
Gene Expression Profile of the Cerebral Cortex of Niemann-Pick Disease Type C Mutant Mice


 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of the Tissue
2.3. RNA Extraction and cDNA Synthesis
2.4. Microarray Analysis
2.5. Bioinformatic Analysis
2.6. Gene Expression by qPCR
2.7. Determination of Protein Content by Western Blotting
2.8. Statistical Analysis
3. Results
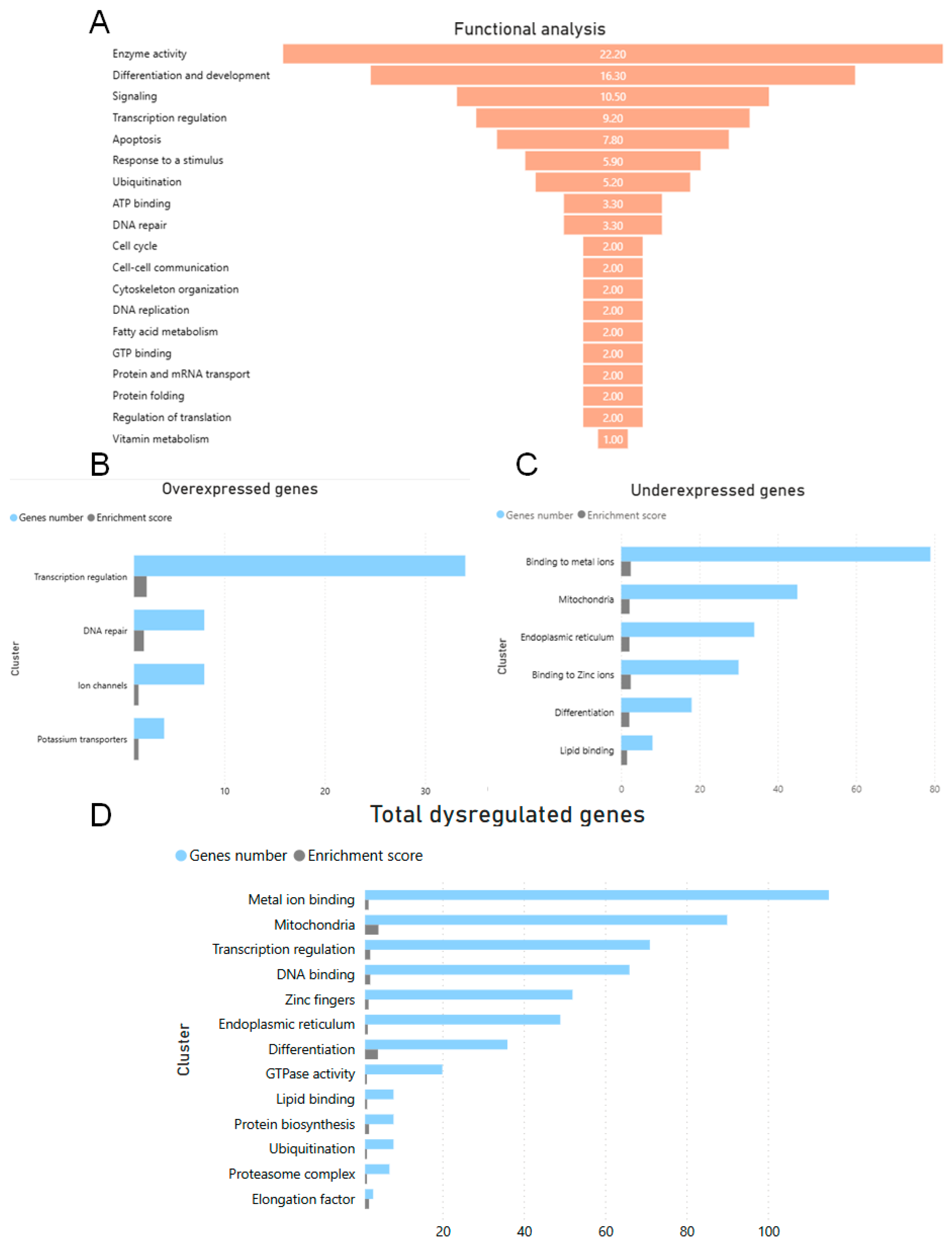
3.1. Functional and Cluster Classification of DEGs in the Cerebral Cortex of the NPC Mouse Mutant
3.2. KEGG Analysis of DEGs in the Cerebral Cortex of the NPC Mouse
3.3. Relevant Genes in Bioinformatics Analysis
3.4. Validation of Microarray Expression Data by qPCR and Determination of Altered Pathways by Western Blot Protein Expression
3.4.1. Expression Analysis Reveals the Dysregulation in Genes Involved in Intrinsic Apoptosis and Alterations in the 1C Metabolic Pathway in the NPC Mouse
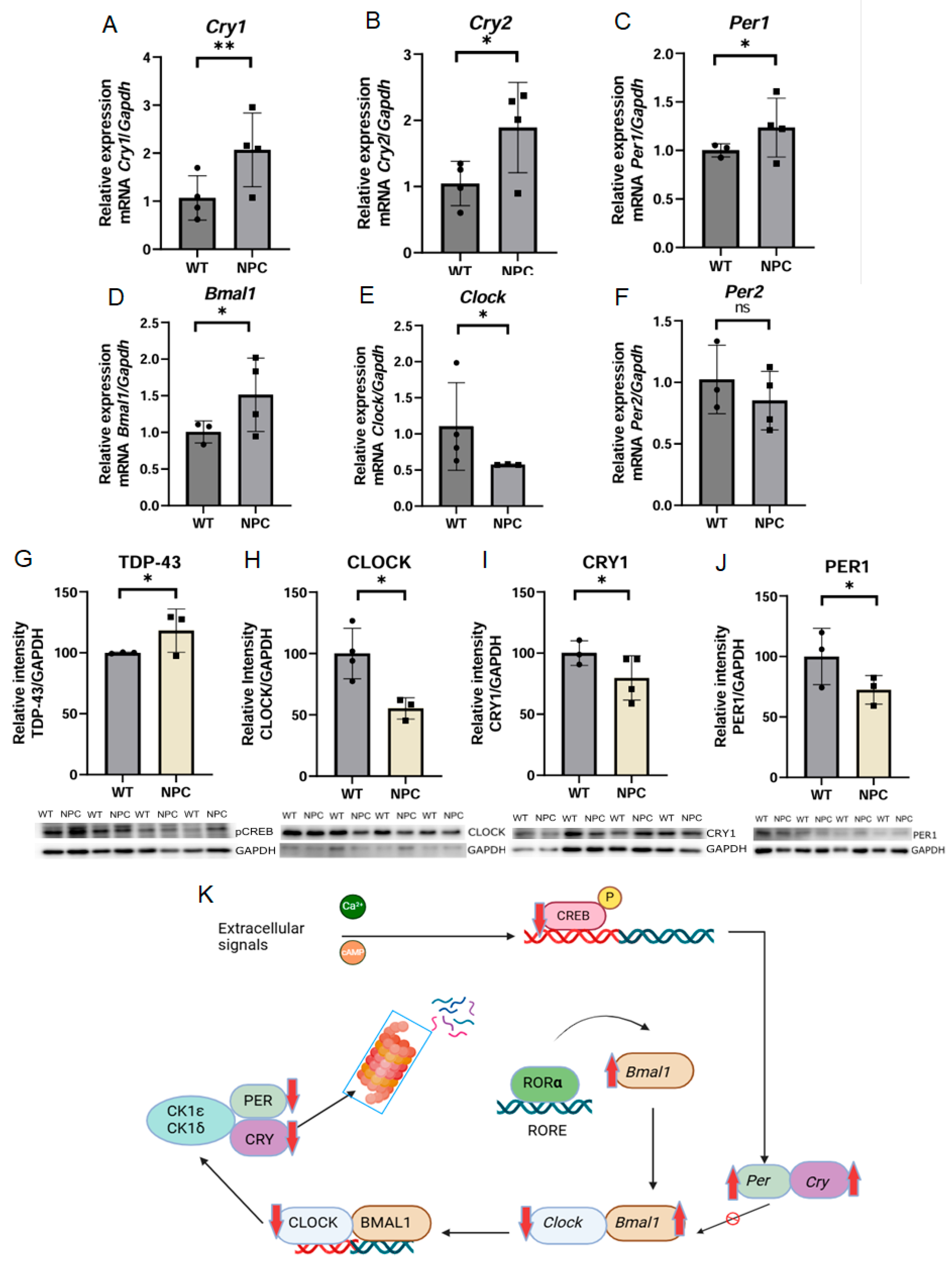
3.4.2. Alteration in the Cortical Circadian Rhythm Pathway in the Cerebral Cortex of NPC Mice
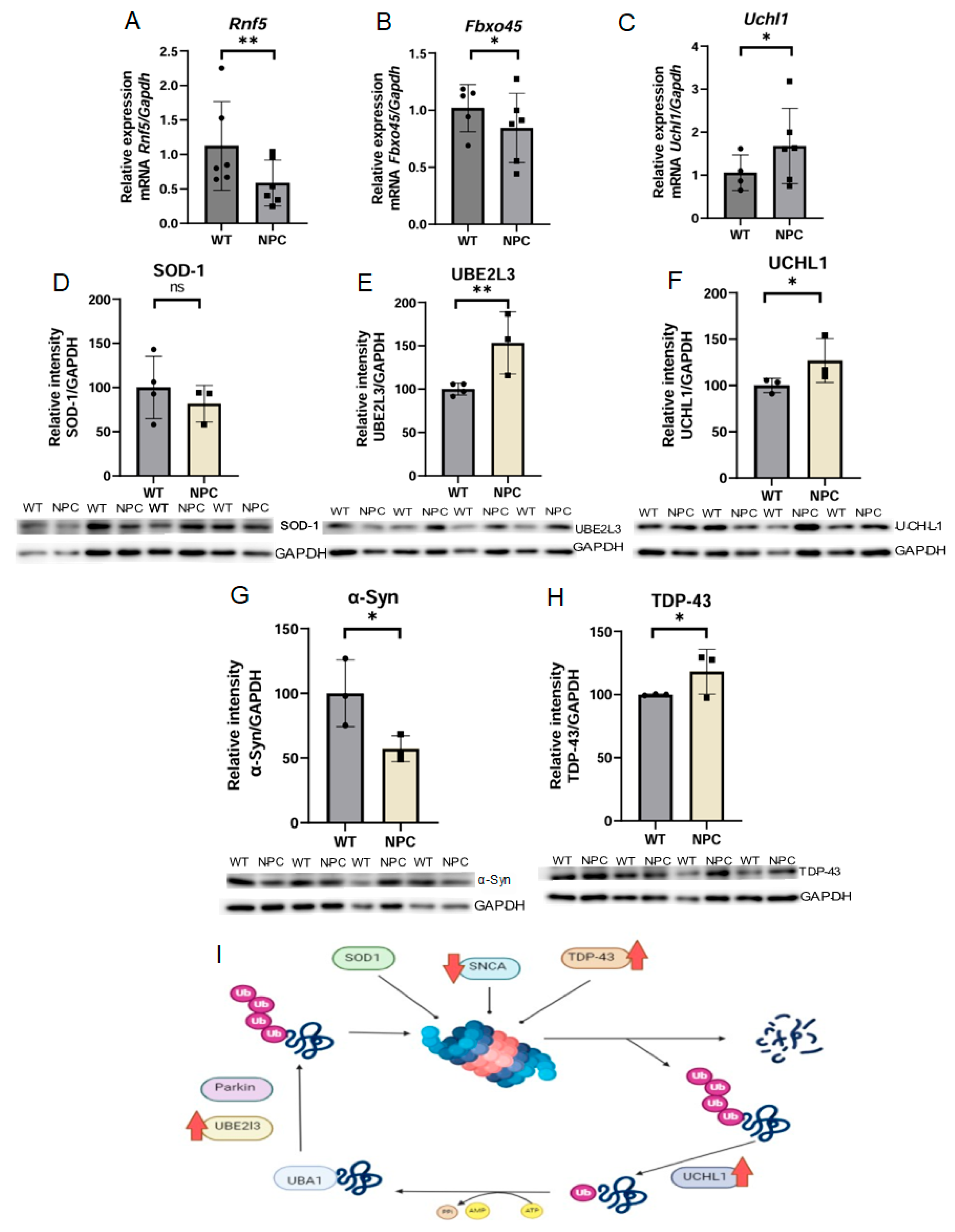
3.4.3. Genes Associated with Ubiquitination Are Altered and Show Increased Levels of TDP-43 in the Cerebral Cortex of the NPC Mouse Mutant
3.4.4. The Cerebral Cortex of the NPC Mouse Shows Changes in Genes Associated with Differentiation and Development
3.4.5. The Cerebral Cortex of NPC Mice Shows a Dysregulation of Proteins Involved in the Beta-Oxidation of VLCFA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCD1 | ATP-binding cassette, sub-family D (ALD), member 1 |
| ACAA1 | Acetyl-CoA Acyltransferase 1 |
| ACOX1 | Acyl-CoA Oxidase 1 |
| BBS9 | Bardet–Biedl syndrome 9 |
| BDNF | Brain-Derived Neurotrophic Factor |
| BMAL1 | Basic Helix-Loop-Helix ARNT Like 1 |
| BMP4 | Bone morphogenetic protein 4 |
| cDNA | Complementary DNA |
| CNS | Central Nervous System |
| CREB | CAMP Responsive Element Binding Protein |
| CRY1 | Cryptochrome Circadian Regulator 1 |
| CRY2 | Cryptochrome Circadian Regulator 2 |
| Ct | Cycling threshold |
| CYP24A1 | Cytochrome P450, family 24, subfamily a, polypeptide 1 |
| DEGs | Differentially expressed genes |
| dUTP | Deoxyuridine Triphosphate |
| FBS | Fetal bovine serum |
| FBXO45 | F-box protein 45 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GHITM | Growth hormone inducible transmembrane protein |
| GHRH | Growth hormone releasing hormone |
| GnRH | Gonadotropin-Releasing Hormone |
| GPAM | Glycerol-3-phosphate acyltransferase, mitochondrial |
| HIV | Human immunodeficiency virus |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LSD | Lysosomal Storage Diseases |
| MGI | Mouse Genome Informatics |
| MHFDL2 | Methylenetetrahydrofolate Dehydrogenase NADP+ Dependent 2 Like |
| mRNA | Messenger ribonucleic acid |
| MTHD1 | Methylenetetrahydrofolate Dehydrogenase, Cyclohydrolase and Formyltetrahydrofolate Synthetase 1 |
| NPC | Niemann Pick type C |
| NPC1 | NPC Intracellular Cholesterol Transporter 1 protein |
| NPC2 | NPC Intracellular Cholesterol Transporter 2 protein |
| PER1 | Period Circadian Regulator 1 |
| PPM1F | Protein phosphatase 1F |
| PVDF | Polyvinylidene difluoride |
| qPCR | Quantitative polymerase chain reaction |
| RARG | Retinoic acid receptor, gamma |
| RNA | Ribonucleic acid |
| RNF5 | Ring finger protein 5 |
| SEM | Standard error of the mean |
| SFXN1 | Sideroflexin 1 |
| SGPL1 | Sphingosine- 1-Phosphate Lyase 1 |
| SLC39A1 | Solute carrier family 39 zinc transporter member 1 |
| SMPD1 | Sphingomyelin phosphodiesterase 1, acid lysosomal |
| SOD- 1 | Superoxide Dismutase 1 |
| SPHK1 | Sphingosine Kinase 1 |
| TBS | Tris-buffered saline |
| TDP-43 | TAR DNA-binding protein 43 |
| TYMS | Thymidylate synthase |
| UBE2L3 | Ubiquitin Conjugating Enzyme E2 L3 |
| UCHL1 | Ubiquitin carboxy-terminal hydrolase L1 |
| VLCFA | Very Long Chain Fatty Acids |
| WT | Wild type |
| α-Syn | α-synuclein |
| PGP | Phosphatidylglycerophosphate |
| PG | Phosphatidylglycerol |
| X-ALD | X-linked adrenoleukodystrophy |
| CALD | Cerebral adrenoleukodystrophy |
| AMN | Adrenomyeloneuropathy |
References
- Geberhiwot, T.; Moro, A.; Dardis, A.; Ramaswami, U.; Sirrs, S.; Marfa, M.P.; Vanier, M.T.; Walterfang, M.; Bolton, S.; Dawson, C.; et al. International Niemann-Pick Disease Registry (INPDR) Consensus clinical management guidelines for Niemann-Pick disease type C. Orphanet J. Rare Dis. 2018, 13, 50. [Google Scholar] [CrossRef] [PubMed]
- Peake, K.B.; Vance, J.E. Defective cholesterol trafficking in Niemann-Pick C-deficient cells. FEBS Lett. 2010, 584, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Pallottini, V.; Pfrieger, F.W. Understanding and Treating Niemann-Pick Type C Disease: Models Matter. Int. J. Mol. Sci. 2020, 21, E8979. [Google Scholar] [CrossRef] [PubMed]
- Bräuer, A.U.; Kuhla, A.; Holzmann, C.; Wree, A.; Witt, M. Current Challenges in Understanding the Cellular and Molecular Mechanisms in Niemann-Pick Disease Type C1. Int. J. Mol. Sci. 2019, 20, E4392. [Google Scholar] [CrossRef] [PubMed]
- Griñán-Ferré, C.; Companys-Alemany, J.; Jarné-Ferrer, J.; Codony, S.; González-Castillo, C.; Ortuño-Sahagún, D.; Vilageliu, L.; Grinberg, D.; Vázquez, S.; Pallàs, M. Inhibition of Soluble Epoxide Hydrolase Ameliorates Phenotype and Cognitive Abilities in a Murine Model of Niemann Pick Type C Disease. Int. J. Mol. Sci. 2021, 22, 3409. [Google Scholar] [CrossRef] [PubMed]
- Tharkeshwar, A.K.; Trekker, J.; Vermeire, W.; Pauwels, J.; Sannerud, R.; Priestman, D.A.; Vruchte, D.T.; Vints, K.; Baatsen, P.; Decuypere, J.-P.; et al. A novel approach to analyze lysosomal dysfunctions through subcellular proteomics and lipidomics: The case of NPC1 deficiency. Sci. Rep. 2017, 7, 41408. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Hou, Q.; Cheung, N.S.; Li, Q.-T. Neuronal cell death caused by inhibition of intracellular cholesterol trafficking is caspase dependent and associated with activation of the mitochondrial apoptosis pathway. J. Neurochem. 2006, 97, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Grau, M.; Albaigès, J.; Casas, J.; Auladell, C.; Dierssen, M.; Vilageliu, L.; Grinberg, D. New murine Niemann-Pick Type C models bearing a pseudoexon-generating mutation recapitulate the main neurobehavioural. Sci. Rep. 2017, 7, 41931. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Mayorquín, A.E.; Torres-Ruíz, N.M.; Ortuño-Sahagún, D.; Gudiño-Cabrera, G. Microarray analysis of striatal embryonic stem cells induced to differentiate by ensheathing cell conditioned media. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2008, 237, 979–994. [Google Scholar] [CrossRef] [PubMed]
- Ortuño-Sahagún, D.; Rivera-Cervantes, M.C.; Gudiño-Cabrera, G.; Junyent, F.; Verdaguer, E.; Auladell, C.; Pallàs, M.; Camins, A.; Beas-Zárate, C. Microarray analysis of rat hippocampus exposed to excitotoxicity: Reversal Na+/Ca2+ exchanger NCX3 is overexpressed in glial cells. Hippocampus 2010, 22, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Rodríguez, P.; Godínez-Rubí, M.; Guzmán-Brambila, C.; Padilla-Velarde, E.; Orozco-Barocio, A.; Ortuño-Sahagún, D.; Rojas-Mayorquín, A.E. Prenatal Alcohol Exposure in Rats Diminishes Postnatal Cxcl16 Chemokine Ligand Brain Expression. Brain Sci. 2020, 10, 987. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mayen, A.P.; Corral-Guillé, G.; Riego-Ruíz, L.; Coello Coutino, G. The Package “GenArise” (API); Version 3.21, Microarray Analysis Tool; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2006; Available online: https://bioconductor.org/packages/genArise (accessed on 13 June 2025).
- Linares, P.S.; Carvajal, M.A.; Coello, G. Package genArise2. 2010. Available online: https://bioc.r-universe.dev/genArise/doc/manual.html (accessed on 1 August 2024).
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Xiao, W.C.; Li, H.; Hao, Z.Y.; Liu, G.Z.; Zhang, D.H.; Wu, L.M.; Wang, Z.; Zhang, Y.Q.; Huang, Z.; et al. E3 ubiquitin ligase RNF5 attenuates pathological cardiac hypertrophy through STING. Cell Death Dis. 2022, 13, 889. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.A.; Baldarelli, R.; Kadin, J.A.; Richardson, J.E.; Smith, C.L.; Bult, C.J. Mouse Genome Database (MGD): Knowledgebase for mouse-human comparative biology. Nucleic Acids Res. 2021, 49, D981–D987. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.; Taishi, P.; Churchill, L.; Urza, M.J.; Krueger, J.M. Localized suppression of cortical growth hormone-releasing hormone receptors state-specifically attenuates electroencephalographic delta waves. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 4151–4159. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.H.; Yang, C.; Tse, A.P.; Wei, L.; Lee, D.; Zhang, M.S.; Goh, C.C.; Chiu, D.K.; Yuen, V.W.; Law, C.T.; et al. Genome-wide CRISPR-Cas9 knockout library screening identified PTPMT1 in cardiolipin synthesis is crucial to survival in hypoxia in liver cancer. Cell Rep. 2021, 34, 108676. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Tyurin, V.A.; Jiang, J.; Tyurina, Y.Y.; Ritov, V.B.; Amoscato, A.A.; Osipov, A.N.; Belikova, N.A.; Kapralov, A.A.; Kini, V.; et al. Cytochrome c acts as a cardiolipin oxygenase required for release of proapoptotic factors. Nat. Chem. Biol. 2005, 1, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.A.; Ryan, R.O. Studies of the cardiolipin interactome. Prog. Lipid Res. 2022, 88, 101195. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Sayano, T.; Tamai, S.; Yokota, S.; Kato, H.; Fujii, G.; Mihara, K. Identification of a novel protein MICS1 that is involved in maintenance of mitochondrial morphology and apoptotic release of cytochrome c. Mol. Biol. Cell 2008, 19, 2597–2608. [Google Scholar] [CrossRef] [PubMed]
- Acoba, M.G.; Alpergin, E.S.S.; Renuse, S.; Fernández-Del-Río, L.; Lu, Y.W.; Khalimonchuk, O.; Clarke, C.F.; Pandey, A.; Wolfgang, M.J.; Claypool, S.M. The mitochondrial carrier SFXN1 is critical for complex III integrity and cellular metabolism. Cell Rep. 2021, 34, 108869. [Google Scholar] [CrossRef] [PubMed]
- Kory, N.; Wyant, G.A.; Prakash, G.; Uit de Bos, J.; Bottanelli, F.; Pacold, M.E.; Chan, S.H.; Lewis, C.A.; Wang, T.; Keys, H.R.; et al. SFXN1 is a mitochondrial serine transporter required for one-carbon metabolism. Science 2018, 362, eaat9528. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, A.J.; Perry, C.A.; Girnary, H.H.; Gao, D.; Allen, R.H.; Stabler, S.P.; Shane, B.; Stover, P.J. MTHFD1 is an essential gene in mice and alters biomarkers of impaired one-carbon metabolism. J. Biol. Chem. 2009, 284, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Moran, R.G.; Goldman, I.D. Pemetrexed: Biochemical and cellular pharmacology, mechanisms, and clinical applications. Mol. Cancer Ther. 2007, 6, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Racanelli, A.C.; Rothbart, S.B.; Heyer, C.L.; Moran, R.G. Therapeutics by cytotoxic metabolite accumulation: Pemetrexed causes ZMP accumulation, AMPK activation, and mammalian target of rapamycin inhibition. Cancer Res. 2009, 69, 5467–5474. [Google Scholar] [CrossRef] [PubMed]
- Servín Muñoz, I.V.; Ortuño-Sahagún, D.; Griñán-Ferré, C.; Pallàs, M.; González-Castillo, C. Alterations in Proteostasis Mechanisms in Niemann-Pick Type C Disease. Int. J. Mol. Sci. 2024, 25, 3806. [Google Scholar] [CrossRef] [PubMed]
- Cawley, N.X.; Giddens, S.; Farhat, N.M.; Luke, R.A.; Scott, K.E.J.; Mohamed, H.O.; Dang Do, A.; Berry-Kravis, E.; Cologna, S.M.; Liu, F.; et al. Elevated cerebrospinal fluid ubiquitin C-terminal hydrolase-L1 levels correlate with phenotypic severity and therapeutic response in Niemann-Pick disease, type C1. Mol. Genet. Metab. 2023, 140, 107656. [Google Scholar] [CrossRef] [PubMed]
- Tramutola, A.; Di Domenico, F.; Barone, E.; Perluigi, M.; Butterfield, D.A. It Is All about (U)biquitin: Role of Altered Ubiquitin-Proteasome System and UCHL1 in Alzheimer Disease. Oxidative Med. Cell Longev. 2016, 2016, 2756068. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Vollmer, S.; Golombek, S.; Kahle, P.J. The ubiquitin-conjugating enzymes UBE2N, UBE2L3 and UBE2D2/3 are essential for Parkin-dependent mitophagy. J. Cell Sci. 2014, 127, 3280–3293. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.J.; Cui, P.; Li, H.; Lang, W.J.; Liu, G.Y.; Ma, X.F. Shared genes between Alzheimer’s disease and ischemic stroke. CNS Neurosci. Ther. 2019, 25, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Dardis, A.; Zampieri, S.; Canterini, S.; Newell, K.L.; Stuani, C.; Murrell, J.R.; Ghetti, B.; Fiorenza, M.T.; Bembi, B.; Buratti, E. Altered localization and functionality of TAR DNA Binding Protein 43 (TDP-43) in niemann- pick disease type C. Acta Neuropathol. Commun. 2016, 4, 52. [Google Scholar] [CrossRef] [PubMed]
- Chiba, Y.; Komori, H.; Takei, S.; Hasegawa-Ishii, S.; Kawamura, N.; Adachi, K.; Nanba, E.; Hosokawa, M.; Enokido, Y.; Kouchi, Z.; et al. Niemann-Pick disease type C1 predominantly involving the frontotemporal region, with cortical and brainstem Lewy bodies: An autopsy case. Neuropathol. Off. J. Jpn. Soc. Neuropathol. 2014, 34, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Ribas, G.S.; Pires, R.; Coelho, J.C.; Rodrigues, D.; Mescka, C.P.; Vanzin, C.S.; Biancini, G.B.; Negretto, G.; Wayhs, C.A.; Wajner, M.; et al. Oxidative stress in Niemann-Pick Type C patients: A protective role of N-butyl-deoxynojirimycin therapy. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2012, 30, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Adir, O.; Bening-Abu-Shach, U.; Arbib, S.; Henis-Korenblit, S.; Broday, L. Inactivation of the Caenorhabditis elegans RNF-5 E3 ligase promotes IRE-1-independent ER functions. Autophagy 2021, 17, 2401–2414. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.; Calvo-Jiménez, E.; Kon, E.; Cao, H.; Jossin, Y.; Cooper, J.A. Fbxo45 Binds SPRY Motifs in the Extracellular Domain of N-Cadherin and Regulates Neuron Migration during Brain Development. Mol. Cell Biol. 2020, 40, e00539-19. [Google Scholar] [CrossRef] [PubMed]
- Saiga, T.; Fukuda, T.; Matsumoto, M.; Tada, H.; Okano, H.J.; Okano, H.; Nakayama, K.I. Fbxo45 forms a novel ubiquitin ligase complex and is required for neuronal development. Mol. Cell Biol. 2009, 29, 3529–3543. [Google Scholar] [CrossRef] [PubMed]
- Manor, J.; Chung, H.; Bhagwat, P.K.; Wangler, M.F. ABCD1 and X-linked adrenoleukodystrophy: A disease with a markedly variable phenotype showing conserved neurobiology in animal models. J. Neurosci. Res. 2021, 99, 3170–3181. [Google Scholar] [CrossRef] [PubMed]
- Buda, A.; Forss-Petter, S.; Hua, R.; Jaspers, Y.; Lassnig, M.; Waidhofer-Söllner, P.; Kemp, S.; Kim, P.; Weinhofer, I.; Berger, J. ABCD1 Transporter Deficiency Results in Altered Cholesterol Homeostasis. Biomolecules 2023, 13, 1333. [Google Scholar] [CrossRef] [PubMed]
- Weinhofer, I.; Forss-Petter, S.; Kunze, M.; Zigman, M.; Berger, J. X-linked adrenoleukodystrophy mice demonstrate abnormalities in cholesterol metabolism. FEBS Lett. 2005, 579, 5512–5516. [Google Scholar] [CrossRef] [PubMed]
- Veleri, S.; Bishop, K.; Dalle Nogare, D.E.; English, M.A.; Foskett, T.J.; Chitnis, A.; Sood, R.; Liu, P.; Swaroop, A. Knockdown of Bardet-Biedl syndrome gene BBS9/PTHB1 leads to cilia defects. PLoS ONE 2012, 7, e34389. [Google Scholar] [CrossRef] [PubMed]
- Tsang, S.H.; Aycinena, A.R.P.; Sharma, T. Ciliopathy: Bardet-Biedl Syndrome. Adv. Exp. Med. Biol. 2018, 1085, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Meyers, E.A.; Kessler, J.A. TGF-β Family Signaling in Neural and Neuronal Differentiation, Development, and Function. Cold Spring Harb. Perspect. Biol. 2017, 9, a022244. [Google Scholar] [CrossRef] [PubMed]
- Bakrania, P.; Efthymiou, M.; Klein, J.C.; Salt, A.; Bunyan, D.J.; Wyatt, A.; Ponting, C.P.; Martin, A.; Williams, S.; Lindley, V.; et al. Mutations in BMP4 cause eye, brain, and digit developmental anomalies: Overlap between the BMP4 and hedgehog signaling pathways. Am. J. Hum. Genet. 2008, 82, 304–319. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Gu, X.; Zhang, Z.; Li, W.; Wang, X. Retinoic acid receptor gamma is targeted by microRNA-124 and inhibits neurite outgrowth. Neuropharmacology 2020, 163, 107657. [Google Scholar] [CrossRef] [PubMed]
- Bering, T.; Carstensen, M.B.; Wörtwein, G.; Weikop, P.; Rath, M.F. The Circadian Oscillator of the Cerebral Cortex: Molecular, Biochemical and Behavioral Effects of Deleting the Arntl Clock Gene in Cortical Neurons. Cereb. Cortex 2018, 28, 644–657. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.A.; Polino, A.J.; King, M.W.; Musiek, E.S. Circadian clock protein BMAL1 broadly influences autophagy and endolysosomal function in astrocytes. Proc. Natl. Acad. Sci. USA 2023, 120, e2220551120. [Google Scholar] [CrossRef] [PubMed]
- Maronde, E.; Schomerus, C.; Stehle, J.H.; Korf, H.W. Control of CREB phosphorylation and its role for induction of melatonin synthesis in rat pinealocytes. Biol. Cell 1997, 89, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Rosenwasser, A.M.; Turek, F.W. Neurobiology of Circadian Rhythm Regulation. Sleep Med. Clin. 2015, 10, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Bering, T.; Gadgaard, C.; Vorum, H.; Honoré, B.; Rath, M.F. Diurnal proteome profile of the mouse cerebral cortex: Conditional deletion of the Bmal1 circadian clock gene elevates astrocyte protein levels and cell abundance in the neocortex and hippocampus. Glia 2023, 71, 2623–2641. [Google Scholar] [CrossRef] [PubMed]
- Won, E.; Na, K.S.; Kim, Y.K. Associations between Melatonin, Neuroinflammation, and Brain Alterations in Depression. Int. J. Mol. Sci. 2021, 23, 305. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Meng, F.; Wang, W.; Wu, M.; Zhang, Y.; Cui, M.; Qiu, C.; Hu, F.; Zhao, D.; Wang, D.; et al. Medial prefrontal cortical PPM1F alters depression-related behaviors by modifying p300 activity via the AMPK signaling pathway. CNS Neurosci. Ther. 2023, 29, 3624–3643. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Analysis | Cluster Analysis | KEGG Pathway Analysis |
|---|---|---|
| Enzyme activity | Transcription regulation | Ubiquitination |
| Differentiation | Differentiation | Estrogen signaling |
| Transcription | Mitochondria | Ferroptosis |
| Apoptosis | Ubiquitination | GnRH secretion |
| Ubiquitination | Lipid binding | Toll-like receptors |
| Fatty acid metabolism | Proteosome complex |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Servín-Muñoz, I.V.; Ortuño-Sahagún, D.; Reyes-Mata, M.P.; Griñán-Ferré, C.; Pallàs, M.; González-Castillo, C. Gene Expression Profile of the Cerebral Cortex of Niemann-Pick Disease Type C Mutant Mice. Genes 2025, 16, 865. https://doi.org/10.3390/genes16080865
Servín-Muñoz IV, Ortuño-Sahagún D, Reyes-Mata MP, Griñán-Ferré C, Pallàs M, González-Castillo C. Gene Expression Profile of the Cerebral Cortex of Niemann-Pick Disease Type C Mutant Mice. Genes. 2025; 16(8):865. https://doi.org/10.3390/genes16080865
Chicago/Turabian StyleServín-Muñoz, Iris Valeria, Daniel Ortuño-Sahagún, María Paulina Reyes-Mata, Christian Griñán-Ferré, Mercè Pallàs, and Celia González-Castillo. 2025. "Gene Expression Profile of the Cerebral Cortex of Niemann-Pick Disease Type C Mutant Mice" Genes 16, no. 8: 865. https://doi.org/10.3390/genes16080865
APA StyleServín-Muñoz, I. V., Ortuño-Sahagún, D., Reyes-Mata, M. P., Griñán-Ferré, C., Pallàs, M., & González-Castillo, C. (2025). Gene Expression Profile of the Cerebral Cortex of Niemann-Pick Disease Type C Mutant Mice. Genes, 16(8), 865. https://doi.org/10.3390/genes16080865