
Circular RNA circ_0001591 Contributes to Melanoma Cell Migration Through AXL and FRA1 Proteins by Targeting miR-20a-3p and miR-34a-5p


 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results
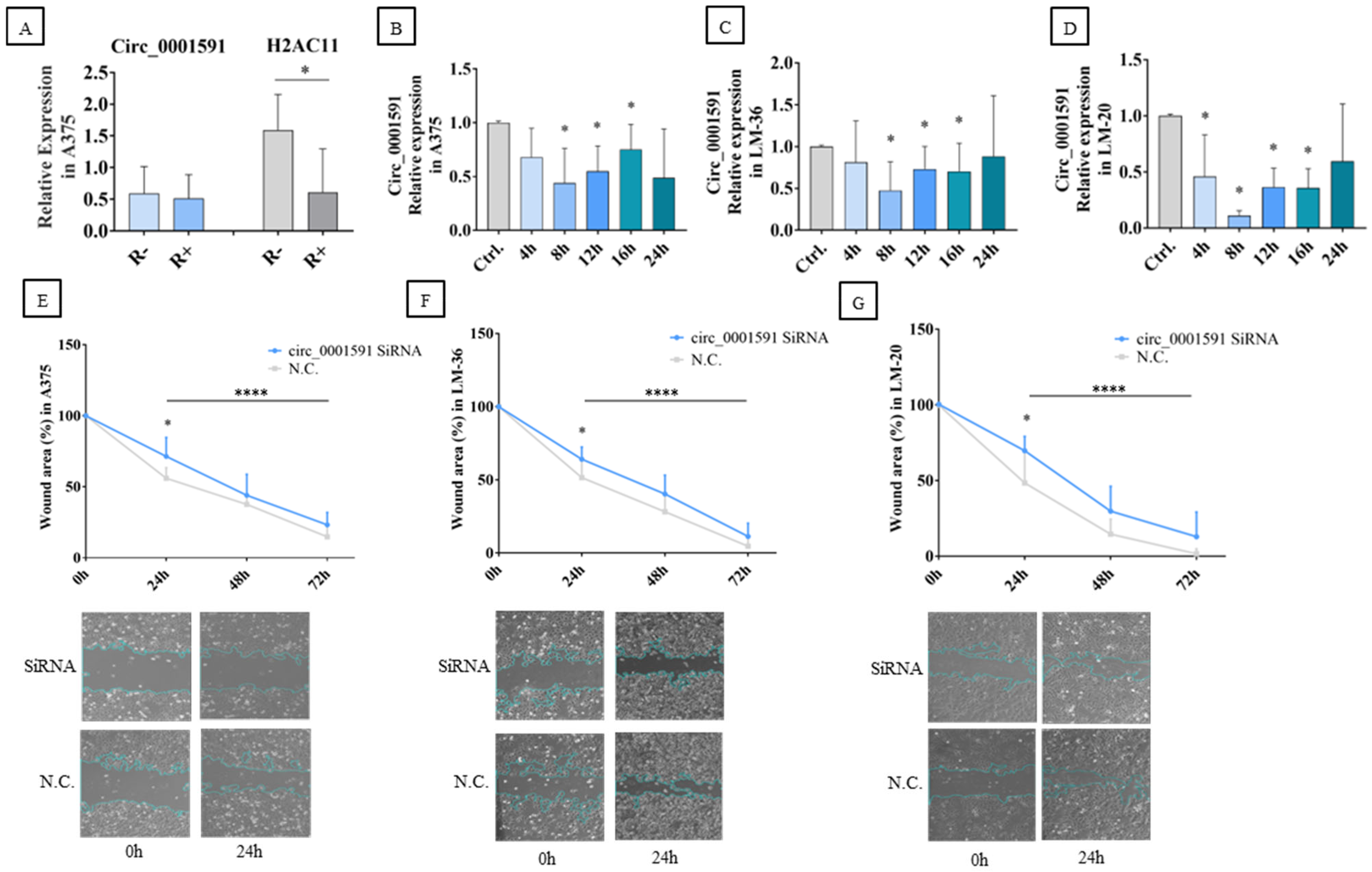
2.1. Circ_0001591 in Melanoma Cell Migration
2.2. Circ_0001591 Targeted miR-20a-3p and miR-34a-5p
2.3. miR-20a-3p and miR-34a-5p in Melanoma Cell Migration
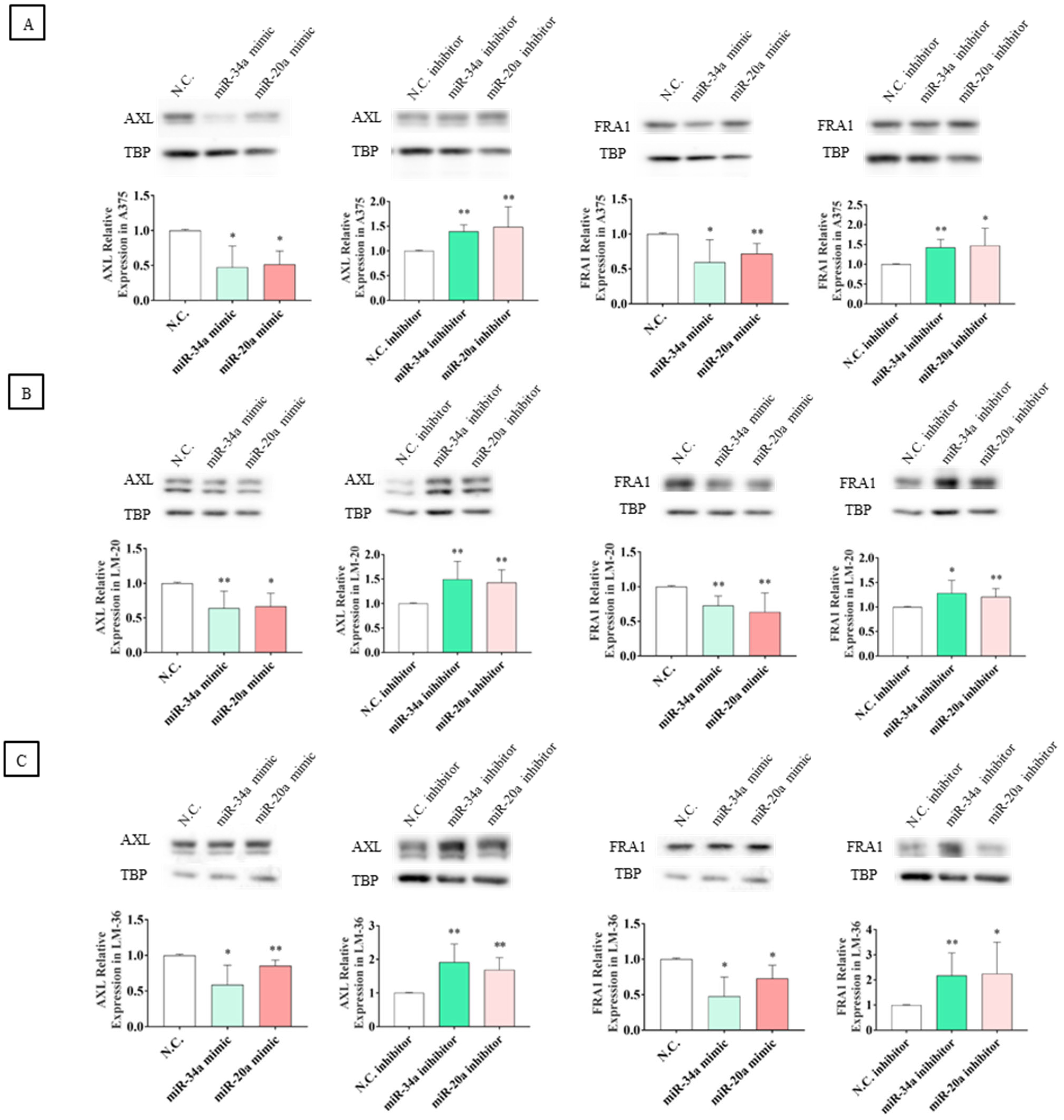
2.4. miR-20a-3p and miR-34a-5p Target AXL and FOSL1 Genes and Negatively Regulate Axl and Fra1 Proteins
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. siRNAs, Mimics, and Inhibitors Transfections
4.3. RNA Extraction and RNase R Treatment
4.4. cDNA Synthesis and Quantitative Real-Time PCR
4.5. Protein Extraction and Western Blot Analysis
4.6. Wound Healing Assay
4.7. Plasmid Construction and Mutagenesis
4.8. Dual Luciferase Reporter Assay
4.9. In Silico Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CircRNAs/Circ | CircularRNAs |
| MiRs/MiRNAs | MicroRNAs |
| ROCK1 | Rho Associated Coiled-Coil Containing Protein Kinase 1 |
| PI3K | Phosphoinositide 3-kinase |
| AKT | Protein kinase B |
| AXL/Axl | AXL Receptor Tyrosine Kinase |
| TAM | TYRO3, AXL, and MERTK receptor tyrosine kinases family |
| MAPK/Erk | Mitogen-Activated Protein Kinase 1 |
| Fra1/FOSL1 | FOS-related antigen 1 |
| AP-1 | Activator protein-1 |
| qRT-PCR | Quantitative Real-Time PCR |
| H2AC11 | H2A Clustered Histone 11 |
| SiRNA | Silencing RNA |
| 3’UTR | 3’ untranslated region |
| NcRNAs | Non-coding RNAs |
| N.C. | Negative control |
| BRAF | B-Raf Proto-Oncogene |
| MEK | Mitogen-activated protein kinase kinase |
| P53 | Tumor protein P53 |
| c-KIT | KIT Proto-Oncogene |
| RPMI | Roswell Park Memorial Institute 1640 |
| NRAS | NRAS Proto-Oncogene |
| EMT | Epithelial–mesenchymal transition |
| SNAIL1 | Snail Family Transcriptional Repressor 1 |
| TRIB2 | Tribbles Pseudokinase 2 |
| CHAF1B | Chromatin Assembly Factor 1 Subunit B |
| MCAM | Melanoma Cell Adhesion Molecule |
| STAT3 | Signal Transducer And Activator Of Transcription 3 |
| c-Met | MET Proto-Oncogene, Receptor Tyrosine Kinase |
| TBP | Tata box protein |
References
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Barsouk, A. Epidemiology of Melanoma. Med. Sci. 2021, 9, 63. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Natarelli, N.; Aleman, S.J.; Mark, I.M.; Tran, J.T.; Kwak, S.; Botto, E.; Aflatooni, S.; Diaz, M.J.; Lipner, S.R. A Review of Current and Pipeline Drugs for Treatment of Melanoma. Pharmaceuticals 2024, 17, 214. [Google Scholar] [CrossRef]
- Gracia-Hernandez, M.; Munoz, Z.; Villagra, A. Enhancing Therapeutic Approaches for Melanoma Patients Targeting Epigenetic Modifiers. Cancers 2021, 13, 6180. [Google Scholar] [CrossRef]
- Teixido, C.; Castillo, P.; Martinez-Vila, C.; Arance, A.; Alos, L. Molecular Markers and Targets in Melanoma. Cells 2021, 10, 2320. [Google Scholar] [CrossRef] [PubMed]
- Corno, C.; Gatti, L.; Lanzi, C.; Zaffaroni, N.; Colombo, D.; Perego, P. Role of the Receptor Tyrosine Kinase Axl and Its Targeting in Cancer Cells. Curr. Med. Chem. 2016, 23, 1496–1512. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, L.; Ren, D.; Liu, C.; Gong, Y.; Wang, N.; Zhang, X.; Zhao, Y. Axl Mediates Tumor Invasion and Chemosensitivity through PI3K/Akt Signaling Pathway and Is Transcriptionally Regulated by Slug in Breast Carcinoma. IUBMB Life 2014, 66, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Boshuizen, J.; Koopman, L.A.; Krijgsman, O.; Shahrabi, A.; Van Den Heuvel, E.G.; Ligtenberg, M.A.; Vredevoogd, D.W.; Kemper, K.; Kuilman, T.; Song, J.Y.; et al. Cooperative Targeting of Melanoma Heterogeneity with an AXL Antibody-Drug Conjugate and BRAF/MEK Inhibitors. Nat. Med. 2018, 24, 203–212. [Google Scholar] [CrossRef]
- Basbous, J.; Chalbos, D.; Hipskind, R.; Jariel-Encontre, I.; Piechaczyk, M. Ubiquitin-Independent Proteasomal Degradation of Fra-1 Is Antagonized by Erk1/2 Pathway-Mediated Phosphorylation of a Unique C-Terminal Destabilizer. Mol. Cell Biol. 2007, 27, 3936–3950. [Google Scholar] [CrossRef]
- Jiang, X.; Xie, H.; Dou, Y.; Yuan, J.; Zeng, D.; Xiao, S. Expression and Function of FRA1 Protein in Tumors. Mol. Biol. Rep. 2020, 47, 737–752. [Google Scholar] [CrossRef]
- Caramel, J.; Papadogeorgakis, E.; Hill, L.; Browne, G.J.; Richard, G.; Wierinckx, A.; Saldanha, G.; Osborne, J.; Hutchinson, P.; Tse, G.; et al. A Switch in the Expression of Embryonic EMT-Inducers Drives the Development of Malignant Melanoma. Cancer Cell 2013, 24, 466–480. [Google Scholar] [CrossRef]
- Di Agostino, S.; Riccioli, A.; De Cesaris, P.; Fontemaggi, G.; Blandino, G.; Filippini, A.; Fazi, F. Circular RNAs in Embryogenesis and Cell Differentiation With a Focus on Cancer Development. Front. Cell Dev. Biol. 2020, 8, 389. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in Human Diseases and Potential Use as Biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef]
- Misir, S.; Wu, N.; Yang, B.B. Specific Expression and Functions of Circular RNAs. Cell Death Differ. 2022, 29, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, X.; Yan, M.; Li, H. Emerging Role of Circular RNAs in Cancer. Front. Oncol. 2020, 10, 663. [Google Scholar] [CrossRef] [PubMed]
- Luan, W.; Shi, Y.; Zhou, Z.; Xia, Y.; Wang, J. Corrigendum to CircRNA_0084043 Promote Malignant Melanoma Progression via MiR-153-3p/Snail Axis. Biochem. Biophys. Res. Commun. 2018, 502, 22–29, Erratum in Biochem. Biophys. Res. Commun. 2022, 587, 168–169. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Dong, D.; Yang, Z.; Xia, R.; Tao, S.; Piao, M. CircMYC Regulates Glycolysis and Cell Proliferation in Melanoma. Cell Biochem. Biophys. 2020, 78, 77–88. [Google Scholar] [CrossRef]
- Tian, S.; Han, G.; Lu, L.; Meng, X. Circ-FOXM1 Contributes to Cell Proliferation, Invasion, and Glycolysis and Represses Apoptosis in Melanoma by Regulating MiR-143-3p/FLOT2 Axis. World J. Surg. Oncol. 2020, 18, 56. [Google Scholar] [CrossRef]
- Bian, D.; Wu, Y.; Song, G. Novel Circular RNA, Hsa_circ_0025039 Promotes Cell Growth, Invasion and Glucose Metabolism in Malignant Melanoma via the MiR-198/CDK4 Axis. Biomed. Pharmacother. 2018, 108, 165–176. [Google Scholar] [CrossRef]
- Wei, C.Y.; Zhu, M.X.; Lu, N.H.; Liu, J.Q.; Yang, Y.W.; Zhang, Y.; Shi, Y.D.; Feng, Z.H.; Li, J.X.; Qi, F.Z.; et al. Circular RNA Circ_0020710 Drives Tumor Progression and Immune Evasion by Regulating the MiR-370-3p/CXCL12 Axis in Melanoma. Mol. Cancer 2020, 19, 84. [Google Scholar] [CrossRef]
- Hanniford, D.; Ulloa-Morales, A.; Karz, A.; Berzoti-Coelho, M.G.; Moubarak, R.S.; Sánchez-Sendra, B.; Kloetgen, A.; Davalos, V.; Imig, J.; Wu, P.; et al. Epigenetic Silencing of CDR1as Drives IGF2BP3-Mediated Melanoma Invasion and Metastasis. Cancer Cell 2020, 37, 55–70.e15. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Wei, G.; Yang, F.; Sun, X. Circular RNA Has Circ 0001591 Promoted Cell Proliferation and Metastasis of Human Melanoma via ROCK1/PI3K/AKT by Targeting MiR-431-5p. Hum. Exp. Toxicol. 2021, 40, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Gupta, A.; Krawczyk, J.; Gupta, S. The MiR-17-92 Cluster: Yin and Yang in Human Cancers. Cancer Treat. Res. Commun. 2022, 33, 100647. [Google Scholar] [CrossRef] [PubMed]
- Branyan, T.E.; Selvamani, A.; Park, M.J.; Korula, K.E.; Kosel, K.F.; Srinivasan, R.; Sohrabji, F. Functional Assessment of Stroke-Induced Regulation of MiR-20a-3p and Its Role as a Neuroprotectant. Transl. Stroke Res. 2022, 13, 432–448. [Google Scholar] [CrossRef]
- Sampath, D.; Branyan, T.E.; Markowsky, K.G.; Gunda, R.; Samiya, N.; Obenaus, A.; Sohrabji, F. Sex Differences in Cognitive Impairment after Focal Ischemia in Middle-Aged Rats and the Effect of Iv MiR-20a-3p Treatment. Neurobiol. Aging 2023, 129, 168–177. [Google Scholar] [CrossRef]
- Liang, Y.; Li, J.; Li, T.; Li, M.; Liao, H.; Liu, Y.; Yao, Y.; Yang, L.; Lei, X. Colorectal Cancer Cells with High Metastatic Potential Drive Metastasis by Transmitting Exosomal MiR-20a-3p through Modulating NF1/MAPK Pathway. Carcinogenesis 2024, 45, 773–785. [Google Scholar] [CrossRef]
- Thomas, P.; Ramani, P.; Ramasubramanian, A.; Sekar, D. Characterizing MiR-20a-3p Expression in the Progression of Oral Squamous Cell Carcinoma. J. Adv. Oral. Res. 2025, 16, 34–42. [Google Scholar] [CrossRef]
- Plata-Bello, J.; Fariña-Jerónimo, H.; Betancor, I.; Salido, E. High Expression of FOXP2 Is Associated with Worse Prognosis in Glioblastoma. World Neurosurg. 2021, 150, e253–e278. [Google Scholar] [CrossRef]
- Pan, W.; Chai, B.; Li, L.; Lu, Z.; Ma, Z. P53/MicroRNA-34 Axis in Cancer and Beyond. Heliyon 2023, 9, e15155. [Google Scholar] [CrossRef]
- Oda, T.; Tsutsumi, K.; Obata, T.; Ueta, E.; Kikuchi, T.; Ako, S.; Fujii, Y.; Yamazaki, T.; Uchida, D.; Matsumoto, K.; et al. MicroRNA-34a-5p: A Pivotal Therapeutic Target in Gallbladder Cancer. Mol. Ther. Oncol. 2024, 32, 200765. [Google Scholar] [CrossRef]
- Chen, S.; Yuan, M.; Chen, H.; Wu, T.; Wu, T.; Zhang, D.; Miao, X.; Shi, J. MiR-34a-5p Suppresses Cutaneous Squamous Cell Carcinoma Progression by Targeting SIRT6. Arch. Dermatol. Res. 2024, 316, 299. [Google Scholar] [CrossRef]
- Yahya, S.M.M.; Nabih, H.K.; Elsayed, G.H.; Mohamed, S.I.A.; Elfiky, A.M.; Salem, S.M. Restoring MicroRNA-34a Overcomes Acquired Drug Resistance and Disease Progression in Human Breast Cancer Cell Lines via Suppressing the ABCC1 Gene. Breast Cancer Res. Treat. 2024, 204, 133–149. [Google Scholar] [CrossRef]
- He, T.; Huang, J.; Liang, H.; Zhong, B.; Xu, G.; Zhu, X. Circular RNA Hsa_circ_0005239 Contributes to Hepatocellular Carcinoma Cell Migration, Invasion, and Angiogenesis by Targeting the MiR-34a-5p/PD-L1 Axis. Cell Biol. Int. 2023, 47, 1519–1534. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Zhu, B.; Han, J.; Huang, J.; Wu, Y. Circ-0036602 Acts As a Sponge of MiR-34a-5p and MiR-431-5p to Promote Cervical Cancer Proliferation and Invasion. J. Genom. 2022, 10, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Nie, D.; Li, B.; Gui, S.B.; Li, C.Z.; Zhang, Y.Z.; Zhao, P. CircNFIX Promotes Progression of Pituitary Adenoma via CCNB1 by Sponging MiR-34a -5p. Mol. Cell. Endocrinol. 2021, 525, 111140. [Google Scholar] [CrossRef] [PubMed]
- Pavri, S.N.; Clune, J.; Ariyan, S.; Narayan, D. Malignant Melanoma: Beyond the Basics. Plast. Reconstr. Surg. 2016, 138, 330e–340e. [Google Scholar] [CrossRef]
- Curti, B.D.; Faries, M.B. Recent Advances in the Treatment of Melanoma. N. Engl. J. Med. 2021, 384, 2229–2240. [Google Scholar] [CrossRef]
- Akbani, R.; Akdemir, K.C.; Aksoy, B.A.; Albert, M.; Ally, A.; Amin, S.B.; Arachchi, H.; Arora, A.; Auman, J.T.; Ayala, B.; et al. Genomic Classification of Cutaneous Melanoma. Cell 2015, 161, 1681–1696. [Google Scholar] [CrossRef]
- Karami Fath, M.; Azargoonjahromi, A.; Soofi, A.; Almasi, F.; Hosseinzadeh, S.; Khalili, S.; Sheikhi, K.; Ferdousmakan, S.; Owrangi, S.; Fahimi, M.; et al. Current Understanding of Epigenetics Role in Melanoma Treatment and Resistance. Cancer Cell Int. 2022, 22, 313. [Google Scholar] [CrossRef]
- Shain, A.H.; Joseph, N.M.; Yu, R.; Benhamida, J.; Liu, S.; Prow, T.; Ruben, B.; North, J.; Pincus, L.; Yeh, I.; et al. Genomic and Transcriptomic Analysis Reveals Incremental Disruption of Key Signaling Pathways during Melanoma Evolution. Cancer Cell 2018, 34, 45–55.e4. [Google Scholar] [CrossRef]
- Lee, J.J.; Murphy, G.F.; Lian, C.G. Melanoma Epigenetics: Novel Mechanisms, Markers, and Medicines. Lab. Investig. 2014, 94, 822–838. [Google Scholar] [CrossRef]
- Casalino, L.; Talotta, F.; Cimmino, A.; Verde, P. The Fra-1/AP-1 Oncoprotein: From the “Undruggable” Transcription Factor to Therapeutic Targeting. Cancers 2022, 14, 1480. [Google Scholar] [CrossRef]
- Nyakas, M.; Fleten, K.G.; Haugen, M.H.; Engedal, N.; Sveen, C.; Farstad, I.N.; Flørenes, V.A.; Prasmickaite, L.; Mælandsmo, G.M.; Seip, K. AXL Inhibition Improves BRAF-Targeted Treatment in Melanoma. Sci. Rep. 2022, 12, 5076. [Google Scholar] [CrossRef]
- Tang, K.; Zhang, H.; Li, Y.; Sun, Q.; Jin, H. Circular RNA as a Potential Biomarker for Melanoma: A Systematic Review. Front. Cell Dev. Biol. 2021, 9, 638548. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, J.; Wa, Q.; He, M.; Wang, X.; Zhou, J.; Cen, Y. Knockdown of Circ_0084043 Suppresses the Development of Human Melanoma Cells through MiR-429/Tribbles Homolog 2 Axis and Wnt/β-Catenin Pathway. Life Sci. 2020, 243, 117323. [Google Scholar] [CrossRef] [PubMed]
- De Tomi, E.; Orlandi, E.; Belpinati, F.; Patuzzo, C.; Trabetti, E.; Gomez-Lira, M.; Malerba, G. New Axes of Interaction in Circ_0079593/MiR-516b-5p Network in Melanoma Metastasis Cell Lines. Genes 2024, 15, 1647. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Selvakumar, S.C.; Auxzilia Preethi, K.; Ramasubramanian, A.; Ramani, P.; Sekar, D. MiRNA-20a: A Dual Regulator of Cell Migration and Apoptosis in Oral Squamous Cell Carcinoma:–An In Vitro Study. J. Orofac. Sci. 2023, 15, 167–174. [Google Scholar] [CrossRef]
- Stope, M.; Ahrend, H.; Daeschlein, G.; Grove, E.; Paditz, M.; Mustea, A.; Burchardt, M. MicroRNA-20a-3p and MicroRNA-20a-5p Exhibit Anti-Proliferative Activities in a Melanoma in Vitro Model. SDRP J. Cell. Mol. Physiol. 2019, 3, 164–170. [Google Scholar] [CrossRef]
- Yan, D.; Zhou, X.; Chen, X.; Hu, D.N.; Da Dong, X.; Wang, J.; Lu, F.; Tu, L.L.; Qu, J. MicroRNA-34a Inhibits Uveal Melanoma Cell Proliferation and Migration through Downregulation of c-Met. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1559–1565. [Google Scholar] [CrossRef]
- MicroRNA-34a Regulates Epithelial-Mesenchymal Transition and Cancer Stem Cell Phenotype of Head and Neck Squamous Cell Carcinoma In Vitro. Available online: https://www.spandidos-publications.com/10.3892/ijo.2015.3142 (accessed on 16 September 2024).
- Adams, B.D.; Wali, V.B.; Cheng, C.J.; Inukai, S.; Booth, C.J.; Agarwal, S.; Rimm, D.L.; Györffy, B.; Santarpia, L.; Pusztai, L.; et al. MiR-34a Silences c-SRC to Attenuate Tumor Growth in Triple-Negative Breast Cancer. Cancer Res. 2016, 76, 927–939. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, Y.; Xie, C.; Zhang, Y. MiR-34a Exerts as a Key Regulator in the Dedifferentiation of Osteosarcoma via PAI-1–Sox2 Axis. Cell Death Dis. 2018, 9, 777. [Google Scholar] [CrossRef]
- Ma, G.; Li, G.; Fan, W.; Xu, Y.; Song, S.; Guo, K.; Liu, Z. Circ-0005105 Activates COL11A1 by Targeting MiR-20a-3p to Promote Pancreatic Ductal Adenocarcinoma Progression. Cell Death Dis. 2021, 12, 656. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Wang, W.; Zhao, G.; Jiang, H.; Zhang, Y.; Zhao, D.; Jin, H.; Yu, H.; Xu, H. Circular RNA Circitga7 Accelerates Glioma Progression via MiR-34a-5p/VEGFA Axis. Aging 2021, 13, 13138–13152. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Teramae, D.; Wells, A. Axl Contributes to Efficient Migration and Invasion of Melanoma Cells. PLoS ONE 2023, 18, e0283749. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.Y.; Huang, J.S.; Shiah, S.G.; Chung, S.Y.; Lay, J.D.; Yang, Y.Y.; Lai, G.M.; Cheng, A.L.; Chen, L.T.; Chuang, S.E. Negative Feedback Regulation of AXL by MiR-34a Modulates Apoptosis in Lung Cancer Cells. RNA 2016, 22, 303–315. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Liu, R.; Kasinski, A.L.; Shen, H.; Slack, F.J.; Tang, D.G. MicroRNA-34a: Potent Tumor Suppressor, Cancer Stem Cell Inhibitor, and Potential Anticancer Therapeutic. Front. Cell Dev. Biol. 2021, 9, 640587. [Google Scholar] [CrossRef]
- Su, Y.F.; Lin, C.S.; Shen, P.C.; Chuang, S.E.; Dai, Y.H.; Huang, T.W.; Lin, C.Y.; Hung, Y.J.; Shieh, Y.S. MiR-34a Functions as a Tumor Suppressor in Oral Cancer through the Inhibition of the Axl/Akt/GSK-3β Pathway. J. Dent. Sci. 2024, 19, 428–437. [Google Scholar] [CrossRef]
- Daniotti, M.; Oggionni, M.; Ranzani, T.; Vallacchi, V.; Campi, V.; Di Stasi, D.; Della Torre, G.; Perrone, F.; Luoni, C.; Suardi, S.; et al. BRAF Alterations Are Associated with Complex Mutational Profiles in Malignant Melanoma. Oncogene 2004, 23, 5968–5977. [Google Scholar] [CrossRef]
- Xiao, M.-S.; Wilusz, J.E. An Improved Method for Circular RNA Purification Using RNase R That Efficiently Removes Linear RNAs Containing G-Quadruplexes or Structured 3 Ends. Nucleic Acids Res. 2019, 47, 8755–8769. [Google Scholar] [CrossRef]
- Fochi, S.; Orlandi, E.; Ceccuzzi, L.; Rodolfo, M.; Vergani, E.; Turco, A.; Romanelli, M.G.; Gomez-Lira, M. Identification of Suitable MRNAs and MicroRNAs as Reference Genes for Expression Analyses in Skin Cells under Sex Hormone Exposure. Gene 2021, 769, 145336. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SiRNA | Sequence F (5′-3′) | Sequence R (5′-3′) |
|---|---|---|
| Hsa_circ_0001591_1 | AGGCGGCAAAGCCCGCGCUAATT | UUAGCGCGGGCUUUGCCGCCUTT |
| Hsa_circ_0001591_2 | GUGGCAAGCAAGGCGGCAAAGTT | CUUUGCCGCCUUGCUUGCCACTT |
| Negative Control (NC) | UUCUCCGAACGUGUCACGUTT | ACGUGACACGUUCGGAGAATT |
| Gene/miR | Primers/Probe | Bp/Assay |
|---|---|---|
| AXL | F: 5′ ATTGGCTTCGGGATGGACAG 3′ | 125 |
| R: 5′ AAGCTCCAGGGAGGTGATTC 3′ | ||
| FOSL1 | F: 5′ GCCCTTGTGAACAGATCAGC 3′ | 137 |
| R: 5′ CAGTTTGTCAGTCTCCGCCT 3′ | ||
| Circ_0001591 | F: 5′ TCGCCCCCAATAGTGTTTAT 3′ | 136 |
| R: 5′ GCCCCTAGAAGAGCGAGTCT 3′ | ||
| H2AC11 | F: 5′ AAGCCCAAGACTCGCTCTTC 3′ | 82 |
| R: 5′ CATAGTTGCCTTTGCGGAGC 3′ | ||
| TBP | F: 5′ TGTATCCACAGTGAATCTTGG 3′ | 102 |
| R: 5′ ATGATTACCGCAGCAAACC 3′ | ||
| hsa-miR-20a-3p | ACUGCAUUAUGAGCACUUAAAG | 478317_mir |
| hsa-miR-34a-5p | UGGCAGUGUCUUAGCUGGUUGU | 478048_mir |
| hsa-miR-191-5p | CAACGGAAUCCCAAAAGCAGCUG | 477952_mir |
| Plasmid | Primers (5′-3′) | Bp |
|---|---|---|
| Circ_0001591_1 | F: CCTAAGGT TCTAGA GTCATGGCACTGGTAGGAGT | 304 |
| R: CCTAAGGT TCTAGA AGCAAACTTTGTCAGAGGCG | ||
| Circ_0001591_2 | F: CCTAAGGT TCTAGA ACAGCAAGTTACAGCCAGTCA | 317 |
| R: ACCTTAGG TCTAGA TGAGTTGGTTAGTCAGCACAA | ||
| Circ_0001591_3 | F: CCTAAGGT TCTAGA GTCCCCTCCCCCCAGAGGGACTTC | 337 |
| R: ACCTTAGG TCTAGA GAAGTCCCTCTGGGGGGAGGGGAC | ||
| Circ_0001591_4 | F: CCTAAGGT TCTAGA AAGTCAGGAGTTCAGGACCAG | 389 |
| R: ACCTTAGG TCTAGAG GCAAATCTCACAACTCAATGCC | ||
| AXL_miR-20a-3p | F: CCTAAGGT TCTAGA GTGCCCCTCTCCTTCTTAGC | 215 |
| R: ACCTTAGG TCTAGA TATGTGACCTGAGCCCCTCT | ||
| AXL_miR-34a-5p | F: CCTAAGGT TCTAGA AGACAACGCTCCACCTGGTA | 164 |
| R: ACCTTAGG TCTAGA TGCTACTCCACAGAGAAGGG | ||
| FOSL1 | F: CCTAAGGT TCTAGA AACCCTCCTCGCTTTGTGAG | 893 |
| R: ACCTTAGG TCTAGA AGCCTCTTCGCTTTTACCCC |
| Plasmid | Primers (5′-3′) | Mutagenesis |
|---|---|---|
| Circ_0001591_1 | F: CTTAAGCCTCTGGG CAGCCCAGTAGGCC | Δ GAAT |
| R: GGCCTACTGGGCTGCCCAGAGGCTTAAG | ||
| Circ_0001591_2 | F: TGGAATACTTTGAAAATACTTCAAGGCTGTAAGGTACTCATTAAAATAAAG | Δ CACT |
| R: CTTTATTTTAATGAGTACCTTACAGCCTTGAAGTATTTTCAAAGTATTCCAC | ||
| Circ_0001591_3 | F: GTCCCCTCCCCCCAGAGGGACTTC | Δ AATG |
| R: GAAGTCCCTCTGGGGGGAGGGGAC | ||
| Circ_0001591_4 | F: AGCTGACACAGTGCGCACTCCAGCCTTG | Δ CACT |
| R: CAAGGCTGGAGTGCGCACTGTGTCAGCT | ||
| AXL_miR-20a-3p | F: CCTTCAAGCCTGTGCAAT TAGGGATGCCTCCTTT | Δ GCAT |
| R: AAAGGAGGCATCCCTAATTGCACAGGCTTGAAGG | ||
| AXL_miR-34a-5p | F: GATCCAAGCTAAGCACTCTGGGGAAAACTCCACC | Δ GCCA |
| R: GGTGGAGTTTTCCCCAGAGTGCTTAGCTTGGATC | ||
| FOSL1_1 | F: CCCCTTCCAGATCATATCTGCCACACTCTCC | Ins TAT |
| R: GGAGAGTGTGGCAGATATGATCTGGAAGGGG | ||
| FOSL1_2 | F: ACTCACCAGCCCCACTGCGAGCAGCAGCAGGT | C→G |
| R: ACCTGCTGCTGCTCGCAGTGGGGCTGGTGAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlandi, E.; De Tomi, E.; Belpinati, F.; Menegazzi, M.; Gomez-Lira, M.; Romanelli, M.G.; Trabetti, E. Circular RNA circ_0001591 Contributes to Melanoma Cell Migration Through AXL and FRA1 Proteins by Targeting miR-20a-3p and miR-34a-5p. Genes 2025, 16, 921. https://doi.org/10.3390/genes16080921
Orlandi E, De Tomi E, Belpinati F, Menegazzi M, Gomez-Lira M, Romanelli MG, Trabetti E. Circular RNA circ_0001591 Contributes to Melanoma Cell Migration Through AXL and FRA1 Proteins by Targeting miR-20a-3p and miR-34a-5p. Genes. 2025; 16(8):921. https://doi.org/10.3390/genes16080921
Chicago/Turabian StyleOrlandi, Elisa, Elisa De Tomi, Francesca Belpinati, Marta Menegazzi, Macarena Gomez-Lira, Maria Grazia Romanelli, and Elisabetta Trabetti. 2025. "Circular RNA circ_0001591 Contributes to Melanoma Cell Migration Through AXL and FRA1 Proteins by Targeting miR-20a-3p and miR-34a-5p" Genes 16, no. 8: 921. https://doi.org/10.3390/genes16080921
APA StyleOrlandi, E., De Tomi, E., Belpinati, F., Menegazzi, M., Gomez-Lira, M., Romanelli, M. G., & Trabetti, E. (2025). Circular RNA circ_0001591 Contributes to Melanoma Cell Migration Through AXL and FRA1 Proteins by Targeting miR-20a-3p and miR-34a-5p. Genes, 16(8), 921. https://doi.org/10.3390/genes16080921