. Adenoviral expression of p53 represses telomerase activity through down-regulation of human telomerase reverse transcriptase transcription. Clin. Cancer Res. 2000, 6, 1239–1247. [Google Scholar] [PubMed]
- Saito, T.; Akaike, K.; Kurisaki-Arakawa, A.; Toda-Ishii, M.; Mukaihara, K.; Suehara, Y.; Takagi, T.; Kaneko, K.; Yao, T. Promoter mutations are rare in bone and soft tissue sarcomas of japanese patients. Mol. Clin. Oncol. 2016, 4, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Wang, X.; Popov, N.; Zhang, A.; Zhao, X.; Zhou, R.; Zetterberg, A.; Bjorkholm, M.; Henriksson, M.; Gruber, A.; et al. The histone deacetylase inhibitor trichostatin a derepresses the telomerase reverse transcriptase (hTERT) gene in human cells. Exp. Cell Res. 2002, 274, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Takakura, M.; Kyo, S.; Sowa, Y.; Wang, Z.; Yatabe, N.; Maida, Y.; Tanaka, M.; Inoue, M. Telomerase activation by histone deacetylase inhibitor in normal cells. Nucleic Acids Res. 2001, 29, 3006–3011. [Google Scholar] [CrossRef] [PubMed]
- Varshney, A.; Ramakrishnan, S.K.; Sharma, A.; Santosh, B.; Bala, J.; Yadava, P.K.; Jaiswal, R.K. Global expression profile of telomerase-associated genes in HeLa cells. Gene 2014, 547, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.P.; Hoare, S.F.; Glasspool, R.M.; Keith, W.N. Lack of telomerase gene expression in alternative lengthening of telomere cells is associated with chromatin remodeling of the hTR and hTERT gene promoters. Cancer Res. 2005, 65, 7585–7590. [Google Scholar] [PubMed]
- Wang, Y.; Chen, T.; Yan, H.; Qi, H.; Deng, C.; Ye, T.; Zhou, S.; Li, F.R. Role of histone deacetylase inhibitors in the aging of human umbilical cord mesenchymal stem cells. J. Cell. Biochem. 2013, 114, 2231–2239. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Guarente, L.P. Mammalian sirtuins—Emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006, 20, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Jeng, Y.M.; Yuan, R.H.; Hsu, H.C.; Chen, Y.L. SIRT1 promotes tumorigenesis and resistance to chemotherapy in hepatocellular carcinoma and its expression predicts poor prognosis. Ann. Surg. Oncol. 2012, 19, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Zhao, G.; Lv, X.; Chen, H.Z.; Xue, Z.; Yang, B.; Liu, D.P.; Liang, C.C. SIRT1 deacetylates c-Myc and promotes c-Myc /max association. Int. J. Biochem. Cell Biol. 2011, 43, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, B.; Wong, N.; Lo, A.W.; To, K.F.; Chan, A.W.; Ng, M.H.; Ho, C.Y.; Cheng, S.H.; Lai, P.B.; et al. Sirtuin 1 is upregulated in a subset of hepatocellular carcinomas where it is essential for telomere maintenance and tumor cell growth. Cancer Res. 2011, 71, 4138–4149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, J.; Cheng, A.S.; Ko, B.C. Depletion of sirtuin 1 (SIRT1) leads to epigenetic modifications of telomerase (TERT) gene in hepatocellular carcinoma cells. PLoS ONE 2014, 9, e84931. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Selker, E.U. Gene silencing: Repeats that count. Cell 1999, 97, 157–160. [Google Scholar] [CrossRef]
- Devereux, T.R.; Horikawa, I.; Anna, C.H.; Annab, L.A.; Afshari, C.A.; Barrett, J.C. DNA methylation analysis of the promoter region of the human telomerase reverse transcriptase (hTERT) gene. Cancer Res. 1999, 59, 6087–6090. [Google Scholar] [PubMed]
- Iliopoulos, D.; Satra, M.; Drakaki, A.; Poultsides, G.A.; Tsezou, A. Epigenetic regulation of hTERT promoter in hepatocellular carcinomas. Int. J. Oncol. 2009, 34, 391–399. [Google Scholar] [PubMed]
- Zinn, R.L.; Pruitt, K.; Eguchi, S.; Baylin, S.B.; Herman, J.G. hTERT is expressed in cancer cell lines despite promoter DNA methylation by preservation of unmethylated DNA and active chromatin around the transcription start site. Cancer Res. 2007, 67, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Guilleret, I.; Benhattar, J. Demethylation of the human telomerase catalytic subunit (hTERT) gene promoter reduced hTERT expression and telomerase activity and shortened telomeres. Exp. Cell Res. 2003, 289, 326–334. [Google Scholar] [CrossRef]
- Pettigrew, K.A.; Armstrong, R.N.; Colyer, H.A.; Zhang, S.D.; Rea, I.M.; Jones, R.E.; Baird, D.M.; Mills, K.I. Differential TERT promoter methylation and response to 5-Aza-2’-deoxycytidine in acute myeloid leukemia cell lines: TERT expression, telomerase activity, telomere length, and cell death. Genes Chromosomes Cancer 2012, 51, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Dessain, S.K.; Yu, H.; Reddel, R.R.; Beijersbergen, R.L.; Weinberg, R.A. Methylation of the human telomerase gene cpg island. Cancer Res. 2000, 60, 537–541. [Google Scholar] [PubMed]
- Guilleret, I.; Yan, P.; Grange, F.; Braunschweig, R.; Bosman, F.T.; Benhattar, J. Hypermethylation of the human telomerase catalytic subunit (hTERT) gene correlates with telomerase activity. Int. J. Cancer 2002, 101, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, P.; Choufani, S.; Mack, S.; Gallagher, D.; Zhang, C.; Lipman, T.; Zhukova, N.; Walker, E.J.; Martin, D.; Merino, D.; et al. Methylation of the TERT promoter and risk stratification of childhood brain tumours: An integrative genomic and molecular study. Lancet. Oncol. 2013, 14, 534–542. [Google Scholar] [CrossRef]
- Peifer, M.; Hertwig, F.; Roels, F.; Dreidax, D.; Gartlgruber, M.; Menon, R.; Kramer, A.; Roncaioli, J.L.; Sand, F.; Heuckmann, J.M.; et al. Telomerase activation by genomic rearrangements in high-risk neuroblastoma. Nature 2015, 526, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.L.; Theodorescu, D.; Vogelstein, B.; Papadopoulos, N.; Cech, T.R. Mutation of the TERT promoter, switch to active chromatin, and monoallelic TERT expression in multiple cancers. Genes Dev. 2015, 29, 2219–2224. [Google Scholar] [CrossRef] [PubMed]
- Ojesina, A.I.; Lichtenstein, L.; Freeman, S.S.; Pedamallu, C.S.; Imaz-Rosshandler, I.; Pugh, T.J.; Cherniack, A.D.; Ambrogio, L.; Cibulskis, K.; Bertelsen, B.; et al. Landscape of genomic alterations in cervical carcinomas. Nature 2014, 506, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Kandoth, C.; McLellan, M.D.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A.; et al. Mutational landscape and significance across 12 major cancer types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Weinhold, N.; Jacobsen, A.; Schultz, N.; Sander, C.; Lee, W. Genome-wide analysis of noncoding regulatory mutations in cancer. Nat. Genet. 2014, 46, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly recurrent TERT promoter mutations in human melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. Tert promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz, L.A., Jr.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar] [CrossRef] [PubMed]
- Quaas, A.; Oldopp, T.; Tharun, L.; Klingenfeld, C.; Krech, T.; Sauter, G.; Grob, T.J. Frequency of TERT promoter mutations in primary tumors of the liver. Virchows Arch. 2014, 465, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Kinde, I.; Munari, E.; Faraj, S.F.; Hruban, R.H.; Schoenberg, M.; Bivalacqua, T.; Allaf, M.; Springer, S.; Wang, Y.; Diaz, L.A., Jr.; et al. TERT promoter mutations occur early in urothelial neoplasia and are biomarkers of early disease and disease recurrence in urine. Cancer Res. 2013, 73, 7162–7167. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Schilling, B.; Murali, R.; Bielefeld, N.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Hillen, U.; Schaller, J.; Brenn, T.; et al. TERT promoter mutations are frequent in atypical fibroxanthomas and pleomorphic dermal sarcomas. Mod. Pathol. 2014, 27, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Adeberg, S.; Bernhardt, D.; Harrabi, S.B.; Diehl, C.; Koelsche, C.; Rieken, S.; Unterberg, A.; von Deimling, A.; Debus, J. Radiotherapy plus concomitant temozolomide in primary gliosarcoma. J. Neurooncol. 2016, 128, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.E.; Ohta, T.; Nonoguchi, N.; Satomi, K.; Capper, D.; Pierscianek, D.; Sure, U.; Vital, A.; Paulus, W.; Mittelbronn, M.; et al. Genetic alterations in gliosarcoma and giant cell glioblastoma. Brain Pathol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Sahm, F.; Capper, D.; Reuss, D.; Sturm, D.; Jones, D.T.; Kool, M.; Northcott, P.A.; Wiestler, B.; Bohmer, K.; et al. Distribution of TERT promoter mutations in pediatric and adult tumors of the nervous system. Acta Neuropathol. 2013, 126, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.S.; Wang, Z.; He, X.J.; Diplas, B.H.; Yang, R.; Killela, P.J.; Meng, Q.; Ye, Z.Y.; Wang, W.; Jiang, X.T.; et al. Recurrent TERT promoter mutations identified in a large-scale study of multiple tumour types are associated with increased TERT expression and telomerase activation. Eur. J. Cancer 2015, 51, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Renner, M.; Hartmann, W.; Brandt, R.; Lehner, B.; Waldburger, N.; Alldinger, I.; Schmitt, T.; Egerer, G.; Penzel, R.; et al. TERT promoter hotspot mutations are recurrent in myxoid liposarcomas but rare in other soft tissue sarcoma entities. J. Exp. Clin. Cancer Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Vail, E.; Zheng, X.; Zhou, M.; Yang, X.; Fallon, J.T.; Epstein, J.I.; Zhong, M. Telomerase reverse transcriptase promoter mutations in glandular lesions of the urinary bladder. Ann. Diagn. Pathol. 2015, 19, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Nonoguchi, N.; Ohta, T.; Oh, J.E.; Kim, Y.H.; Kleihues, P.; Ohgaki, H. TERT promoter mutations in primary and secondary glioblastomas. Acta Neuropathol. 2013, 126, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Narita, Y.; Fukushima, S.; Tateishi, K.; Matsushita, Y.; Yoshida, A.; Miyakita, Y.; Ohno, M.; Collins, V.P.; Kawahara, N.; et al. Upregulating mutations in the TERT promoter commonly occur in adult malignant gliomas and are strongly associated with total 1p19q loss. Acta Neuropathol. 2013, 126, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Mosrati, M.A.; Malmstrom, A.; Lysiak, M.; Krysztofiak, A.; Hallbeck, M.; Milos, P.; Hallbeck, A.L.; Bratthall, C.; Strandeus, M.; Stenmark-Askmalm, M.; et al. TERT promoter mutations and polymorphisms as prognostic factors in primary glioblastoma. Oncotarget 2015, 6, 16663–16673. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Hosen, I.; Gousias, K.; Rachakonda, S.; Heidenreich, B.; Gessi, M.; Schramm, J.; Hemminki, K.; Waha, A.; Kumar, R. TERT promoter mutations: A novel independent prognostic factor in primary glioblastomas. Neuro-Oncology 2015, 17, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Labussiere, M.; Boisselier, B.; Mokhtari, K.; Di Stefano, A.L.; Rahimian, A.; Rossetto, M.; Ciccarino, P.; Saulnier, O.; Paterra, R.; Marie, Y.; et al. Combined analysis of TERT, EGFR, and IDH status defines distinct prognostic glioblastoma classes. Neurology 2014, 83, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hosen, I.; Volz, F.; Hemminki, K.; Weyerbrock, A.; Kumar, R. TERT promoter mutations and telomere length in adult malignant gliomas and recurrences. Oncotarget 2015, 6, 10617–10633. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, G.; Shan, Y.; Hartmann, C.; von Deimling, A.; Xing, M. Highly prevalent TERT promoter mutations in bladder cancer and glioblastoma. Cell Cycle 2013, 12, 1637–1638. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Almeida, A.; Populo, H.; Batista, R.; Lyra, J.; Pinto, V.; Coelho, R.; Celestino, R.; Prazeres, H.; Lima, L.; et al. Frequency of TERT promoter mutations in human cancers. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, N.J.; Ny, L.; Nilsson, J.A.; Larsson, E. Systematic analysis of noncoding somatic mutations and gene expression alterations across 14 tumor types. Nat. Genet. 2014, 46, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Batista, R.; Cruvinel-Carloni, A.; Vinagre, J.; Peixoto, J.; Catarino, T.A.; Campanella, N.C.; Menezes, W.; Becker, A.P.; de Almeida, G.C.; Matsushita, M.M.; et al. The prognostic impact of TERT promoter mutations in glioblastomas is modified by the rs2853669 single nucleotide polymorphism. Int. J. Cancer 2016, 139, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Nencha, U.; Rahimian, A.; Giry, M.; Sechi, A.; Mokhtari, K.; Polivka, M.; Schmitt, Y.; Di Stefano, A.L.; Alentorn, A.; Labussiere, M.; et al. TERT promoter mutations and rs2853669 polymorphism: Prognostic impact and interactions with common alterations in glioblastomas. J. Neurooncol. 2016, 126, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Spiegl-Kreinecker, S.; Lotsch, D.; Ghanim, B.; Pirker, C.; Mohr, T.; Laaber, M.; Weis, S.; Olschowski, A.; Webersinke, G.; Pichler, J.; et al. Prognostic quality of activating TERT promoter mutations in glioblastoma: Interaction with the rs2853669 polymorphism and patient age at diagnosis. Neuro-Oncology 2015, 17, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Pirozzi, C.J.; Healy, P.; Reitman, Z.J.; Lipp, E.; Rasheed, B.A.; Yang, R.; Diplas, B.H.; Wang, Z.; Greer, P.K.; et al. Mutations in IDH1, IDH2, and in the TERT promoter define clinically distinct subgroups of adult malignant gliomas. Oncotarget 2014, 5, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Li, G.; Qu, Y.; Wang, M.; Cui, B.; Ji, M.; Shi, B.; Hou, P. TERT promoter mutations and long telomere length predict poor survival and radiotherapy resistance in gliomas. Oncotarget 2016, 7, 8712–8725. [Google Scholar] [PubMed]
- Labussiere, M.; Di Stefano, A.L.; Gleize, V.; Boisselier, B.; Giry, M.; Mangesius, S.; Bruno, A.; Paterra, R.; Marie, Y.; Rahimian, A.; et al. TERT promoter mutations in gliomas, genetic associations and clinico-pathological correlations. Br. J. Cancer 2014, 111, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Puig-Butille, J.A.; Schilling, B.; Livingstone, E.; Potrony, M.; Carrera, C.; Schimming, T.; Moller, I.; Schwamborn, M.; et al. TERT promoter mutation status as an independent prognostic factor in cutaneous melanoma. J. Natl. Cancer Inst. 2014. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, P.S.; Hosen, I.; de Verdier, P.J.; Fallah, M.; Heidenreich, B.; Ryk, C.; Wiklund, N.P.; Steineck, G.; Schadendorf, D.; Hemminki, K.; et al. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by a common polymorphism. Proc. Natl. Acad. Sci. USA 2013, 110, 17426–17431. [Google Scholar] [CrossRef] [PubMed]
- Hurst, C.D.; Platt, F.M.; Knowles, M.A. Comprehensive mutation analysis of the TERT promoter in bladder cancer and detection of mutations in voided urine. Eur. Urol. 2014, 65, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Allory, Y.; Beukers, W.; Sagrera, A.; Flandez, M.; Marques, M.; Marquez, M.; van der Keur, K.A.; Dyrskjot, L.; Lurkin, I.; Vermeij, M.; et al. Telomerase reverse transcriptase promoter mutations in bladder cancer: High frequency across stages, detection in urine, and lack of association with outcome. Eur. Urol. 2014, 65, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Liu, C.; Meng, Y.; Yuan, X.; Liu, L.; Ge, N.; Liu, J.; Wang, C.; Ren, H.; et al. TERT promoter mutations and TERT mrna but not FGFR3 mutations are urinary biomarkers in han chinese patients with urothelial bladder cancer. Oncologist 2015, 20, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Liu, C.; Wang, K.; Liu, L.; Liu, T.; Ge, N.; Kong, F.; Yang, L.; Bjorkholm, M.; Fan, Y.; et al. The genetic difference between western and chinese urothelial cell carcinomas: Infrequent FGFR3 mutation in han chinese patients. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, P.; Li, C.; Huang, Y.; Li, X.; Wang, Y.; Chen, C.; Lv, Z.; Tang, A.; Sun, X.; et al. Telomerase reverse transcriptase gene promoter mutations help discern the origin of urogenital tumors: A genomic and molecular study. Eur. Urol. 2014, 65, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Kurtis, B.; Zhuge, J.; Ojaimi, C.; Ye, F.; Cai, D.; Zhang, D.; Fallon, J.T.; Zhong, M. Recurrent TERT promoter mutations in urothelial carcinoma and potential clinical applications. Ann. Diagn. Pathol. 2016, 21, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Davidson, D.D.; Wang, M.; Lopez-Beltran, A.; Montironi, R.; Wang, L.; Tan, P.H.; MacLennan, G.T.; Williamson, S.R.; Zhang, S. Telomerase reverse transcriptase (TERT) promoter mutation analysis of benign, malignant and reactive urothelial lesions reveals a subpopulation of inverted papilloma with immortalizing genetic change. Histopathology 2015, 69, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.; Xi, L.; Zaug, A.J.; Powell, N.M.; Dancik, G.M.; Cohen, S.B.; Costello, J.C.; Theodorescu, D.; Cech, T.R. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science 2015, 347, 1006–1010. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Tian, W.; Zhuge, J.; Zheng, X.; Huang, T.; Cai, D.; Zhang, D.; Yang, X.J.; Argani, P.; Fallon, J.T.; et al. Distinguishing nested variants of urothelial carcinoma from benign mimickers by TERT promoter mutation. Am. J. Surg. Pathol. 2015, 39, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.A.; Laughlin, T.S.; Rothberg, P.G. Mutations of the TERT promoter are common in basal cell carcinoma and squamous cell carcinoma. Mod. Pathol. 2014, 27, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Schimming, T.; Moller, I.; Moll, I.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Schadendorf, D.; et al. TERT promoter mutations are frequent in cutaneous basal cell carcinoma and squamous cell carcinoma. PLoS ONE 2013, 8, e80354. [Google Scholar] [CrossRef] [PubMed]
- Populo, H.; Boaventura, P.; Vinagre, J.; Batista, R.; Mendes, A.; Caldas, R.; Pardal, J.; Azevedo, F.; Honavar, M.; Guimaraes, I.; et al. TERT promoter mutations in skin cancer: The effects of sun exposure and x-irradiation. J. Investig. Dermatol. 2014, 134, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Liao, S.L.; Hong, J.B.; Chu, C.Y.; Sheen, Y.S.; Jhuang, J.Y.; Tsai, J.H.; Liau, J.Y. TERT promoter mutations in periocular carcinomas: Implications of ultraviolet light in pathogenesis. Br. J. Ophthalmol. 2016, 100, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.A.; Kurtis, B.; Babayeva, S.; Zhuge, J.; Tantchou, I.; Cai, D.; Lafaro, R.J.; Fallon, J.T.; Zhong, M. Heterogeneity of TERT promoter mutations status in squamous cell carcinomas of different anatomical sites. Ann. Diagn. Pathol. 2015, 19, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, Z.; Sun, M.; Chen, K.; Yuan, W.; Jiang, G. The sensitive detection of telomerase reverse transcriptase promoter mutation by amplification refractory mutation system-PCR. Genet. Test. Mol. Biomark. 2016, 20, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bishop, J.; Shan, Y.; Pai, S.; Liu, D.; Murugan, A.K.; Sun, H.; El-Naggar, A.K.; Xing, M. Highly prevalent TERT promoter mutations in aggressive thyroid cancers. Endocr. Relat. Cancer 2013, 20, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Landa, I.; Ganly, I.; Chan, T.A.; Mitsutake, N.; Matsuse, M.; Ibrahimpasic, T.; Ghossein, R.A.; Fagin, J.A. Frequent somatic TERT promoter mutations in thyroid cancer: Higher prevalence in advanced forms of the disease. J. Clin. Endocrinol. Metab. 2013, 98, E1562–E1566. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liu, R.; Qu, S.; Zhu, G.; Bishop, J.; Liu, X.; Sun, H.; Shan, Z.; Wang, E.; Luo, Y.; et al. Association of TERT promoter mutation 1,295,228 c>t with braf v600e mutation, older patient age, and distant metastasis in anaplastic thyroid cancer. J. Clin. Endocrinol. Metab. 2015, 100, E632–E637. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.; da Rocha, A.G.; Vinagre, J.; Batista, R.; Peixoto, J.; Tavares, C.; Celestino, R.; Almeida, A.; Salgado, C.; Eloy, C.; et al. TERT promoter mutations are a major indicator of poor outcome in differentiated thyroid carcinomas. J. Clin. Endocrinol. Metab. 2014, 99, E754–E765. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, N.; Cao, J.; Sofiadis, A.; Dinets, A.; Zedenius, J.; Larsson, C.; Xu, D. The age- and shorter telomere-dependent TERT promoter mutation in follicular thyroid cell-derived carcinomas. Oncogene 2014, 33, 4978–4984. [Google Scholar] [CrossRef] [PubMed]
- Landa, I.; Ibrahimpasic, T.; Boucai, L.; Sinha, R.; Knauf, J.A.; Shah, R.H.; Dogan, S.; Ricarte-Filho, J.C.; Krishnamoorthy, G.P.; Xu, B.; et al. Genomic and transcriptomic hallmarks of poorly differentiated and anaplastic thyroid cancers. J. Clin. Investig. 2016, 126, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Ge, N.; Liu, L.; Yuan, X.; Liu, J.; Kong, F.; Wang, C.; Ren, H.; Yan, K.; et al. TERT promoter mutations are associated with distant metastases in upper tract urothelial carcinomas and serve as urinary biomarkers detected by a sensitive castpcr. Oncotarget 2014, 5, 12428–12439. [Google Scholar] [CrossRef] [PubMed]
- Nault, J.C.; Mallet, M.; Pilati, C.; Calderaro, J.; Bioulac-Sage, P.; Laurent, C.; Laurent, A.; Cherqui, D.; Balabaud, C.; Zucman-Rossi, J. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat. Commun. 2013. [Google Scholar] [CrossRef]
- Cevik, D.; Yildiz, G.; Ozturk, M. Common telomerase reverse transcriptase promoter mutations in hepatocellular carcinomas from different geographical locations. World J. Gastroenterol. 2015, 21, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Jeng, Y.M.; Chang, C.N.; Lee, H.J.; Hsu, H.C.; Lai, P.L.; Yuan, R.H. TERT promoter mutation in resectable hepatocellular carcinomas: A strong association with hepatitis c infection and absence of hepatitis b infection. Int. J. Surg. 2014, 12, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Guo, X.; Chen, Y.; Chen, G.; Ma, Y.; Huang, K.; Zhang, Y.; Zhao, Q.; Winkler, C.A.; An, P.; et al. Telomerase reverse transcriptase promoter mutations in hepatitis B virus-associated hepatocellular carcinoma. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Totoki, Y.; Tatsuno, K.; Covington, K.R.; Ueda, H.; Creighton, C.J.; Kato, M.; Tsuji, S.; Donehower, L.A.; Slagle, B.L.; Nakamura, H.; et al. Trans-ancestry mutational landscape of hepatocellular carcinoma genomes. Nat. Genet. 2014, 46, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- de Unamuno Bustos, B.; Murria Estal, R.; Perez Simo, G.; Oliver Martinez, V.; Llavador Ros, M.; Palanca Suela, S.; Botella Estrada, R. Lack of TERT promoter mutations in melanomas with extensive regression. J. Am. Acad. Dermatol. 2016, 74, 570–572. [Google Scholar] [CrossRef] [PubMed]
- Egberts, F.; Kruger, S.; Behrens, H.M.; Bergner, I.; Papaspyrou, G.; Werner, J.A.; Alkatout, I.; Haag, J.; Hauschild, A.; Rocken, C. Melanomas of unknown primary frequently harbor TERT-promoter mutations. Melanoma Res. 2014, 24, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.K.; Yao, Y.; Zhang, Z.; Chung, N.Y.; Liu, J.S.; Li, K.K.; Shi, Z.; Chan, D.T.; Poon, W.S.; Zhou, L.; et al. TERT promoter mutations contribute to subset prognostication of lower-grade gliomas. Mod. Pathol. 2015, 28, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, A.E.; Ober, K.; Dubbink, H.J.; Paridaens, D.; Naus, N.C.; Belunek, S.; Krist, B.; Post, E.; Zwarthoff, E.C.; de Klein, A.; et al. Prevalence and implications of TERT promoter mutation in uveal and conjunctival melanoma and in benign and premalignant conjunctival melanocytic lesions. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6024–6030. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Scholz, S.; Sucker, A.; Song, M.; Susskind, D.; Grabellus, F.; Zimmer, L.; Hillen, U.; et al. TERT promoter mutations in ocular melanoma distinguish between conjunctival and uveal tumours. Br. J. Cancer 2013, 109, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Nagore, E.; Rachakonda, P.S.; Garcia-Casado, Z.; Requena, C.; Traves, V.; Becker, J.; Soufir, N.; Hemminki, K.; Kumar, R. Telomerase reverse transcriptase promoter mutations in primary cutaneous melanoma. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed]
- Macerola, E.; Loggini, B.; Giannini, R.; Garavello, G.; Giordano, M.; Proietti, A.; Niccoli, C.; Basolo, F.; Fontanini, G. Coexistence of TERT promoter and braf mutations in cutaneous melanoma is associated with more clinicopathological features of aggressiveness. Virchows Arch. 2015, 467, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Qasem, E.; Murugan, A.K.; Al-Hindi, H.; Xing, M.; Almohanna, M.; Alswailem, M.; Alzahrani, A.S. TERT promoter mutations in thyroid cancer: A report from a middle eastern population. Endocr. Relat. Cancer 2015, 22, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Remke, M.; Ramaswamy, V.; Peacock, J.; Shih, D.J.; Koelsche, C.; Northcott, P.A.; Hill, N.; Cavalli, F.M.; Kool, M.; Wang, X.; et al. TERT promoter mutations are highly recurrent in shh subgroup medulloblastoma. Acta Neuropathol. 2013, 126, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, J.C.; Schwalbe, E.C.; Potluri, S.; Bailey, S.; Williamson, D.; Clifford, S.C. TERT promoter mutation and aberrant hypermethylation are associated with elevated expression in medulloblastoma and characterise the majority of non-infant shh subgroup tumours. Acta Neuropathol. 2014, 127, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Kim, Y.H.; Oh, J.E.; Satomi, K.; Nonoguchi, N.; Keyvani, K.; Pierscianek, D.; Sure, U.; Mittelbronn, M.; Paulus, W.; et al. Alterations of the RRAS and ERCC1 genes at 19q13 in gemistocytic astrocytomas. J. Neuropathol. Exp. Neurol. 2014, 73, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Alentorn, A.; Daniau, M.; Labussiere, M.; Rahimian, A.; Tabouret, E.; Polivka, M.; Jouvet, A.; Adam, C.; Figarella-Branger, D.; et al. TERT promoter mutations in primary central nervous system lymphoma are associated with spatial distribution in the splenium. Acta Neuropathol. 2015, 130, 439–440. [Google Scholar] [CrossRef] [PubMed]
- Muzza, M.; Colombo, C.; Rossi, S.; Tosi, D.; Cirello, V.; Perrino, M.; De Leo, S.; Magnani, E.; Pignatti, E.; Vigo, B.; et al. Telomerase in differentiated thyroid cancer: Promoter mutations, expression and localization. Mol. Cell. Endocrinol. 2015, 399, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, T.; Sofiadis, A.; Juhlin, C.C.; Zedenius, J.; Hoog, A.; Larsson, C.; Xu, D. TERT promoter mutation as an early genetic event activating telomerase in follicular thyroid adenoma (FTA) and atypical FTA. Cancer 2014, 120, 2965–2979. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Lim, J.A.; Choi, H.; Won, J.K.; Moon, J.H.; Cho, S.W.; Lee, K.E.; Park, Y.J.; Yi, K.H.; Park do, J.; et al. Prognostic effects of TERT promoter mutations are enhanced by coexistence with BRAF or RAS mutations and strengthen the risk prediction by the ATA or TNM staging system in differentiated thyroid cancer patients. Cancer 2016, 122, 1370–1379. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.S.; Kim, Y.; Jeon, S.; Kim, S.H.; Kim, T.J.; Lee, S.; Kim, M.H.; Lim, D.J.; Lee, Y.S.; Jung, C.K. Clinical utility of TERT promoter mutations and ALK rearrangement in thyroid cancer patients with a high prevalence of the BRAF V600E mutation. Diagn. Pathol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.N.; Chiang, Y.C.; Cheng, W.F.; Chen, C.A.; Lin, M.C.; Kuo, K.T. Molecular alterations in endometrial and ovarian clear cell carcinomas: Clinical impacts of telomerase reverse transcriptase promoter mutation. Mod. Pathol. 2015, 28, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.C.; Ayhan, A.; Maeda, D.; Kim, K.R.; Clarke, B.A.; Shaw, P.; Chui, M.H.; Rosen, B.; Shih Ie, M.; Wang, T.L. Frequent somatic mutations of the telomerase reverse transcriptase promoter in ovarian clear cell carcinoma but not in other major types of gynaecological malignancy. J. Pathol. 2014, 232, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Chindris, A.M.; Casler, J.D.; Bernet, V.J.; Rivera, M.; Thomas, C.; Kachergus, J.M.; Necela, B.M.; Hay, I.D.; Westphal, S.A.; Grant, C.S.; et al. Clinical and molecular features of hurthle cell carcinoma of the thyroid. J. Clin. Endocrinol. Metab. 2015, 100, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Tallet, A.; Nault, J.C.; Renier, A.; Hysi, I.; Galateau-Salle, F.; Cazes, A.; Copin, M.C.; Hofman, P.; Andujar, P.; Le Pimpec-Barthes, F.; et al. Overexpression and promoter mutation of the TERT gene in malignant pleural mesothelioma. Oncogene 2014, 33, 3748–3752. [Google Scholar] [CrossRef] [PubMed]
- Akaike, K.; Kurisaki-Arakawa, A.; Hara, K.; Suehara, Y.; Takagi, T.; Mitani, K.; Kaneko, K.; Yao, T.; Saito, T. Distinct clinicopathological features of NAB2-STAT6 fusion gene variants in solitary fibrous tumor with emphasis on the acquisition of highly malignant potential. Hum. Pathol. 2015, 46, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Liau, J.Y.; Tsai, J.H.; Jeng, Y.M.; Chu, C.Y.; Kuo, K.T.; Liang, C.W. TERT promoter mutation is uncommon in acral lentiginous melanoma. J. Cutan. Pathol. 2014, 41, 504–508. [Google Scholar] [CrossRef] [PubMed]
- George, J.R.; Henderson, Y.C.; Williams, M.D.; Roberts, D.B.; Hei, H.; Lai, S.Y.; Clayman, G.L. Association of TERT promoter mutation, but not BRAF mutation, with increased mortality in PTC. J. Clin. Endocrinol. Metab. 2015, 100, E1550–E1559. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Liu, R.; Liu, X.; Murugan, A.K.; Zhu, G.; Zeiger, M.A.; Pai, S.; Bishop, J. BRAF V600E and TERT promoter mutations cooperatively identify the most aggressive papillary thyroid cancer with highest recurrence. J. Clin. Oncol. 2014, 32, 2718–2726. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Xing, M. Diagnostic and prognostic TERT promoter mutations in thyroid fine-needle aspiration biopsy. Endocr. Relat. Cancer 2014, 21, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qu, S.; Liu, R.; Sheng, C.; Shi, X.; Zhu, G.; Murugan, A.K.; Guan, H.; Yu, H.; Wang, Y.; et al. TERT promoter mutations and their association with BRAF V600E mutation and aggressive clinicopathological characteristics of thyroid cancer. J. Clin. Endocrinol. Metab. 2014, 99, E1130–E1136. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, M.S.; Schmitt, A.; Steinert, H.; Capper, D.; Moch, H.; Komminoth, P.; Perren, A. Tall cell papillary thyroid carcinoma: New diagnostic criteria and mutations in braf and TERT. Endocr. Relat. Cancer 2015, 22, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Chen, E.; Dong, S.; Cai, Y.; Zhang, X.; Zhou, Y.; Zeng, R.; Yang, F.; Pan, C.; Liu, Y.; et al. BRAF and TERT promoter mutations in the aggressiveness of papillary thyroid carcinoma: A study of 653 patients. Oncotarget 2016, 7, 18346–18355. [Google Scholar] [CrossRef] [PubMed]
- Myung, J.K.; Kwak, B.K.; Lim, J.A.; Lee, M.C.; Kim, M.J. TERT Promoter mutations and tumor persistence/recurrence in papillary thyroid cancer. Cancer Res. Treat. 2016, 48, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.J.; Kim, W.G.; Sim, S.; Lim, S.; Kwon, H.; Kim, T.Y.; Shong, Y.K.; Kim, W.B. Low prevalence of somatic TERT promoter mutations in classic papillary thyroid carcinoma. Endocrinol. Metab. 2016, 31, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Bullock, M.; Ren, Y.; O’Neill, C.; Gill, A.; Aniss, A.; Sywak, M.; Sidhu, S.; Delbridge, L.; Learoyd, D.; de Vathaire, F.; et al. TERT promoter mutations are a major indicator of recurrence and death due to papillary thyroid carcinomas. Clin. Endocrinol. 2016, 85, 283–190. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Shi, L.; Wang, D.; Zhang, B.; Yang, Q.; Ji, M.; Shi, B.; Hou, P. Low frequency of TERT promoter mutations in a large cohort of gallbladder and gastric cancers. Int. J. Cancer 2014, 134, 2993–2994. [Google Scholar] [CrossRef] [PubMed]
- Papathomas, T.G.; Oudijk, L.; Zwarthoff, E.C.; Post, E.; Duijkers, F.A.; van Noesel, M.M.; Hofland, L.J.; Pollard, P.J.; Maher, E.R.; Restuccia, D.F.; et al. Telomerase reverse transcriptase promoter mutations in tumors originating from the adrenal gland and extra-adrenal paraganglia. Endocr. Relat. Cancer 2014, 21, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Brown, T.C.; Juhlin, C.C.; Andreasson, A.; Wang, N.; Backdahl, M.; Healy, J.M.; Prasad, M.L.; Korah, R.; Carling, T.; et al. The activating TERT promoter mutation C228T is recurrent in subsets of adrenal tumors. Endocr. Relat. Cancer 2014, 21, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, T.; Liu, L.; Liu, J.; Liu, C.; Wang, C.; Ge, N.; Ren, H.; Yan, K.; Hu, S.; et al. TERT promoter mutations in renal cell carcinomas and upper tract urothelial carcinomas. Oncotarget 2014, 5, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Jangard, M.; Zebary, A.; Ragnarsson-Olding, B.; Hansson, J. TERT promoter mutations in sinonasal malignant melanoma: A study of 49 cases. Melanoma Res. 2015, 25, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Barnhill, R.L.; Dummer, R.; Dalton, J.; Wu, J.; Pappo, A.; Bahrami, A. TERT promoter mutations are predictive of aggressive clinical behavior in patients with spitzoid melanocytic neoplasms. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Goutagny, S.; Nault, J.C.; Mallet, M.; Henin, D.; Rossi, J.Z.; Kalamarides, M. High incidence of activating TERT promoter mutations in meningiomas undergoing malignant progression. Brain Pathol. 2014, 24, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Sahm, F.; Schrimpf, D.; Olar, A.; Koelsche, C.; Reuss, D.; Bissel, J.; Kratz, A.; Capper, D.; Schefzyk, S.; Hielscher, T.; et al. TERT promoter mutations and risk of recurrence in meningioma. J. Natl. Cancer Inst. 2016. [Google Scholar] [CrossRef]
- Carcano, F.M.; Vidal, D.O.; van Helvoort Lengert, A.; Neto, C.S.; Queiroz, L.; Marques, H.; Baltazar, F.; da Silva Martinelli, C.M.; Soares, P.; da Silva, E.C.; et al. Hotspot TERT promoter mutations are rare events in testicular germ cell tumors. Tumour Biol. 2016, 37, 4901–4907. [Google Scholar] [CrossRef]
- Campanella, N.C.; Celestino, R.; Pestana, A.; Scapulatempo-Neto, C.; de Oliveira, A.T.; Brito, M.J.; Gouveia, A.; Lopes, J.M.; Guimaraes, D.P.; Soares, P.; et al. Low frequency of TERT promoter mutations in gastrointestinal stromal tumors (GISTs). Eur. J. Hum. Genet. 2015, 23, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Akaike, K.; Toda-Ishii, M.; Suehara, Y.; Mukaihara, K.; Kubota, D.; Mitani, K.; Takagi, T.; Kaneko, K.; Yao, T.; Saito, T. TERT promoter mutations are a rare event in gastrointestinal stromal tumors. Springerplus 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gao, Y.; Chen, Z.; Hu, X.; Zhou, F.; He, J. Low frequency of TERT promoter somatic mutation in 313 sporadic esophageal squamous cell carcinomas. Int. J. Cancer 2014, 134, 493–494. [Google Scholar] [CrossRef]
- Dono, M.; Angelini, G.; Cecconi, M.; Amaro, A.; Esposito, A.I.; Mirisola, V.; Maric, I.; Lanza, F.; Nasciuti, F.; Viaggi, S.; et al. Mutation frequencies of GNAQ, GNA11, BAP1, SF3B1, EIF1AX and TERT in uveal melanoma: Detection of an activating mutation in the TERT gene promoter in a single case of uveal melanoma. Br. J. Cancer 2014, 110, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Hosen, I.; Rachakonda, P.S.; Heidenreich, B.; de Verdier, P.J.; Ryk, C.; Steineck, G.; Hemminki, K.; Kumar, R. Mutations in TERT promoter and FGFR3 and telomere length in bladder cancer. Int. J. Cancer 2015, 137, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Dang, S.; Wu, K.; Shao, Y.; Yang, Q.; Ji, M.; Shi, B.; Hou, P. TERT promoter mutations predict worse survival in laryngeal cancer patients. Int. J. Cancer 2014, 135, 1008–1010. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Ruden, M.; Puri, N. Novel anticancer therapeutics targeting telomerase. Cancer Treat. Rev. 2013, 39, 444–456. [Google Scholar] [CrossRef] [PubMed]
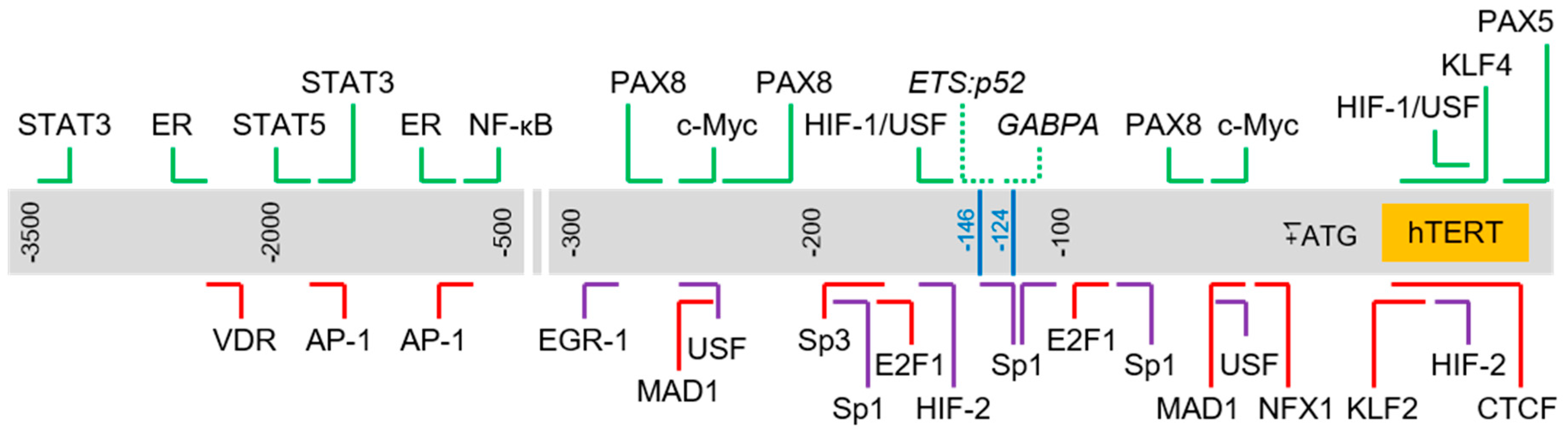
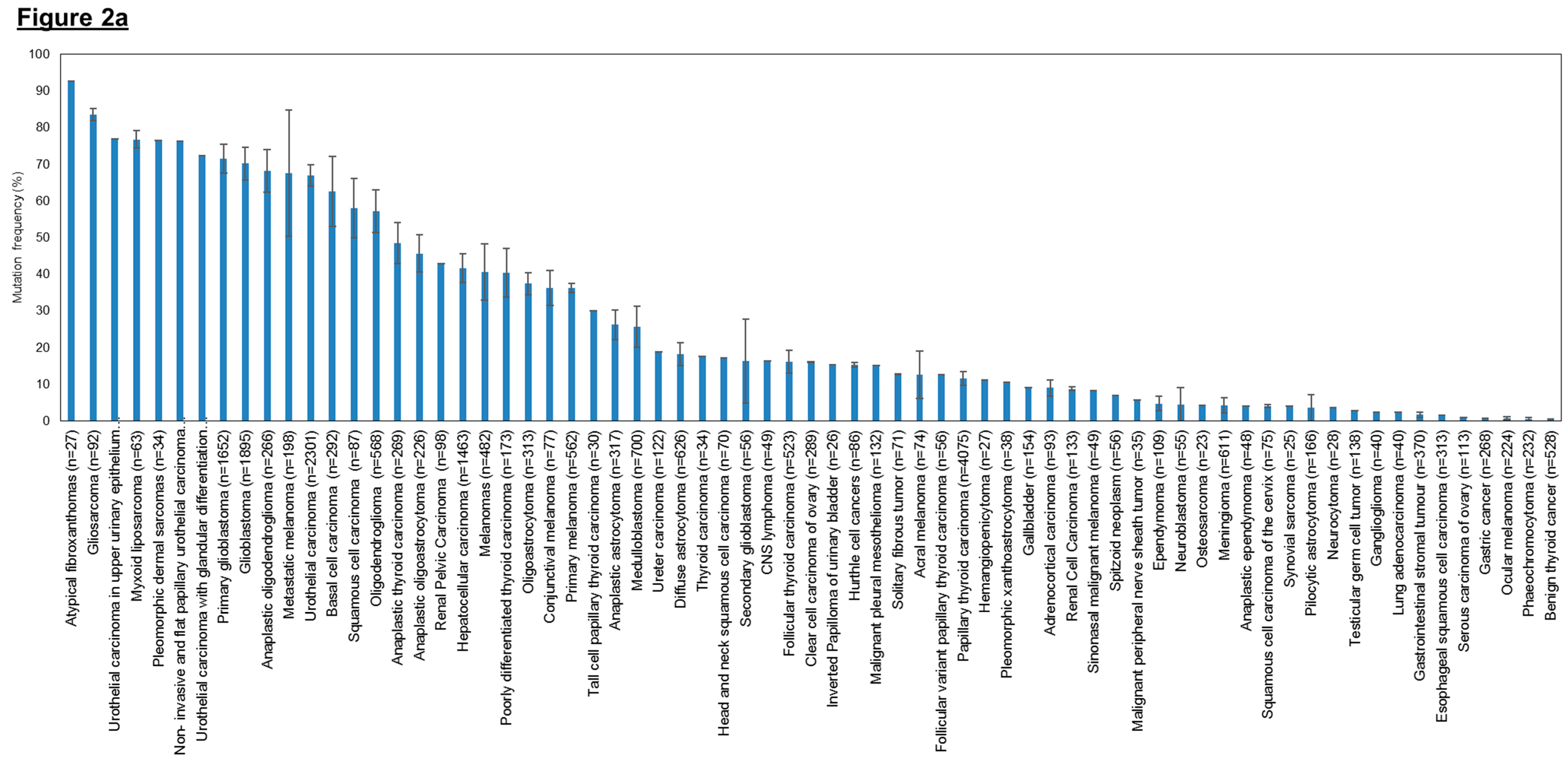
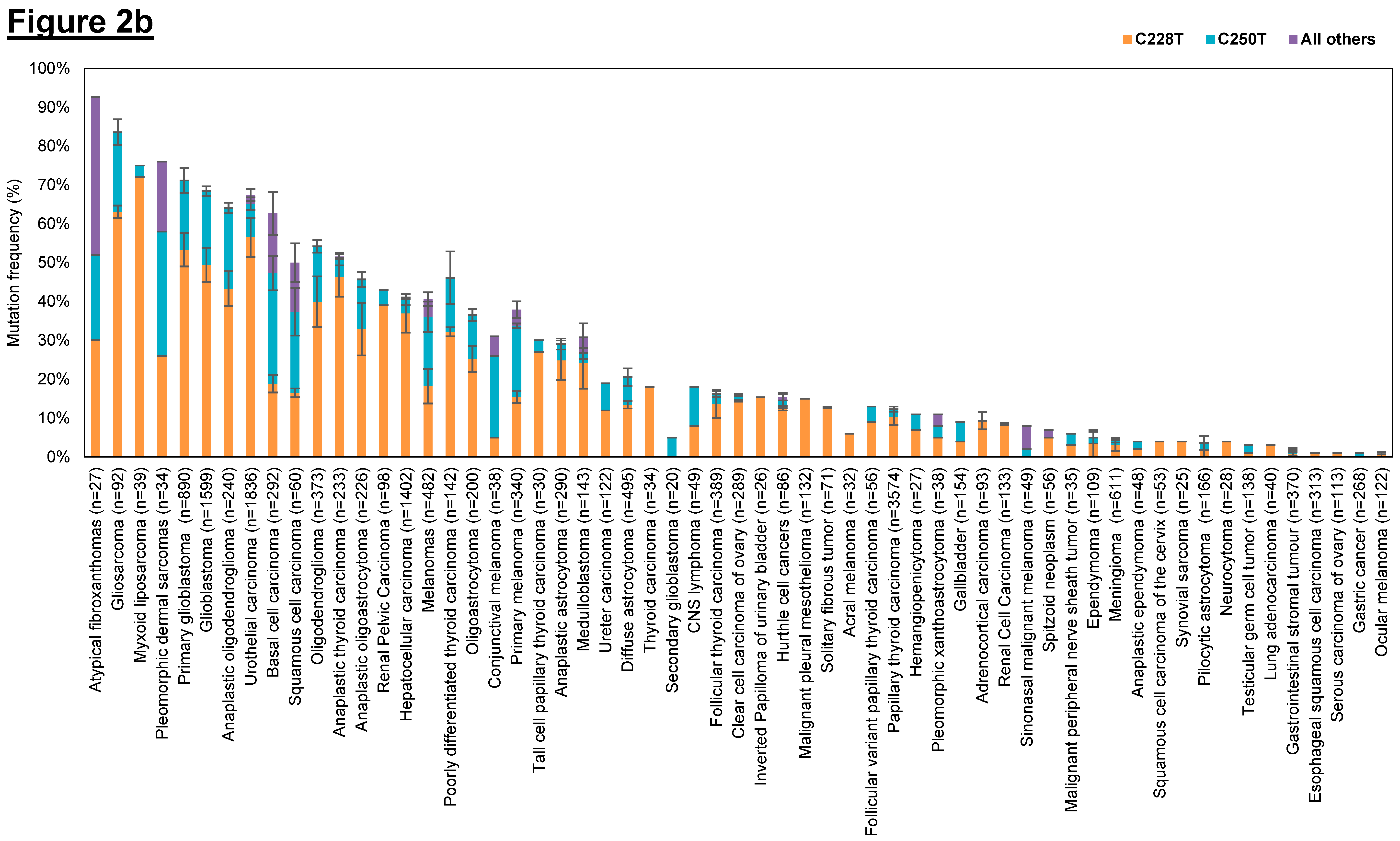

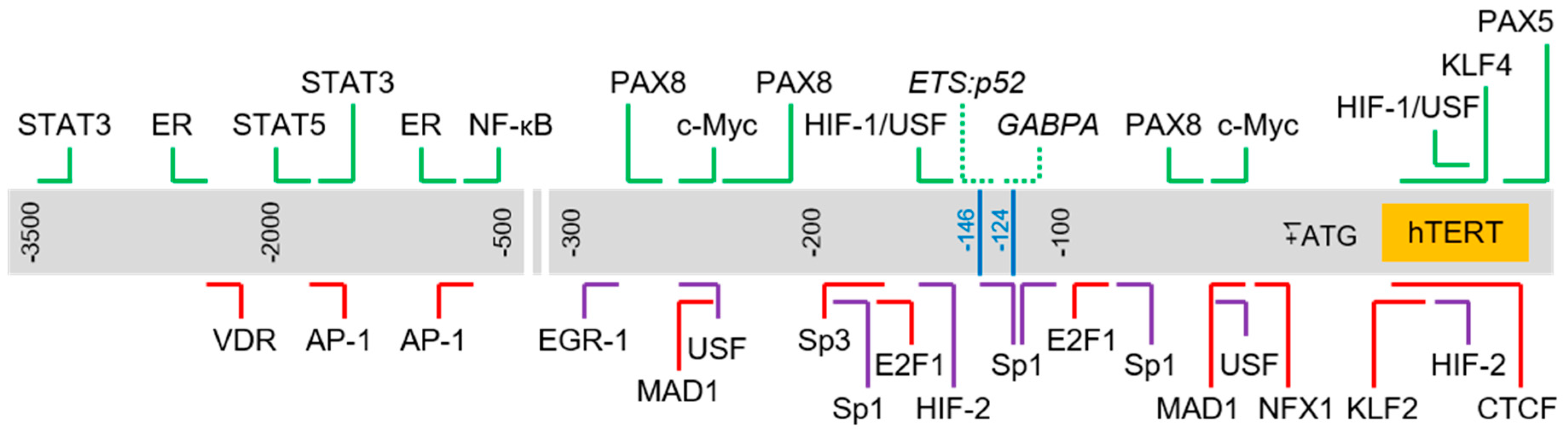{kind=link}
{kind=link}
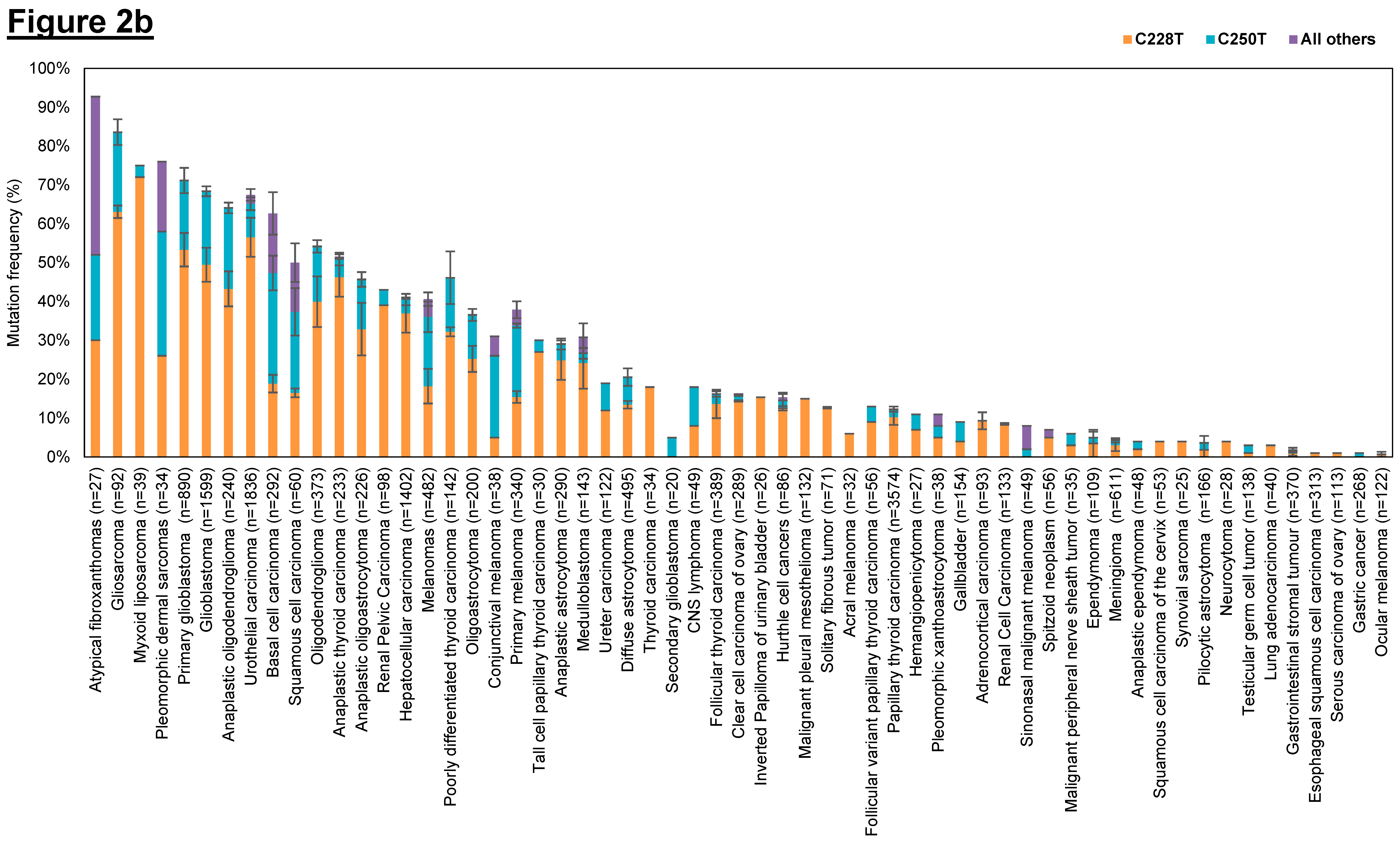{kind=link}
| Regulator | Activator/Repressor | Cell Tested | Binding Site | Binding Assay | Reference | Co-Regulator | Positive Regulators | Negative Regulators |
|---|---|---|---|---|---|---|---|---|
| Androgen receptor | Repressor | LNCaP, PC3 | ND | ChIP | [33] | |||
| AP-1 | Repressor | HeLa | −1655; −718 | EMSA, ChIP | [34] | WT1 [35] | ||
| Activator | HeLa | −378/+1 | ChIP | [36] | HBZ/Sp1 [36] | |||
| AP-2β | Activator | H1299, H322, HBE, WI-38 | −43/−18 | EMSA, ChIP | [37] | BBR [38]; WT1 [35] | ||
| ARID1A | Repressor | OVISE | ND | ChIP | [39] | SIN3A [39] | ||
| β-catenin | Activator | NTera2, SW480, 293T, HCT116, MCF7 | −255/+40; −659/−653 | EMSA, ChIP | [40,41] | Klf4 [40] | TCF1 [40] | |
| Cbfa1 | Repressor | BMSSC | −888/−865; −2815/−2792 | EMSA, ChIP | [42] | |||
| c-Myc | Activator | U937, HL60, A549 | −242/−237; −34/−29 | EMSA, ChIP | [43,44,45] | Sp1 [46];E6 [47]; NFX1-123 [48]; p300 [49];E7 [50]; Max [51]; Ets2 [52] | E6 [53,54,55]; Max [44]; BRCA1/Nmi [56]; NF-κB [57,58]; FOXO3a [59]; MMP-9 [60]; Tax [58]; estrogen [51]; Aurora-A [61]; LMP1 [62]; survivin [63]; PKCθ [64]; SMYD3 [65]; STAGA complex [66]; EGF [67]; CSF1R [68]; GSK3 [69]; DJ-1 [70]; leptin [71]; BDNF [72]; SIRT1 [73]; saquinavir [74]; bile acids [75]; IRF-4 [76] | HIF-1α [77,78,79,80]; BRCA1 [81,82]; cordyceptin [83]; 15d-PGJ2 [84,85]; rosiglitazone [85]; Mad [57,86,87]; ceramide [45]; p53 [88]; Tax [89]; 5-aza-CR [90]; DAC [91]; TMPyP4 [92]; p27KIP1 [93]; genistein [94]; reFIP-gts [95]; 4-hydroxynonena [96]; gambogic acid [97]; butein [98]; wogonin [99]; WT1 [35]; E2F1 [100]; ATO [101]; DADS [102] |
| COUP-TFII | Repressor | HeLa | −201/+35 | EMSA | [103] | |||
| CPSF4 | Activator | H1299, A549, H322, WI-38, HBE | −378/+60 | ChIP | [104] | |||
| CTCF | Repressor | HeLa, SW480, BJ | +16/+126 | EMSA, ChIP | [105] | ROS [106]; TGF-β [107] | Sulforaphane [108] | |
| DEK | Repressor | HeLa | ND | ChIP | [109] | Tax [109] | ||
| E2F1 | Repressor | SCC25 | −174/−170; −98/−94 | EMSA | [110] | TGFβ/Smad3 [111]; PPARα/p16 [112] | ||
| E6 | Activator | HK | ND | ChIP | [113] | c-Myc [113] | ||
| EGR-1 | Activator | JAR, JEG-3 | −281/−273 | EMSA | [114] | |||
| Repressor | CaSki, SiHa | −281/−273 | EMSA | [115] | ||||
| eNOS | Activator | HUVEC | −949/−935 | ChIP | [116] | ERα [116] | ||
| ER81 | Activator | 293T | +288/+291; +390/+393 | EMSA | [117] | HER2/Neu, ERK/MAPK, Ras/Raf [117] | ||
| Estrogen receptor | Activator | MCF7, SiHa, NHK, MDA-MB231, HeLa, OVCA-433, HUVEC | −2677; −949/−935 | EMSA, ChIP | [51,116,118] | eNOS [116] | Sp1 [118]; MAPK [119]; leptin [120] | I3C [121]; 15d-PGJ2 [84]; progesterone [122]; miR-498 [120] |
| ETS | Activator | A549, H1299, MCF7, NIH3T3, A-431, MCF7, ME180 | −246/−243; −99/−96; −23/−13 | EMSA, ChIP | [52,123,124,125] | c-Myc [52]; EWS [126] | EGF/MAPK [123,124] | PPARγ [127] |
| Repressor | U937, K562 | −351 | EMSA | [125] | WT1 [35] | |||
| GLI1/2 | Activator | HT29, 293T | −1226/+438 | ChIP | [128] | |||
| GRHL2 | Activator | SCC4, NHOK, SCC15 | −21/−19 | ChIP, PMP | [129,130] | |||
| HIF-1 | Activator | ME180, JEG-3, JAR | −165/−158; +44/+51 | EMSA, ChIP | [131,132] | LPA [133]; PI3K [133] | ||
| HIF-2 | Activator | A498 | −165/−158; +44/+51 | ChIP | [134] | |||
| Repressor | U251 | −165/−158; +44/+51 | ChIP | [134] | ||||
| hnRNP D | Activator | NHOK, SCC15, SCC4 | −188/−42 | ChIP, PMP | [130] | |||
| hnRNP K | Activator | NHOK, SCC15, SCC4 | −188/−42 | [130] | ||||
| Hsp90 | Activator | SCC4 | −465/−341; −188/+5 | ChIP | [135] | |||
| KLF2 | Repressor | Kit 225, primary human T cells | +9/+30 | EMSA, ChIP | [136] | |||
| KLF4 | Activator | FaDu | +18/+77 | EMSA, ChIP | [137] | |||
| LSD1 | Repressor | HL60 | ND | ChIP | [138] | |||
| MAD1 | Repressor | WI38, 293T, U937 | −243/−238; −34/−29 | EMSA, ChIP | [86,139] | Max [140] | Sulforaphane [108]; PPARγ ligands [85]; DADS [102] | miR-202 [140]; MMP-9 [60]; c-Myc [57,86]; miR-202 [140] |
| Maz | Repressor | HFK | ND | ChIP | [141] | HBx [142] | E6 [141] | |
| MCPH1 | Repressor | HeLa | +4/+68 | EMSA | [143] | |||
| Menin | Repressor | MCF-7, C33A, HeLa | Sequence-independent | EMSA | [144] | JunD [145] | HBZ [145] | |
| MSH2 | Activator | NHOK, SCC15, SCC4 | −377/−207 | ChIP, PMP | [130] | |||
| MZF-2 | Repressor | C33A, SiHa, HeLa, ME100, K562 | −687/−680 | EMSA | [146] | |||
| NF-κB | Activator | U937, SH-SY5Y, SK-N-MC | −650/−638 | ChIP, EMSA | [147,148] | STAT3 [149]; Tax [58,150]; PI3K/Akt [151] | ATO [101]; IκBα [152]; curcumin [148]; pelitinib [153] | |
| NFAT1 | Activator | MCF7 | −775/−771; −40/−36 | ChIP | [154] | Sp1 [154] | ||
| NFX1 | Repressor | HFK | −28/−19 | EMSA | [48] | Sin3A/HDAC [155] | E6/E6-AP [48] | |
| Activator | HFK | −28/−19 | EMSA | [48] | c-Myc [48] | E6 [55] | WT1 [35] | |
| PAX5 | Activator | Raji, Nalm6 | +110/+137; +489/+516 | EMSA: ChIP | [156] | |||
| PAX8 | Activator | LN18, SF268, U87MG | −272/−268; −236/−219; −217/−202; −57/−41 | EMSA | [157] | |||
| PITX1 | Repressor | A2058 | −1366/−1361; −1347/−1342; −1325/−1320 | EMSA, ChIP | [158] | miR-19b [159] | ||
| PreS2 | Activator | HepG2.2.15 | −407/−387 | EMSA | [160] | |||
| Reptin | Activator | AGS, HGC-27 | ND | ChIP | [161] | c-Myc [161] | ||
| RFPL3 | Activator | H1299, A549, WI-38, HBE, | ND | ChIP | [162] | CBP [163] | ||
| SNAI1 | Repressor | HaCaT, HEK293 | −242/−237; −34/−29 | ChIP | [80] | TGFβ [80] | ||
| Sp1 | Activator | C33A, ME180, SiHa, HeLa | −182; −160; −128; −108; −79 | EMSA | [30,46] | CBP [164]; c-Myc [46] | E6 [53,141]; HBZ/JunD [36]; LANA [165,166]; Tax/NF-κB [58]; genistein [167]; survivin [63]; BDNF [72]; PKCθ/NF-κB [64]; SMYD3 [65]; IRF-4 [76]; MCAF1 [168]; NFAT [154]; HBx [169]; HMGA2 [170] | p53 [171]; p73 [172,173]; cordyceptin [83]; 15d-PGJ2 [84]; p27KIP1 [93]; arsenic/ROS [25]; ceramide ([174]; ATO [101]; DADS [102]; E2 [175]; E2F-1 [176]; p16 [177]; butylidenephthalide [178]; indole-3-carbinol [121]; nilotinib/dasatinib [179]; CDDO-Me [180]; homocysteine [181]; triptolide [182] |
| Repressor | IMR90, WI38, HFF | −182; −160; −128; −108; −79 | EMSA, ChIP | [183] | HDAC [183]; CtBP [184] | TAK1 [185] | E1A [184] | |
| Sp3 | Repressor | A549, IMR90, WI38, HFF | −195/−168 | EMSA, ChIP | [174,183,186] | Ceramide [174] | ||
| SPT5 | Activator | SW620, HT29, Colo320, RKO, HCE8693 | −378/+60 | ChIP | [187] | |||
| STAT3 | Activator | HepG2, MCF-7, DU-145, K562, A172, HS27 | −3308/−3300; −1587/−1579 | ChIP | [71,167,188,189,190] | miR-21 [191]; leptin [71,188]; GSK3 [69]; HCVc [192] | ||
| STAT5 | Activator | ILT-Hod, K562-ADM, K562 | −1872/−1864 | ChIP | [189,193,194] | IL-2/JAK [193]; EPO [195,196] | ||
| TAL1 | Repressor | HeLa | ND | ChIP | [197] | Tax [197] | HBZ/JunD [197] | |
| TEIF | Activator | HeLa, 293, HT1080, 293T | −120/+90 | EMSA | [198,199] | MSP58 [199] | ||
| USF | Activator | SKBR3, MDA-MB-231, MCF7, OVCAR3, 293T, SKOV3, Wi38, HFF, BJ, lymphocytes, HEK293 | −242/−237; −165/−160; −34/−29; +44/+49 | EMSA | [200,201] | p300, p38/MAPK [200]; IRF-4 [76] | Truncated USF2 [201]; ATO [101] | |
| Repressor | OEC-M1, HFK | −242/−237; −34/−29 | EMSA, ChIP | [54,202] | ||||
| VDR | Repressor | PC3, LNCaP | −2530/−2516 | EMSA | [203] | RXR [203] | ||
| WT1 | Repressor | 293T | −423/−307 | EMSA | [204] | PI3K pathway [150] |
| Region Tested | Methylation Status | Cell Line/Tissue | Technique to Detect Methylation | Reference |
|---|---|---|---|---|
| −500 to +50 (72 CpG sites) | Complete | Telomerase-positive (CMV, SUSM-1) | Bisulfite genomic sequencing | [238] |
| −500 to +50 (72 CpG sites) | Partial | Telomerase-positive (HTB 182, HTB178, CaLu1, CaLu3, CaLu6, HTB57, HCT 116) | Bisulfite genomic sequencing | [238] |
| −500 to +50 (72 CpG sites) | Unmethylated | Telomerase-positive (A549, HTB183) and telomerase-negative (NHF, MRC-5 p27) | Bisulfite genomic sequencing | [238] |
| Promoter region | Unmethylated | Telomerase-negative (WI38, HA-1 pre-crisis cell strain, JFCF-6T/5K pre-crisis cell strain, IMR90, BJ fibroblast, telomerase-negative adrenal carcinoma) and telomerase-positive (CT1485) | Methylation-specific PCR-based assay | [243] |
| Promoter region | Partial or complete | U2OS, GM847, VA13, telomerase-negative breast carcinoma, Co1310 | Methylation-specific PCR-based assay | [243] |
| −441 to +218; with respect to ATG (27 CpG sites) | Hypo- or unmethylated | Telomerase-negative tissue(bladder, brain, heart, kidney, muscle, placenta, skin, testis) | Bisulfite genomic sequencing | [244] |
| −441 to +218; with respect to ATG (27 CpG sites) | Hypermethylated | Telomerase-positive (MCF-7, A431, HeLa, Co115, HT29, SW480, HS520, SW2, PC3, Saos-2, U2-OS) and two paraffin-embedded fixed tumor tissue (colon, kidney) | Bisulfite genomic sequencing | [244] |
| −100 and +100 | 75% to 100% methylated | HeLa, SW480, 8 clones from tumor tissues (breast, bladder and cervix) | Bisulfite genomic sequencing | [105] |
| −100 and +100 | Partial | Telomerase-negative( BJ, HLF) | Bisulfite genomic sequencing | [105] |
| −80 to −165 | Unmethylated | Telomerase-positive (HeLa, SW480) and telomerase negative (BJ, HLF) | Bisulfite genomic sequencing | [105] |
| −600 region | Complete or partial | Caco-2, HCT116, RKO, SW480, MCF7, MDA-MB-231, MDA-MB-435S, MDA-MB-453, H82, H157, H209, H146, H358, H417, H549, H747, H1299, U1752, DMS53, HL-60, KG-1a, Jurkat, Raji, LCL, VA13 | Methylation-specific PCR and bisulfite sequencing | [240] |
| TSS region | Complete, partial or unmethylated | |||
| Promoter region (27 CpG sites) | Hypermethylated | Downregulated hTERT expression in 12 HCC samples and normal tissue | Bisulfite genomic sequencing | [239] |
| Promoter region (27 CpG sites) | Unmethylated | Downregulated hTERT expression in HCC samples | Bisulfite genomic sequencing | [239] |
| Upstream of the transcription start site (UTSS) | Hypermethylated (5 sites) | Malignant pediatric brain tumor samples | Methylation arrays | [245] |
| Upstream of the transcription start site (UTSS) | Densely methylated | Neuroblastoma samples | Methylation arrays | [246] |
| Cancer type | References |
|---|---|
| Atypical fibroxanthomas | [256] |
| Gliosarcoma | [257,258,259] |
| Urothelial carcinoma in upper urinary epithelium | [260] |
| Myxoid liposarcoma | [253,261] |
| Pleomorphic dermal sarcomas | [256] |
| Non- invasive and flat papillary urothelial carcinoma | [255] |
| Urothelial carcinoma with glandular differentiation | [262] |
| Primary glioblastoma | [253,259,260,263,264,265,266,267,268] |
| Glioblastoma | [260,269,270,271,272,273,274,275,276,277,278] |
| Anaplastic oligodendroglioma | [253,259,264,270,272,278] |
| Metastatic melanoma | [252,279] |
| Urothelial carcinoma | [253,255,260,262,269,270,271,280,281,282,283,284,285,286,287,288,289] |
| Basal cell carcinoma | [290,291,292,293] |
| Squamous cell carcinoma | [290,291,294] |
| Oligodendroglioma | [259,264,265,268,270,272,276,277,278,295] |
| Anaplastic thyroid carcinoma | [296,297,298,299,300,301] |
| Anaplastic oligoastrocytoma | [259,264,278] |
| Renal Pelvic Carcinoma | [284,302] |
| Hepatocellular carcinoma | [253,254,260,303,304,305,306,307] |
| Melanomas | [251,270,271,292,308,309] |
| Poorly differentiated thyroid carcinoma | [297,299,301] |
| Oligoastrocytoma | [264,268,276,277,278,310] |
| Conjunctival melanoma | [311,312] |
| Primary melanoma | [252,279,313,314] |
| Tall cell papillary thyroid carcinoma | [315] |
| Anaplastic astrocytoma | [253,259,264,278,310] |
| Medulloblastoma | [253,259,316,317] |
| Ureter carcinoma | [302] |
| Diffuse astrocytoma | [259,260,264,270,272,277,310,318] |
| Thyroid carcinoma | [271] |
| Head and neck squamous cell carcinoma | [276] |
| Secondary glioblastoma | [263] [259] |
| CNS lymphoma | [319] |
| Follicular thyroid carcinoma | [270,296,299,300,320,321,322,323] |
| Clear cell carcinoma of ovary | [324,325] |
| Inverted Papilloma of urinary bladder | [287] |
| Hurthle cell cancers | [297,326] |
| Malignant pleural mesothelioma | [327] |
| Solitary fibrous tumor | [261,328] |
| Acral melanoma | [279,329] |
| Follicular variant papillary thyroid carcinoma | [315] |
| Papillary thyroid carcinoma | [270,296,297,299,300,315,320,322,323,330,331,332,333,334,335,336,337,338] |
| Hemangiopenicytoma | [259] |
| Pleomorphic xanthoastrocytoma | [259] |
| Gallbladder | [339] |
| Adrenocortical carcinoma | [285,340,341] |
| Renal Cell Carcinoma | [342] |
| Sinonasal malignant melanoma | [343] |
| Spitzoid neoplasm | [344] |
| Malignant peripheral nerve sheath tumor | [261] |
| Ependymoma | [253,259] |
| Neuroblastoma | [253,340] |
| Osteosarcoma | [253] |
| Meningioma | [253,259,260,345,346] |
| Anaplastic ependymoma | [259] |
| Squamous cell carcinoma of the cervix | [253,325] |
| Synovial sarcoma | [261] |
| Pilocytic astrocytoma | [259,272] |
| Neurocytoma | [259] |
| Testicular germ cell tumor | [347] |
| Ganglioglioma | [259] |
| Lung adenocarcinoma | [271] |
| Gastrointestinal stromal tumor | [260,270,340,348,349] |
| Esophageal squamous cell carcinoma | [350] |
| Serous carcinoma of ovary | [325] |
| Gastric cancer | [339] |
| Ocular melanoma | [270,311,312,351] |
| Phaeochromocytoma | [340,341] |
| Benign thyroid cancer | [270,296,299,321,332,333] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramlee, M.K.; Wang, J.; Toh, W.X.; Li, S. Transcription Regulation of the Human Telomerase Reverse Transcriptase (hTERT) Gene. Genes 2016, 7, 50. https://doi.org/10.3390/genes7080050
Ramlee MK, Wang J, Toh WX, Li S. Transcription Regulation of the Human Telomerase Reverse Transcriptase (hTERT) Gene. Genes. 2016; 7(8):50. https://doi.org/10.3390/genes7080050
Chicago/Turabian StyleRamlee, Muhammad Khairul, Jing Wang, Wei Xun Toh, and Shang Li. 2016. "Transcription Regulation of the Human Telomerase Reverse Transcriptase (hTERT) Gene" Genes 7, no. 8: 50. https://doi.org/10.3390/genes7080050
APA StyleRamlee, M. K., Wang, J., Toh, W. X., & Li, S. (2016). Transcription Regulation of the Human Telomerase Reverse Transcriptase (hTERT) Gene. Genes, 7(8), 50. https://doi.org/10.3390/genes7080050