
Direct Oral Anticoagulant-Related Bleeding in Atrial Fibrillation Patients Leads to ADAMTS7 Promoter Demethylation

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
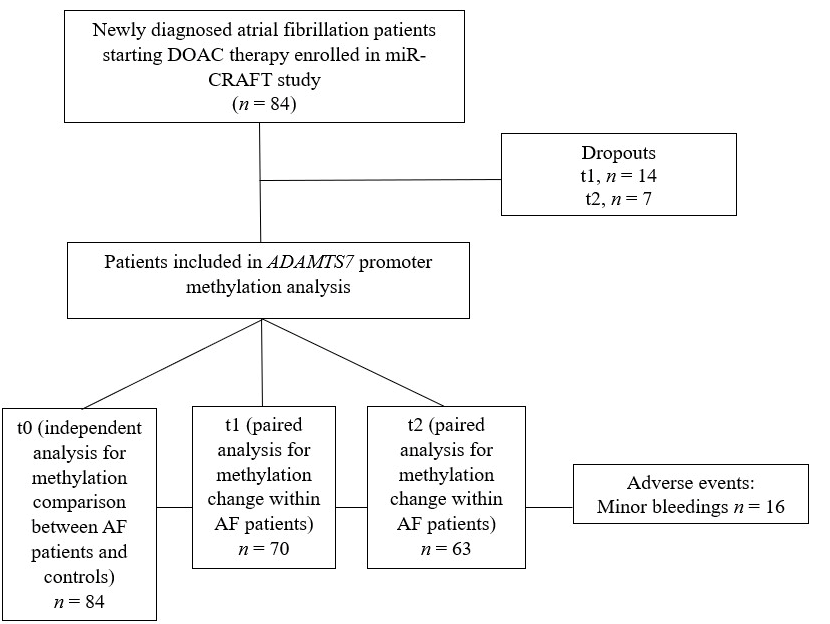
2.1. Study Population
2.2. Genomic DNA Purification and Bisulfite Conversion
2.3. Analysis of DNA Methylation
2.4. Statistics
3. Results
3.1. MIP and qMSP Validation
3.2. Population Characteristics
3.3. ADAMTS7 Promoter Methylation
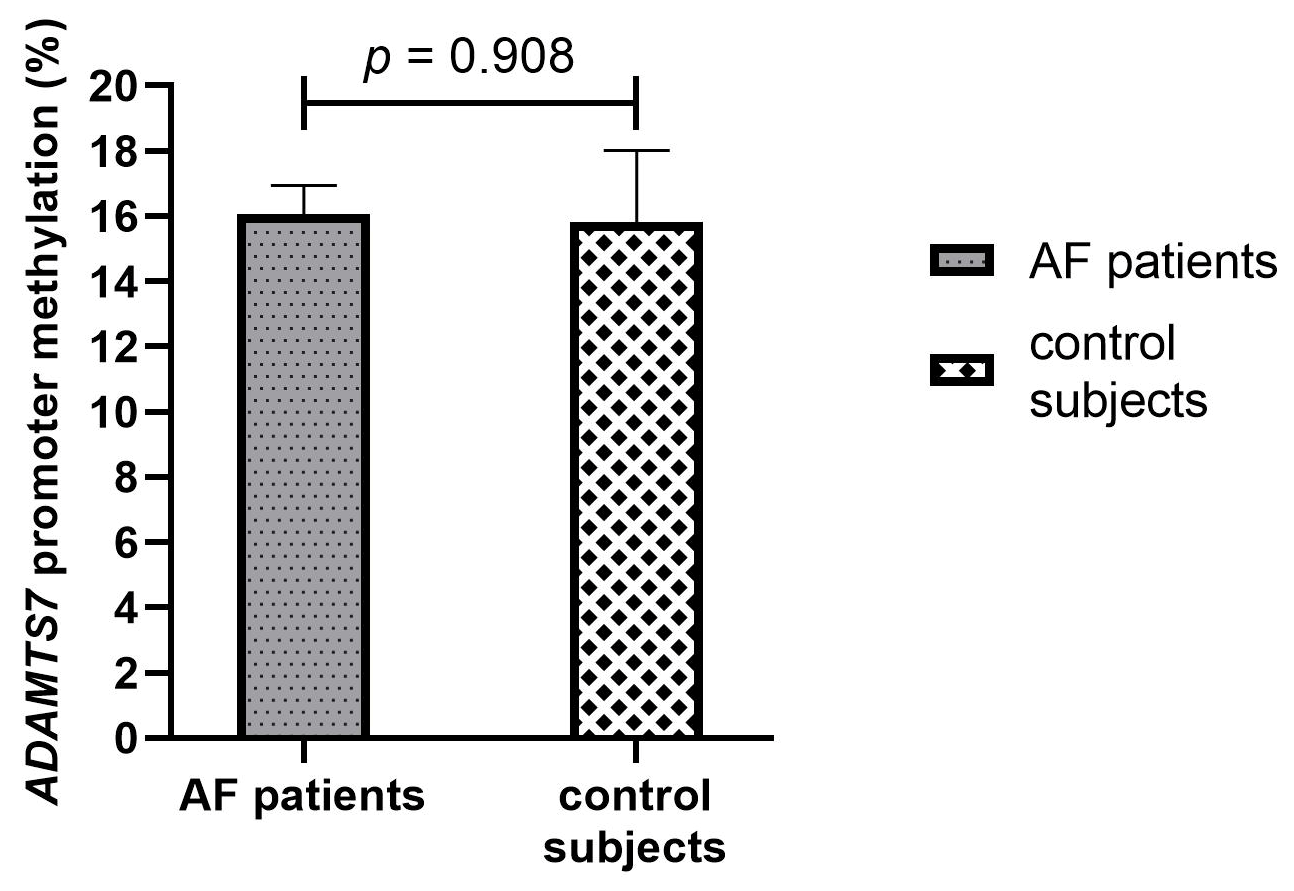
3.3.1. ADAMTS7 Promoter Methylation in AF Patients and Control Subjects
3.3.2. ADAMTS7 Promoter Methylation in DOAC-Treated AF Patients
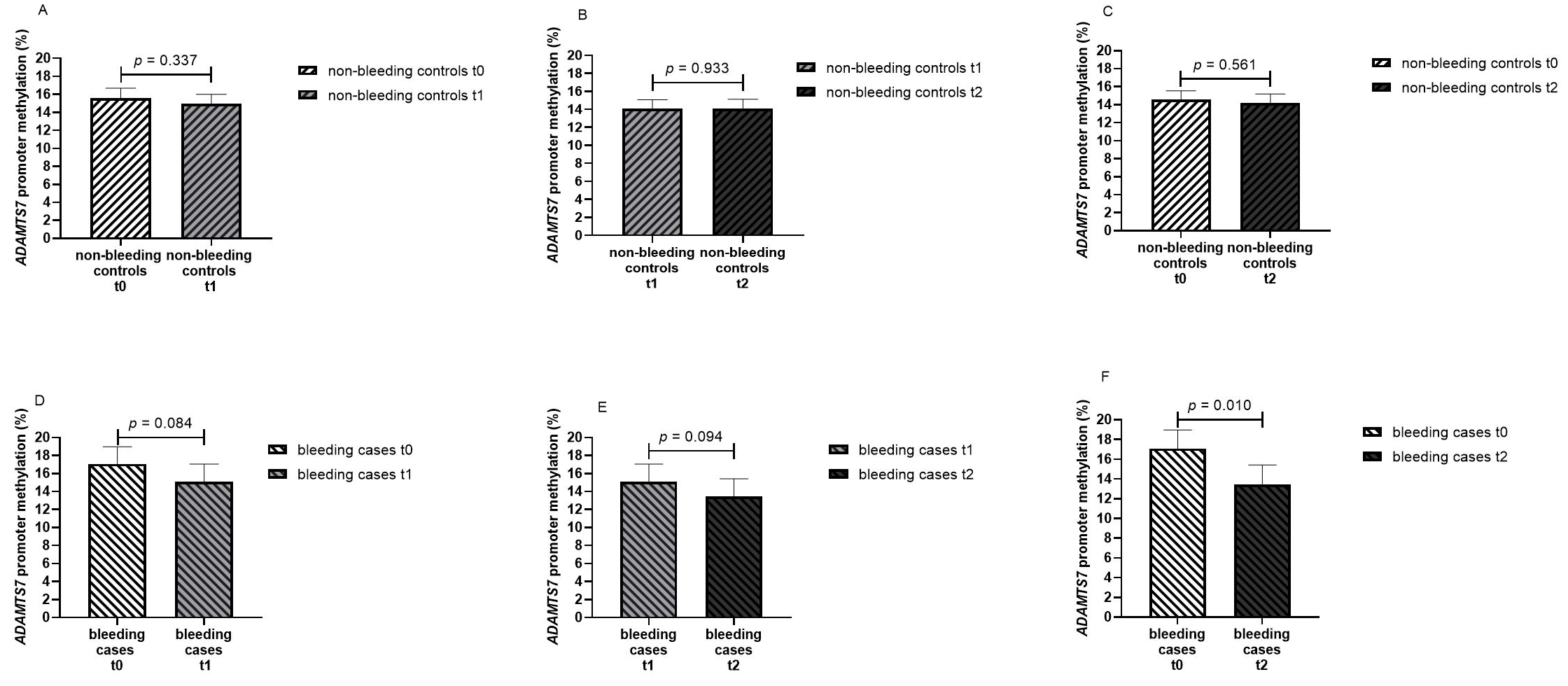
3.3.3. ADAMTS7 Promoter Methylation in Bleeding Cases and Non-Bleeding Controls
3.3.4. Effect of Sex and Co-Morbidities on ADAMTS7 Promoter Methylation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADAMTS7 | A disintegrin and metalloproteinase with thrombospondin motifs 7 |
| DOAC | Direct oral anticoagulant |
| COMP | The cartilage oligomeric protein |
| TSP-1 | Thrombospondin-1 |
| TIMP-1 | Tissue inhibitor of metalloproteinases-1 |
| AF | Atrial fibrillation |
| MMPs | Matrix metalloproteinases |
| GFR | Glomerular filtration rate |
| CI | Confidence intervals |
| TNFa | Tumor necrosis factor alpha |
| PDGF-BB | Platelet-derived growth factor homodimer BB |
References
- Atzemian, N.; Mohammed, S.; Di Venanzio, L.; Gorica, E.; Costantino, S.; Ruschitzka, F.; Paneni, F. Cardiometabolic disease management: Influences from epigenetics. Epigenomics 2025, 17, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Rose, K.W.J.; Taye, N.; Karoulias, S.Z.; Hubmacher, D. Regulation of ADAMTS Proteases. Front. Mol. Biosci. 2021, 8, 701959. [Google Scholar] [CrossRef] [PubMed]
- Mead, T.J.; Apte, S.S. ADAMTS proteins in human disorders. Matrix Biol. 2018, 71–72, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, S.; de Groot, R. ADAMTS proteases in cardiovascular physiology and disease. Open Biol. 2020, 10, 200333. [Google Scholar] [CrossRef]
- Colige, A.; Monseur, C.; Crawley, J.T.B.; Santamaria, S.; de Groot, R. Proteomic discovery of substrates of the cardiovascular protease ADAMTS7. J. Biol. Chem. 2019, 294, 8037–8045. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Keshishian, H.; Mundorff, C.C.; Arduini, A.; Lai, D.; Bendinelli, K.; Popp, N.R.; Bhandary, B.; Clauser, K.R.; Specht, H.; et al. TAILS Identifies Candidate Substrates and Biomarkers of ADAMTS7, a Therapeutic Protease Target in Coronary Artery Disease. Mol. Cell Proteom. 2022, 21, 100223. [Google Scholar] [CrossRef]
- Liu, C.J.; Kong, W.; Ilalov, K.; Yu, S.; Xu, K.; Prazak, L.; Fajardo, M.; Sehgal, B.; Di Cesare, P.E. ADAMTS-7: A metalloproteinase that directly binds to and degrades cartilage oligomeric matrix protein. Faseb J. 2006, 20, 988–990. [Google Scholar] [CrossRef]
- Kessler, T.; Zhang, L.; Liu, Z.; Yin, X.; Huang, Y.; Wang, Y.; Fu, Y.; Mayr, M.; Ge, Q.; Xu, Q.; et al. ADAMTS-7 inhibits re-endothelialization of injured arteries and promotes vascular remodeling through cleavage of thrombospondin-1. Circulation 2015, 131, 1191–1201. [Google Scholar] [CrossRef]
- Sharifi, M.A.; Wierer, M.; Dang, T.A.; Milic, J.; Moggio, A.; Sachs, N.; von Scheidt, M.; Hinterdobler, J.; Müller, P.; Werner, J.; et al. ADAMTS-7 Modulates Atherosclerotic Plaque Formation by Degradation of TIMP-1. Circ. Res. 2023, 133, 674–686. [Google Scholar] [CrossRef]
- Liang, Y.; Fu, Y.; Qi, R.; Wang, M.; Yang, N.; He, L.; Yu, F.; Zhang, J.; Yun, C.H.; Wang, X.; et al. Cartilage oligomeric matrix protein is a natural inhibitor of thrombin. Blood 2015, 126, 905–914. [Google Scholar] [CrossRef]
- Liu, B.; Yang, H.; Song, Y.S.; Sorenson, C.M.; Sheibani, N. Thrombospondin-1 in vascular development, vascular function, and vascular disease. Semin. Cell Dev. Biol. 2024, 155, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv Wound Care (New Rochelle) 2015, 4, 225–234. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Liang, A.; Peng, Y.; Sun, J.; Huang, D.; Xu, K.; Ye, W. Regulation of a disintegrins and metalloproteinase with thrombospondin motifs 7 during inflammation in nucleus pulposus (NP) cells: Role of AP-1, Sp1 and NF-κB signaling. Inflamm. Res. 2016, 65, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, S.; Wang, M.; Liu, G.; Yang, Z.; Wang, L. miR-654-5p Suppresses Migration and Proliferation of Vascular Smooth Muscle Cells by Targeting ADAMTS-7. Cells Tissues Organs 2023, 212, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Liang, L.; Li, Y.; Wei, F.Y.; Mu, N.; Zhang, L.; He, W.; Cao, Y.; Xiong, D.; Li, H. Upregulation of miR-423 improves autologous vein graft restenosis via targeting ADAMTS-7. Int. J. Mol. Med. 2020, 45, 532–542. [Google Scholar] [CrossRef]
- Du, Y.; Gao, C.; Liu, Z.; Wang, L.; Liu, B.; He, F.; Zhang, T.; Wang, Y.; Wang, X.; Xu, M.; et al. Upregulation of a disintegrin and metalloproteinase with thrombospondin motifs-7 by miR-29 repression mediates vascular smooth muscle calcification. Arter. Thromb. Vasc. Biol. 2012, 32, 2580–2588. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, F.; Li, C.; Li, H.; Tang, Q.; Chen, Y.; Yao, Y.; Ding, Z.; Xu, Y.; Chen, A.; et al. Hypomethylation of DNA promoter upregulates ADAMTS7 and contributes to HTR-8/SVneo and JEG-3 cells abnormalities in pre-eclampsia. Placenta 2020, 93, 26–33. [Google Scholar] [CrossRef]
- Ragia, G.; Thomopoulos, T.; Chalikias, G.; Trikas, A.; Tziakas, D.N.; Manolopoulos, V.G. Circulating microRNAs and DNA Methylation as Regulators of Direct Oral Anticoagulant Response in Atrial Fibrillation and Key Elements for the Identification of the Mechanism of Action (miR-CRAFT): Study Design and Patient Enrolment. J. Pers. Med. 2024, 14, 562. [Google Scholar] [CrossRef]
- Ragia, G.; Pallikarou, M.; Michou, C.; Thomopoulos, T.; Chalikias, G.; Trikas, A.; Tziakas, D.N.; Manolopoulos, V.G. Direct oral anticoagulants do not affect miR-27a-3p expression, a regulator of coagulation cascade, in atrial fibrillation patients. J. Thromb. Thrombolysis 2025, 2015, 1–10. [Google Scholar] [CrossRef]
- Lu, L.; Katsaros, D.; de la Longrais, I.A.; Sochirca, O.; Yu, H. Hypermethylation of let-7a-3 in epithelial ovarian cancer is associated with low insulin-like growth factor-II expression and favorable prognosis. Cancer Res. 2007, 67, 10117–10122. [Google Scholar] [CrossRef]
- Kessler, T.; Schunkert, H. Targeting ADAMTS-7: A Vaccination Against Atherosclerosis-and Its Complications? Circulation 2023, 147, 743–745. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Feng, Q.; Kong, W. ECM Microenvironment in Vascular Homeostasis: New Targets for Atherosclerosis. Physiology 2024, 39, 324–344. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Kong, W. Cartilage Oligomeric Matrix Protein: Matricellular and Matricrine Signaling in Cardiovascular Homeostasis and Disease. Curr. Vasc. Pharmacol. 2017, 15, 186–196. [Google Scholar] [CrossRef]
- Leberzammer, J.; von Hundelshausen, P. Chemokines, molecular drivers of thromboinflammation and immunothrombosis. Front. Immunol. 2023, 14, 1276353. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, J.; Bai, X.; Liu, B.; Liu, C.J.; Xu, Q.; Zhu, Y.; Wang, N.; Kong, W.; Wang, X. ADAMTS-7 mediates vascular smooth muscle cell migration and neointima formation in balloon-injured rat arteries. Circ. Res. 2009, 104, 688–698. [Google Scholar] [CrossRef]
- Bauer, K.A.; ten Cate, H.; Barzegar, S.; Spriggs, D.R.; Sherman, M.L.; Rosenberg, R.D. Tumor necrosis factor infusions have a procoagulant effect on the hemostatic mechanism of humans. Blood 1989, 74, 165–172. [Google Scholar] [CrossRef]
- Ernofsson, M.; Siegbahn, A. Platelet-derived growth factor-BB and monocyte chemotactic protein-1 induce human peripheral blood monocytes to express tissue factor. Thromb. Res. 1996, 83, 307–320. [Google Scholar] [CrossRef]
- Lawler, J. Counter regulation of tumor angiogenesis by vascular endothelial growth factor and thrombospondin-1. Semin. Cancer Biol. 2022, 86, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Krishnaswamy, V.R.; Mintz, D.; Sagi, I. Matrix metalloproteinases: The sculptors of chronic cutaneous wounds. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2220–2227. [Google Scholar] [CrossRef]
- Wang, X.; Khalil, R.A. Matrix Metalloproteinases, Vascular Remodeling, and Vascular Disease. Adv. Pharmacol. 2018, 81, 241–330. [Google Scholar] [CrossRef] [PubMed]
- Kandhwal, M.; Behl, T.; Singh, S.; Sharma, N.; Arora, S.; Bhatia, S.; Al-Harrasi, A.; Sachdeva, M.; Bungau, S. Role of matrix metalloproteinase in wound healing. Am. J. Transl. Res. 2022, 14, 4391–4405. [Google Scholar] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Kanaya, T.; Kyo, S.; Maida, Y.; Yatabe, N.; Tanaka, M.; Nakamura, M.; Inoue, M. Frequent hypermethylation of MLH1 promoter in normal endometrium of patients with endometrial cancers. Oncogene 2003, 22, 2352–2360. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Z.; Ding, Y.; Wang, L.; Wang, S.; Wang, H.; Qin, Y. DNA Methylation: From Cancer Biology to Clinical Perspectives. Front. Biosci. 2022, 27, 326. [Google Scholar] [CrossRef] [PubMed]
- Bhootra, S.; Jill, N.; Shanmugam, G.; Rakshit, S.; Sarkar, K. DNA methylation and cancer: Transcriptional regulation, prognostic, and therapeutic perspective. Med. Oncol. 2023, 40, 71. [Google Scholar] [CrossRef]
- Manolopoulos, V.G.; Ragia, G. Fluoropyrimidine Toxicity: The Hidden Secrets of DPYD. Curr. Drug Metab. 2024, 25, 91–95. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | miR-CRAFT Population (n = 84) | Bleeding Cases (n = 16) | Non-Bleeding Controls (n = 68) |
|---|---|---|---|
| Demographic | |||
| Male (n, %) | 39 (46.4) | 8 (50) | 31 (45.6) |
| Age (years, mean ± SD) | 69 (11) | 71 (9) | 69 (12) |
| Weight (kg, median, 25, 75 percentiles) | 79 (65, 92) | 72 (60, 92) | 80 (65, 93.7) |
| Height (cm, median, 25, 75 percentiles) | 170 (160, 176) | 164 (156, 175) | 170 (160, 177) |
| Smokers (n, %) | 14 (16.7) | 3 (18.7) | 11 (16.2) |
| Biochemical | |||
| Hemoglobin (gr%, median, 25, 75 percentiles) | 13.9 (12.5, 15.1) | 14.6 (13.2, 14.9) | 13.7 (12.5, 15.2) |
| Platelets (100/μL, mean ± SD) | 232 (69) | 232 (64) | 232 (71) |
| Urea (mg/dL, median, 25, 75 percentiles) | 40 (33, 52.5) | 41 (37.2, 56.7) | 40 (32, 50.7) |
| Creatinine (mg/dL, median, 25, 75 percentiles) | 0.94 (0.8, 1.1) | 0.89 (0.77, 1.01) | 1.00 (0.80, 1.20) |
| Creatinine Clearance (mL/min, median, 25, 75 percentiles) | 78.8 (52.1, 97.8) | 66.8 (50.9, 108.2) | 80.1 (57.7, 97.8) |
| SGOT (U/L, median, 25, 75 percentiles) | 22 (18, 30) | 19 (17, 26) | 22.5 (18, 32.2) |
| SGOT > 40 U/L (n, %) | 9 (10.7) | 1 (6.3) | 8 (11.7) |
| SGPT (U/L, median, 25, 75 percentiles) | 25 (19, 36) | 25 (20, 30) | 25 (19, 37) |
| SGPT > 56 U/L (n, %) | 8 (9.5) | 1 (6.3) | 7 (10.3) |
| Clinical | |||
| CHA2DS2-Vasc score (median, 25, 75 percentiles) | 3 (2, 4) | 3 (2, 4) | 3 (1.25, 4) |
| HAS-BLED score (median, 25, 75 percentiles) | 0 (0, 1) | 0 (0, 1) | 0.5 (0, 1) |
| Renal function based on GFR (data for 58 patients) | |||
| Normal (n, %) | 16 (27.6) | 4 (25.0) | 12 (17.6) |
| Mildly decreased (n, %) | 26 (44.8) | 5 (31.3) | 21 (30.9) |
| Moderately decreased (n, %) | 14 (24.1) | 6 (37.5) | 8 (11.8) |
| Severely decreased (n, %) | 2 (3.4) | - | 2 (2.9) |
| Failure (n, %) | - | - | - |
| DOAC therapy | |||
| Rivaroxaban (n, %) | 28 (33.3) | 6 (37.5) | 22 (32.4) |
| Apixaban (n, %) | 36 (42.9) | 7 (43.8) | 29 (42.6) |
| Dabigatran (n, %) | 20 (23.8) | 3 (18.7) | 17 (25.0) |
| Bleeding, Minor (n, %) | 16 (19.0) | 16 | - |
| Co-morbidities | |||
| Hypertension (n, %) | 45 (53.6) | 15 (93.7) | 30 (44.1) * |
| Type 2 Diabetes (n, %) | 19 (22.6) | 7 (43.7) | 12 (17.6) |
| Dyslipidemia (n, %) | 37 (44.0) | 12 (75.0) | 25 (36.7) ** |
| Co-medications | |||
| Salicylates (n,%) | 3 (3.6) | 1 (6.3) | 2 (3.0) |
| Antiplatelets (n,%) | - | - | - |
| NSAIDs (n,%) | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragia, G.; Thomopoulos, T.; Pallikarou, M.; Atzemian, N.; Maslarinou, A.; Chalikias, G.; Trikas, A.; Tziakas, D.N.; Manolopoulos, V.G. Direct Oral Anticoagulant-Related Bleeding in Atrial Fibrillation Patients Leads to ADAMTS7 Promoter Demethylation. Genes 2025, 16, 698. https://doi.org/10.3390/genes16060698
Ragia G, Thomopoulos T, Pallikarou M, Atzemian N, Maslarinou A, Chalikias G, Trikas A, Tziakas DN, Manolopoulos VG. Direct Oral Anticoagulant-Related Bleeding in Atrial Fibrillation Patients Leads to ADAMTS7 Promoter Demethylation. Genes. 2025; 16(6):698. https://doi.org/10.3390/genes16060698
Chicago/Turabian StyleRagia, Georgia, Thomas Thomopoulos, Myria Pallikarou, Natalia Atzemian, Anthi Maslarinou, Georgios Chalikias, Athanasios Trikas, Dimitrios N. Tziakas, and Vangelis G. Manolopoulos. 2025. "Direct Oral Anticoagulant-Related Bleeding in Atrial Fibrillation Patients Leads to ADAMTS7 Promoter Demethylation" Genes 16, no. 6: 698. https://doi.org/10.3390/genes16060698
APA StyleRagia, G., Thomopoulos, T., Pallikarou, M., Atzemian, N., Maslarinou, A., Chalikias, G., Trikas, A., Tziakas, D. N., & Manolopoulos, V. G. (2025). Direct Oral Anticoagulant-Related Bleeding in Atrial Fibrillation Patients Leads to ADAMTS7 Promoter Demethylation. Genes, 16(6), 698. https://doi.org/10.3390/genes16060698