
Genome-Wide Identification of the LEA Gene Family in Myricaria laxiflora and Its Responses to Abiotic Stress


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experiment Design
2.2. Search and Identification of LEA Gene Sequence in M. laxiflora
2.3. Nomenclature and Phylogenetic Tree Analysis of LEA Family Genes in M. laxiflora
2.4. Conserved Domain and Gene Structure Analysis and Cis-Element Analysis of LEA Gene in M. laxiflora
2.5. Chromosomal Mapping, Gene Duplication, and Collinearity Analysis
2.6. Differential Expression Analysis of LEA Gene Family in Myricaria laxiflora
2.7. Statistical Analysis
3. Results
3.1. Identification of LEA Gene in M. laxiflora
3.2. Analysis of Structural Domains and Physicochemical Properties of LEA Gene Family Members in M. laxiflora
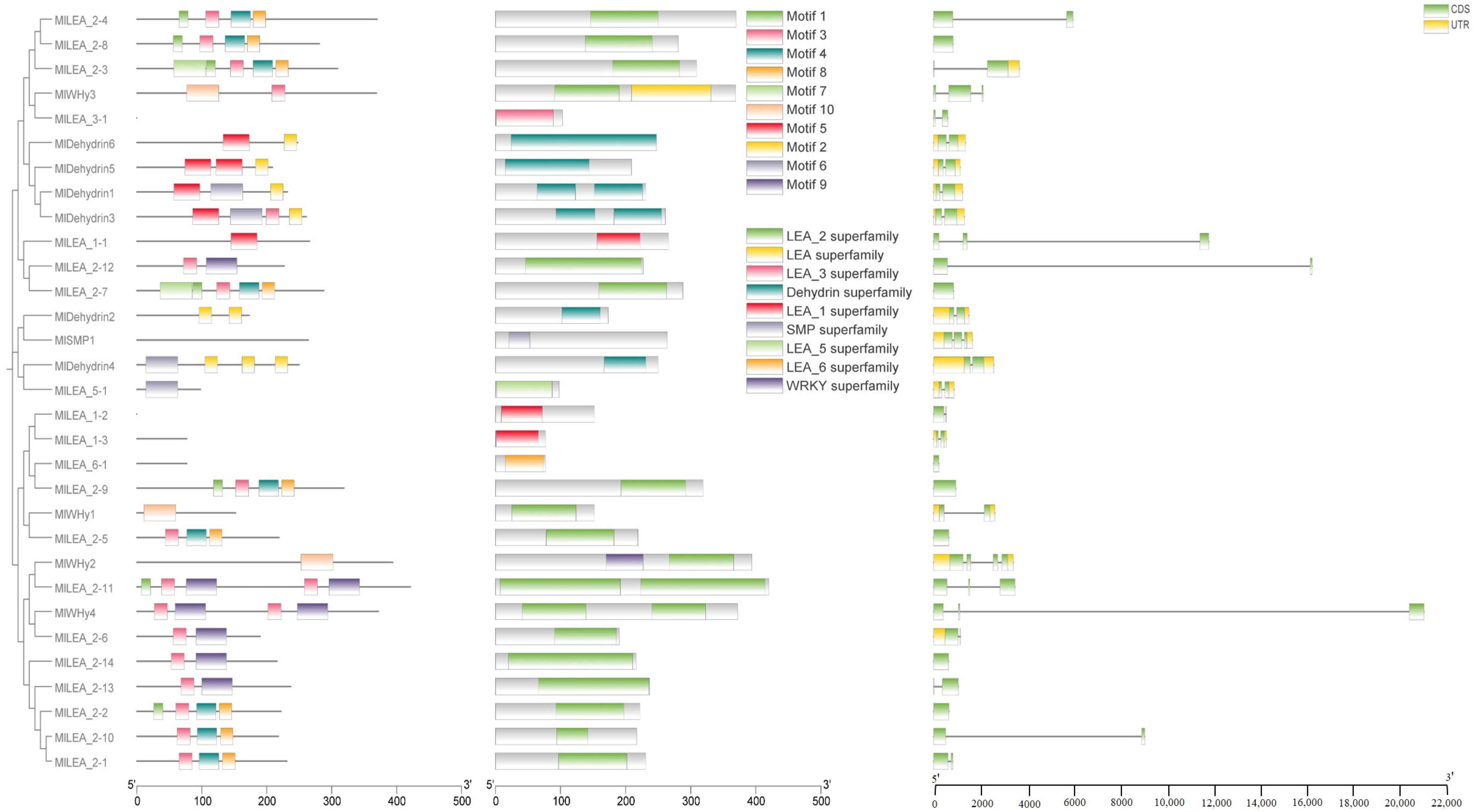
3.3. Phylogenetic Tree Analysis of LEA Gene Family Members in M. laxiflora
3.4. The Domain, Motif Distribution Pattern, and Gene Structure of LEA Gene Family Members in M. laxiflora
3.5. The Cis-Acting Elements of LEA Gene in M. laxiflora
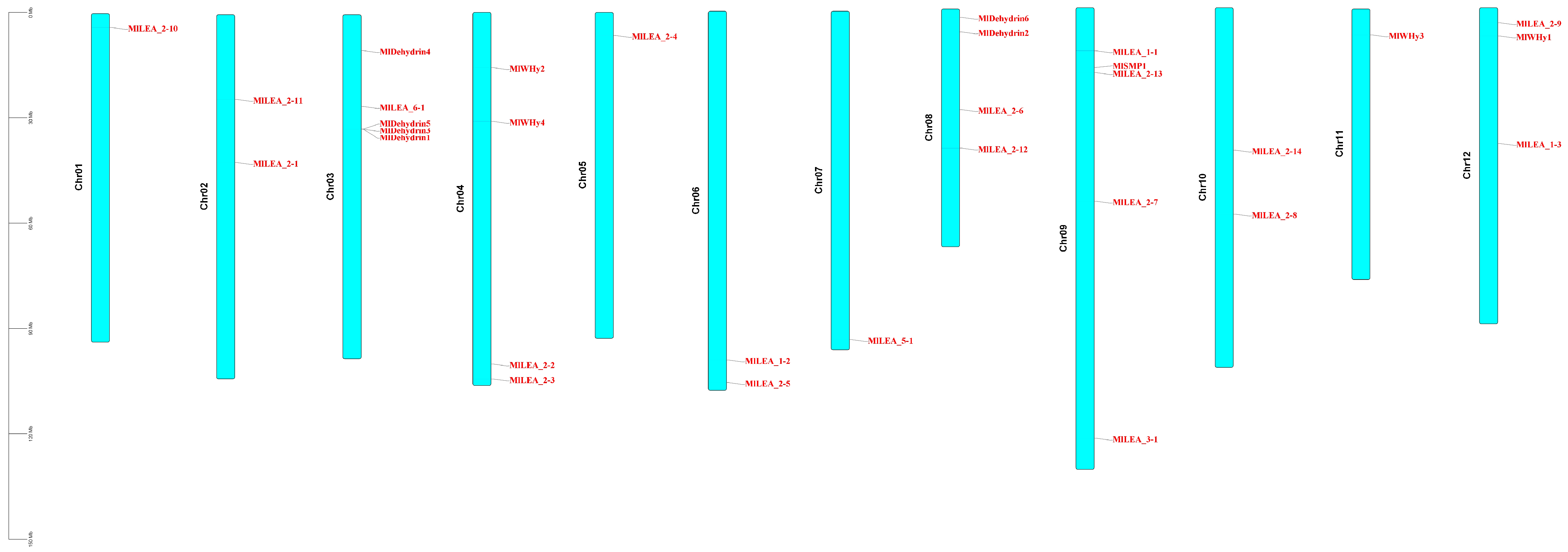
3.6. Chromosomal Distribution and Synteny Analysis of LEA Genes
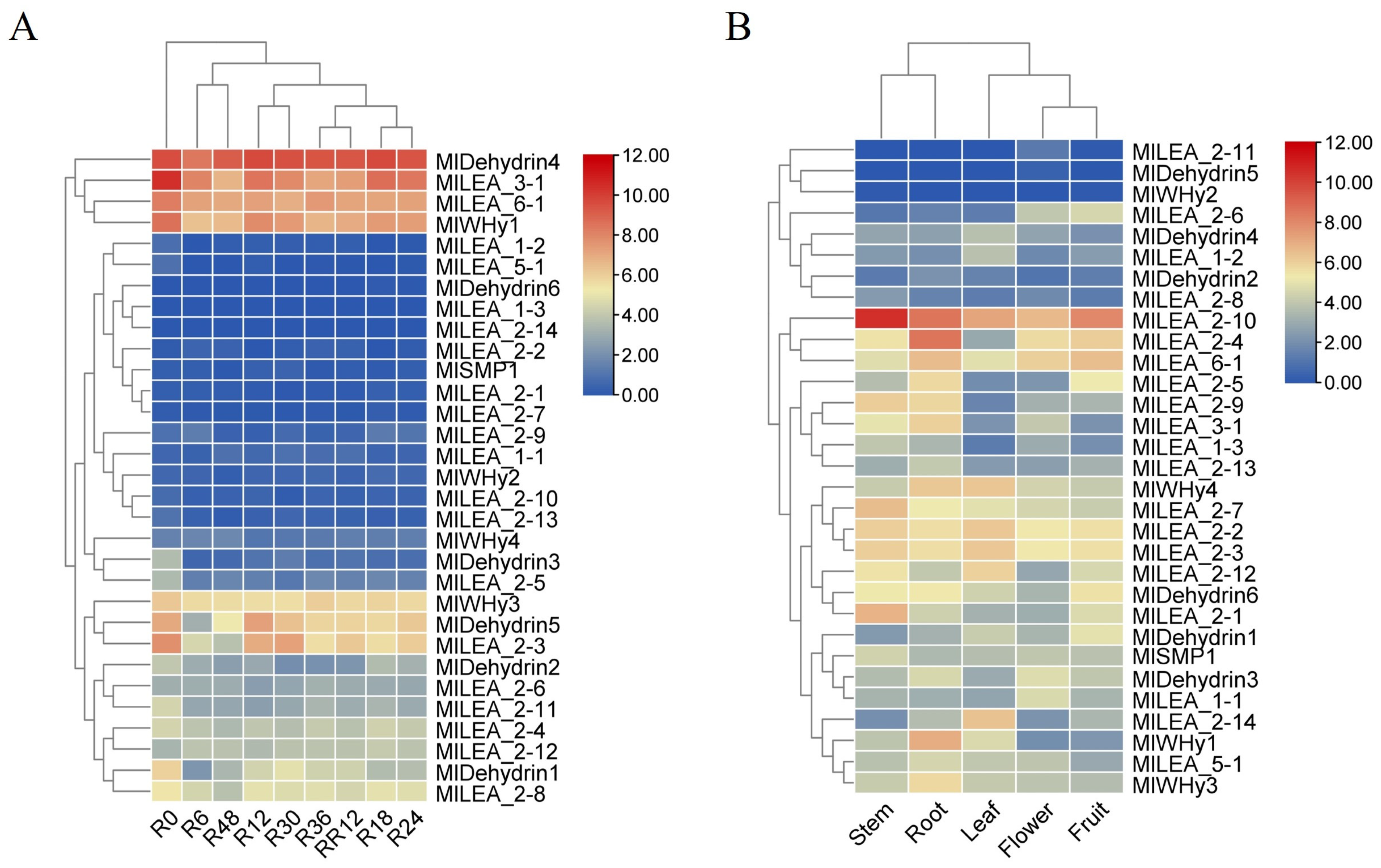
3.7. Expression Profiling of M. laxiflora LEA Genes with RNA-seq
4. Discussion
4.1. The Significant Expansion and Adaptive Evolution of LEA Gene Family
4.2. Diversified Regulation of Promoter Cis-Acting Elements
4.3. Relationship Between Expression Pattern and Flooding Stress Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tian, W.; Bi, Y.H.; Zeng, W.; Jiang, W.; Xue, Y.H.; Wang, G.X.; Liu, S.P. Diversity of Endophytic Fungi of Myricaria laxiflora Grown Under Pre- and Post-Flooding Conditions. Genet. Mol. Res. 2015, 14, 10849–10862. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wang, N.; Zhou, J.; Zhao, Z.; Lv, K.; Huang, Y.; Huang, G.; Qiu, L. Summer Dormancy of Myricaria laxiflora to Escape Flooding Stress: Changes in Phytohormones and Enzymes Induced by Environmental Factors. Plant Physiol. Biochem. 2022, 193, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Huang, H. High Interpopulation Genetic Differentiation and Unidirectional Linear Migration Patterns in Myricaria laxiflora (Tamaricaceae), an Endemic Riparian Plant in the Three Gorges Valley of the Yangtze River. Am. J. Bot. 2006, 93, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, D.; Zhen, Q.; Zhang, J.; Qiu, L.; Huang, G.; Yang, C. Morphological Structures and Histochemistry of Roots and Shoots in Myricaria laxiflora (Tamaricaceae). Open Life Sci. 2021, 16, 455–463. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, Y.-B.; Wang, J.-C.; Tian, M.-Y.; Yuan, X.-H.; Yang, Z.-J.; Zuo, Y.-W.; Deng, H.-P. Transcriptome Proffling, Physiological and Biochemical Analyses Reveal Comprehensive Insights into Cadmium Stress in Myricaria laxiflora. Plants 2024, 13, 3433. [Google Scholar] [CrossRef]
- Xie, Y. Progress and Application of IUCN Red List of Threatened Species. Biodivers. Sci. 2022, 30, 22445. [Google Scholar] [CrossRef]
- Li, L.; Huang, G.; Xiang, W.; Zhu, H.; Zhang, H.; Zhang, J.; Ding, Z.; Liu, J.; Wu, D. Integrated Transcriptomic and Proteomic Analyses Uncover the Regulatory Mechanisms of Myricaria laxiflora Under Flooding Stress. Front. Plant Sci. 2022, 13, 924490. [Google Scholar] [CrossRef]
- Li, L.; Su, Y.; Xiang, W.; Huang, G.; Liang, Q.; Dun, B.; Zhang, H.; Xiao, Z.; Qiu, L.; Zhang, J.; et al. Transcriptomic Network Underlying Physiological Alterations in the Stem of Myricaria laxiflora in Response to Waterlogging Stress. Ecotoxicol. Environ. Safety 2024, 284, 116991. [Google Scholar] [CrossRef]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The Enigmatic LEA Proteins and Other Hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef]
- Candat, A.; Paszkiewicz, G.; Neveu, M.; Gautier, R.; Logan, D.C.; Avelange-Macherel, M.-H.; Macherel, D. The Ubiquitous Distribution of Late Embryogenesis Abundant Proteins Across Cell Compartments in Arabidopsis Offers Tailored Protection Against Abiotic Stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) Proteins and Their Encoding Genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef]
- Jia, C.; Guo, B.; Wang, B.; Li, X.; Yang, T.; Li, N.; Wang, J.; Yu, Q. The LEA Gene Family in Tomato and Its Wild Relatives: Genome-Wide Identification, Structural Characterization, Expression Profiling, and Role of SlLEA6 in Drought Stress. BMC Plant Biol. 2022, 22, 596. [Google Scholar] [CrossRef]
- Wang, Q.; Lei, X.; Wang, Y.; Di, P.; Meng, X.; Peng, W.; Rong, J.; Wang, Y. Genome-Wide Identification of the LEA Gene Family in Panax Ginseng: Evidence for the Role of PgLEA2-50 in Plant Abiotic Stress Response. Plant Physiol. Biochem. 2024, 212, 108742. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data Without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant Responses to Drought, Salinity and Extreme Temperatures: Towards Genetic Engineering for Stress Tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Cao, D.; Wang, Z.; Ma, L.; Tian, K.; Liu, Y.; Gong, Z.; Zhu, X.; Jiang, C.; Li, Y. Genome-Wide Identification and Expression Analyses of the LEA Protein Gene Family in Tea Plant Reveal Their Involvement in Seed Development and Abiotic Stress Responses. Sci. Rep. 2019, 9, 14123. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, S.; Chen, W.; Zhang, J.; Zhang, L.; Sun, W.; Wang, Z. Plant Dehydrins: Expression, Regulatory Networks, and Protective Roles in Plants Challenged by Abiotic Stress. Int. J. Mol. Sci. 2021, 22, 12619. [Google Scholar] [CrossRef] [PubMed]
- Mertens, J.; Aliyu, H.; Cowan, D.A. LEA Proteins and the Evolution of the WHy Domain. Appl. Environ. Microbiol. 2018, 84, e00539-18. [Google Scholar] [CrossRef] [PubMed]
- Abdul Aziz, M.; Sabeem, M.; Mullath, S.K.; Brini, F.; Masmoudi, K. Plant Group II LEA Proteins: Intrinsically Disordered Structure for Multiple Functions in Response to Environmental Stresses. Biomolecules 2021, 11, 1662. [Google Scholar] [CrossRef]
- Ghanmi, S.; Graether, S.P.; Hanin, M. The Halophyte Dehydrin Sequence Landscape. Biomolecules 2022, 12, 330. [Google Scholar] [CrossRef] [PubMed]
- Bies-Ethève, N.; Gaubier-Comella, P.; Debures, A.; Lasserre, E.; Jobet, E.; Raynal, M.; Cooke, R.; Delseny, M. Inventory, Evolution and Expression Profiling Diversity of the LEA (Late Embryogenesis Abundant) Protein Gene Family in Arabidopsis thaliana. Plant Mol. Biol. 2008, 67, 107–124. [Google Scholar] [CrossRef]
- Danquah, A.; de Zelicourt, A.; Colcombet, J.; Hirt, H. The Role of ABA and MAPK Signaling Pathways in Plant Abiotic Stress Responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The Roles of Methyl Jasmonate to Stress in Plants. Funct. Plant Biol. 2019, 46, 197–212. [Google Scholar] [CrossRef]
- Soliman, S.; Wang, Y.; Han, Z.; Pervaiz, T.; El-Kereamy, A. Strigolactones in Plants and Their Interaction with the Ecological Microbiome in Response to Abiotic Stress. Plants 2022, 11, 3499. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.C.J.; Perata, P.; van Dongen, J.T. Oxygen Sensing in Plants Is Mediated by an N-End Rule Pathway for Protein Destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The Role and Regulation of ABI5 (ABA-Insensitive 5) in Plant Development, Abiotic Stress Responses and Phytohormone Crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef]
- Zamora-Briseño, J.A.; de Jiménez, E.S. A LEA 4 Protein Up-Regulated by ABA Is Involved in Drought Response in Maize Roots. Mol. Biol. Rep. 2016, 43, 221–228. [Google Scholar] [CrossRef]
- Král, D.; Šenkyřík, J.B.; Ondřej, V. Expression of Genes Involved in ABA and Auxin Metabolism and LEA Gene During Embryogenesis in Hemp. Plants 2022, 11, 2995. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant Hormone-Mediated Regulation of Stress Responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Nguyen, C.T.T.; Jung, C.; Cheong, J.-J. AtMYB44 Suppresses Transcription of the Late Embryogenesis Abundant Protein Gene AtLEA4-5. Biochem. Biophys. Res. Commun. 2019, 511, 931–934. [Google Scholar] [CrossRef]
- Xiao, B.; Huang, Y.; Tang, N.; Xiong, L. Over-Expression of a LEA Gene in Rice Improves Drought Resistance Under the Field Conditions. Theor. Appl. Genet. 2007, 115, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, Q.; Qin, J.; Xiao, G.; Zhu, S.; Hu, T. OsLEA1a Overexpression Enhances Tolerance to Diverse Abiotic Stresses by Inhibiting Cell Membrane Damage and Enhancing ROS Scavenging Capacity in Transgenic Rice. Funct. Plant Biol. 2021, 48, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, D.; Agoston, B.; Tompa, P. Disordered Plant LEA Proteins as Molecular Chaperones. Plant Signal. Behav. 2008, 3, 710–713. [Google Scholar] [CrossRef]
- Chakrabortee, S.; Tripathi, R.; Watson, M.; Schierle, G.S.K.; Kurniawan, D.P.; Kaminski, C.F.; Wise, M.J.; Tunnacliffe, A. Intrinsically Disordered Proteins as Molecular Shields. Mol. Biosyst. 2012, 8, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Romero-Pérez, P.S.; Martínez-Castro, L.V.; Linares, A.; Arroyo-Mosso, I.; Sánchez-Puig, N.; Cuevas-Velazquez, C.L.; Sukenik, S.; Guerrero, A.; Covarrubias, A.A. Self-Association and Multimer Formation in AtLEA4-5, a Desiccation-Induced Intrinsically Disordered Protein from Plants. Protein Sci. 2024, 33, e5192. [Google Scholar] [CrossRef]
- Artur, M.A.S.; Rienstra, J.; Dennis, T.J.; Farrant, J.M.; Ligterink, W.; Hilhorst, H. Structural Plasticity of Intrinsically Disordered LEA Proteins from Xerophyta Schlechteri Provides Protection In Vitro and In Vivo. Front. Plant Sci. 2019, 10, 1272. [Google Scholar] [CrossRef]
- Hu, M.; Li, Z.; Lin, X.; Tang, B.; Xing, M.; Zhu, H. Comparative Analysis of the LEA Gene Family in Seven Ipomoea Species, Focuses on Sweet Potato (Ipomoea batatas L.). BMC Plant Biol. 2024, 24, 1256. [Google Scholar] [CrossRef]
- Andersson, J.M.; Pham, Q.D.; Mateos, H.; Eriksson, S.; Harryson, P.; Sparr, E. The Plant Dehydrin Lti30 Stabilizes Lipid Lamellar Structures in Varying Hydration Conditions. J. Lipid Res. 2020, 61, 1014–1024. [Google Scholar] [CrossRef]
- Mishra, M.K.; Tiwari, S.; Misra, P. Overexpression of WssgtL3.1 Gene from Withania somnifera Confers Salt Stress Tolerance in Arabidopsis. Plant Cell Rep. 2021, 40, 2191–2204. [Google Scholar] [CrossRef]
- Gao, C.; Wang, C.; Zheng, L.; Wang, L.; Wang, Y. A LEA Gene Regulates Cadmium Tolerance by Mediating Physiological Responses. Int. J. Mol. Sci. 2012, 13, 5468–5481. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Domain Name | Domain ID | Start | End | Length | E-Value |
|---|---|---|---|---|---|---|
| MlDehydrin1 | Dehydrin | PF00257 | 64 | 123 | 60 | 1.56 × 10−7 |
| Dehydrin | PF00257 | 152 | 226 | 75 | 1.26 × 10−6 | |
| MlDehydrin2 | Dehydrin | PF00257 | 102 | 161 | 60 | 1.91 × 10−9 |
| MlDehydrin3 | Dehydrin | PF00257 | 93 | 153 | 61 | 2.78 × 10−7 |
| Dehydrin | PF00257 | 182 | 255 | 74 | 1.92 × 10−3 | |
| MlDehydrin4 | Dehydrin | PF00257 | 167 | 231 | 65 | 3.85 × 10−7 |
| MlDehydrin5 | Dehydrin | PF00257 | 15 | 144 | 130 | 1.39 × 10−12 |
| MlDehydrin6 | Dehydrin | PF00257 | 24 | 247 | 224 | 1.42 × 10−9 |
| MlLEA_1-1 | lLEA_1 | PF03760 | 156 | 222 | 67 | 5.65 × 10−21 |
| MlLEA_1-2 | lLEA_1 | PF03760 | 9 | 72 | 64 | 2.67 × 10−12 |
| MlWHy1 | WHy | PF08267 | 25 | 124 | 100 | 3.67 × 10−22 |
| MlWHy2 | WHy | PF08267 | 267 | 366 | 100 | 5.79 × 10−24 |
| MlWHy3 | WHy | PF08267 | 91 | 190 | 100 | 7.51 × 10−28 |
| WHy | PF08267 | 209 | 331 | 123 | 5.98 × 10−21 | |
| MlLEA_3-1 | LEA_3 | PF03242 | 1 | 89 | 89 | 1.16 × 10−30 |
| MlLEA_5-1 | LEA_5 | PF00477 | 2 | 87 | 86 | 2.61 × 10−26 |
| MlLEA_6-1 | LEA_6 | PF10714 | 15 | 74 | 60 | 3.53 × 10−21 |
| MlSMP1 | SMP | PF04927 | 21 | 53 | 33 | 7.80 × 10−4 |
| MlLEA_1-3 | lLEA_1 | PF03760 | 1 | 66 | 66 | 3.60 × 10−5 |
| MlLEA_2-10 | LEA_2 Superfamily | PF03168 | 94 | 142 | 49 | 9.28 × 10−3 |
| MlLEA_2-11 | LEA_2 Superfamily | PF03168 | 7 | 192 | 186 | 2.18 × 10−39 |
| LEA_2 Superfamily | PF03168 | 223 | 414 | 192 | 2.77 × 10−24 | |
| MlLEA_2-1 | LEA_2 | PF03168 | 97 | 202 | 106 | 5.73 × 10−5 |
| MlWHy4 | WHy | PF08267 | 240 | 323 | 84 | 6.90 × 10−16 |
| WHy | PF08267 | 41 | 139 | 99 | 1.55 × 10−15 | |
| MlLEA_2-2 | LEA_2 | PF03168 | 93 | 197 | 105 | 1.27 × 10−3 |
| MlLEA_2-3 | LEA_2 | PF03168 | 180 | 283 | 104 | 4.04 × 10−7 |
| MlLEA_2-4 | LEA_2 | PF03168 | 146 | 250 | 105 | 2.45 × 10−8 |
| MlLEA_2-5 | LEA_2 | PF03168 | 78 | 182 | 105 | 2.22 × 10−8 |
| MlLEA_2-6 | LEA_2 | PF03168 | 91 | 186 | 96 | 2.71 × 10−8 |
| MlLEA_2-12 | LEA_2 super family | PF03168 | 46 | 224 | 179 | 8.20 × 10−12 |
| MlLEA_2-13 | LEA_2 super family | PF03168 | 66 | 236 | 171 | 1.09 × 10−7 |
| MlLEA_2-7 | LEA_2 | PF03168 | 159 | 263 | 105 | 2.09 × 10−3 |
| MlLEA_2-14 | LEA_2 super family | PF03168 | 20 | 211 | 192 | 2.75 × 10−22 |
| MlLEA_2-8 | LEA_2 | PF03168 | 138 | 241 | 104 | 5.22 × 10−7 |
| MlLEA_2-9 | LEA_2 | PF03168 | 193 | 292 | 100 | 6.14 × 10−3 |
| Gene Name | Gene ID | Subfamily | Subcellular Location | CDS Size | Protein Size | MW/KDa | pI | Instability Index | Aliphatic Index | GRAVY |
|---|---|---|---|---|---|---|---|---|---|---|
| MlDehydrin1 | Myl03g00992.1 | Dehydrin | Chloroplast | 696 | 231 | 24.87 | 9.54 | 25.44 | 37.6 | −1.392 |
| MlDehydrin2 | Myl08g00301.1 | Dehydrin | Chloroplast | 519 | 172 | 19.1 | 6.46 | 55.21 | 42.6 | −1.369 |
| MlDehydrin3 | Myl03g00991.1 | Dehydrin | Chloroplast | 783 | 260 | 27.71 | 9.43 | 25.9 | 36.7 | −1.326 |
| MlDehydrin4 | Myl03g00338.1 | Dehydrin | Chloroplast | 750 | 249 | 27.82 | 5.47 | 55.85 | 49.9 | −1.378 |
| MlDehydrin5 | Myl03g00989.1 | Dehydrin | Chloroplast | 627 | 208 | 22.14 | 6.71 | 38.97 | 30.4 | −1.378 |
| MlDehydrin6 | Myl08g00106.1 | Dehydrin | Chloroplast | 744 | 247 | 26.53 | 6.68 | 29.65 | 51.1 | −1.002 |
| MlLEA_1-1 | Myl09g00459.1 | LEA_1 | Chloroplast | 798 | 265 | 29.02 | 9.52 | 46.03 | 70 | −0.579 |
| MlLEA_1-2 | Myl06g01621.1 | LEA_1 | Chloroplast | 459 | 152 | 15.76 | 9.13 | 19.26 | 44.5 | −0.774 |
| MlLEA_1-3 | Myl12g00406.1 | LEA_1 | Chloroplast | 231 | 76 | 8.694 | 6.84 | 35.63 | 63.3 | −1.064 |
| MlLEA_2-1 | Myl02g01162.1 | LEA_2 | Chloroplast | 693 | 230 | 26.31 | 9.56 | 57.55 | 84.8 | −0.27 |
| MlLEA_2-2 | Myl04g01868.1 | LEA_2 | Chloroplast | 666 | 221 | 25.31 | 9.35 | 36.44 | 91.7 | −0.196 |
| MlLEA_2-3 | Myl04g02032.1 | LEA_2 | Chloroplast | 927 | 308 | 34 | 9.18 | 42.85 | 85 | −0.207 |
| MlLEA_2-4 | Myl05g00215.1 | LEA_2 | Chloroplast | 1110 | 369 | 40.42 | 10.25 | 48.67 | 87.7 | −0.146 |
| MlLEA_2-5 | Myl06g01866.1 | LEA_2 | Chloroplast | 657 | 218 | 24 | 9.41 | 32.09 | 95.2 | −0.008 |
| MlLEA_2-6 | Myl08g01177.1 | LEA_2 | Chloroplast | 570 | 189 | 21.01 | 9.55 | 50.03 | 114 | 0.138 |
| MlLEA_2-7 | Myl09g01907.1 | LEA_2 | Chloroplast | 864 | 287 | 31.94 | 9.32 | 44.02 | 83.8 | −0.249 |
| MlLEA_2-8 | Myl10g02084.1 | LEA_2 | Chloroplast | 843 | 280 | 30.56 | 10.09 | 41.04 | 80.1 | −0.147 |
| MlLEA_2-9 | Myl12g00097.1 | LEA_2 | Chloroplast | 957 | 318 | 35.7 | 8.5 | 61.28 | 81.5 | −0.297 |
| MlLEA_3-1 | Myl09g02902.1 | LEA_3 | Chloroplast | 309 | 102 | 11.19 | 7.93 | 43.05 | 69.8 | −0.46 |
| MlLEA_5-1 | Myl07g01997.1 | LEA_5 | Chloroplast | 294 | 97 | 10.46 | 8.12 | 63.95 | 45.3 | −1.31 |
| MlLEA_6-1 | Myl03g00855.1 | LEA_6 | Chloroplast | 231 | 76 | 8.08 | 5.58 | 56.29 | 28.3 | −378 |
| MlWHy1 | Myl12g00170.1 | WHy | Chloroplast | 456 | 151 | 16.37 | 4.68 | 16.03 | 108 | 0.148 |
| MlWHy2 | Myl04g00578.1 | WHy | Chloroplast | 1182 | 393 | 43.57 | 5.84 | 38.55 | 76.3 | −0.475 |
| MlWHy3 | Myl11g00251.1 | WHy | Chloroplast | 1107 | 368 | 40.96 | 5.13 | 20.59 | 99.3 | −0.22 |
| MlWHy4 | Myl04g00969.1 | WHy | Chloroplast | 1116 | 371 | 40.79 | 9.67 | 34.73 | 103 | 0.095 |
| MlSMP1 | Myl09g00630.1 | SMP | Chloroplast | 792 | 263 | 28.72 | 9.81 | 42.05 | 81.6 | −0.327 |
| MlLEA_2-10 | Myl01g00162.1 | LEA_2 superfamily | Chloroplast | 654 | 217 | 25.01 | 9.9 | 47.78 | 99.2 | −0.236 |
| MlLEA_2-11 | Myl02g00806.1 | LEA_2 superfamily | Chloroplast | 1263 | 420 | 45.91 | 9.09 | 22.82 | 115 | 0.248 |
| MlLEA_2-12 | Myl08g01489.1 | LEA_2 superfamily | Chloroplast | 681 | 226 | 25.37 | 8.35 | 47.41 | 94 | −0.078 |
| MlLEA_2-13 | Myl09g00692.1 | LEA_2 superfamily | Chloroplast | 711 | 236 | 26.13 | 9.36 | 43.07 | 75.9 | −0.168 |
| MlLEA_2-14 | Myl10g01528.1 | LEA_2 superfamily | Chloroplast | 648 | 215 | 23.45 | 9.51 | 27.41 | 116 | 0.237 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, D.; Zhang, T.; Li, L.; Zhang, H.; Su, Y.; Wu, J.; Wang, J.; Li, C.; Huang, G. Genome-Wide Identification of the LEA Gene Family in Myricaria laxiflora and Its Responses to Abiotic Stress. Genes 2025, 16, 763. https://doi.org/10.3390/genes16070763
Wu D, Zhang T, Li L, Zhang H, Su Y, Wu J, Wang J, Li C, Huang G. Genome-Wide Identification of the LEA Gene Family in Myricaria laxiflora and Its Responses to Abiotic Stress. Genes. 2025; 16(7):763. https://doi.org/10.3390/genes16070763
Chicago/Turabian StyleWu, Di, Tonghua Zhang, Linbao Li, Haibo Zhang, Yang Su, Jinhua Wu, Junchen Wang, Chunlong Li, and Guiyun Huang. 2025. "Genome-Wide Identification of the LEA Gene Family in Myricaria laxiflora and Its Responses to Abiotic Stress" Genes 16, no. 7: 763. https://doi.org/10.3390/genes16070763
APA StyleWu, D., Zhang, T., Li, L., Zhang, H., Su, Y., Wu, J., Wang, J., Li, C., & Huang, G. (2025). Genome-Wide Identification of the LEA Gene Family in Myricaria laxiflora and Its Responses to Abiotic Stress. Genes, 16(7), 763. https://doi.org/10.3390/genes16070763