
Wool Keratin-Associated Protein Genes in Sheep—A Review

 ,
,  , ,
, , 
Abstract
1. Introduction
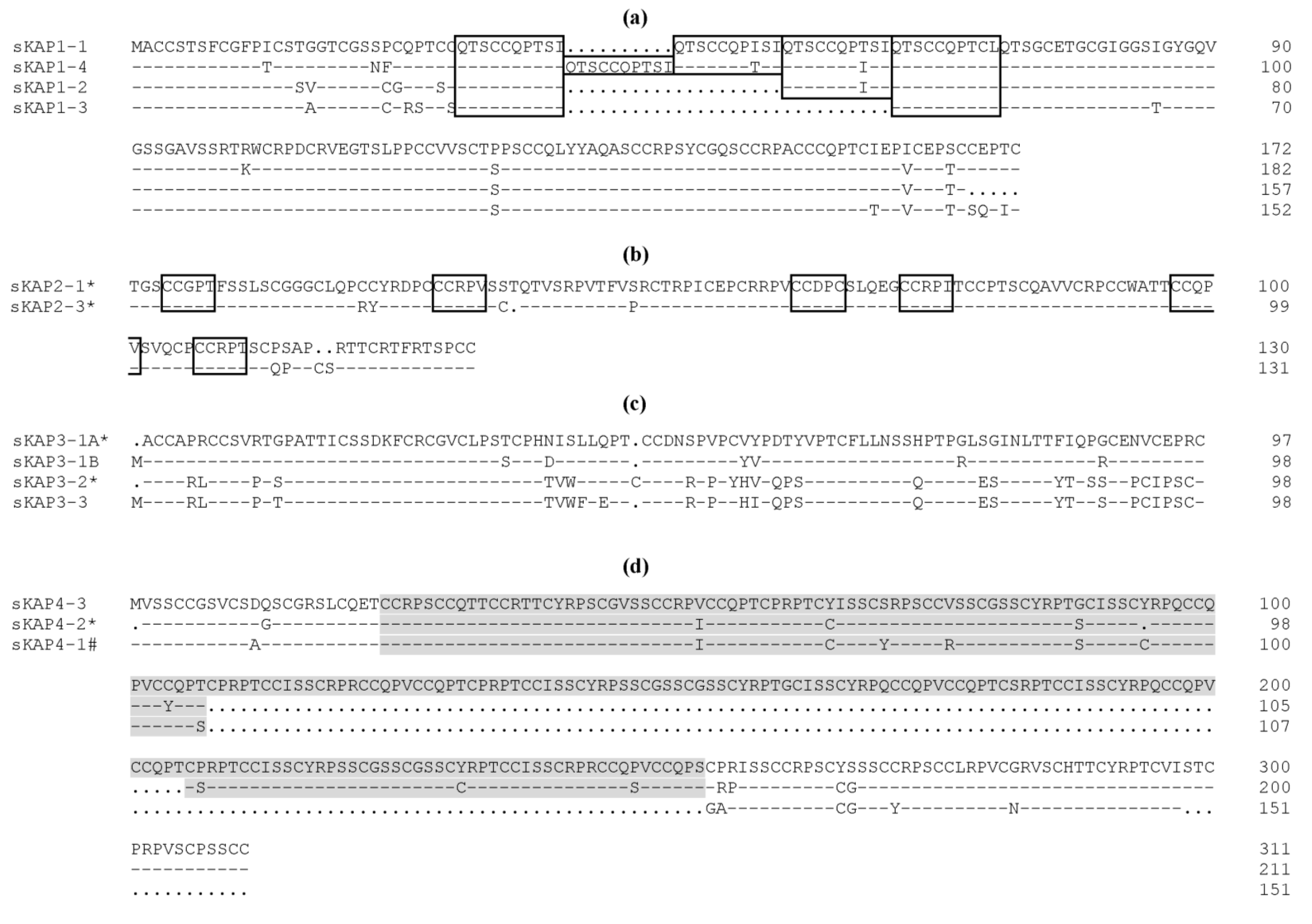
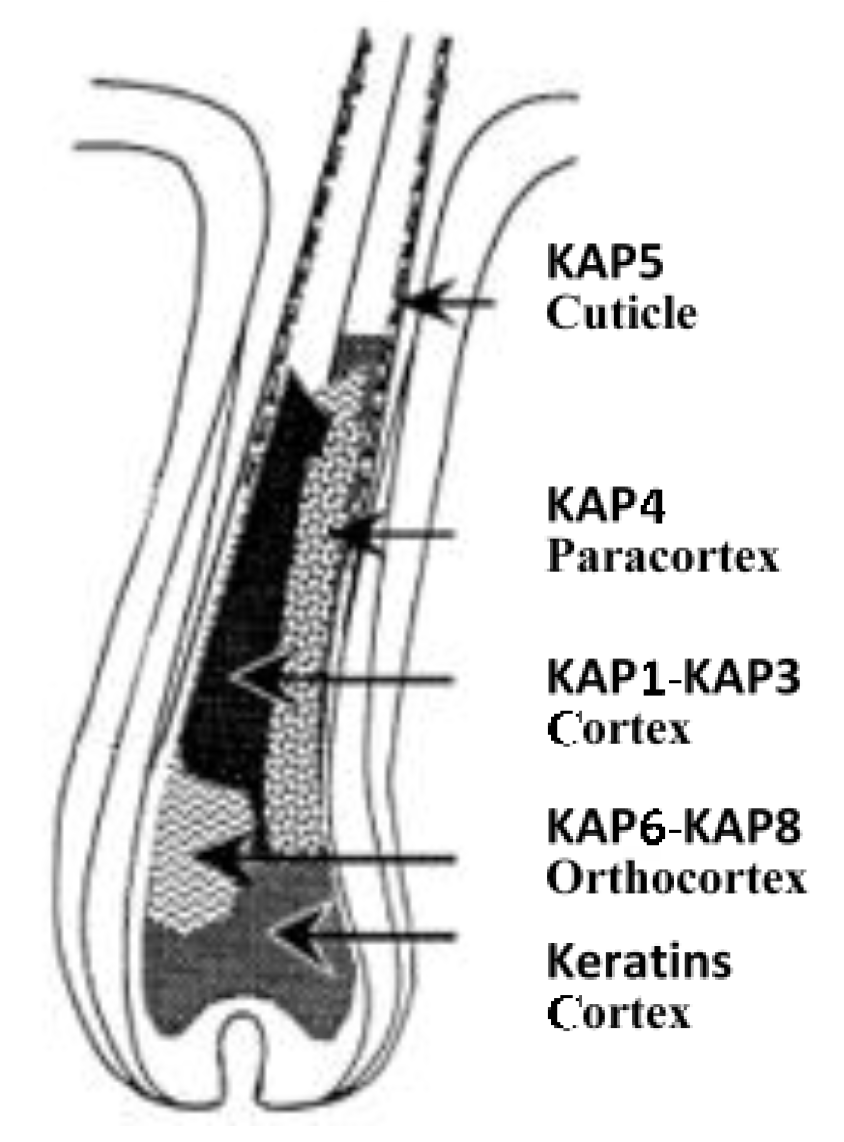
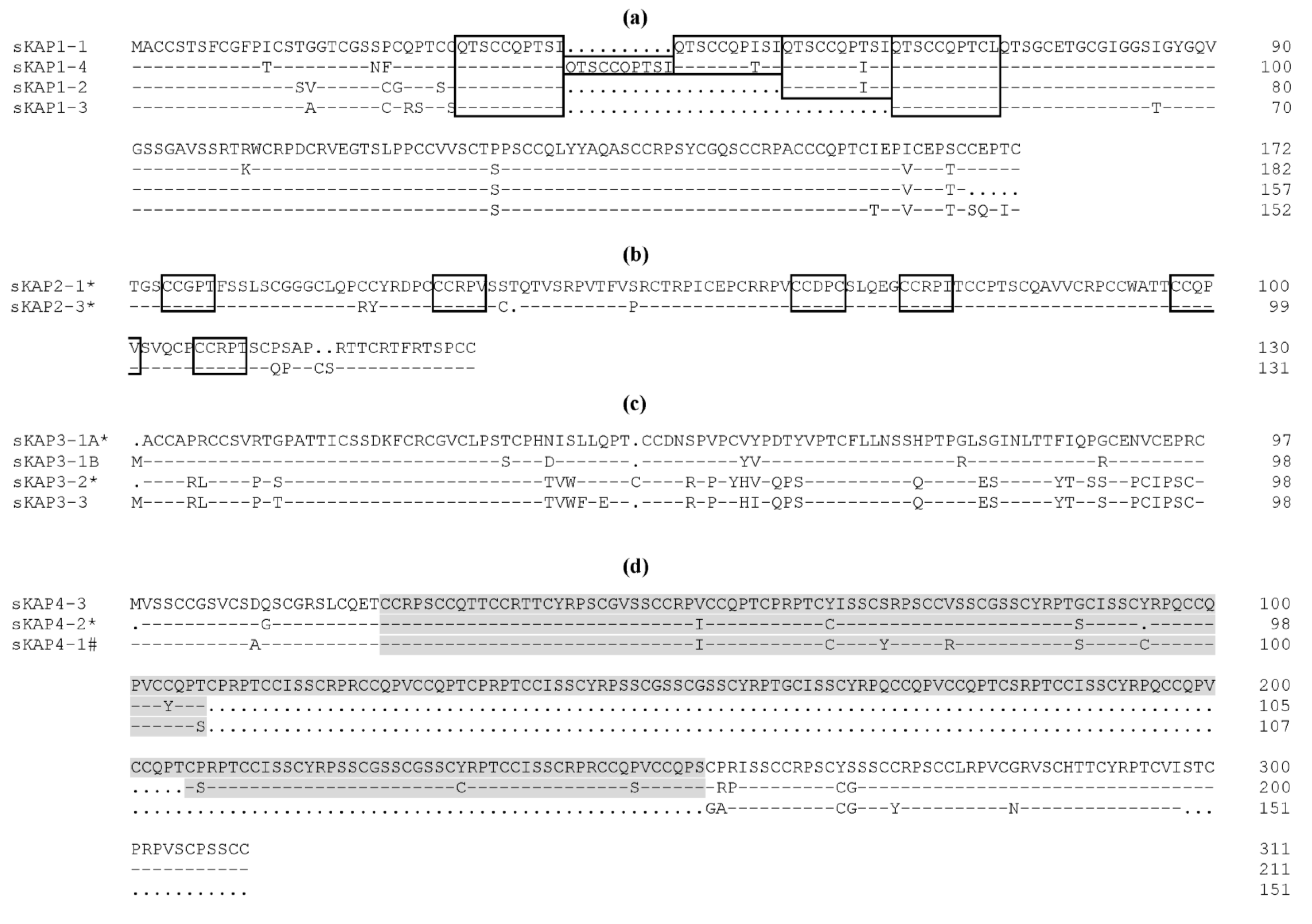
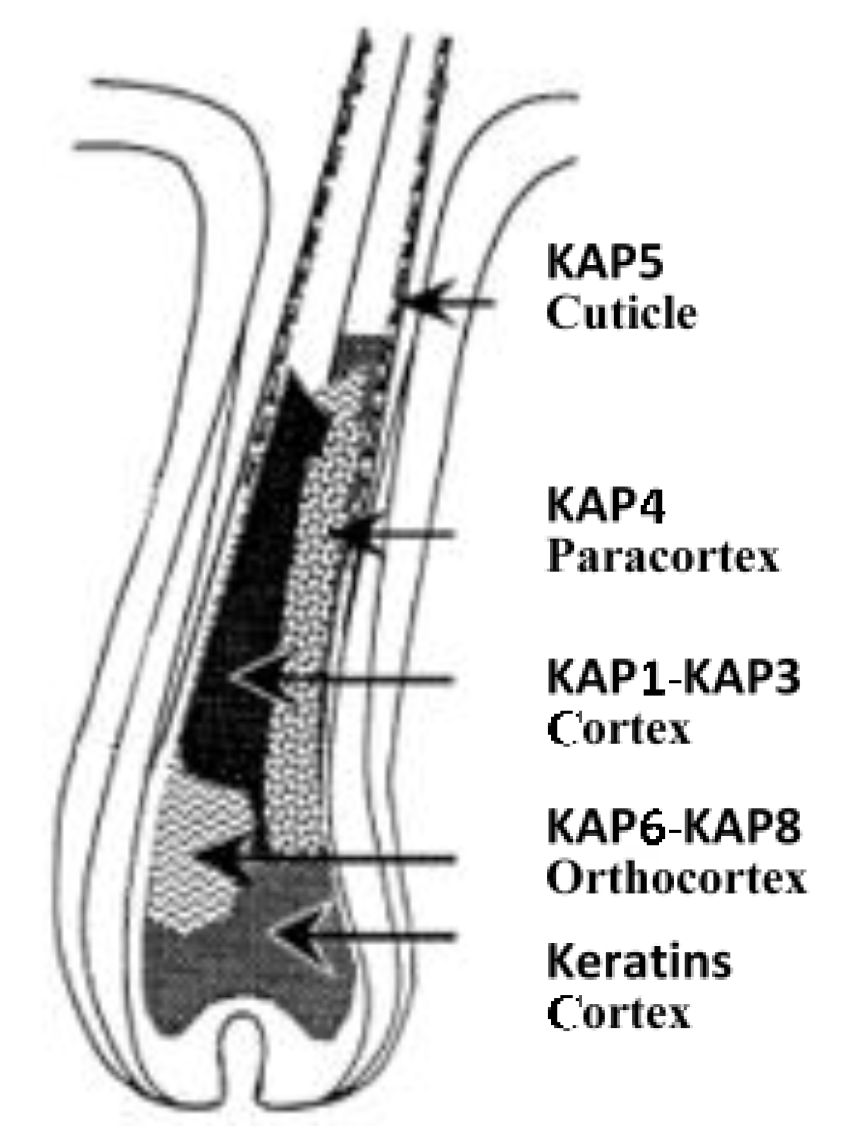
2. The Complexity of Wool KAPs
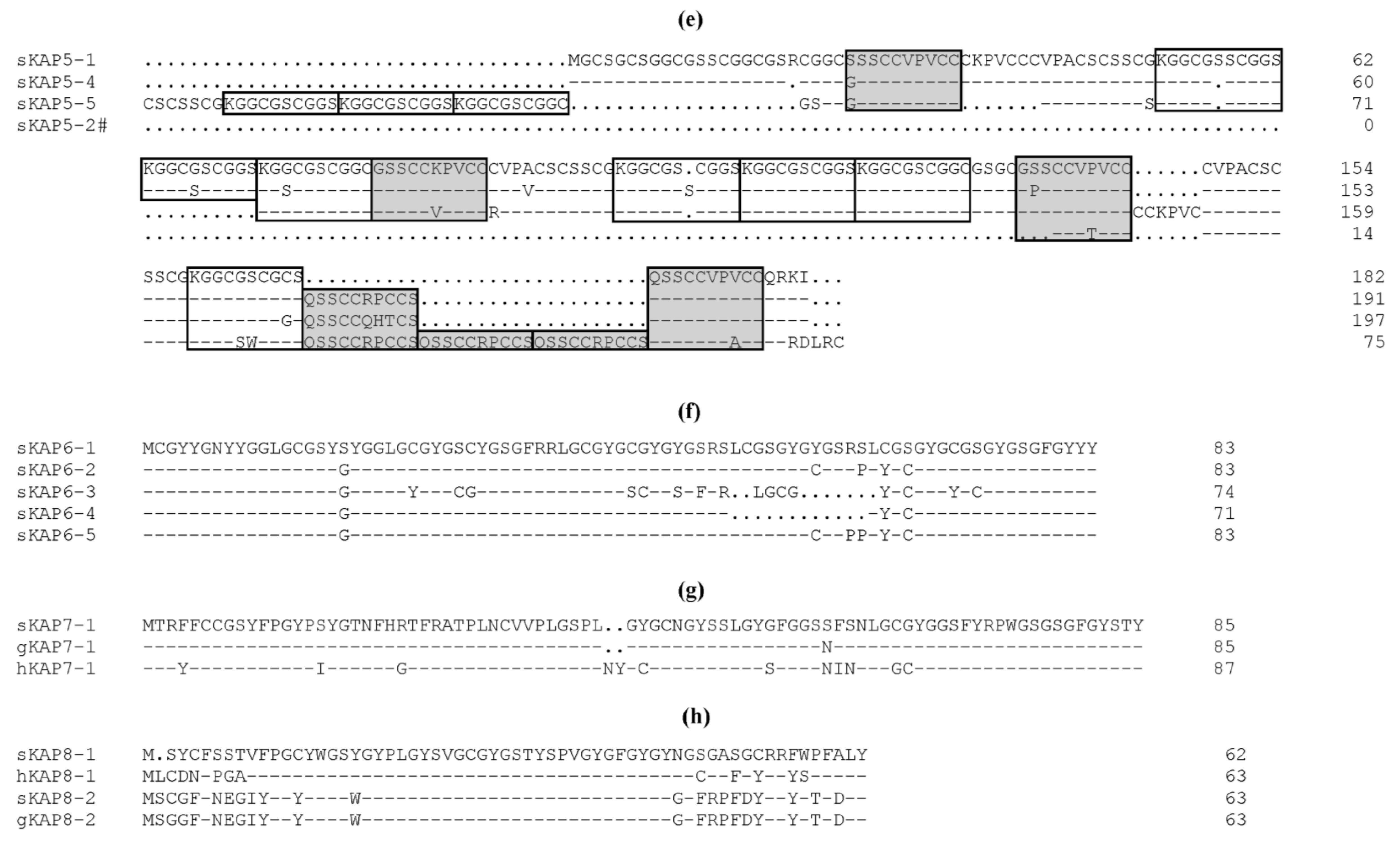
3. The Categorisation and Classification of Wool KAPs
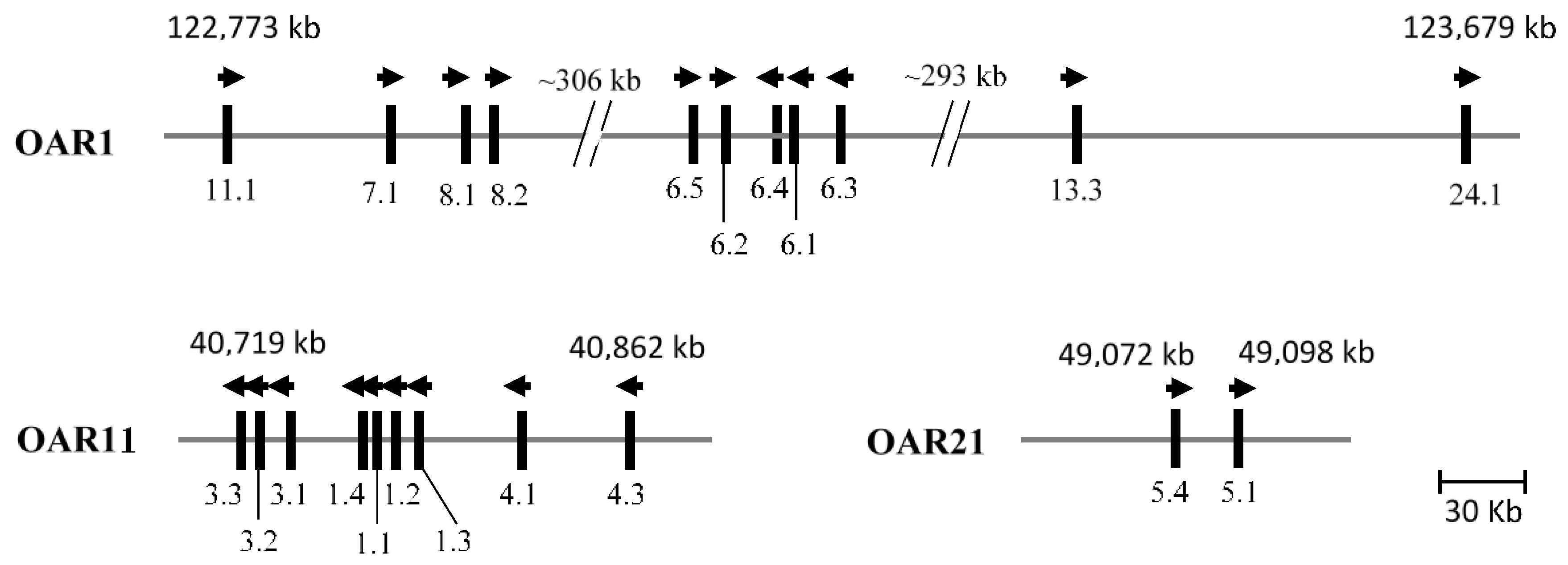
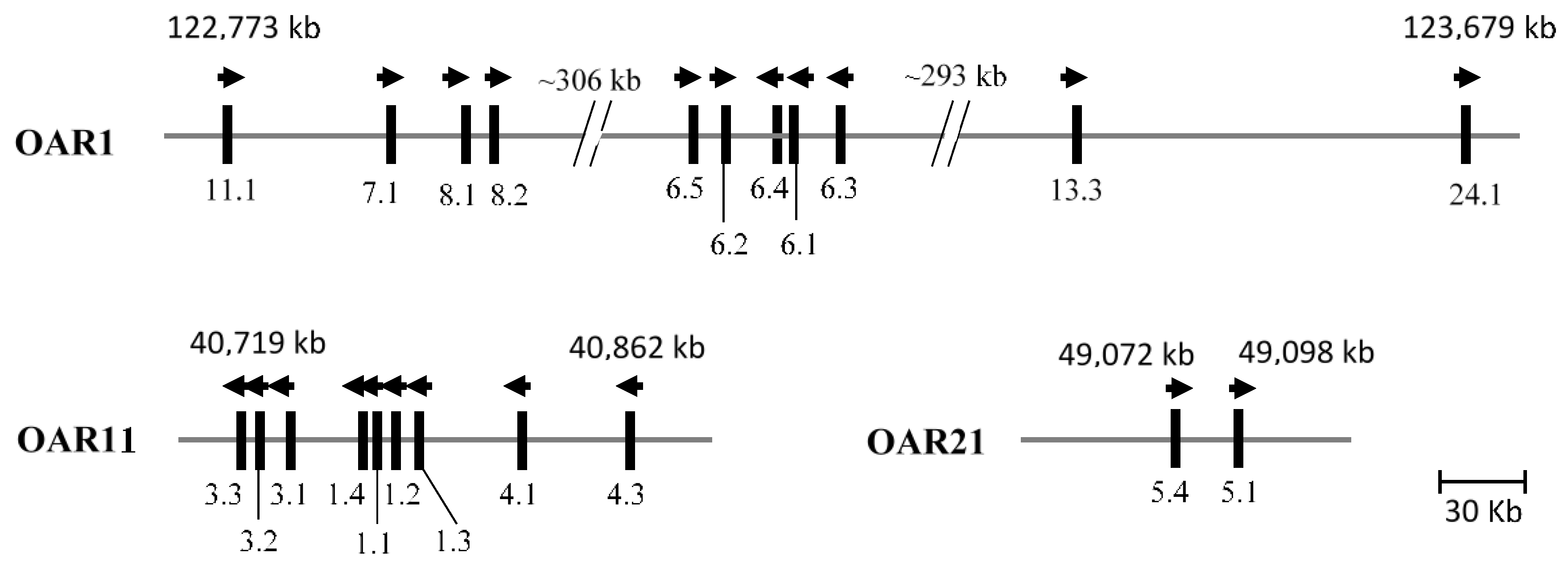
4. The Chromosomal Location of Wool KAP Genes
5. Nucleotide Polymorphism within the KAP Genes
6. The Expression of KAPs
7. KAP Genes and Wool Traits
Acknowledgements
Conflicts of Interest
References
- Leeder, J.D. Wool: Nature’s Wonder Fibre; Australasian Textiles Publishers: Victoria, Australia, 1984. [Google Scholar]
- Popescu, C.; Höcker, H. Hair—The most sophisticated biological composite material. Chem. Soc. Rev. 2007, 36, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Goddard, D.R.; Michaelis, L. Derivatives of keratin. J. Biol. Chem. 1935, 112, 361–371. [Google Scholar]
- Powell, B.C.; Rogers, G.E. Hair keratin: Composition, structure and biogenesis. In Biology of the Integument; Bereiter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin, Germany, 1986; Volume 2, pp. 695–721. [Google Scholar]
- Powell, B.C.; Rogers, G.E. The role of keratin proteins and their genes in the growth, structure and properties of hair. In Formation and Structure of Human Hair; Jollès, P., Zahn, H., Hӧcker, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1997; pp. 59–148. [Google Scholar]
- Rogers, G.E. Hair follicle differentiation and regulation. Int. J. Dev. Biol. 2004, 48, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Plowman, J.E. Proteomic database of wool components. J. Chromatogr. 2003, 787, 63–76. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Winter, H.; Schweizer, J. Human hair keratin-associated proteins (KAPs). Int. Rev. Cytol. 2006, 251, 209–263. [Google Scholar] [PubMed]
- Haylett, T.; Swart, L. Studies on the high-sulfur proteins of reduced Merino wool. Part III: The amino-acid sequence of protein SCMKB-IIIB2. Text. Res. J. 1969, 39, 917–929. [Google Scholar]
- Elleman, T.C. Amino-acid sequence of a high-sulphur protein from wool. Nature 1971, 234, 148–148. [Google Scholar] [CrossRef]
- Haylett, T.; Swart, L.; Parris, D. Studies on the high-sulphur proteins of reduced Merino wool. Amino acid sequence of protein SCMBK-IIIB3. Biochem. J. 1971, 123, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Swart, L.; Haylett, T. Studies on the high-sulphur proteins of reduced Merino wool. Amino acid sequence of protein SCMKB-IIIB4. Biochem. J. 1971, 123, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Elleman, T.C. The amino acid sequence of protein SCMK-B2A from the high-sulphur fraction of wool keratin. Biochem. J. 1972, 130, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Elleman, T.C. The amino acid sequence of protein SCMK-B2C from the high-sulphur fraction of wool keratin. Biochem. J. 1972, 128, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Elleman, T.C.; Dopheide, T.A. The sequence of SCMK-B2B, a high-sulfur protein from wool keratin. J. Biol. Chem. 1972, 247, 3900–3909. [Google Scholar] [PubMed]
- Swart, L.S.; Haylett, T. Studies on the high-sulphur proteins of reduced Merino wool. Amino acid sequence of protein SCMKB-IIIA3. Biochem. J. 1973, 133, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Dopheide, T.A. The primary structure of a protein, component 0.62, rich in glycine and aromatic residues, obtained from wool keratin. Eur. J. Biochem. 1973, 34, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.M. The proteins of hari and other hard α-keartins. In Cellular and Molecular Biology of Intermediate Filaments; Goldman, R.D., Steinert, P.M., Eds.; Plenum: New York, NY, USA, 1990; pp. 95–128. [Google Scholar]
- Powell, B.C.; Sleigh, M.J.; Ward, K.A.; Rogers, G.E. Mammalian keratin gene families: Organisation of genes coding for the B2 high-sulphur proteins of sheep wool. Nucleic Acids Res. 1983, 11, 5327–5346. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, M.J.; Powell, B.C.; Ward, K.A.; Sleigh, M.J.; Rogers, G.E. The keratin BIIIB gene family: Isolation of cDNA clones and structure of a gene and a related pseudogene. Genomics 1989, 4, 182–191. [Google Scholar] [CrossRef]
- MacKinnon, P.; Powell, B.; Rogers, G. Structure and expression of genes for a class of cysteine-rich proteins of the cuticle layers of differentiating wool and hair follicles. J. Cell. Biol. 1990, 111, 2587–2600. [Google Scholar] [CrossRef] [PubMed]
- Fratini, A.; Powell, B.C.; Hynd, P.I.; Keough, R.A.; Rogers, G.E. Dietary cysteine regulates the levels of messenger-rnas encoding a family of cysteine-rich proteins of wool. J. Investig. Dermatol. 1994, 102, 178–185. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jenkins, B.J.; Powell, B.C. Differential expression of genes encoding a cysteine-rich keratin family in the hair cuticle. J. Investig. Dermatol. 1994, 103, 310–317. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kuczek, E.; Rogers, G.E. Sheep keratins: Characterization of cDNA clones for the glycine + tyrosine-rich wool proteins using a synthetic probe. Eur. J. Biochem. 1985, 146, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Kuczek, E.S.; Rogers, G.E. Sheep wool (glycine + tyrosine)-rich keratin genes. Eur. J. Biochem. 1987, 166, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Fratini, A.; Powell, B.C.; Rogers, G.E. Sequence, expression, and evolutionary conservation of a gene encoding a glycine tyrosine-rich keratin-associated protein of hair. J. Biol. Chem. 1993, 268, 4511–4518. [Google Scholar] [PubMed]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G. Identification of the ovine KAP11-1 gene (KRTAP11-1) and genetic variation in its coding sequence. Mol. Biol. Rep. 2011, 38, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Dyer, J.M.; Plowman, J.E.; Hickford, J.G.H. Identification of the keratin-associated protein 13-3 (KAP13-3) gene in sheep. Open J. Genet. 2011, 1, 60–64. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Yu, Z.; Dyer, J.; Plowman, J.E.; Hickford, J.G.H. Identification of the ovine keratin-associated protein KAP1-2 gene (KRTAP1-2). Exp. Dermatol. 2011, 20, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gong, H.; Yan, W.; Luo, Y.; Hickford, J.G.H. Identification and sequence analysis of the keratin-associated protein 24-1 (KAP24-1) gene homologue in sheep. Gene 2012, 511, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Gordon, S.W.; Nixon, A.J.; Bawden, C.S.; Rogers, M.A.; Wildermoth, J.E.; Maqbool, N.J.; Pearson, A.J. Expression patterns of keratin intermediate filament and keratin associated protein genes in wool follicles. Differentiation 2009, 77, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hickford, J.G.H. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G.H. The sheep KAP8-2 gene, a new KAP8 family member that is absent in humans. SpringerPlus 2014, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gong, H.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G.H. Identification of four new gene members of the KAP6 gene family in sheep. Sci. Rep. 2016, 6, 24074. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Aoki, N.; Schweizer, J.; Langbein, L.; Rogers, M.A.; Winter, H.; Ito, M. Polymorphisms in the human high sulfur hair keratin-associated protein 1, KAP1, gene family. J. Biol. Chem. 2002, 277, 45493–45501. [Google Scholar] [CrossRef] [PubMed]
- Kariya, N.; Shimomura, Y.; Ito, M. Size polymorphisms in the human ultrahigh sulfur hair keratin-associated protein 4, KAP4, gene family. J. Investig. Dermatol. 2005, 124, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Schweizer, J. Human KAP genes, only the half of it? Extensive size polymorphisms in hair keratin-associated protein genes. J. Investig. Dermatol. 2005, 124, vii–ix. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Winter, H.; Langbein, L.; Wollschläger, A.; Praetzel-Wunder, S.; Jave-Suarez, L.F.; Schweizer, J. Characterization of human KAP24. 1, a cuticular hair keratin-associated protein with unusual amino-acid composition and repeat structure. J. Investig. Dermatol. 2007, 127, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Praetzel Wunder, S.; Giehl, K. Characterization and expression analysis of the hair keratin associated protein KAP26.1. Br. J. Dermatol. 2008, 159, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Winter, H.; Ehmann, C.; Praetzel, S.; Korn, B.; Schweizer, J. Characterization of a cluster of human high/ultrahigh sulfur keratin-associated protein genes embedded in the type I keratin gene domain on chromosome 17q12-21. J. Biol. Chem. 2001, 276, 19440–19451. [Google Scholar] [CrossRef] [PubMed]
- Rogers, G.R.; Hickford, J.G.H.; Bickerstaffe, R. Polymorphism in two genes for B2 high sulfur proteins of wool. Anim. Genet. 1994, 25, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Itenge-Mweza, T.O.; Forrest, R.H.; McKenzie, G.W.; Hogan, A.; Abbott, J.; Amoafo, O.; Hickford, J.G.H. Polymorphism of the KAP1.1, KAP1.3 and K33 genes in Merino sheep. Mol. Cell. Probs 2007, 21, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hickford, J.G. Polymorphism of the ovine keratin-associated protein 1–4 gene (KRTAP1-4). Mol. Biol. Rep. 2010, 37, 3377–3380. [Google Scholar] [CrossRef] [PubMed]
- Swart, L.; Haylett, T.; Joubert, F. Studies on the high-sulfur proteins of reduced Merino wool. Part II1: The isolation of a homogeneous protein. Text. Res. J. 1969, 39, 912–917. [Google Scholar]
- Rogers, M.A.; Langbein, L.; Winter, H.; Ehmann, C.; Praetzel, S.; Schweizer, J. Characterization of a first domain of human high glycine-tyrosine and high sulfur keratin-associated protein (KAP) genes on chromosome 21q22.1. J. Biol. Chem. 2002, 277, 48993–49002. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Winter, H.; Langbein, L.; Bleiler, R.; Schweizer, J. The human type I keratin gene family: Characterization of new hair follicle specific members and evaluation of the chromosome 17q21.2 gene domain. Differentiation 2004, 72, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.E.; Reeves, R.H. A cluster of keratin-associated proteins on mouse chromosome 10 in the region of conserved linkage with human chromosome 21. Genomics 1998, 54, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, F.; Lassing, C.; Range, A.; Mueller, M.; Hunziker, T.; Ziemiecki, A.; Andres, A.C. Pmg-1 and pmg-2 constitute a novel family of KAP genes differentially expressed during skin and mammary gland development. Mech. Dev. 1999, 86, 193–196. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Winter, H.; Beckmann, I.; Praetzel, S.; Schweizer, J. Hair keratin associated proteins: Characterization of a second high sulfur KAP gene domain on human chromosome 21. J. Investig. Dermatol. 2004, 122, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Obayashi, I.; Asakawa, S.; Minoshima, S.; Kudoh, J.; Shimizu, N. A cluster of 21 keratin-associated protein genes within introns of another gene on human chromosome 21q22.3. Genomics 2004, 83, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Yahagi, S.; Shibuya, K.; Obayashi, I.; Masaki, H.; Kurata, Y.; Kudoh, J.; Shimizu, N. Identification of two novel clusters of ultrahigh-sulfur keratin-associated protein genes on human chromosome 11. Biochem. Biophys. Res. Commun. 2004, 318, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.D.; Irwin, D.; Zhang, Y.P. Molecular evolution of the keratin associated protein gene family in mammals, role in the evolution of mammalian hair. BMC Evol. Biol. 2008, 8, 241. [Google Scholar] [CrossRef] [PubMed]
- McLaren, R.J.; Rogers, G.R.; Davies, K.P.; Maddox, J.F.; Montgomery, G.W. Linkage mapping of wool keratin and keratin-associated protein genes in sheep. Mamm. Genome 1997, 8, 938–940. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; McKenzie, G.W.; Yu, Z.; Clerens, S.; Dyer, J.M.; Plowman, J.E.; Wright, M.W.; Arora, R.; Bawden, C.S. An Updated Nomenclature for Keratin-Associated Proteins (KAPs). Int. J. Biol. Sci. 2012, 8, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hodge, S.; Dyer, J.M.; Hickford, J.G.H. Association of wool traits with variation in the ovine KAP1–2 gene in Merino cross lambs. Small Rum. Res. 2015, 124, 24–29. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Plowman, J.E.; Dyer, J.M.; Hickford, J.G.H. Analysis of variation in the ovine ultra-high sulphur keratin-associated protein KAP5-4 gene using PCR-SSCP technique. Electrophoresis 2010, 31, 3545–3547. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gong, H.; Li, S.; Luo, Y.; Hickford, J.G.H. A 57-bp deletion in the ovine KAP6-1 gene affects wool fibre diameter. J. Anim. Breed. Genet. 2015, 132, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Plowman, J.E.; Dyer, J.M.; Hickford, J.G.H. Search for variation in the ovine KAP7-1 and KAP8-1 genes using polymerase chain reaction–single-stranded conformational polymorphism screening. DNA Cell. Biol. 2011, 31, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Kunhareang, S.; Gong, H.; Fang, Q.; Hu, J.; Luo, Y.; Hickford, J.G.H. Detection of sequence variation and genotyping of polymorphic genes using polymerase chain reaction stem—Loop conformational polymorphism analysis. Anal. Biochem. 2011, 408, 340–341. [Google Scholar] [CrossRef] [PubMed]
- Miklos, G.L.G.; Maleszka, R. Protein functions and biological contexts. Proteomics 2001, 1, 169–178. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, J.M.; Darskus, R.L. Relation between the tyrosine content of various wools and their content of a class of proteins rich in tyrosine and glycine. Aust. J. Biol. Sci. 1971, 24, 1189–1198. [Google Scholar] [PubMed]
- Rogers, G.; Kuczek, E.; MacKinnon, P.; Presland, R.; Fietz, M. Special biochemical features of the hair follicle. In The Biology of Wool and Hair; Rogers, G.E., Reis, P.J., Ward, K.A., Marshall, R.C., Eds.; Chapman & Hall: London, UK, 1989; pp. 69–85. [Google Scholar]
- Short, B. A dominant felting lustre mutant fleece-type in the Australian Merino sheep. Nature 1958, 181, 1414–1415. [Google Scholar] [CrossRef] [PubMed]
- Blair, H. Inheritance of a major gene for excessively lustrous wool in sheep. J. Hered. 1990, 81, 222–224. [Google Scholar]
- Li, S.W.; Ouyang, H.S.; Rogers, G.E.; Bawden, C.S. Characterization of the structural and molecular defects in fibres and follicles of the merino felting lustre mutant. Exp. Dermatol. 2009, 18, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, H.; Yang, H.; Wang, S.; Rong, E.; Pei, W.; Li, H.; Wang, N. Genome-wide association study for wool production traits in a Chinese Merino sheep population. PLoS ONE 2014, 9, e107101. [Google Scholar] [CrossRef] [PubMed]
- Parsons, Y.M.; Cooper, D.W.; Piper, L.R. Evidence of linkage between high-glycine-tyrosine keratin gene loci and wool fiber diameter in a merino half-sib family. Anim. Genet. 1994, 25, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Parsons, Y.M.; Piper, L.R.; Cooper, D.W. Linkage relationships between keratin-associated protein (KRTAP) genes and growth hormone in sheep. Genomics 1994, 20, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Beh, K.J.; Callaghan, M.J.; Leish, Z.; Hulme, D.J.; Lenane, I.; Maddox, J.F. A genome scan for QTL affecting fleece and wool traits in Merino sheep. Wool Technol. Sheep Breed. 2001, 49, 88–97. [Google Scholar]
- Roldan, D.; Dodero, A.; Bidinost, F.; Taddeo, H.; Allain, D.; Poli, M.; Elsen, J. Merino sheep: A further look at quantitative trait loci for wool production. Animal 2010, 4, 1330–1340. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Allain, D.; Schibler, L.; Mura, L.; Barillet, F.; Sechi, T.; Rupp, R.; Casu, S.; Cribiu, E.; Carta, A. QTL detection with DNA markers for wool traits in a sheep backcross Sarda × Lacaune resource population. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Minas Gerais, Brazil, 13–18 August 2006; Instituto Prociência: Minas Gerais, Brazil, 2006; pp. 5–7. [Google Scholar]
- Rogers, G.; Hickford, J.; Bickerstaffe, R. A Potential QTL for Wool Strength Located on Ovine Chromosome 11. In Proceedings of the 5th World Congress on Genetics Applied to Livestock Production, Guelph, ON, Canada, 7–12 August 1994; pp. 291–294.
- Itenge-Mweza, T.O. Identification of Genetic Markers Associated with Wool Quality Traits in Merino Sheep. Ph.D. Thesis, Lincoln University, Christchurch, New Zealand, 2007. [Google Scholar]
- Zhang, Y.; He, Y.; Xue, P.; Chen, Y. Sequence variant in the KAP1.1 gene associate with cashmere trait in two cashmere goat breeds. Afr. J. Agr. Res. 2011, 6, 2889–2894. [Google Scholar]
- Raadsma, H.W.; Jonas, E.; Fleet, M.R.; Fullard, K.; Gongora, J.; Cavanagh, C.R.; Tammen, I.; Thomson, P.C. QTL and association analysis for skin and fibre pigmentation in sheep provides evidence of a major causative mutation and epistatic effects. Anim. Genet. 2013, 44, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Phuaa, S.H.; Scobieb, D.R.; O’Connellb, D.; Henrya, H.; Doddsa, K.G.; Brauninga, R.; Clerensb, S. Preliminary linkage studies in sheep of keratin and keratin-associated protein genes with fleece weight, wool fibre diameter and fibre curvature. Proc. N. Z. Soc. Anim. Prod. 2015, 75, 101–105. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KAP Family | Category | KAP Member | Old Name | Acidity/Basicity (Isoelectronic Point) | Sequence Type | Sequence Accession Number | Reference |
|---|---|---|---|---|---|---|---|
| KAP1 | HS | KAP1-1 | B2A | Acidic (5.9) | Protein * | P02438 | [13] |
| DNA | X01610 # | [19] | |||||
| KAP1-2 | B2B | Neutral (7.0) | Protein | P02439 | [15] | ||
| DNA | HQ897973 # | [29] | |||||
| KAP1-3 | B2C | Neutral (7.3) | Protein | 711148A | [14] | ||
| DNA | X02925 # | [19] | |||||
| KAP1-4 | B2D | Acidic (5.9) | DNA | X01610 # | [19] | ||
| KAP2 | HS | KAP2-1 | BIIIA3A, KAP2.12 | Basic (8.1) | Protein | P02443 | [16] |
| KAP2-3 | BIIIA3 | Basic (8.1) | Protein | P02441 | [16] | ||
| DNA * | U60024 | Unpublished | |||||
| KAP3 | HS | KAP3-1A | BIIIB2 | Neutral (7.0) | Protein | P02446 | [9] |
| KAP3-1B | BIIIB2 | Neutral (7.4) | DNA | M21099 | [20] | ||
| KAP3-2 | BIIIB3 | Neutral (7.0) | Protein | P02444 | [11] | ||
| DNA * | M21100 | [20] | |||||
| KAP3-3 | BIIIB4 | Neutral (7.0) | Protein | P02445 | [12] | ||
| DNA | M21103 | [20] | |||||
| KAP4 | UHS | KAP4-1 | ND | DNA * | X73462 | [22] | |
| KAP4-2 | Basic (8.1) | Protein | [5] | ||||
| KAP4-3 | Basic (8.2) | DNA | EU239778 | [31] | |||
| KAP5 | UHS | KAP5-1 | Basic (7.9) | DNA | X55294 | [21] | |
| KAP5-2 | ND | DNA * | [4] | ||||
| KAP5-4 | Basic (7.9) | DNA | X73434 # | [23] | |||
| KAP5-5 | ND | DNA * | X73435 | [23] | |||
| KAP6 | HGT | KAP6-1 | HGT type II | Basic (8.3) | DNA | M95719 # | [22] |
| KAP6-2 | HGT type II | Basic (8.2) | Protein * | [18] | |||
| DNA | KT725827 # | [34] | |||||
| KAP6-3 | Basic (8.4) | DNA | KT725833 # | [34] | |||
| KAP6-4 | Basic (8.2) | DNA | KT725838 # | [34] | |||
| KAP6-5 | Basic (8.1) | DNA | KT725841 # | [34] | |||
| KAP7 | HGT | KAP7-1 | HGT-C2 | Basic (8.7) | DNA | X05638 | [25] |
| KAP8 | HGT | KAP8-1 | HGT-F | Basic (8.3) | Protein | P02448 | [17] |
| DNA | X05639 # | [25] | |||||
| KAP8-2 | Acid (6.3) | DNA | KF220646 # | [33] | |||
| KAP11 | HS | KAP11-1 | Basic (8.0) | DNA | HQ595347 # | [27] | |
| KAP13 | HS | KAP13-3 | Basic (9.4) | DNA | JN377429 # | [28] | |
| KAP24 | HS | KAP24-1 | Basic (8.5) | DNA | JX112014 # | [30] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, H.; Zhou, H.; Forrest, R.H.J.; Li, S.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G.H. Wool Keratin-Associated Protein Genes in Sheep—A Review. Genes 2016, 7, 24. https://doi.org/10.3390/genes7060024
Gong H, Zhou H, Forrest RHJ, Li S, Wang J, Dyer JM, Luo Y, Hickford JGH. Wool Keratin-Associated Protein Genes in Sheep—A Review. Genes. 2016; 7(6):24. https://doi.org/10.3390/genes7060024
Chicago/Turabian StyleGong, Hua, Huitong Zhou, Rachel H. J. Forrest, Shaobin Li, Jiqing Wang, Jolon M. Dyer, Yuzhu Luo, and Jon G. H. Hickford. 2016. "Wool Keratin-Associated Protein Genes in Sheep—A Review" Genes 7, no. 6: 24. https://doi.org/10.3390/genes7060024
APA StyleGong, H., Zhou, H., Forrest, R. H. J., Li, S., Wang, J., Dyer, J. M., Luo, Y., & Hickford, J. G. H. (2016). Wool Keratin-Associated Protein Genes in Sheep—A Review. Genes, 7(6), 24. https://doi.org/10.3390/genes7060024