
New Oligonucleotide Probes for ND-FISH Analysis to Identify Barley Chromosomes and to Investigate Polymorphisms of Wheat Chromosomes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Oligonucleotide Probe Development
2.3. ND-FISH Analysis
2.4. Cloning of Original Repetitive DNA Sequences
2.5. Denaturing FISH
3. Results
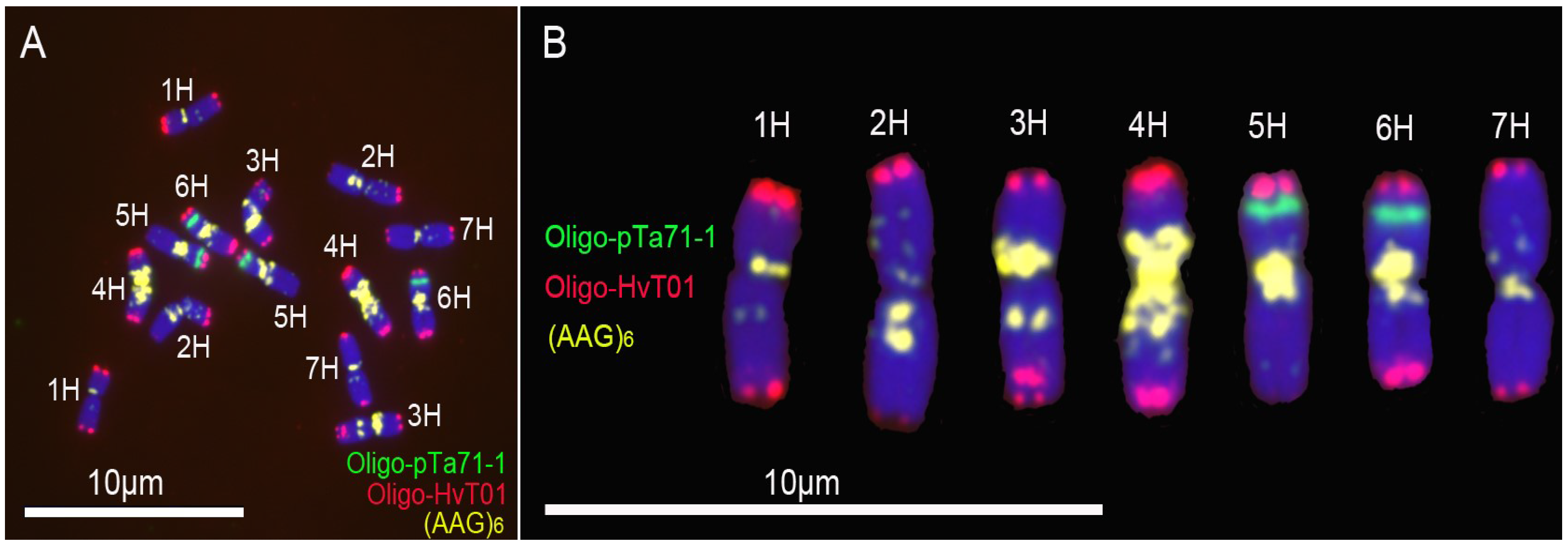
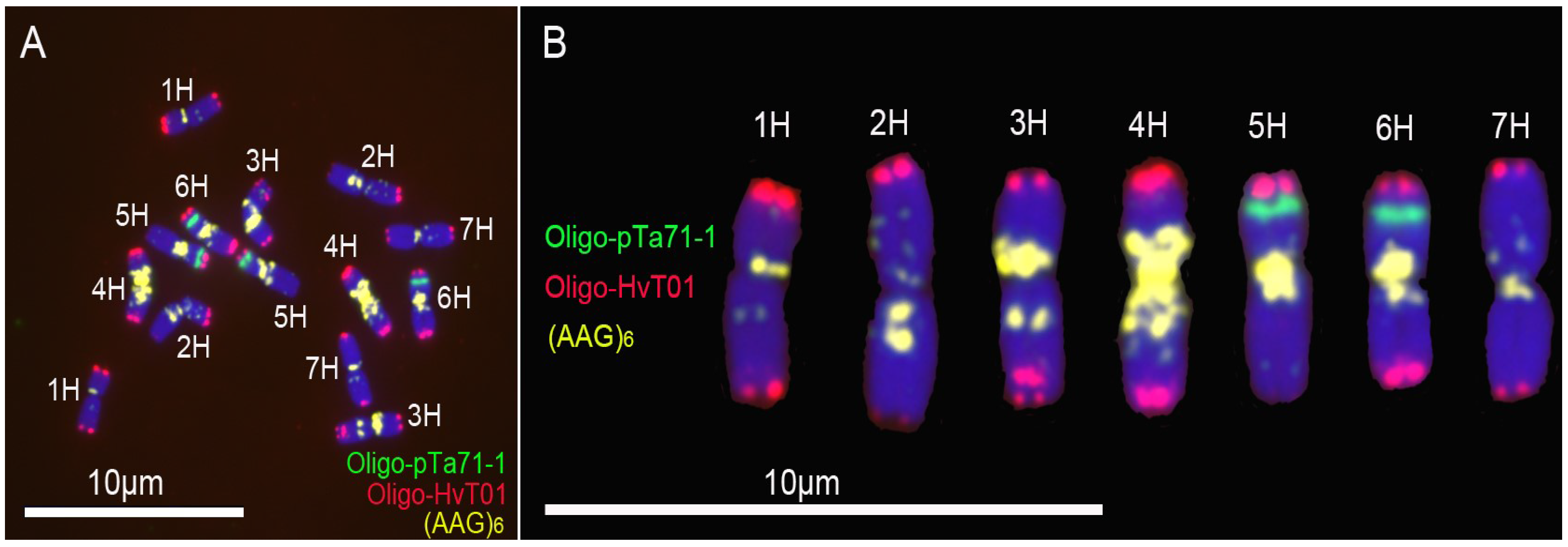
3.1. New Oligonucleotide Probes for Identifying Barley Chromosomes
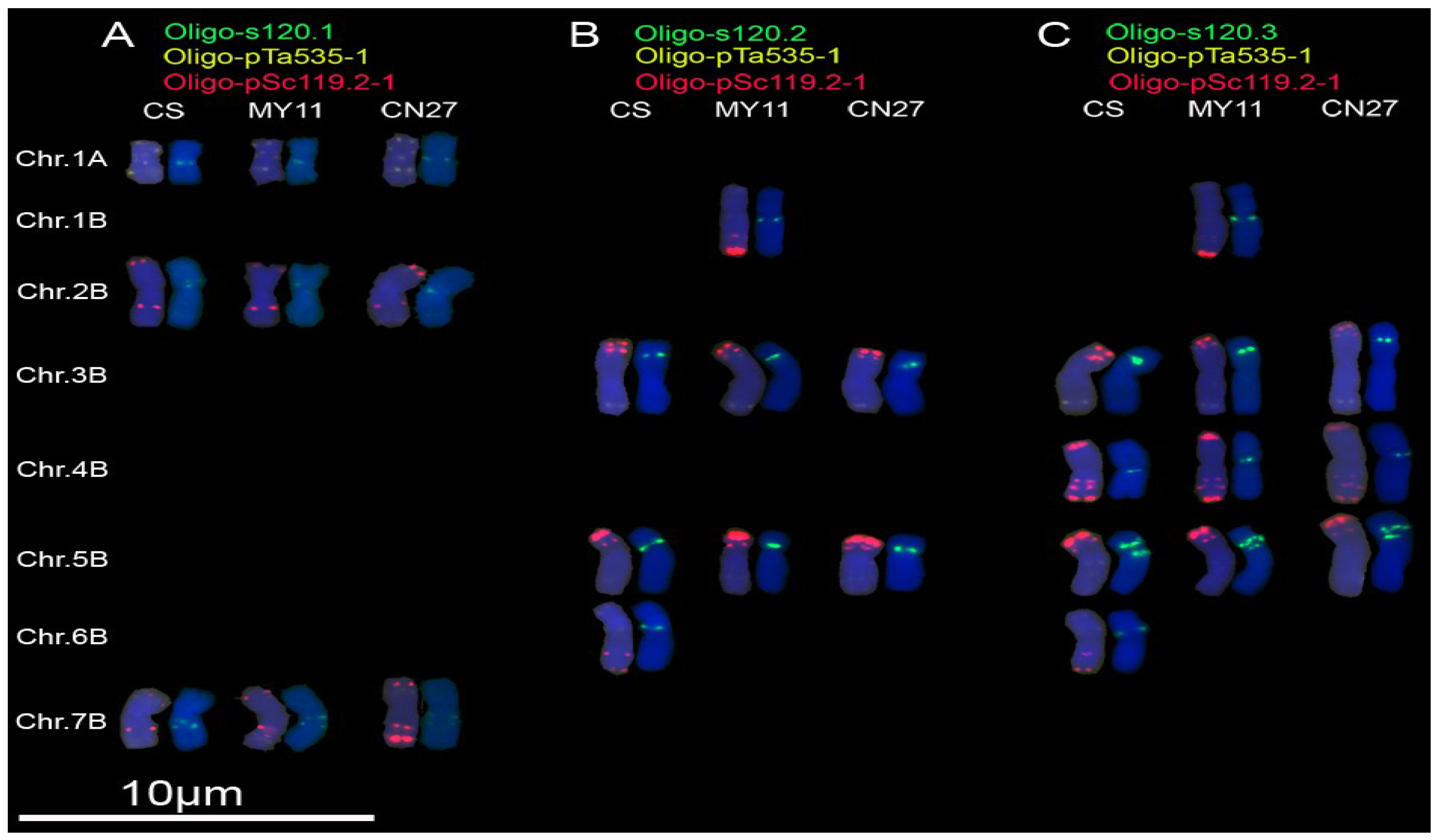
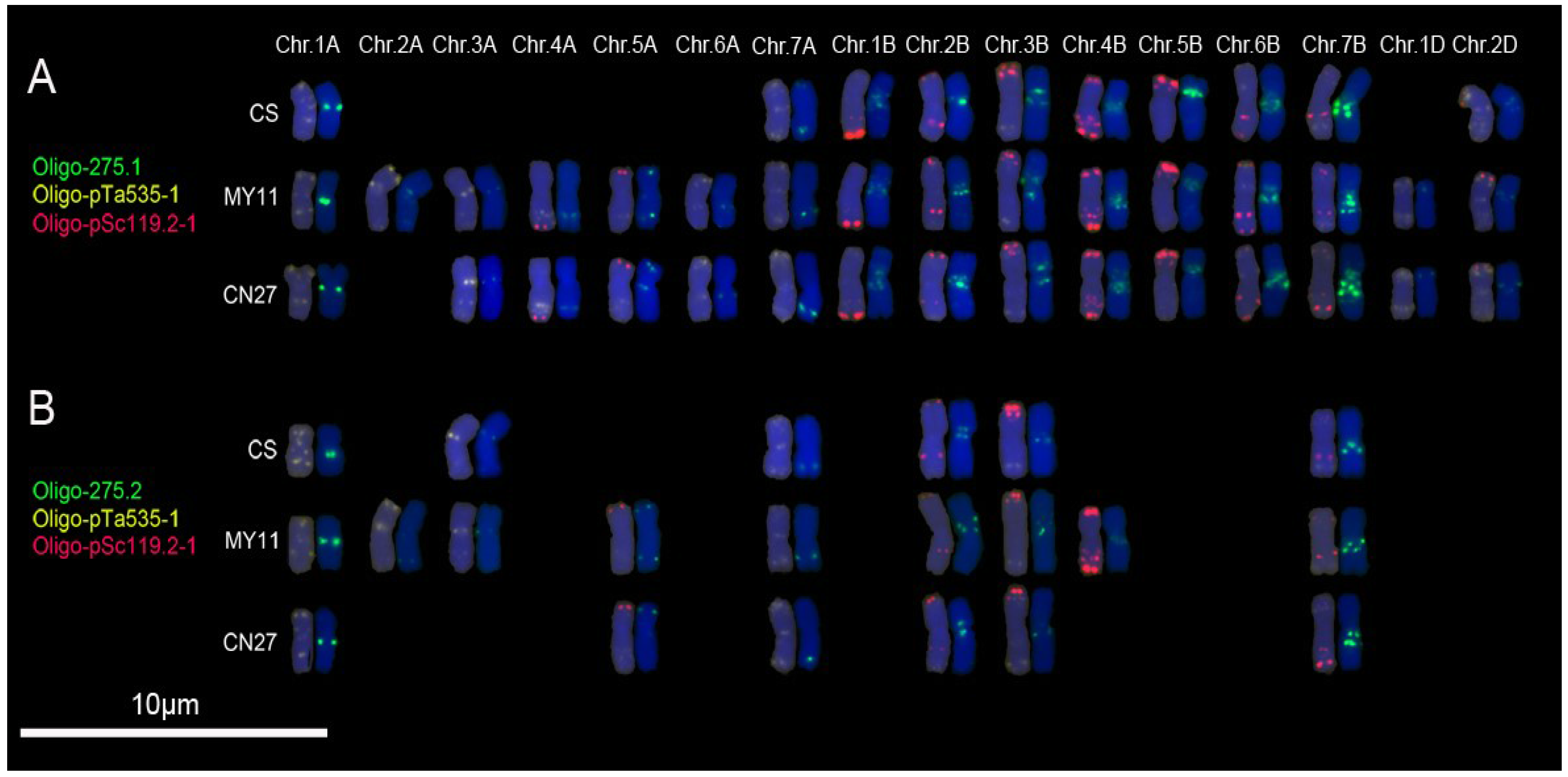
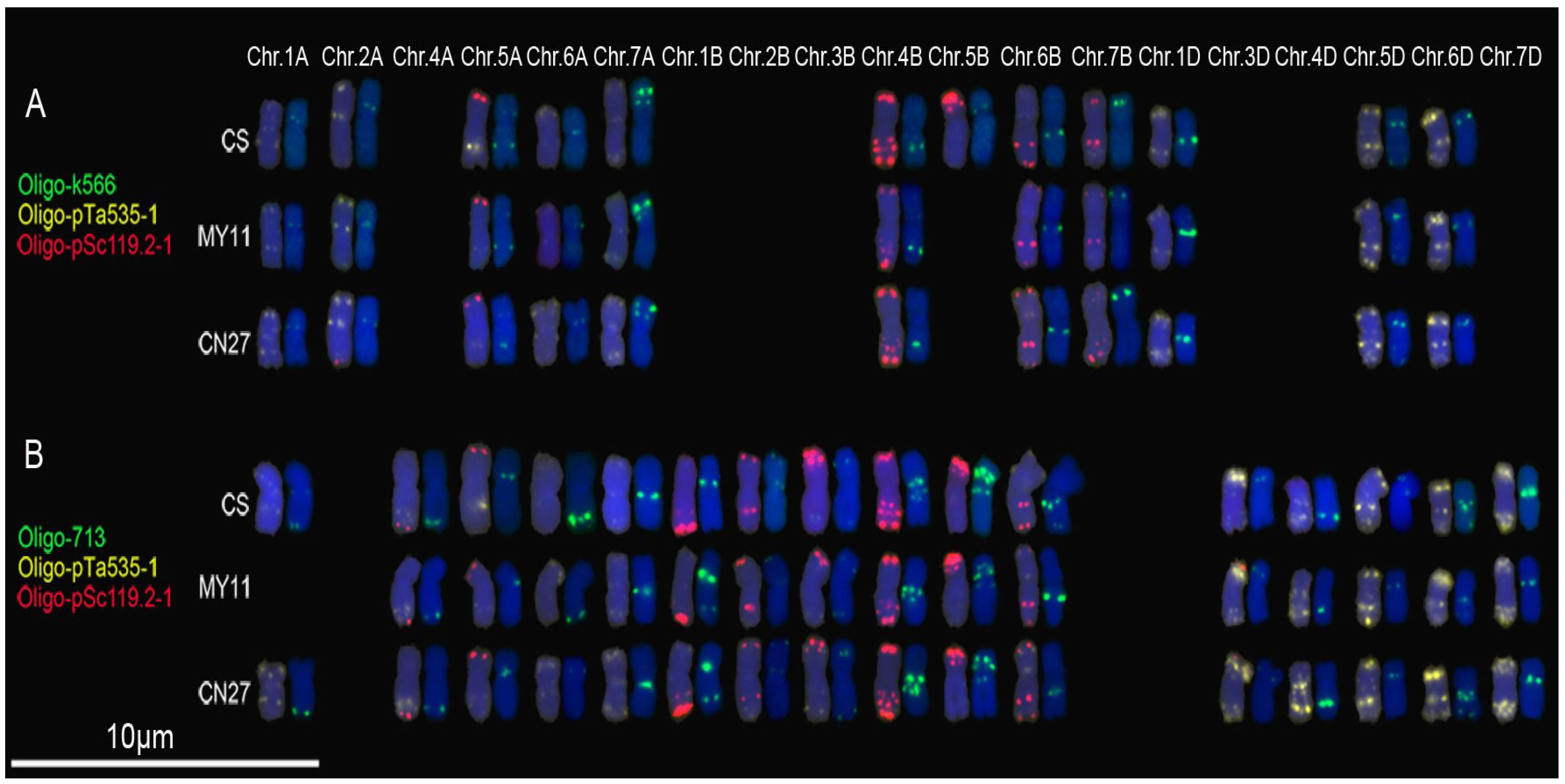
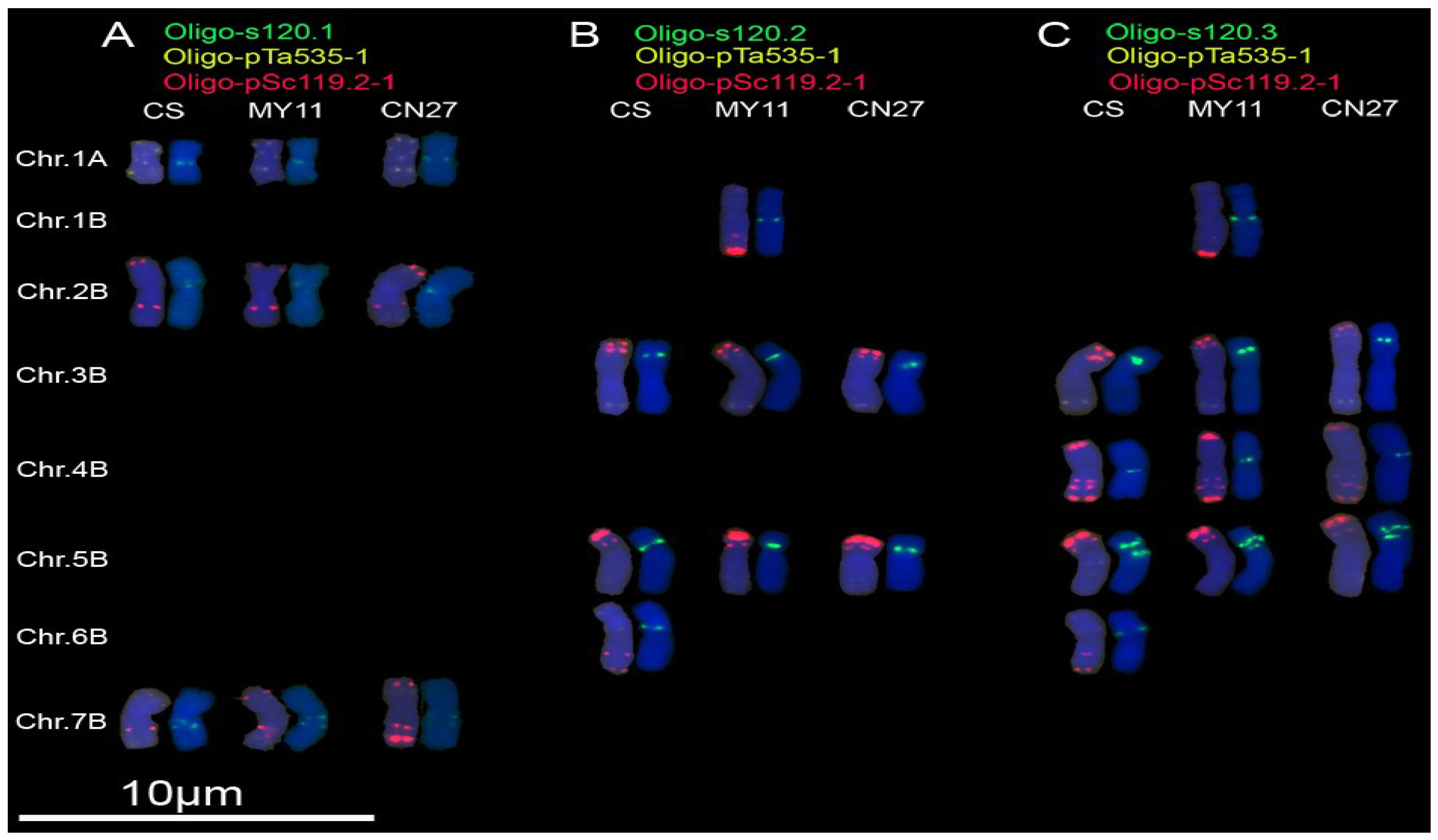
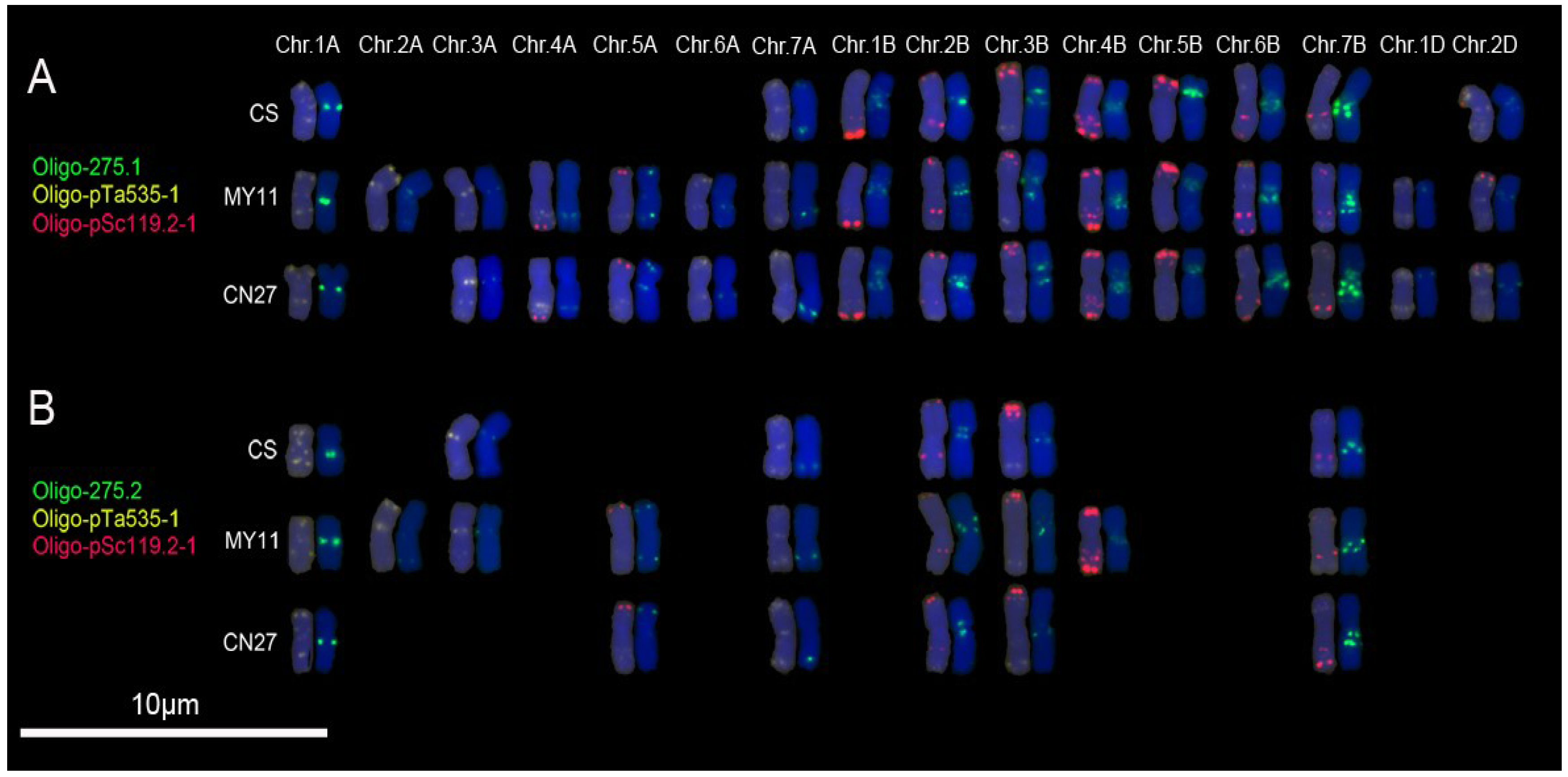
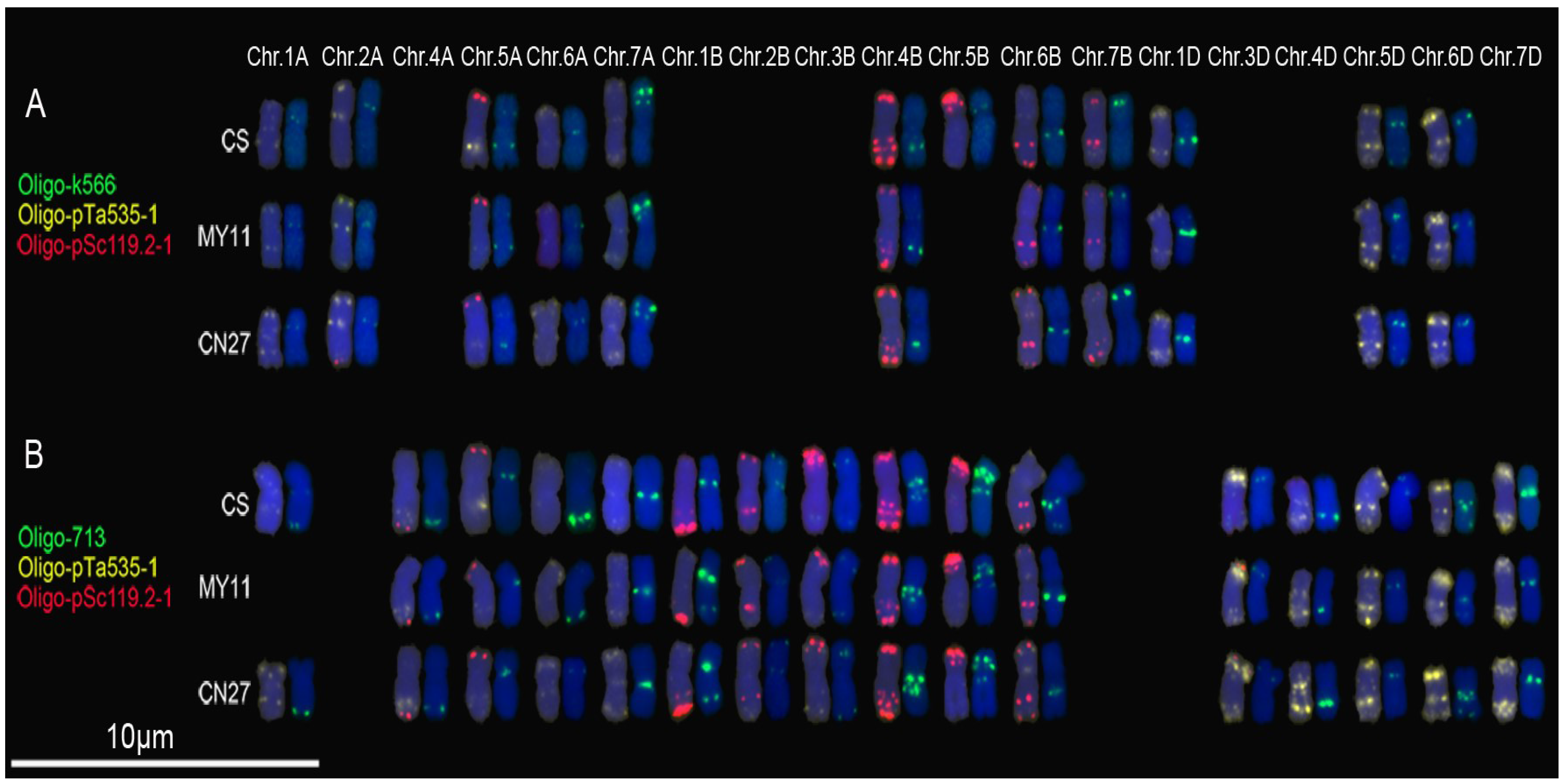
3.2. New Oligonucleotide Probes for Identifying Wheat Chromosomes
3.3. Cloning of Original Repetitive Sequences
3.4. Hybridization Patterns of Oligonucleotide Probes and Their Corresponding Repetitive Sequence Families
4. Discussion
4.1. Convenience of Oligonucleotide Probes in Analyzing Barley and Wheat Chromosomes
4.2. Oligonucleotide Probes Displaying Polymorphisms of Wheat Chromosomes
4.3. Oligonucleotide Probes Implying Different Structural Status of Tandem Repeats
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jiang, J.M.; Gill, B.S. Current status and the future of fluorescence in situ hybridization (FISH) in plant genome research. Genome 2006, 49, 1057–1068. [Google Scholar] [PubMed]
- Pedersen, C.; Langridge, P. Identification of the entire chromosome complement of bread wheat by two-colour FISH. Genome 1997, 40, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Linc, G.; Molnár-Láng, M. Fluorescence in situ hybridization polymorphism using two repetitive DNA clones in different cultivars of wheat. Plant Breed. 2003, 122, 396–400. [Google Scholar] [CrossRef]
- Danilova, T.V.; Friebe, B.; Gill, B.S. Single-copy gene fluorescence in situ hybridization and genome analysis: Acc-2 loci mark evolutionary chromosomal rearrangements in wheat. Chromosoma 2012, 121, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Ruban, A.S.; Zoshchuk, S.A.; Surzhikov, S.A.; Knüpffer, H.; Kilian, B. Molecular cytogenetic characterization of Triticum timopheevii chromosomes provides new insight on genome evolution of T. zhukovskyi. Plant Syst. Evol. 2016, 302, 943–956. [Google Scholar] [CrossRef]
- Patokar, C.; Sepsi, A.; Schwarzacher, T.; Kishii, M.; Heslop-Harrison, J.S. Molecular cytogenetic characterization of novel wheat-Thinopyrum bessarabicum recombinant lines carrying intercalary translocations. Chromosoma 2016, 125, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome 2013, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.X.; Yang, Z.J.; Fu, S.L. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.L.; Chen, L.; Wang, Y.Y.; Li, M.; Yang, Z.J.; Qiu, L.; Yan, B.J.; Ren, Z.L.; Tang, Z.X. Oligonucleotide probes for ND-FISH analysis to identify rye and wheat chromosomes. Sci. Rep. 2015, 5, 10552. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.; Carvalho, A.; Martín, A.C.; Martín, A.; Lima-Brito, J. Use of the synthetic Oligo-pTa535 and Oligo-pAs1 probes for identification of Hordeum chilense-origin chromosomes in hexaploid tritordeum. Genet. Resour. Crop. Evol. 2016, 63, 945–951. [Google Scholar] [CrossRef]
- Cuadrado, Á.; Golczyk, H.; Jouve, N. A novel, simple and rapid nondenaturing FISH (ND-FISH) technique for the detection of plant telomeres. Potential used and possible target structures detected. Chromosome Res. 2009, 17, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, Á.; Jouve, N. Chromosomal detection of simple sequence repeats (SSRs) using nondenaturing FISH (ND-FISH). Chromosoma 2010, 119, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, Á.; Carmona, A.; Jouve, N. Chromosomal characterization of the three subgenomes in the polyploids of Hordeum murinum L.: New insight into the evolution of this complex. PLoS ONE 2013, 8, e81385. [Google Scholar] [CrossRef] [PubMed]
- Cabo, S.; Carvalho, A.; Martin, A.; Lima-Brito, J. Structural rearrangements detected in newly-formed hexaploid tritordeum after three sequential FISH experiments with repetitive DNA sequences. J. Genet. 2014, 93, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.; Carvalho, A.; Martín, C.; Lima-Brito, J. Genomic reshuffling in advanced lines of hexaploid tritordeum. Genet. Resour. Crop. Evol. 2016. [Google Scholar] [CrossRef]
- Cuadrado, Á.; Jouve, N. Novel simple sequence repeats (SSRs) detected by ND-FISH in heterochromatin of Drosophila melanogaster. BMC Genom. 2011, 12, 205. [Google Scholar] [CrossRef] [PubMed]
- Pavia, I.; Carvalho, A.; Rocha, L.; Gaspar, M.J.; Lima-Brito, J. Physical location of SSR regions and cytogenetic instabilities in Pinus sylvestris chromosomes revealed by ND-FISH. J. Genet. 2014, 93, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.F.; Harberd, N.P.; Jarvis, M.G.; Flavell, R.B. Structure and evolution of the intergenic region in a ribosomal DNA repeat unit of wheat. J. Mol. Biol. 1988, 201, 1–17. [Google Scholar] [CrossRef]
- Schubert, I.; Shi, F.; Fuchs, J.; Endo, T.R. An efficient screening for terminal deletions and translocations of barley chromosomes added to common wheat. Plant J. 1998, 14, 489–495. [Google Scholar] [CrossRef]
- Han, F.P.; Lamb, J.C.; Birchler, A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Tang, Z.X.; Qiu, L.; Wang, Y.Y.; Tang, S.Y.; Fu, S.L. Identification and physical mapping of new PCR-based markers specific for the long arm of rye (Secale cereale L.) chromosome 6. J. Genet. Genom. 2016, 43, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.X.; Fu, S.L.; Ren, Z.L.; Zhang, H.Q.; Yang, Z.J.; Yan, B.J.; Zhang, H.Y. Production of new wheat cultivar with a different 1B.1R translocation with resistance to powdery mildew and stripe rust. Cereal Res. Commun. 2008, 36, 451–460. [Google Scholar] [CrossRef]
- Szakács, É.; Kruppa, K.; Molnár-Láng, M. Analysis of chromosomal polymorphism in barley (Hordeum vulgare L. ssp. vulgare) and between H. vulgare and H. chilense using three-color fluorescence in situ hybridization (FISH). J. Appl. Genet. 2013, 54, 427–433. [Google Scholar]
- Serizawa, N.; Nasuda, S.; Shi, F.; Endo, T.R.; Prodanovic, S.; Schubert, L.; Künzel, G. Deletion-based physical mapping of barley chromosome 7H. Theor. Appl. Genet. 2001, 103, 827–834. [Google Scholar]
- Türkösi, E.; Cseh, A.; Darkó, É.; Molnár-Láng, M. Addition of manas barley chromosome arms to the hexaploid wheat genome. BMC Genet. 2016, 17, 87. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.X.; Li, M.; Chen, L.; Wang, Y.Y.; Ren, Z.L.; Fu, S.L. New types of wheat chromosomal structural variations in derivatives of wheat-rye hybrids. PLoS ONE 2014, 9, e110282. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Dedkova, O.S.; Gay, G.; Pukhalskyi, V.A.; Zelenin, A.V.; Bernard, S.; Bernard, M. Chromosomal rearrangements in wheat: Their types and distribution. Genome 2007, 50, 907–926. [Google Scholar] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Name | Sequence and Fluorochrome Label | The Amount of Each Slide (ng/Slide) | Sequences Used to Develop Probes (GenBank Accession Number) |
|---|---|---|---|
| Oligo-HvT01 | Tamra-5′AAACT CGCAT TTTTG GCCTA TTCTG GCTAG TTCTG CATGC TATTG CTCAC TGATT TTGG3′ | 11.6 | Barley relic DNA HVT01, tandemly repeated sequence (X16095.1) |
| Oligo-pTa71-1 | 6-FAM-5′GACGC GCGCC ATGGA AAACT3′ | 10.8 | Wheat ribosomal DNA (rDNA) 25S-18S intergenic region EcoRI-BamHI fragment (X07841.1) |
| Oligo-s120.1 | 6-FAM-5′TATCG AGTGC GAGTG AGATA TGCAT GTGTA TGTGT GACCC AGGTG GATGG AGAGT TTGA3′ | 21.8 | Triticum aestivum clone pTa-s120 FISH-positive repetitive sequence (KC290913.1) |
| Oligo-s120.2 | 6-FAM-5′GGAGA GGGAT GAACA AGGTT TTTGT GTCGG ATGCA TGCGA CAGAA TTGAA GATTG TCGG3′ | 21.4 | |
| Oligo-s120.3 | 6-FAM-5′GAGAG AGAGA GAGAG AGAGA GAGAG AGAGA GAGAG AGAGA GAGAG AGAGA GA3′ | 19.0 | |
| Oligo-275.1 | 6-FAM-5′TGCTA CTGCT GCTAC TGCTG CTACT CCTGC TACTG CTACT ACTGC TACTC CTGCT ACTC3′ | 24.6 | Triticum aestivum clone pTa-275 FISH-positive repetitive sequence (KC290911.1) |
| Oligo-275.2 | 6-FAM-5′CTACT ACTAC TACTA CTACT ACTAC TACTA CTACT ACTAC TACTA CTACT ACTAC TACT3′ | 22.6 | |
| Oligo-k566 | 6-FAM-5′ATCCT ACCGA GTGGA GAGCG ACCCT CCCAC TCGGG GGCTT AGCTG CAGTC CAGTA CTCG3′ | 23.3 | Triticum aestivum clone pTa-k566 FISH-positive repetitive sequence (KC290904.1) |
| Oligo-713 | 6-FAM-5′GTCGC GGTAG CGACG ACGGA CGCCG AGACG AGCAC GTGAC ACCAT TCCCA CCCTG TCTA3′ | 23.0 | Triticum aestivum clone pTa-713 FISH-positive repetitive sequence (KC290900.1) |
| Primer Name * | Primer Sequence (5′-3′) | Sequences Used to Design Primer (GenBank Accession Number) |
|---|---|---|
| 120F | AGGAGGGAAGAGCTCTGAGA | Triticum aestivum clone pTa-s120 FISH-positive repetitive sequence (KC290913.1) |
| 120R | GCATAAAACACGACCTCCCC | |
| 275F | TGCTACTACTGCTGCTCCT | Triticum aestivum clone pTa-275 FISH-positive repetitive sequence (KC290911.1) |
| 275R | CAGTAGCAGTAGTAGCAGCAG | |
| 275.1F | GCTACTCCTGCTACTCCTGC | |
| 275.1R | CAGTAGCAGTAGTAGCAGCAG | |
| 566F | TGCAATCCAGTACTCGCCTA | Triticum aestivum clone pTa-k566 FISH-positive repetitive sequence (KC290904.1) |
| 566R | GTCGCTCTCCACTCAGTAGG | |
| 566.1F | TCCAGTACTCGCCTAAGTTTGA | |
| 566.1R | CTGGAATGCAGCTAAGCCTC | |
| 713F | CCTCTGCCACCCTGTCTTAG | Triticum aestivum clone pTa-713 FISH-positive repetitive sequence (KC290900.1) |
| 713R | TAGACAGGGTGGGAATGGTG | |
| 713.1F | GCCACCCTGTCTTAGCGTA | |
| 713.1R | GTAAGATAGACAGGGTGGGAATG | |
| HvT01F | CCTATTCTGGCTAGTTCTGCA | Barley relic DNA HVT01, tandemly repeated seq (X16095.1) |
| HvT01R | AGCATACAAAATTGGCTGGAGT |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, S.; Qiu, L.; Xiao, Z.; Fu, S.; Tang, Z. New Oligonucleotide Probes for ND-FISH Analysis to Identify Barley Chromosomes and to Investigate Polymorphisms of Wheat Chromosomes. Genes 2016, 7, 118. https://doi.org/10.3390/genes7120118
Tang S, Qiu L, Xiao Z, Fu S, Tang Z. New Oligonucleotide Probes for ND-FISH Analysis to Identify Barley Chromosomes and to Investigate Polymorphisms of Wheat Chromosomes. Genes. 2016; 7(12):118. https://doi.org/10.3390/genes7120118
Chicago/Turabian StyleTang, Shuyao, Ling Qiu, Zhiqiang Xiao, Shulan Fu, and Zongxiang Tang. 2016. "New Oligonucleotide Probes for ND-FISH Analysis to Identify Barley Chromosomes and to Investigate Polymorphisms of Wheat Chromosomes" Genes 7, no. 12: 118. https://doi.org/10.3390/genes7120118
APA StyleTang, S., Qiu, L., Xiao, Z., Fu, S., & Tang, Z. (2016). New Oligonucleotide Probes for ND-FISH Analysis to Identify Barley Chromosomes and to Investigate Polymorphisms of Wheat Chromosomes. Genes, 7(12), 118. https://doi.org/10.3390/genes7120118