
Identification and Bioinformatics Analysis of the HSP20 Family in the Peony

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of the PsHSP20 Family Genes in the Peony
2.2. Analysis of Physicochemical Properties of the PsHSP20 Family Genes
2.3. Secondary and Tertiary Structural Modeling of the Peony PsHSP20 Proteins
2.4. Phylogenetic Analysis of the PsHSP20 Family Genes
2.5. Analysis of the Structure and Domain of the PsHsp20 Genes in Peony
2.6. Synteny Analysis and Chromosomal Localization of the PsHSP20 Family Genes
2.7. Analysis of Cis-Acting Elements and Transcription Factor Binding Sites in Promoter Regions
2.8. Analysis of Expression Patterns
2.9. Plant Materials and Heat Stress Treatment
3. Results
3.1. Genome-Wide Identification and Physicochemical Characterization of the PsHSP20 Gene Family in the Peony
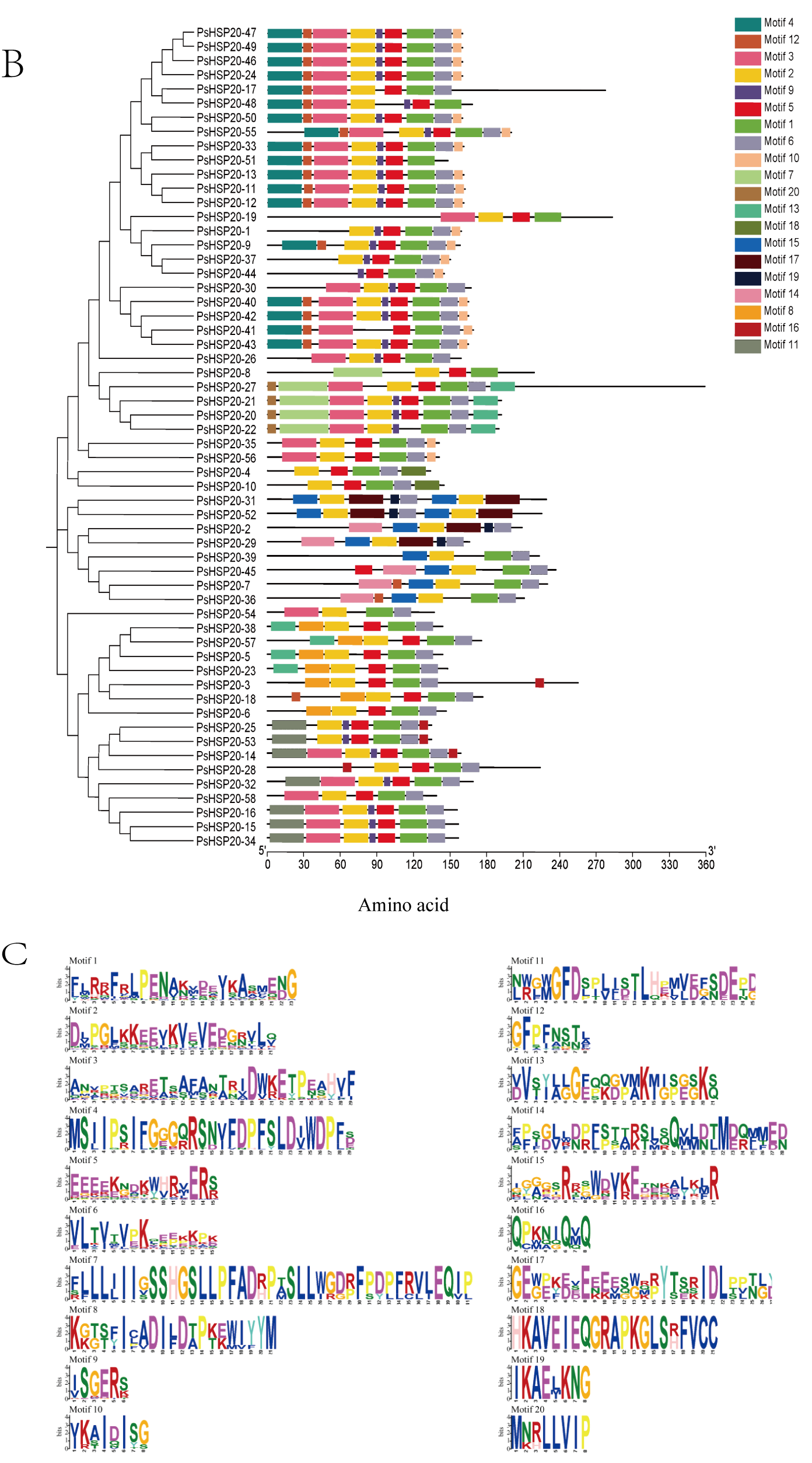
3.2. Structural and Conserved Domain Analysis of the PsHSP20 Genes
3.3. Secondary and Tertiary Structural Features of the PsHSP20 Proteins
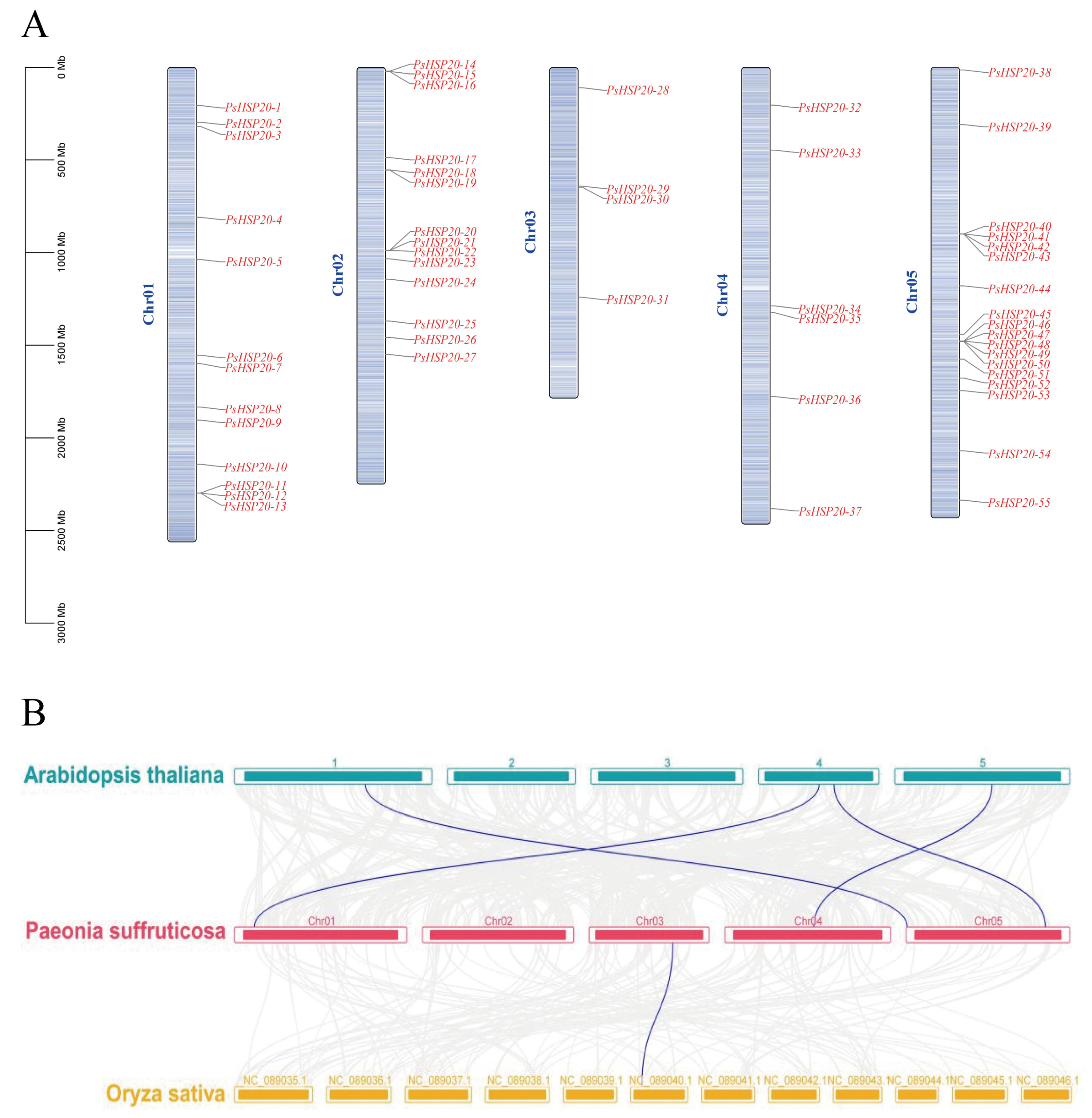
3.4. Chromosomal Localization and Synteny Analysis of the PsHSP20 Genes
3.5. Phylogenetic Classification and Evolutionary Relationships of the PsHSP20 Genes
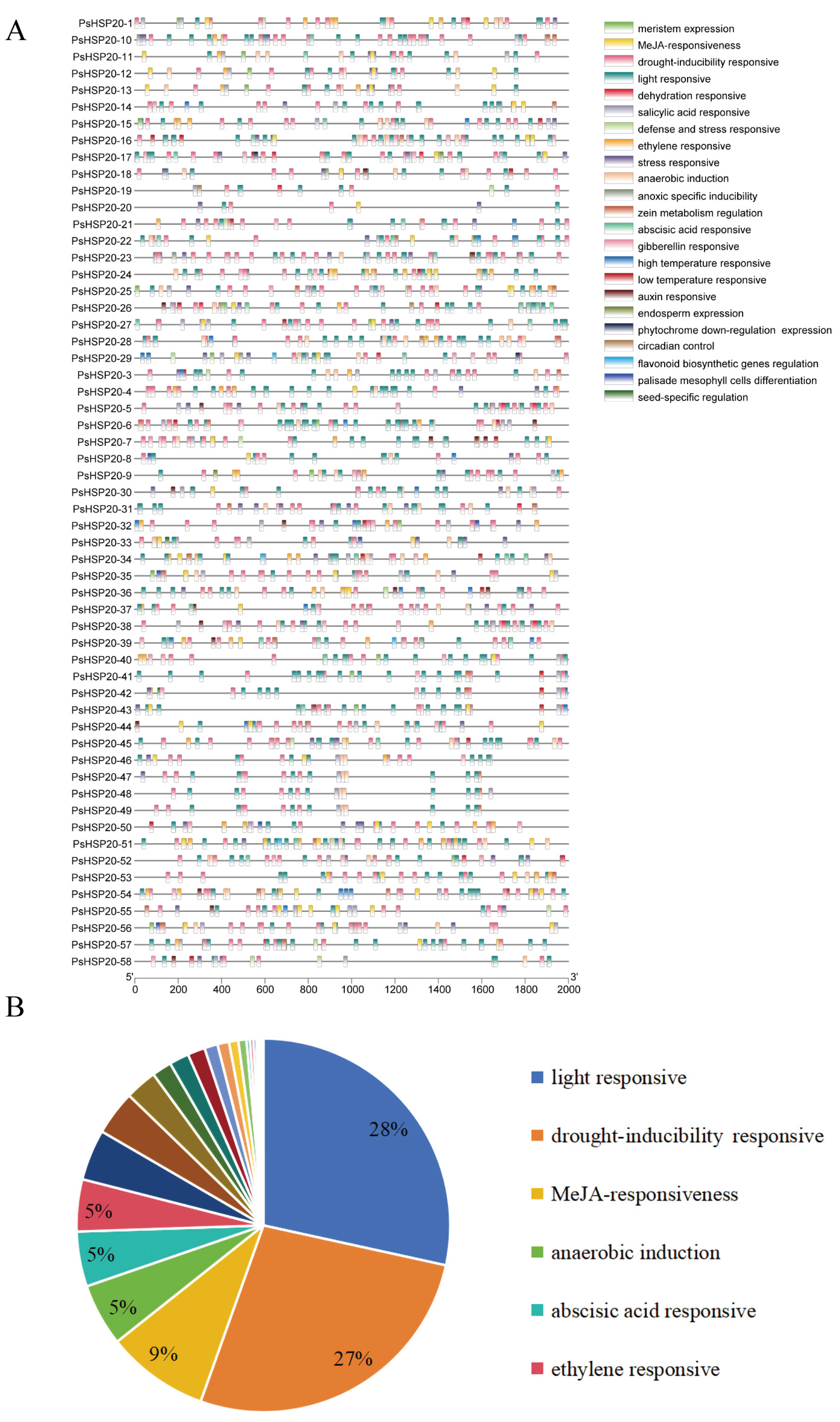
3.6. Promoter Architecture: Cis-Acting Elements and Transcription Factor Binding Sites
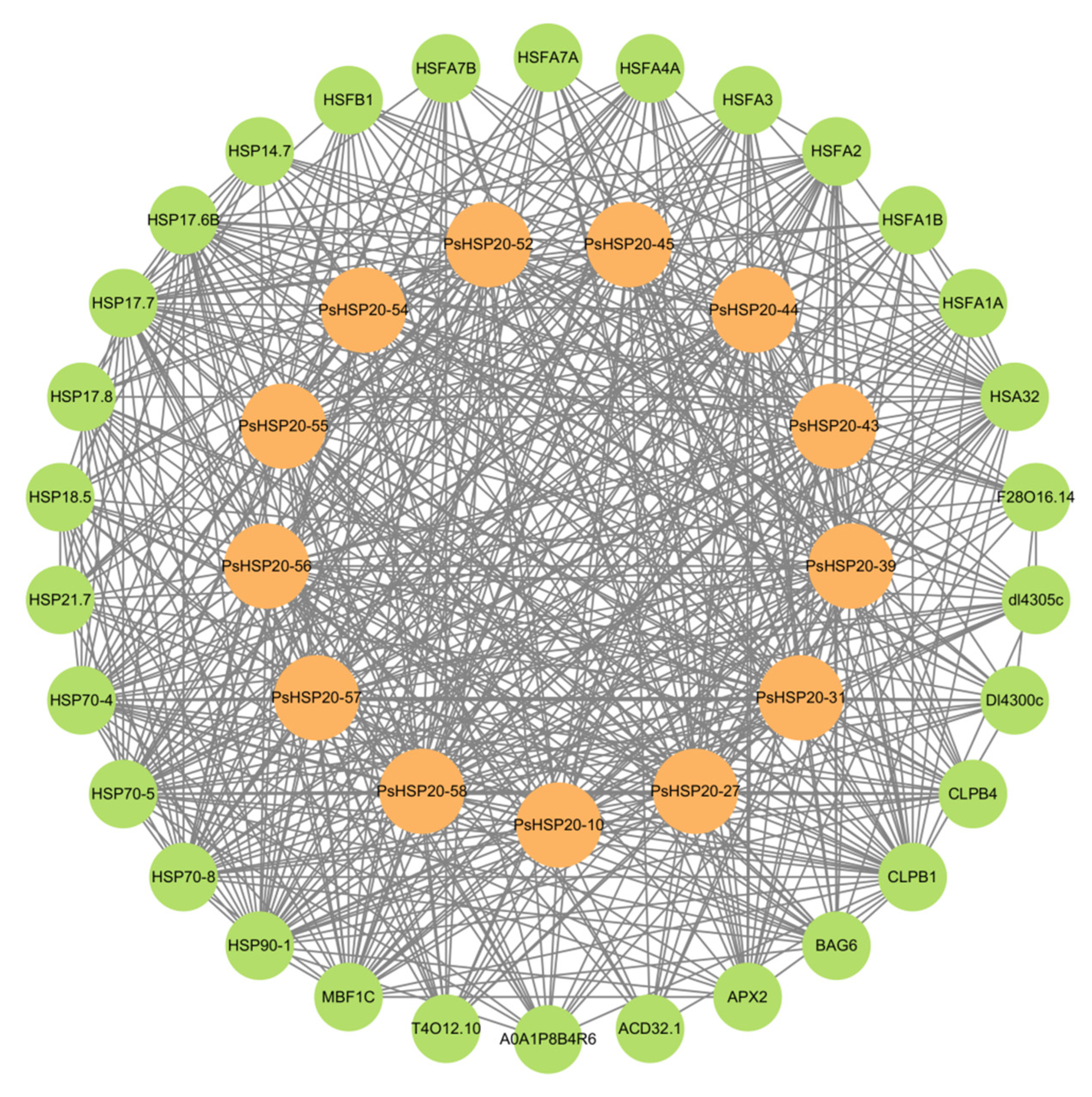
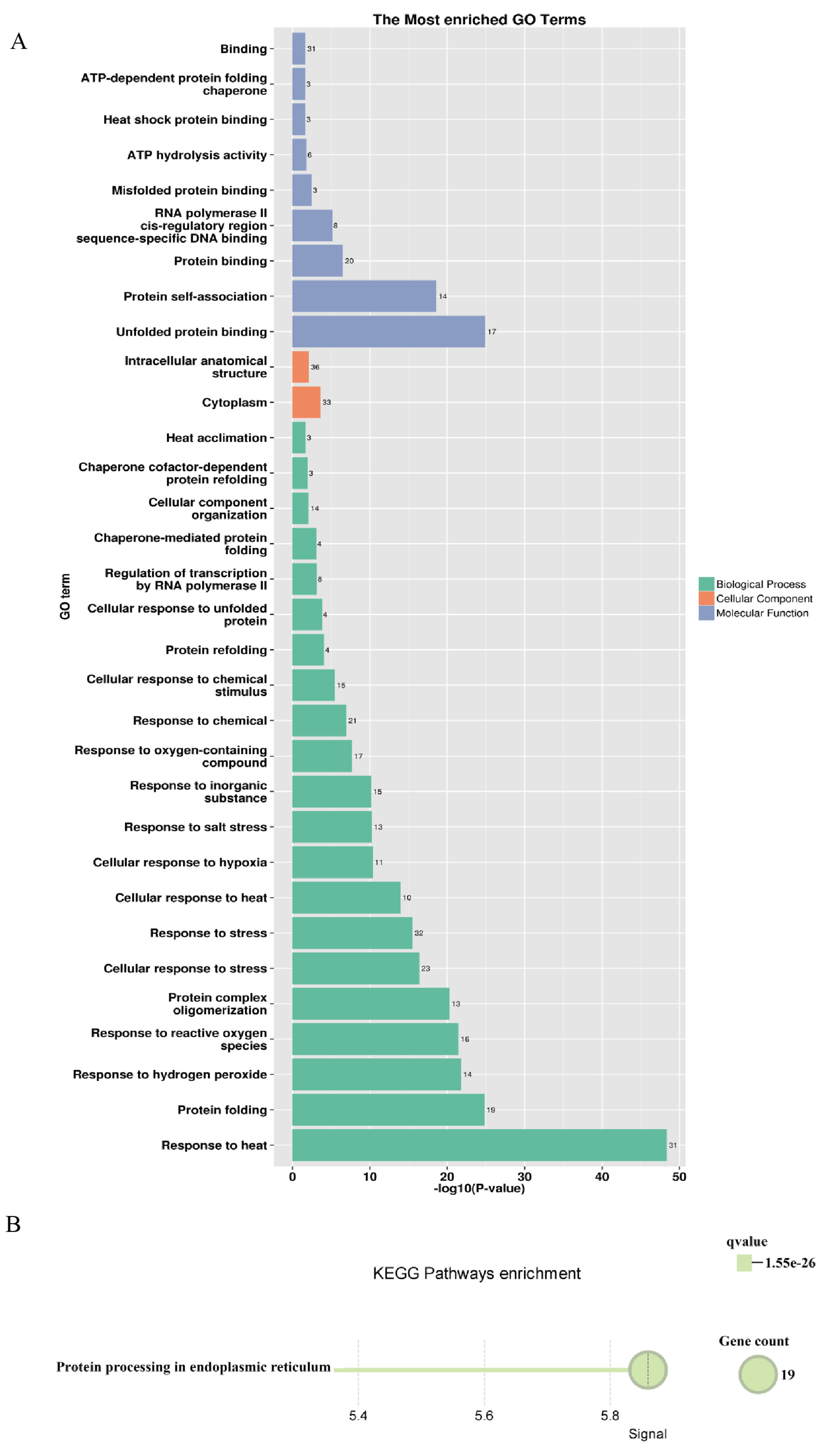
3.7. Regulatory Network and Functional Enrichment of the PsHSP20 Genes
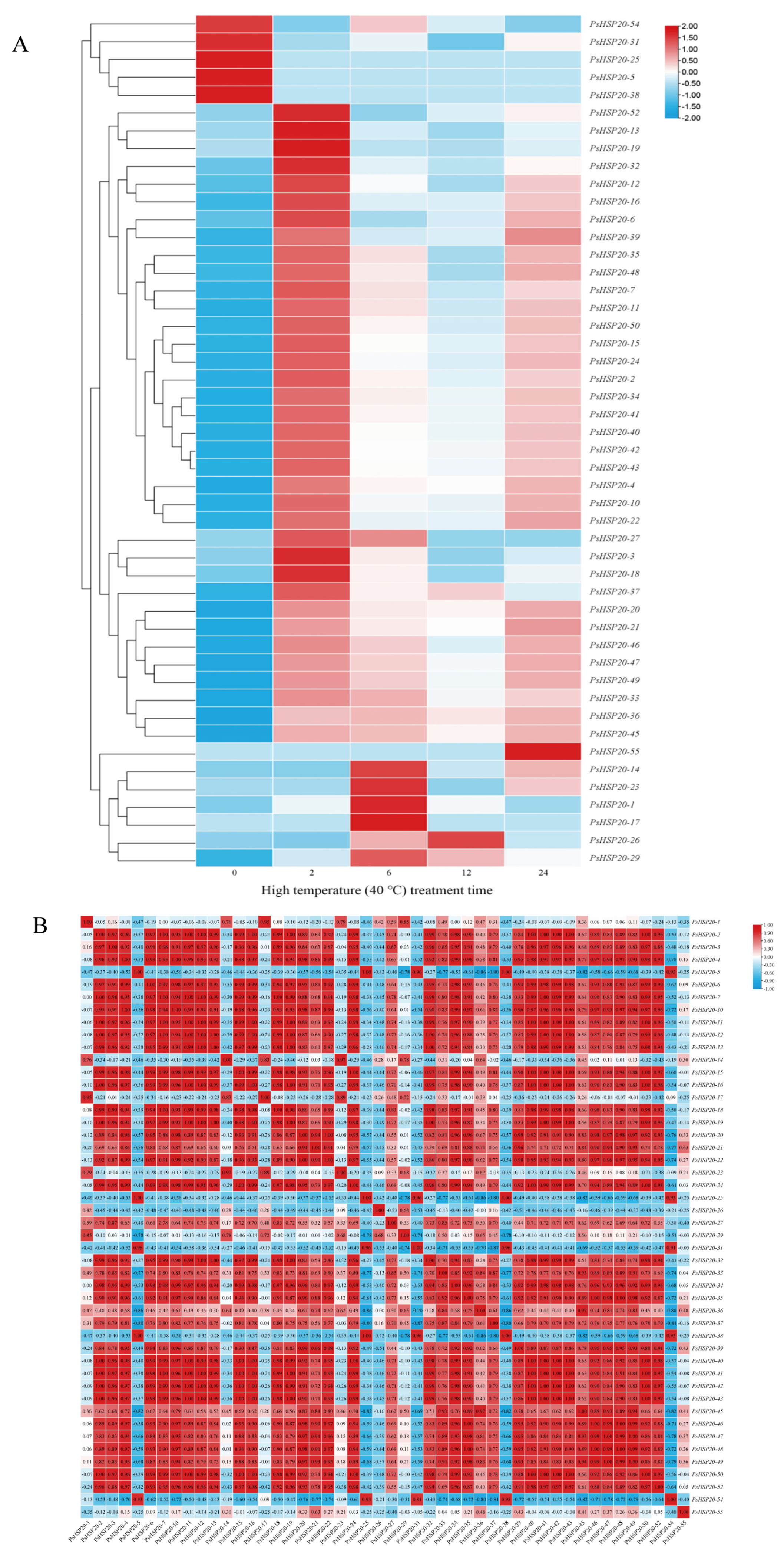
3.8. Dynamic Expression Profiling of the PsHSP20 Genes Under Heat Stress
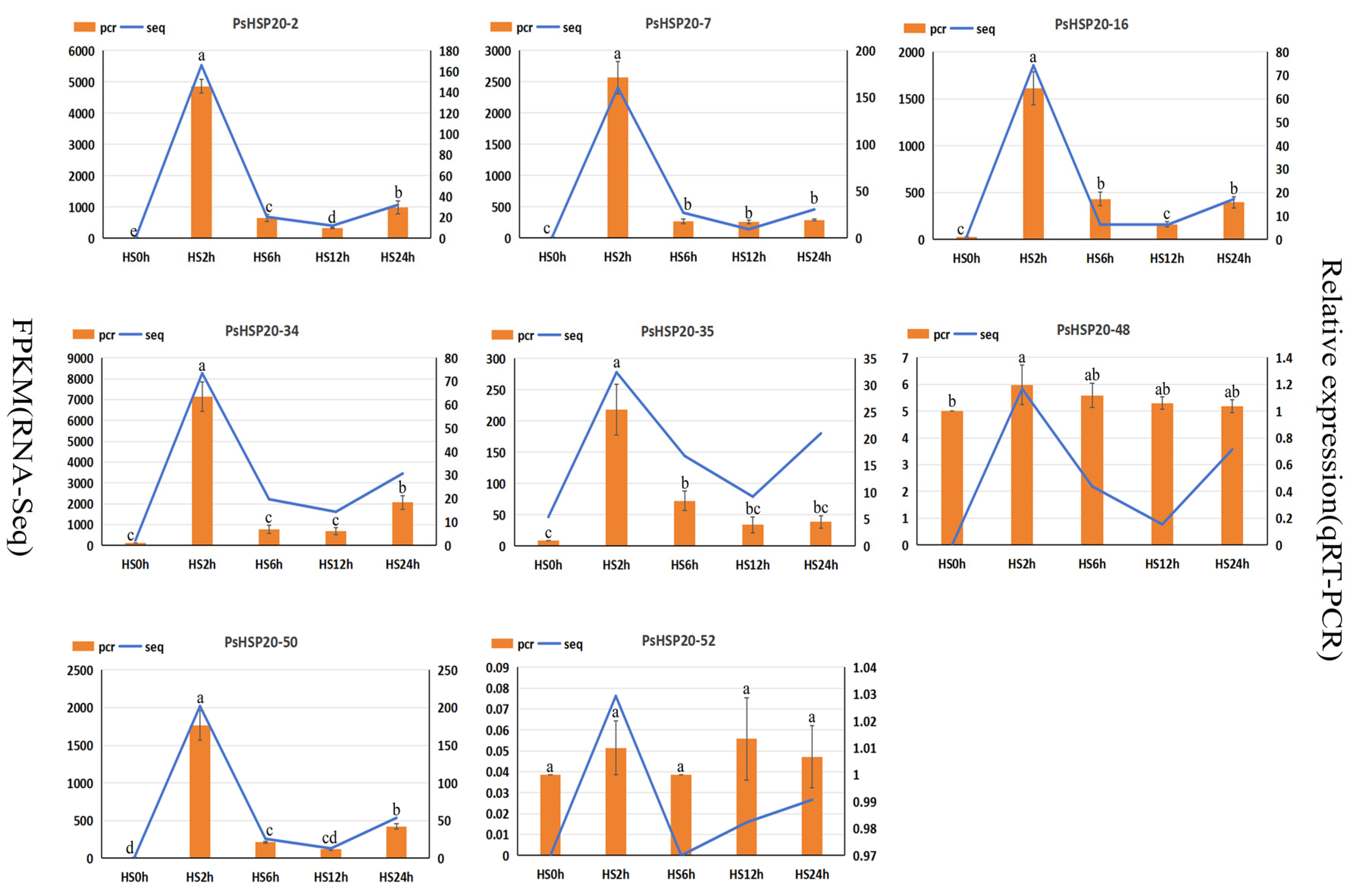
3.9. qRT-PCR Analysis of Target Genes of the PsHSP20 Gene Family
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACD | α-crystallin Domain |
| CDD | Conserved Domain Database |
| CDS | Coding Sequence |
| GO | Gene Ontology |
| HMM | Hidden Markov Model |
| HSE | Heat Shock Elements |
| HSP | Heat Shock Protein |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LCA | Luciferase Complementarity Assays |
| PPI | Protein–Protein Interaction |
| TF | Transcription Factor |
References
- Tong, N.; Shu, Q.; Wang, B.; Peng, L.; Liu, Z. Histology, Physiology, and Transcriptomic and Metabolomic Profiling Reveal the Developmental Dynamics of Annual Shoots in Tree Peonies (Paeonia suffruticosa Andr.). Hortic. Res. 2023, 10, uhad152. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Xia, X.; Su, J.; Wei, M.; Wu, Y.; Tao, J. Overexpression of Herbaceous Peony HSP70 Confers High Temperature Tolerance. BMC Genom. 2019, 20, 70. [Google Scholar] [CrossRef]
- Wang, Q.; Zhou, L.; Yuan, M.; Peng, F.; Zhu, X.; Wang, Y. Genome-Wide Identification of NAC Gene Family Members of Tree Peony (Paeonia suffruticosa Andrews) and Their Expression under Heat and Waterlogging Stress. Int. J. Mol. Sci. 2024, 25, 9312. [Google Scholar] [CrossRef]
- Li, Z.; Ji, W.; Hong, E.; Fan, Z.; Lin, B.; Xia, X.; Chen, X.; Zhu, X. Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics. Horticulturae 2023, 9, 100. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Q.; Wei, L.-L.; Zhao, Y.; Zhang, G.-Z.; Wang, J.; Gu, C.-H. Responses of the Tree Peony (Paeonia suffruticosa, Paeoniaceae) Cultivar “Yu Hong” to Heat Stress Revealed by iTRAQ-Based Quantitative Proteomics. Proteome Sci. 2022, 20, 18. [Google Scholar] [CrossRef] [PubMed]
- Bu, W.; Huang, Y.; Chen, L.; Zhang, M.; Luo, X.; Zheng, T.; Shao, F.; Lei, W.; Xing, W.; Yang, X.; et al. Transcriptome Analysis of Tree Peony under High Temperature Treatment and Functional Verification of PsDREB2A Gene. Plant Physiol. Biochem. 2025, 219, 109405. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Plant Heat Stress: Concepts Directing Future Research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Heat Stress Response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Wang, L.; Hou, J.; Wang, J.; Zhu, Z.; Zhang, W.; Zhang, X.; Shen, H.; Wang, X. Regulatory Roles of HSPA6 in Actinidia chinensis Planch. Root Extract (acRoots)-Inhibited Lung Cancer Proliferation. Clin. Transl. Med. 2020, 10, e46. [Google Scholar] [CrossRef]
- Yang, X.; Huang, Y.; Yao, Y.; Bu, W.; Zhang, M.; Zheng, T.; Luo, X.; Wang, Z.; Lei, W.; Tian, J.; et al. Mining Heat-Resistant Key Genes of Peony Based on Weighted Gene Co-Expression Network Analysis. Genes 2024, 15, 383. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-Z.; Zhou, M.; Ding, Y.-F.; Zhu, C. Gene Networks Involved in Plant Heat Stress Response and Tolerance. Int. J. Mol. Sci. 2022, 23, 11970. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Jiang, S.; Jian, J.; Liu, M.; Yue, Z.; Xu, J.; Li, J.; Xu, C.; Lin, L.; Jing, Y.; et al. Genomic Basis of the Giga-Chromosomes and Giga-Genome of Tree Peony Paeonia ostii. Nat. Commun. 2022, 13, 7328. [Google Scholar] [CrossRef]
- Wang, Q.; Li, B.; Qiu, Z.; Lu, Z.; Hang, Z.; Wu, F.; Chen, X.; Zhu, X. Genome-Wide Identification of MYC Transcription Factors and Their Potential Functions in the Growth and Development Regulation of Tree Peony (Paeonia suffruticosa). Plants 2024, 13, 437. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, W.; Duan, Y.; Xu, Y.; Wang, H.; Hao, J.; Han, Y.; Liu, C. Genome-Wide Identification and Expression Analysis of HSP70 Gene Family Under High-Temperature Stress in Lettuce (Lactuca sativa L.). Int. J. Mol. Sci. 2024, 26, 102. [Google Scholar] [CrossRef]
- Peng, L.; Song, W.; Tan, W.; Liu, Z.; Wang, X.; Li, Y.; Shu, Q. Integration of Genome-Wide Identification, Transcriptome and Association Analysis of HSP20 Gene Family to Revealing Genetic Basis of Floral Organ Number-Related Traits in Tree Peony. Ornam. Plant Res. 2023, 3, 3. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, T.; Liu, Y.; Li, Y.; Wang, M.; Zhu, B.; Liao, D.; Yun, T.; Huang, W.; Zhang, W.; et al. Pumpkin (Cucurbita moschata) HSP20 Gene Family Identification and Expression Under Heat Stress. Front. Genet. 2021, 12, 753953. [Google Scholar] [CrossRef]
- Cho, E.K.; Choi, Y.J. A Nuclear-Localized HSP70 Confers Thermoprotective Activity and Drought-Stress Tolerance on Plants. Biotechnol. Lett. 2009, 31, 597–606. [Google Scholar] [CrossRef]
- Sato, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Complex Plant Responses to Drought and Heat Stress under Climate Change. Plant J. 2024, 117, 1873–1892. [Google Scholar] [CrossRef]
- Huang, B.; Xu, C. Identification and Characterization of Proteins Associated with Plant Tolerance to Heat Stress. J. Integr. Plant Biol. 2008, 50, 1230–1237. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.-M.; Li, J.; He, J.; Liu, H.; Zhang, H.-M. A Class I Cytosolic HSP20 of Rice Enhances Heat and Salt Tolerance in Different Organisms. Sci. Rep. 2020, 10, 1383. [Google Scholar] [CrossRef]
- Huang, J.; Hai, Z.; Wang, R.; Yu, Y.; Chen, X.; Liang, W.; Wang, H. Genome-Wide Analysis of HSP20 Gene Family and Expression Patterns under Heat Stress in Cucumber (Cucumis sativus L.). Front. Plant Sci. 2022, 13, 968418. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Wang, Q.; Li, T.; Gao, H.; Li, H.; Zheng, X.; Wang, X.; Zhang, H.; Cheng, J.; Wang, W.; et al. Phylogenetic and Transcriptional Analyses of the HSP20 Gene Family in Peach Revealed That PpHSP20-32 Is Involved in Plant Height and Heat Tolerance. Int. J. Mol. Sci. 2022, 23, 10849. [Google Scholar] [CrossRef]
- Sarkar, N.K.; Kim, Y.-K.; Grover, A. Rice sHsp Genes: Genomic Organization and Expression Profiling under Stress and Development. BMC Genom. 2009, 10, 393. [Google Scholar] [CrossRef]
- Wang, X.; Yan, B.; Shi, M.; Zhou, W.; Zekria, D.; Wang, H.; Kai, G. Overexpression of a Brassica campestris HSP70 in Tobacco Confers Enhanced Tolerance to Heat Stress. Protoplasma 2016, 253, 637–645. [Google Scholar] [CrossRef]
- Chung, B.Y.W.; Simons, C.; Firth, A.E.; Brown, C.M.; Hellens, R.P. Effect of 5′UTR Introns on Gene Expression in Arabidopsis thaliana. BMC Genom. 2006, 7, 120. [Google Scholar] [CrossRef]
- Lämke, J.; Brzezinka, K.; Bäurle, I. HSFA2 Orchestrates Transcriptional Dynamics after Heat Stress in Arabidopsis thaliana. Transcription 2016, 7, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-A.; Sung, P.-H.; Kuo, Y.-W.; Chen, M.-C.; Jeng, S.-T.; Lin, J.-S. Involvement of microRNA164 in Responses to Heat Stress in Arabidopsis. Plant Sci. 2023, 329, 111598. [Google Scholar] [CrossRef]
- Wang, J.; Gao, X.; Dong, J.; Tian, X.; Wang, J.; Palta, J.A.; Xu, S.; Fang, Y.; Wang, Z. Over-Expression of the Heat-Responsive Wheat Gene TaHSP23.9 in Transgenic Arabidopsis Conferred Tolerance to Heat and Salt Stress. Front. Plant Sci. 2020, 11, 243. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, J.-F.; Kan, Y.; Shan, J.-X.; Ye, W.-W.; Dong, N.-Q.; Guo, T.; Xiang, Y.-H.; Yang, Y.-B.; Li, Y.-C.; et al. A Genetic Module at One Locus in Rice Protects Chloroplasts to Enhance Thermotolerance. Science 2022, 376, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Fang, Y.; Liu, J.; Ye, T.; Li, X.; Tu, H.; Ye, Y.; Wang, Y.; Xiong, L. Stress-Induced Nuclear Translocation of ONAC023 Improves Drought and Heat Tolerance Through Multiple Processes in Rice. Nat. Commun. 2024, 15, 5877. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Huang, Z.; Jiang, H.; Wang, Z.; Wu, F.; Xiong, Y.; Yao, J. A Heat Stress Responsive NAC Transcription Factor Heterodimer Plays Key Roles in Rice Grain Filling. J. Exp. Bot. 2021, 72, 2947–2964. [Google Scholar] [CrossRef]
- Piñero, M.C.; Otálora, G.; Collado, J.; López-Marín, J.; Del Amor, F.M. Foliar Application of Putrescine before a Short-Term Heat Stress Improves the Quality of Melon Fruits (Cucumis melo L.). J. Sci. Food Agric. 2021, 101, 1428–1435. [Google Scholar] [CrossRef]
- Hua, Y.; Liu, Q.; Zhai, Y.; Zhao, L.; Zhu, J.; Zhang, X.; Jia, Q.; Liang, Z.; Wang, D. Genome-Wide Analysis of the HSP20 Gene Family and Its Response to Heat and Drought Stress in Coix (Coix lacryma-jobi L.). BMC Genom. 2023, 24, 478. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Y.; Su, Z.; Shen, Z.; Song, H.; Cai, Z.; Xu, J.; Guo, L.; Zhang, Y.; Guo, S.; et al. Integrated Analysis of HSP20 Genes in the Developing Flesh of Peach: Identification, Expression Profiling, and Subcellular Localization. BMC Plant Biol. 2023, 23, 663. [Google Scholar] [CrossRef]
- Yan, H.; Du, M.; Ding, J.; Song, D.; Ma, W.; Li, Y. Pan-Genome-Wide Investigation and Co-Expression Network Analysis of HSP20 Gene Family in Maize. Int. J. Mol. Sci. 2024, 25, 11550. [Google Scholar] [CrossRef]
- Zhong, L.; Shi, Y.; Xu, S.; Xie, S.; Huang, X.; Li, Y.; Qu, C.; Liu, J.; Liao, J.; Huang, Y.; et al. Heterologous Overexpression of Heat Shock Protein 20 Genes of Different Species of Yellow Camellia in Arabidopsis thaliana Reveals Their Roles in High Calcium Resistance. BMC Plant Biol. 2024, 24, 5. [Google Scholar] [CrossRef]
- Qi, X.; Di, Z.; Li, Y.; Zhang, Z.; Guo, M.; Tong, B.; Lu, Y.; Zhang, Y.; Zheng, J. Genome-Wide Identification and Expression Profiling of Heat Shock Protein 20 Gene Family in Sorbus pohuashanensis (Hance) Hedl under Abiotic Stress. Genes 2022, 13, 2241. [Google Scholar] [CrossRef]
- Ji, H.; Liu, J.; Chen, Y.; Yu, X.; Luo, C.; Sang, L.; Zhou, J.; Liao, H. Bioinformatic Analysis of Codon Usage Bias of HSP20 Genes in Four cruciferous Species. Plants 2024, 13, 468. [Google Scholar] [CrossRef]
- Kan, Y.; Mu, X.-R.; Gao, J.; Lin, H.-X.; Lin, Y. The Molecular Basis of Heat Stress Responses in Plants. Mol. Plant 2023, 16, 1612–1634. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Zhang, J.; Zhao, H.; Tan, S.; Xu, W.; Pan, J.; Yang, F.; Pi, E. ERF Subfamily Transcription Factors and Their Function in Plant Responses to Abiotic Stresses. Front. Plant Sci. 2022, 13, 1042084. [Google Scholar] [CrossRef]
- Cui, F.; Taier, G.; Wang, X.; Wang, K. Genome-Wide Analysis of the HSP20 Gene Family and Expression Patterns of HSP20 Genes in Response to Abiotic Stresses in Cynodon transvaalensis. Front. Genet. 2021, 12, 732812. [Google Scholar] [CrossRef]
- Wang, X.; Tan, N.W.K.; Chung, F.Y.; Yamaguchi, N.; Gan, E.-S.; Ito, T. Transcriptional Regulators of Plant Adaptation to Heat Stress. Int. J. Mol. Sci. 2023, 24, 13297. [Google Scholar] [CrossRef]
- Li, N.; Jiang, M.; Li, P.; Li, X. Identification, Expression, and Functional Analysis of Hsf and Hsp20 Gene Families in Brachypodium Distachyon under Heat Stress. PeerJ 2021, 9, e12267. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, H.; Yuan, H.; Bu, W.; Zhang, M.; Huang, Y.; Hu, J.; Cao, J. Identification and Bioinformatics Analysis of the HSP20 Family in the Peony. Genes 2025, 16, 742. https://doi.org/10.3390/genes16070742
Ma H, Yuan H, Bu W, Zhang M, Huang Y, Hu J, Cao J. Identification and Bioinformatics Analysis of the HSP20 Family in the Peony. Genes. 2025; 16(7):742. https://doi.org/10.3390/genes16070742
Chicago/Turabian StyleMa, Haoran, Heling Yuan, Wenxuan Bu, Minhuan Zhang, Yu Huang, Jian Hu, and Jiwu Cao. 2025. "Identification and Bioinformatics Analysis of the HSP20 Family in the Peony" Genes 16, no. 7: 742. https://doi.org/10.3390/genes16070742
APA StyleMa, H., Yuan, H., Bu, W., Zhang, M., Huang, Y., Hu, J., & Cao, J. (2025). Identification and Bioinformatics Analysis of the HSP20 Family in the Peony. Genes, 16(7), 742. https://doi.org/10.3390/genes16070742