
Examination of Runs of Homozygosity Distribution Patterns and Relevant Candidate Genes of Potential Economic Interest in Russian Goat Breeds Using Whole-Genome Sequencing

 ,
,  ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Sampling
2.2. DNA Extraction and Whole-Genome Sequencing
2.3. Data Processing and in Silico Analyses
- (1)
- windowSize is the size of the sliding window (i.e., number of SNP loci; default = 15);
- (2)
- threshold is the threshold of overlapping windows of the same state (homozygous/heterozygous) to call an SNP in a RUN (default = 0.05);
- (3)
- minSNP is the minimum number of SNPs in a RUN (default = 3);
- (4)
- ROHet is a heterozygosity/homozygosity parameter for whether runs of heterozygosity (ROHet) or homozygosity (ROHom) are detected (default = FALSE);
- (5)
- maxOppWindow is the maximum number of homozygous/heterozygous SNPs in the sliding window (default = 1);
- (6)
- maxMissWindow is the maximum number of missing SNPs in the sliding window (default = 1);
- (7)
- maxGap is the maximum distance between consecutive SNPs to be still considered a potential run (default = 106 bp);
- (8)
- minLengthBps is the minimum length of a run in bp (defaults to 1000 bp = 1 Kb);
- (9)
- minDensity is the minimum number of SNPs per Kb (defaults to 0.1 = 1 SNP every 10 Kb);
- (10)
- maxOppRun is the maximum number of opposite genotype SNPs in the run (optional);
- (11)
- maxMissRun is the maximum number of missing SNPs in the run (optional).
3. Results
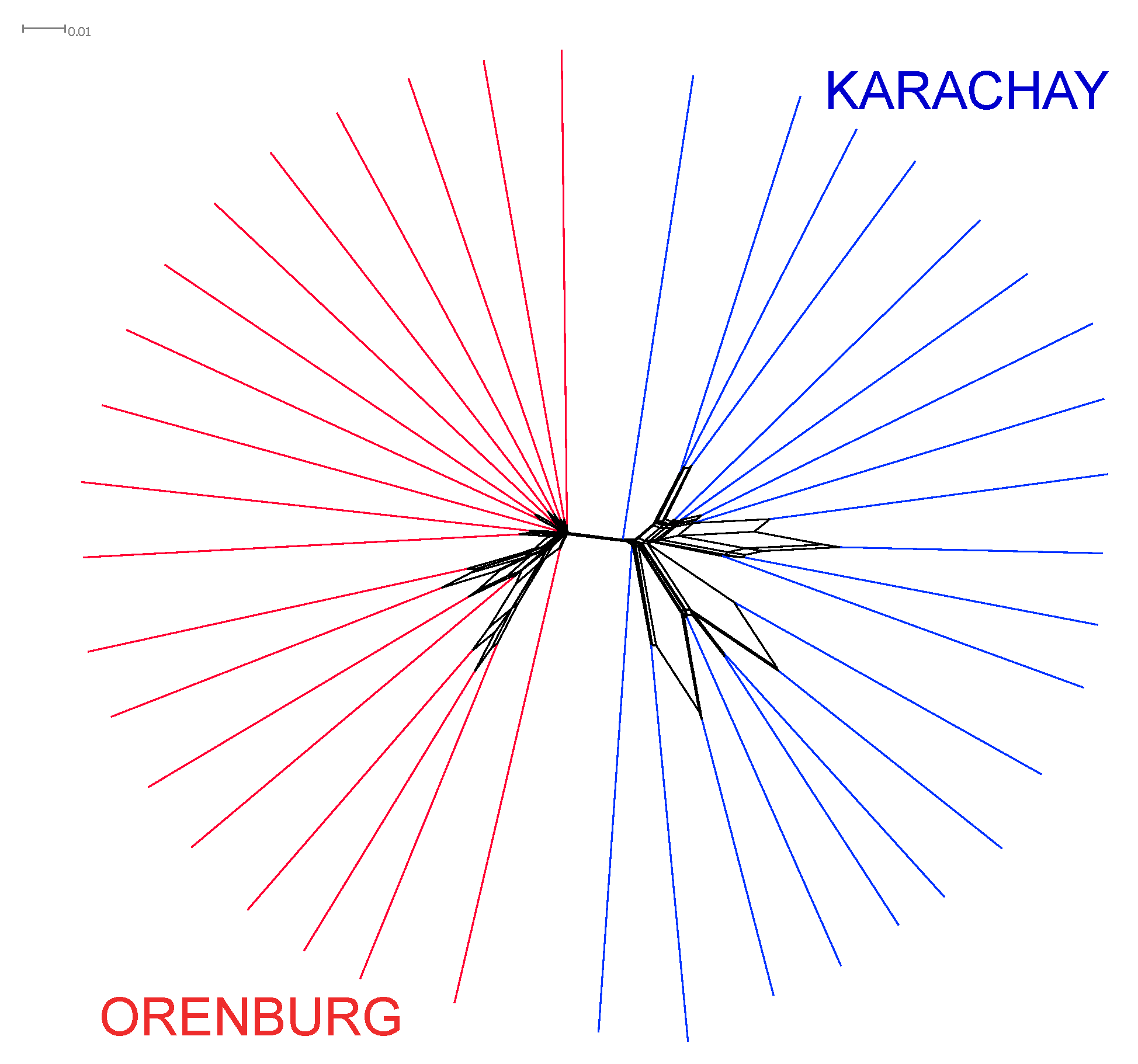
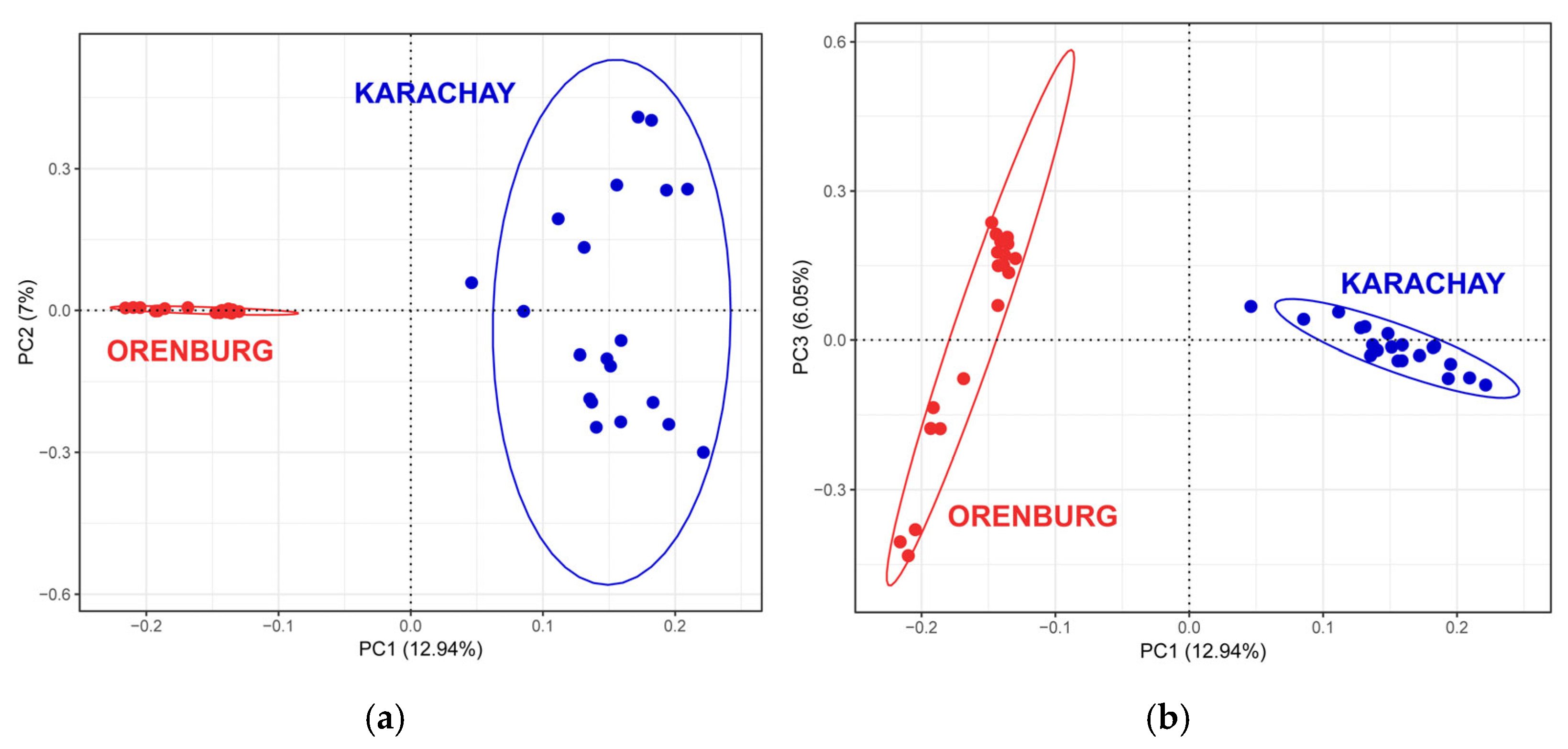
3.1. Between-Breed Genetic Differentiation
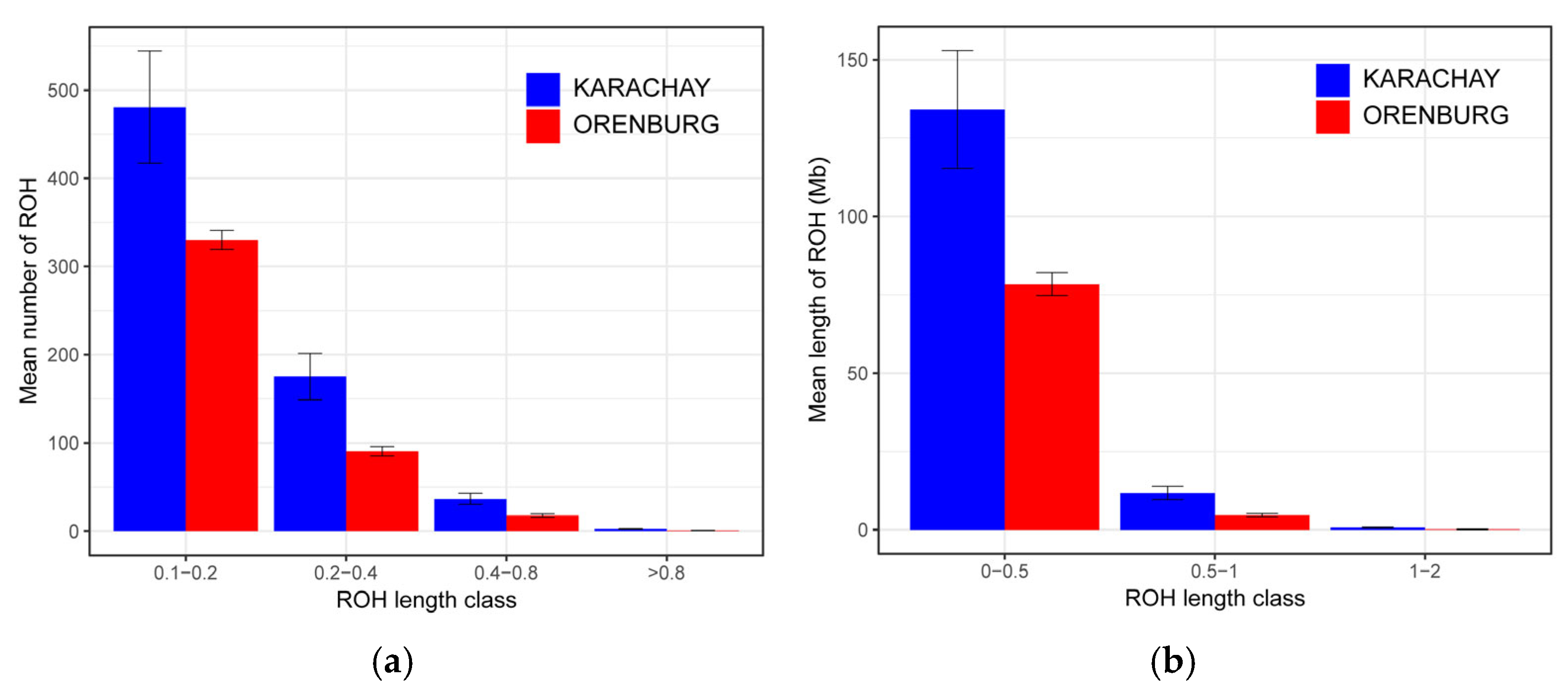
3.2. Runs of Homozogosity Patterns
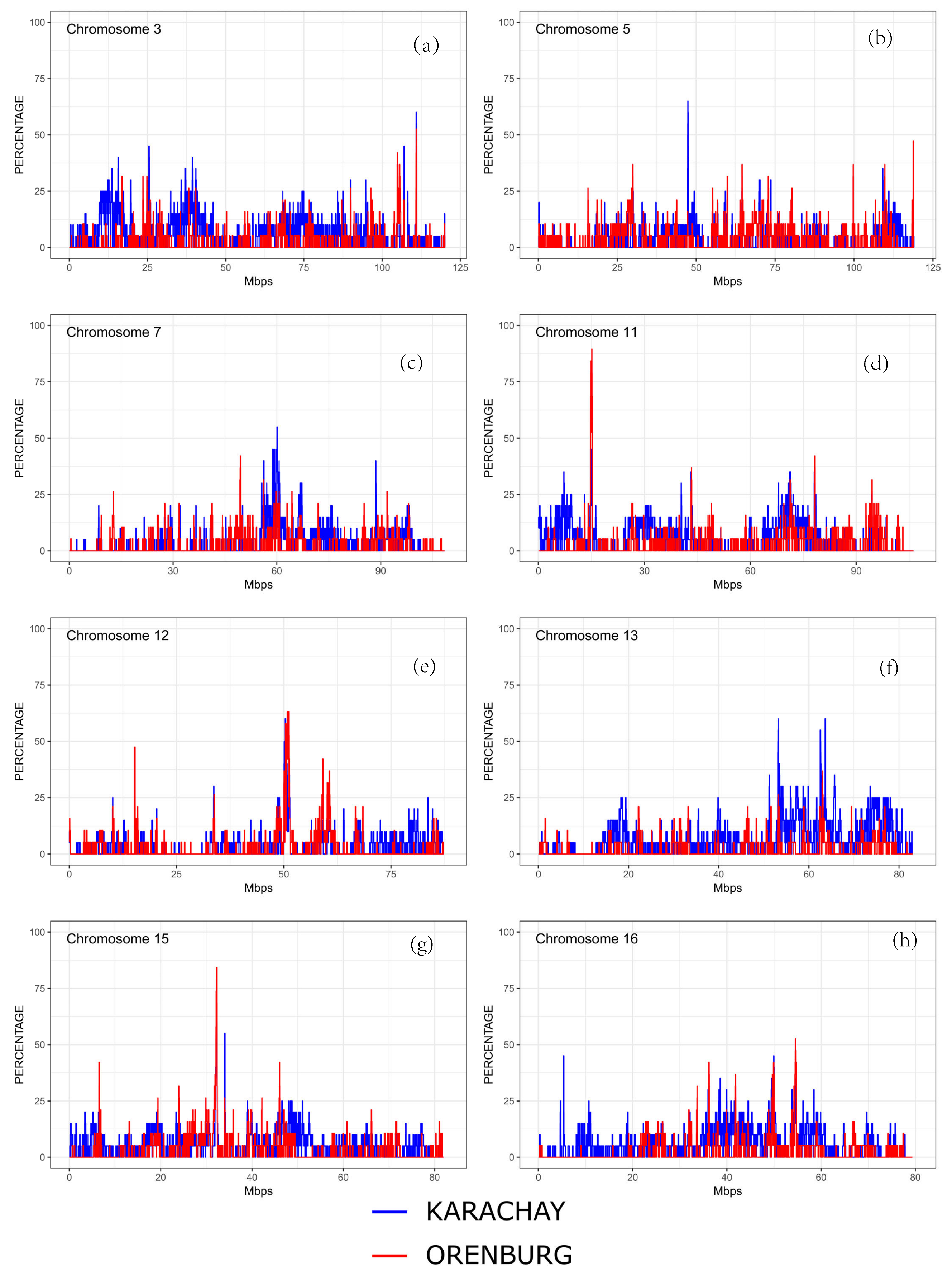
3.3. Distribution of ROH Islands and Candidate Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Signer-Hasler, H.; Henkel, J.; Bangerter, E.; Bulut, Z.; Drögemüller, C.; Leeb, T.; Flury, C. Runs of homozygosity in Swiss goats reveal genetic changes associated with domestication and modern selection. Genet. Sel. Evol. 2022, 54, 6. [Google Scholar] [CrossRef] [PubMed]
- Saif, R.; Henkel, J.; Mahmood, T.; Ejaz, A.; Ahmad, F.; Zia, S. Detection of whole genome selection signatures of Pakistani Teddy goat. Mol. Biol. Rep. 2021, 48, 7273–7280. [Google Scholar] [CrossRef] [PubMed]
- Dan, H.; Zhong, H.; Akhatayeva, Z.; Lin, K.; Xu, S. Whole-genome selective scans detect genes associated with cashmere traits and climatic adaptation in cashmere goats (Capra hircus) in China. Genes 2025, 16, 292. [Google Scholar] [CrossRef]
- Gudra, D.; Valdovska, A.; Jonkus, D.; Kairisa, D.; Galina, D.; Ustinova, M.; Viksne, K.; Fridmanis, D.; Kalnina, I. Genetic characterization of the Latvian local goat breed and genetic traits associated with somatic cell count. Animal 2024, 18, 101154. [Google Scholar] [CrossRef]
- Liu, J.; Dong, S.; Lv, J.; Li, Y.; Sun, B.; Guo, Y.; Deng, M.; Liu, D.; Liu, G. Screening of SNP loci related to leg length trait in Leizhou goats based on whole-genome resequencing. Int. J. Mol. Sci. 2024, 25, 12450. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Bao, J.; Hu, W.; Shang, M.; Zhang, L. Whole-genome resequencing reveals genetic diversity and selection characteristics of dairy goat. Front. Genet. 2023, 13, 1044017. [Google Scholar] [CrossRef]
- Sheriff, O.; Ahbara, A.M.; Haile, A.; Alemayehu, K.; Han, J.L.; Mwacharo, J.M. Whole-genome resequencing reveals genomic variation and dynamics in Ethiopian indigenous goats. Front. Genet. 2024, 15, 1353026. [Google Scholar] [CrossRef]
- Wang, J.J.; Li, Z.D.; Zheng, L.Q.; Zhang, T.; Shen, W.; Lei, C.Z. Genome-wide detection of selective signals for fecundity traits in goats (Capra hircus). Gene 2022, 818, 146221. [Google Scholar] [CrossRef]
- He, S.; Di, J.; Han, B.; Chen, L.; Liu, M.; Li, W. Genome-wide scan for runs of homozygosity identifies candidate genes related to economically important traits in Chinese Merino. Animals 2020, 10, 524. [Google Scholar] [CrossRef]
- Zinovieva, N.A.; Dotsev, A.V.; Sermyagin, A.A.; Deniskova, T.E.; Abdelmanova, A.S.; Kharzinova, V.R.; Sölkner, J.; Reyer, H.; Wimmers, K.; Brem, G. Selection signatures in two oldest Russian native cattle breeds revealed using high-density single nucleotide polymorphism analysis. PLoS ONE 2020, 15, e0242200. [Google Scholar] [CrossRef]
- Abdelmanova, A.S.; Dotsev, A.V.; Romanov, M.N.; Stanishevskaya, O.I.; Gladyr, E.A.; Rodionov, A.N.; Vetokh, A.N.; Volkova, N.A.; Fedorova, E.S.; Gusev, I.V.; et al. Unveiling comparative genomic trajectories of selection and key candidate genes in egg-type Russian White and meat-type White Cornish chickens. Biology 2021, 10, 876. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Liu, J.; Ma, X.; Yang, J. Genome-wide runs of homozygosity reveal inbreeding levels and trait-associated candidate genes in diverse sheep breeds. Genes 2025, 16, 316. [Google Scholar] [CrossRef] [PubMed]
- Romanov, M.N.; Shakhin, A.V.; Abdelmanova, A.S.; Volkova, N.A.; Efimov, D.N.; Fisinin, V.I.; Korshunova, L.G.; Anshakov, D.V.; Dotsev, A.V.; Griffin, D.K.; et al. Dissecting selective signatures and candidate genes in grandparent lines subject to high selection pressure for broiler production and in a local Russian chicken breed of Ushanka. Genes 2024, 15, 524. [Google Scholar] [CrossRef]
- Liu, D.; Chen, Z.; Zhao, W.; Guo, L.; Sun, H.; Zhu, K.; Liu, G.; Shen, X.; Zhao, X.; Wang, Q.; et al. Genome-wide selection signatures detection in Shanghai Holstein cattle population identified genes related to adaption, health and reproduction traits. BMC Genom. 2021, 22, 747. [Google Scholar] [CrossRef]
- Fedorova, E.S.; Dementieva, N.V.; Shcherbakov, Y.S.; Stanishevskaya, O.I. Identification of key candidate genes in runs of homozygosity of the genome of two chicken breeds, associated with cold adaptation. Biology 2022, 11, 547. [Google Scholar] [CrossRef]
- Liu, X.; Peng, Y.; Zhang, X.; Chen, W.; Chen, Y.; Wei, L.; Zhu, Q.; Khan, M.Z.; Wang, C. Potential genetic markers associated with environmental adaptability in herbivorous livestock. Animals 2025, 15, 748. [Google Scholar] [CrossRef]
- Dementieva, N.V.; Kudinov, A.A.; Larkina, T.A.; Mitrofanova, O.V.; Dysin, A.P.; Terletsky, V.P.; Tyshchenko, V.I.; Griffin, D.K.; Romanov, M.N. Genetic variability in local and imported germplasm chicken populations as revealed by analyzing runs of homozygosity. Animals 2020, 10, 1887. [Google Scholar] [CrossRef] [PubMed]
- Romanov, M.N.; Abdelmanova, A.S.; Fisinin, V.I.; Gladyr, E.A.; Volkova, N.A.; Koshkina, O.A.; Rodionov, A.N.; Vetokh, A.N.; Gusev, I.V.; Anshakov, D.V.; et al. Selective footprints and genes relevant to cold adaptation and other phenotypic traits are unscrambled in the genomes of divergently selected chicken breeds. J. Anim. Sci. Biotechnol. 2023, 14, 35. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; Fang, Y.; Cao, C.; Zhang, Z.; Pan, Y.; Wang, Q. Runs of homozygosity revealed reproductive traits of Hu sheep. Genes 2022, 13, 1848. [Google Scholar] [CrossRef]
- Zhao, F.; Xie, R.; Fang, L.; Xiang, R.; Yuan, Z.; Liu, Y.; Wang, L. Analysis of 206 whole-genome resequencing reveals selection signatures associated with breed-specific traits in Hu sheep. Evol. Appl. 2024, 17, e13697. [Google Scholar] [CrossRef]
- Davoudi, P.; Do, D.N.; Rathgeber, B.; Colombo, S.; Sargolzaei, M.; Plastow, G.; Wang, Z.; Miar, Y. Characterization of runs of homozygosity islands in American mink using whole-genome sequencing data. J. Anim. Breed. Genet. 2024, 141, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Ayalew, W.; Wu, X.; Tarekegn, G.M.; Sisay Tessema, T.; Naboulsi, R.; Van Damme, R.; Bongcam-Rudloff, E.; Edea, Z.; Enquahone, S.; Yan, P. Whole-genome resequencing reveals selection signatures of Abigar cattle for local adaptation. Animals 2023, 13, 3269. [Google Scholar] [CrossRef]
- Sun, X.; Guo, J.; Li, L.; Zhong, T.; Wang, L.; Zhan, S.; Lu, J.; Wang, D.; Dai, D.; Liu, G.E.; et al. Genetic diversity and selection signatures in Jianchang Black goats revealed by whole-genome sequencing data. Animals 2022, 12, 2365. [Google Scholar] [CrossRef] [PubMed]
- Orekhov, A.A. Goats. In Animal Genetic Resources of the USSR; FAO Animal Production and Health Paper; Dmitriev, N.G., Ernst, L.K., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 1989; Volume 65, pp. 344–365. Available online: https://www.fao.org/4/ah759e/AH759E14.htm (accessed on 25 April 2025).
- Tarasova, E.I.; Frolov, A.N.; Lebedev, S.V.; Romanov, M.N. Istoriya, razvedeniye, selektsiya i genetika orenburgskoy porody koz [History, Breeding, Selection and Genetics of the Orenburg Goat Breed]. In Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Materials of the 3rd International Scientific and Practical Conference, Moscow, Russia, 30 September 2021; Pozyabin, S.V., Kochish, I.I., Romanov, M.N., Eds.; Ministry of Agriculture of the Russian Federation, Federal State Budgetary Educational Institution of Higher Education “Moscow State Academy of Veterinary Medicine and Biotechnology–MVA named after K.I. Scriabin”; Sel’skokhozyaistvennye tekhnologii: Moscow, Russia, 2021; pp. 450–454. [Google Scholar] [CrossRef]
- Tarasova, E.I.; Frolov, A.N.; Lebedev, S.V.; Romanov, M.N. Landmark native breed of the Orenburg goats: Progress in its breeding and genetics and future prospects. Anim. Biotechnol. 2023, 34, 5139–5154. [Google Scholar] [CrossRef] [PubMed]
- Royal Commission. Official Descriptive and Illustrated Catalogue of the Great Exhibition of the Works of Industry of All Nations; Spicer Brothers: London, UK, 1851; Volume 3, p. 1383. [Google Scholar]
- Petrov, N.I. Produktivnost’ i nasledovanie masti potomstvom orenburgskikh koz [Productivity and inheritance of color type by the progeny of Orenburg goats]. Vestn. Myasn. Skotovod. [Her. Beef Cattle Breed.] 2015, 4, 47–50. Available online: https://www.elibrary.ru/item.asp?id=25286682 (accessed on 25 April 2025). (In Russian with English summary).
- Belkov, G.I.; Panin, V.A. Kachestvennyye i kolichestvennyye pokazateli myasnoy produktivnosti koz orenburgskoy porody [Qualitative and quantitative indicators of meat productivity of Orenburg goats]. Izv. Orenburg. Gos. Agrar. Un-Ta [Izv. Orenburg State Agrar. Univ.] 2021, 3, 304–307, (In Russian with English summary). [Google Scholar]
- Kharlamov, A.V.; Panin, V.A. Kompleksnaya otsenka pokazateley produktivnosti koz orenburgskoy porody v usloviyakh Orenburgskogo regiona [Comprehensive assessment of productivity indicators of Orenburg goats in the conditions of the Orenburg region]. Nauka i obraz. [Sci. Educ.] 2023, 1–2, 123–132, (In Russian with English summary). [Google Scholar]
- Abdelmanova, A.S.; Deniskova, T.E.; Petrov, S.N.; Frolov, A.N.; Platonov, S.A.; Gladyr, E.A.; Gusev, I.V.; Lebedev, S.V.; Zinovieva, N.A. Otsenka dinamiki geneticheskogo raznoobraziya populyatsiy orenburgskoy porody koz s ispol’zovaniyem mikrosatellitnykh markerov [Assessment of the dynamics of genetic diversity of Orenburg goat breed populations by microsatellite markers]. Dostizheniya Nauki I Tekhniki APK [Achiev. Sci. Technol. AIC] 2024, 38, 50–56. [Google Scholar]
- Deniskova, T.E.; Dotsev, A.V.; Abdelmanova, A.S.; Petrov, S.N.; Frolov, A.N.; Platonov, S.A.; Gladyr, E.A.; Gusev, I.V.; Selionova, M.I.; Rodionov, A.N.; et al. Genetic diversity in the Orenburg goat breed revealed by single-nucleotide polymorphism (SNP) analysis: Initial steps in saving a threatened population. Genes 2024, 15, 1375. [Google Scholar] [CrossRef]
- Scherf, B.D. (Ed.) World Watch List for Domestic Animal Diversity, 2nd ed.; FAO, UNEP: Rome, Italy, 1995; Available online: https://www.fao.org/faostat/en/ (accessed on 26 November 2015).
- Aybazov, M.M.; Selionova, M.I.; Mamontova, T.V. Ekster’yernyye i nekotoryye biologicheskiye pokazateli karachayevskikh koz [Exterior and some biological indices of Karachai goats]. Zootehniâ [Zootech] 2019, 12, 5–9, (In Russian with English summary). [Google Scholar]
- Selionova, M.; Aibazov, M.; Sermyagin, A.; Belous, A.; Deniskova, T.; Mamontova, T.; Zharkova, E.; Zinovieva, N. Genome-wide association and pathway analysis of carcass and meat quality traits in Karachai young goats. Animals 2023, 13, 3237. [Google Scholar] [CrossRef] [PubMed]
- Selionova, M.; Trukhachev, V.; Aibazov, M.; Sermyagin, A.; Belous, A.; Gladkikh, M.; Zinovieva, N. Genome-wide association study of milk composition in Karachai goats. Animals 2024, 14, 327. [Google Scholar] [CrossRef]
- Selionova, M.I.; Trukhachev, V.I.; Aybazov, A.-M.M.; Stolpovsky, Y.A.; Zinovieva, N.A. Genetic markers of goats (review). Sel’skokhozyaistvennaya Biol. [Agric. Biol.] 2021, 56, 1031–1048. [Google Scholar] [CrossRef]
- Deniskova, T.E.; Dotsev, A.V.; Selionova, M.I.; Reyer, H.; Sölkner, J.; Fornara, M.S.; Aybazov, A.M.; Wimmers, K.; Brem, G.; Zinovieva, N.A. SNP-based genotyping provides insight into the West Asian origin of Russian local goats. Front. Genet. 2021, 12, 708740. [Google Scholar] [CrossRef]
- Sermyagin, A.A.; Deniskova, T.E.; Gusev, I.V.; Petrov, S.N.; Rodionov, A.N.; Dotsev, A.V.; Zinovieva, N.A. Identification of SNPs associated with growth and development traits of goats (Capra hircus Linnaeus, 1758) from the resource population in age dynamics. Sel’skokhozyaistvennaya Biol. [Agric. Biol.] 2024, 59, 633–648. [Google Scholar] [CrossRef]
- Selionova, M.I.; Mamontova, T.V.; Aibazov, A.-M.M. Osobennosti reproduktivnoy funktsii karachayevskikh koz v zavisimosti ot raznykh geograficheskikh rayonov razvedeniya [Features of the reproductive function of Karachay goats depending on different geographical areas of breeding]. Izv. Timiryazevsk. S-Kh. Akad. [Izv. Timiryazev Agric. Acad.] 2021, 2, 114–122, (In Russian with English summary). [Google Scholar]
- Mamontova, T.V.; Gadzhiev, Z.K.; Aibazov, A.-M.M. Produktivnyye i vosproizvoditel’nyye osobennosti mestnykh karachayevskikh koz [Productive and reproductive features of local Karachay goats]. Sb. Nauch. Tr. Stavropol’skogo Nauch.-Issled. In-Ta Zhivotnovodstva I Kormoproizvodstva [Collect. Sci. Pap. Stavropol Res. Inst. Anim. Husb. Forage Prod.] 2011, 1, 15–17. Available online: https://www.elibrary.ru/item.asp?id=17294528 (accessed on 25 April 2025).
- Aybazov, M.M.; Selionova, M.I.; Seitov, M.S.; Bikteev, S.M. Avtokhtonnaya karachayevskaya poroda koz: Genotipy i nekotoryye ekster’yernyye i inter’yernyye pokazateli [Autochthonous Karachay breed of goats: Genotypes and some exterior and interior indicators]. Izv. Orenburg. Gos. Agrar. Un-Ta [Izv. Orenburg State Agrar. Univ.]. 2022, 5, 300–306, (In Russian with English summary). [Google Scholar]
- Deniskova, T.E.; Dotsev, A.V.; Selionova, M.I.; Aibazov, A.-M.M.; Zinovieva, N.A. Search for signatures of selection in the genomes of domestic goats (Capra hircus L.) raised in Russia using detection of ROH islands. Sel’skokhozyaistvennaya Biol. [Agric. Biol.] 2024, 59, 620–632. [Google Scholar] [CrossRef]
- Becker, R.A.; Wilks, A.R.; Brownrigg, R.; Minka, T.P.; Deckmyn, A. Maps: Draw Geographical Maps. R Package Version 3.3.0. 2018. Available online: https://CRAN.R-project.org/package=maps (accessed on 25 April 2025).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; ISBN 978-0-387-98141-3. [Google Scholar] [CrossRef]
- Vasimuddin, M.; Misra, S.; Li, H.; Aluru, S. Efficient Architecture-aware Acceleration of BWAMEM for Multicore Systems. In Proceedings of the 2019 IEEE International Parallel and Distributed Processing Symposium (IPDPS), Rio de Janeiro, Brazil, 20–24 May 2019; IEEE Xplore: Piscataway, NJ, USA, 2019; pp. 314–324. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Biscarini, F.; Paolo Cozzi, P.; Gaspa, G.; Marras, G. detectRUNS: Detect Runs of Homozygosity and Runs of Heterozygosity in Diploid Genomes; R Package Version 0.9.6; The Comprehensive R Archive Network (CRAN); Institute for Statistics and Mathematics, Vienna University of Economics and Business: Vienna, Austria, 2019; Available online: https://cran.r-project.org/web/packages/detectRUNS/index.html (accessed on 25 April 2025).
- Romanov, M.N.; Sölkner, J.; Zinovieva, N.A.; Wimmers, K.; Weigend, S. Editorial: Traditional and up-to-date genomic insights into domestic animal diversity. Front. Genet. 2023, 13, 1117708. [Google Scholar] [CrossRef] [PubMed]
- Moiseyeva, I.G. Otechestvennyye porody kur [Native breeds of domestic fowl]. In Geneticheskie Nesursy Sel’skokhozyajstvennykh Zhivothykh: Redkie I Ischezayushchie Otechestvennye Porody [Farm Animal Genetic Resources: Rare and Endangered Native Breeds]; Moiseyeva, I.G., Zakharov, I.A., Mitichashvili, R.S., Eds.; Nauka: Moscow, Russia, 1992; pp. 11–112. Available online: https://agris.fao.org/search/en/providers/122621/records/647396783ed73003714cc15a (accessed on 25 April 2025). (In Russian)
- Dehghanzadeh, H.; Mirhosseini, S.Z.; Romanov, M.N.; Ghorbani, A. Evaluation of genetic variability and distances among five Iranian native chicken populations using RAPD markers. Pak. J. Biol. Sci. 2009, 12, 866–871. [Google Scholar] [CrossRef]
- Larkina, T.A.; Barkova, O.Y.; Peglivanyan, G.K.; Mitrofanova, O.V.; Dementieva, N.V.; Stanishevskaya, O.I.; Vakhrameev, A.B.; Makarova, A.V.; Shcherbakov, Y.S.; Pozovnikova, M.V.; et al. Evolutionary subdivision of domestic chickens: Implications for local breeds as assessed by phenotype and genotype in comparison to commercial and fancy breeds. Agriculture 2021, 11, 914. [Google Scholar] [CrossRef]
- Kulibaba, R.O.; Sakhatskyi, M.I.; Griffin, D.K.; Romanov, M.N. Molecular diversity of Ukrainian native chicken breeds: A review. Worlds Poult. Sci. J. 2024, 80, 1265–1292. [Google Scholar] [CrossRef]
- Bertolini, F.; Cardoso, T.F.; Marras, G.; Nicolazzi, E.L.; Rothschild, M.F.; Amills, M.; AdaptMap consortium. Genome-wide patterns of homozygosity provide clues about the population history and adaptation of goats. Genet. Sel. Evol. 2018, 50, 59. [Google Scholar] [CrossRef]
- Wang, R.; Wang, X.; Qi, Y.; Li, Y.; Na, Q.; Yuan, H.; Rong, Y.; Ao, X.; Guo, F.; Zhang, L.; et al. Genetic diversity analysis of Inner Mongolia cashmere goats (Erlangshan subtype) based on whole genome re-sequencing. BMC Genom. 2024, 25, 698. [Google Scholar] [CrossRef]
- An, Z.X.; Shi, L.G.; Hou, G.Y.; Zhou, H.L.; Xun, W.J. Genetic diversity and selection signatures in Hainan black goats revealed by whole-genome sequencing data. Animal 2024, 18, 101147. [Google Scholar] [CrossRef]
- Peng, W.; Zhang, Y.; Gao, L.; Wang, S.; Liu, M.; Sun, E.; Lu, K.; Zhang, Y.; Li, B.; Li, G.; et al. Investigation of selection signatures of dairy goats using whole-genome sequencing data. BMC Genom. 2025, 26, 234. [Google Scholar] [CrossRef]
- Curik, I.; Ferencakovic, M.; Sölkner, J. Inbreeding and runs of homozygosity: A possible solution to an old problem. Livest. Sci. 2014, 166, 26–34. [Google Scholar] [CrossRef]
- Bian, C.; Luo, Y.; Li, J.; Cheng, H.; He, F.; Duan, H.; Ahmed, Z.; Lei, C.; Yi, K. Inference of genetic diversity, population structure, and selection signatures in Xiangxi white buffalo of China through whole-genome resequencing. Genes 2024, 15, 1450. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhong, J.; Li, L.; Zhong, T.; Wang, L.; Song, T.; Zhang, H. Comparative genome analyses reveal the unique genetic composition and selection signals underlying the phenotypic characteristics of three Chinese domestic goat breeds. Genet. Sel. Evol. 2019, 51, 70. [Google Scholar] [CrossRef]
- Wan, X.; Jing, J.N.; Wang, D.F.; Lv, F.H. Whole-genome selective scans detect genes associated with important phenotypic traits in goat (Capra hircus). Front. Genet. 2023, 14, 1173017. [Google Scholar] [CrossRef]
- Saif, R.; Mahmood, T.; Zia, S.; Henkel, J.; Ejaz, A. Genomic selection pressure discovery using site-frequency spectrum and reduced local variability statistics in Pakistani Dera-Din-Panah goat. Trop. Anim. Health Prod. 2023, 55, 331. [Google Scholar] [CrossRef]
- Snelling, W.M.; Allan, M.F.; Keele, J.W.; Kuehn, L.A.; Thallman, R.M.; Bennett, G.L.; Ferrell, C.L.; Jenkins, T.G.; Freetly, H.C.; Nielsen, M.K.; et al. Partial-genome evaluation of postweaning feed intake and efficiency of crossbred beef cattle. J. Anim. Sci. 2011, 89, 1731–1741. [Google Scholar] [CrossRef]
- Lindholm-Perry, A.K.; Kern, R.J.; Kuehn, L.A.; Snelling, W.M.; Miles, J.R.; Oliver, W.T.; Freetly, H.C. Differences in transcript abundance of genes on BTA15 located within a region associated with gain in beef steers. Gene 2015, 572, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Wang, X.; Li, M.; Li, Y.; Yang, Z.; Wang, X.; Pan, X.; Gong, M.; Zhang, Y.; Guo, Y.; et al. The origin of domestication genes in goats. Sci. Adv. 2020, 6, eaaz5216. [Google Scholar] [CrossRef]
- Kim, E.S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.; Sayre, B.; Mwacharo, J.M.; Rothschild, M.F. Multiple genomic signatures of selection in goats and sheep indigenous to a hot arid environment. Heredity 2016, 116, 255–264. [Google Scholar] [CrossRef]
- Dadousis, C.; Cecchi, F.; Ablondi, M.; Fabbri, M.C.; Stella, A.; Bozzi, R. Keep Garfagnina alive. An integrated study on patterns of homozygosity, genomic inbreeding, admixture and breed traceability of the Italian Garfagnina goat breed. PLoS ONE 2021, 16, e0232436. [Google Scholar] [CrossRef]
- Pegolo, S.; Bisutti, V.; Mota, L.F.M.; Cecchinato, A.; Amalfitano, N.; Dettori, M.L.; Pazzola, M.; Vacca, G.M.; Bittante, G. Genome-wide landscape of genetic diversity, runs of homozygosity, and runs of heterozygosity in five Alpine and Mediterranean goat breeds. J. Anim. Sci. Biotechnol. 2025, 16, 33. [Google Scholar] [CrossRef] [PubMed]
- Graham-Paquin, A.L.; Saini, D.; Sirois, J.; Hossain, I.; Katz, M.S.; Zhuang, Q.K.; Kwon, S.Y.; Yamanaka, Y.; Bourque, G.; Bouchard, M.; et al. ZMYM2 is essential for methylation of germline genes and active transposons in embryonic development. Nucleic Acids Res. 2023, 51, 7314–7329. [Google Scholar] [CrossRef] [PubMed]
- Bo, D.; Jiang, X.; Liu, G.; Xu, F.; Hu, R.; Wassie, T.; Chong, Y.; Ahmed, S.; Liu, C.; Girmay, S. Multipathway synergy promotes testicular transition from growth to spermatogenesis in early-puberty goats. BMC Genom. 2020, 21, 372. [Google Scholar] [CrossRef]
- Serranito, B.; Cavalazzi, M.; Vidal, P.; Taurisson-Mouret, D.; Ciani, E.; Bal, M.; Rouvellac, E.; Servin, B.; Moreno-Romieux, C.; Tosser-Klopp, G.; et al. Local adaptations of Mediterranean sheep and goats through an integrative approach. Sci. Rep. 2021, 11, 21363. [Google Scholar] [CrossRef]
- Deniskova, T.E.; Dotsev, A.V.; Zinovieva, N.A. [Identification of candidate genes associated with economically significant traits based on the analysis of homozygosity islands in the genome of sheep breeds bred in Russia]. Dostizheniya Nauki I Tekhniki APK [Achiev. Sci. Technol. AIC] 2023, 37, 80–86, (In Russian with English summary). [Google Scholar]
- Pandya, A.; Arnos, K.S.; Xia, X.J.; Welch, K.O.; Blanton, S.H.; Friedman, T.B.; Garcia Sanchez, G.; Liu, X.Z.; Morell, R.; Nance, W.E. Frequency and distribution of GJB2 (connexin 26) and GJB6 (connexin 30) mutations in a large North American repository of deaf probands. Genet. Med. 2003, 5, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Onzima, R.B.; Upadhyay, M.R.; Doekes, H.P.; Brito, L.F.; Bosse, M.; Kanis, E.; Groenen, M.A.M.; Crooijmans, R.P.M.A. Genome-wide characterization of selection signatures and runs of homozygosity in Ugandan goat breeds. Front. Genet. 2018, 9, 318. [Google Scholar] [CrossRef]
- Li, G.; Tang, J.; Huang, J.; Jiang, Y.; Fan, Y.; Wang, X.; Ren, J. Genome-wide estimates of runs of homozygosity, heterozygosity, and genetic load in two Chinese indigenous goat breeds. Front. Genet. 2022, 13, 774196. [Google Scholar] [CrossRef]
- Abied, A.; Xu, L.; Sahlu, B.W.; Xing, F.; Ahbara, A.; Pu, Y.; Lin, J.; Berihulay, H.; Islam, R.; He, X.; et al. Genome-wide analysis revealed homozygosity and demographic history of five Chinese sheep breeds adapted to different environments. Genes 2020, 11, 1480. [Google Scholar] [CrossRef]
- Liu, J.; Shi, L.; Li, Y.; Chen, L.; Garrick, D.; Wang, L.; Zhao, F. Estimates of genomic inbreeding and identification of candidate regions that differ between Chinese indigenous sheep breeds. J. Anim. Sci. Biotechnol. 2021, 12, 95. [Google Scholar] [CrossRef]
- Quan, K.; Shi, H.; Wei, C.; Li, J.; Liu, K.; Wang, H.; Sun, W.; Han, H. Genetic diversity, reproductive performance, and genetic enhancement strategies in Huang-Huai goats. Front. Genet. 2025, 16, 1549051. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, S.; Qiao, L.; Zhang, S.; Liu, Q.; Yang, K.; Pan, Y.; Liu, J.; Liu, W. Four markers useful for the distinction of intrauterine growth restriction in sheep. Animals 2023, 13, 3305. [Google Scholar] [CrossRef] [PubMed]
- Lakhssassi, K.; Sarto, M.P.; Lahoz, B.; Alabart, J.L.; Folch, J.; Serrano, M.; Calvo, J.H. Blood transcriptome of Rasa Aragonesa rams with different sexual behavior phenotype reveals CRYL1 and SORCS2 as genes associated with this trait. J. Anim. Sci. 2023, 101, skad098. [Google Scholar] [CrossRef]
- Fu, X.; Zhao, B.; Tian, K.; Wu, Y.; Suo, L.; Ba, G.; Ciren, D.; De, J.; Awang, C.; Gun, S.; et al. Integrated analysis of lncRNA and mRNA reveals novel insights into cashmere fineness in Tibetan cash-mere goats. PeerJ 2020, 8, e10217. [Google Scholar] [CrossRef]
- Bai, Z.X.; Xu, Y.N.; Gu, M.; Cai, W.; Zhang, Y.; Qin, Y.; Chen, R.; Sun, Y.; Wu, Y.; Wang, Z. Proteomic analysis of coarse and fine skin tissues of Liaoning cashmere goat. Funct. Integr. Genom. 2022, 22, 503–513. [Google Scholar] [CrossRef]
- Liu, Z.; Li, H.; Luo, Y.; Li, J.; Sun, A.; Ahmed, Z.; Zhang, B.; Lei, C.; Yi, K. Comprehensive whole-genome resequencing unveils genetic diversity and selective signatures of the Xiangdong black goat. Front. Genet. 2024, 15, 1326828. [Google Scholar] [CrossRef] [PubMed]
- San Agustin, J.T.; Pazour, G.J.; Witman, G.B. Intraflagellar transport is essential for mammalian spermiogenesis but is absent in mature sperm. Mol. Biol. Cell 2015, 26, 4358–4372. [Google Scholar] [CrossRef]
- Wei, X.; Wang, X.; Yang, C.; Gao, Y.; Zhang, Y.; Xiao, Y.; Ju, Z.; Jiang, Q.; Wang, J.; Liu, W.; et al. CFAP58 is involved in the sperm head shaping and flagellogenesis of cattle and mice. Development 2024, 151, dev202608. [Google Scholar] [CrossRef]
- Yurchenko, A.A.; Deniskova, T.E.; Yudin, N.S.; Dotsev, A.V.; Khamiruev, T.N.; Selionova, M.I.; Egorov, S.V.; Reyer, H.; Wimmers, K.; Brem, G.; et al. High-density genotyping reveals signatures of selection related to acclimation and economically important traits in 15 local sheep breeds from Russia. BMC Genom. 2019, 20 (Suppl. 3), 294. [Google Scholar] [CrossRef]
- Elder, L.A.; Hinnant, H.R.; Mandella, C.M.; Claus-Walker, R.A.; Parrish, L.M.; Slanzon, G.S.; McConnel, C.S. Differential gene expression in peripheral leukocytes of pre-weaned Holstein heifer calves with respiratory disease. PLoS ONE 2023, 18, e0285876. [Google Scholar] [CrossRef]
- Chen, Z.; Luo, J.; Zhang, C.; Ma, Y.; Sun, S.; Zhang, T.; Loo, J.J. Mechanism of prolactin inhibition of miR-135b via methylation in goat mammary epithelial cells. J. Cell. Physiol. 2018, 233, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.M.; Zhang, T.T.; An, S.Y.; El-Samahy, M.A.; Yang, H.; Wan, Y.J.; Meng, F.X.; Xiao, S.H.; Wang, F.; Lei, Z.H. Expression of Hippo signaling pathway components in Hu sheep male reproductive tract and spermatozoa. Theriogenology 2019, 126, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Plewes, M.R.; Hou, X.; Zhang, P.; Liang, A.; Hua, G.; Wood, J.R.; Cupp, A.S.; Lv, X.; Wang, C.; Davis, J.S. Yes-associated protein 1 is required for proliferation and function of bovine granulosa cells in vitro. Biol. Reprod. 2019, 101, 1001–1017. [Google Scholar] [CrossRef] [PubMed]
- E, G.X.; Zhu, Y.B.; Basang, W.D.; Na, R.S.; Han, Y.G.; Zeng, Y. Comparative and selection sweep analysis of CNV was associated to litter size in Dazu black goats. Anim. Biotechnol. 2021, 32, 792–797. [Google Scholar] [CrossRef]
- Mahar, K.; Gurao, A.; Kumar, A.; Pratap Singh, L.; Chitkara, M.; Gowane, G.R.; Ahlawat, S.; Niranjan, S.K.; Pundir, R.K.; Kataria, R.S.; et al. Genomic inbreeding analysis reveals resilience and genetic diversity in Indian yak populations. Gene 2024, 928, 148787. [Google Scholar] [CrossRef]
- Workman, A.M.; Heaton, M.P.; Webster, D.A.; Harhay, G.P.; Kalbfleisch, T.S.; Smith, T.P.L.; Falkenberg, S.M.; Carlson, D.F.; Sonstegard, T.S. Evaluating large spontaneous deletions in a bovine cell line selected for bovine viral diarrhea virus resistance. Viruses 2021, 13, 2147. [Google Scholar] [CrossRef]
- Zhao, Q.; Huang, C.; Chen, Q.; Su, Y.; Zhang, Y.; Wang, R.; Su, R.; Xu, H.; Liu, S.; Ma, Y.; et al. Genomic inbreeding and runs of homozygosity analysis of cashmere goat. Animals 2024, 14, 1246. [Google Scholar] [CrossRef]
- Salilew-Wondim, D.; Hölker, M.; Rings, F.; Phatsara, C.; Mohammadi-Sangcheshmeh, A.; Tholen, E.; Schellander, K.; Tesfaye, D. Depletion of BIRC6 leads to retarded bovine early embryonic development and blastocyst formation in vitro. Reprod. Fertil. Dev. 2010, 22, 564–579. [Google Scholar] [CrossRef]
- Guan, D.; Luo, N.; Tan, X.; Zhao, Z.; Huang, Y.; Na, R.; Zhang, J.; Zhao, Y. Scanning of selection signature provides a glimpse into important economic traits in goats (Capra hircus). Sci. Rep. 2016, 6, 36372. [Google Scholar] [CrossRef]
- El-Halawany, N.; Zhou, X.; Al-Tohamy, A.F.; El-Sayd, Y.A.; Shawky, A.E.; Michal, J.J.; Jiang, Z. Genome-wide screening of candidate genes for improving fertility in Egyptian native Rahmani sheep. Anim. Genet. 2016, 47, 513. [Google Scholar] [CrossRef]
- Kour, A.; Deb, S.M.; Nayee, N.; Niranjan, S.K.; Raina, V.S.; Mukherjee, A.; Patil, C.S. Novel insights into genome-wide associations in Bos indicus reveal genetic linkages between fertility and growth. Anim. Biotechnol. 2021, 34, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Serão, N.V.; González-Peña, D.; Beever, J.E.; Bollero, G.A.; Southey, B.R.; Faulkner, D.B.; Rodriguez-Zas, S.L. Bivariate genome-wide association analysis of the growth and intake components of feed efficiency. PLoS ONE 2013, 8, e78530. [Google Scholar] [CrossRef]
- Sarkar, J.; Balamurugan, V.; Sen, A.; Saravanan, P.; Sahay, B.; Rajak, K.K.; Rasool, T.J.; Bhanuprakash, V.; Singh, R.K. Sequence analysis of morbillivirus CD150 receptor-signaling lymphocyte activation molecule (SLAM) of different animal species. Virus Genes 2009, 39, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Wang, T.; Li, Z.; Wan, Y.; Yang, B.; Zeng, W.; Zhang, Y.; Wang, J. MicroRNA-218 Regulates signaling lymphocyte activation molecular (SLAM) mediated peste des petits ruminants virus infectivity in goat peripheral blood mononuclear cells. Front. Immunol. 2019, 10, 2201. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, G.; Lin, X.; Zhang, J.; Hou, G.; Zhang, L.; Liu, D.; Li, Y.; Li, J.; Xu, L. Genomic inbreeding and runs of homozygosity analysis of indigenous cattle populations in southern China. PLoS ONE 2022, 17, e0271718. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, C.; Kamalibieke, J.; Gong, P.; Mu, Y.; Zhu, L.; Lv, X.; Wang, W.; Luo, J. Whole genome and transcriptome analyses in dairy goats identify genetic markers associated with high milk yield. Int. J. Biol. Macromol. 2025, 292, 139192. [Google Scholar] [CrossRef]
- Xu, Q.; Xi, Y.; Ma, S.; Wang, J.; Li, J.; Han, C.; Li, L.; Wang, J.; Liu, H. Transcriptome profiling of morphogenetic differences between contour and flight feathers in duck. Br. Poult. Sci. 2022, 63, 597–604. [Google Scholar] [CrossRef]
- Zhang, W.; Li, P.; Wang, S.; Cheng, G.; Wang, L.; Mi, X.; Su, X.; Wang, Y.; Zan, L. TP53INP2 promotes bovine adipocytes differentiation through autophagy activation. Animals 2019, 9, 1060. [Google Scholar] [CrossRef]
- Sabaté-Pérez, A.; Romero, M.; Sànchez-Fernàndez-de-Landa, P.; Carobbio, S.; Mouratidis, M.; Sala, D.; Engel, P.; Martínez-Cristóbal, P.; Villena, J.A.; Virtue, S.; et al. Autophagy-mediated NCOR1 degradation is required for brown fat maturation and thermogenesis. Autophagy 2023, 19, 904–925. [Google Scholar] [CrossRef]
- Sala, D.; Ivanova, S.; Plana, N.; Ribas, V.; Duran, J.; Bach, D.; Turkseven, S.; Laville, M.; Vidal, H.; Karczewska-Kupczewska, M.; et al. Autophagy-regulating TP53INP2 mediates muscle wasting and is repressed in diabetes. J. Clin. Investig. 2014, 124, 1914–1927. [Google Scholar] [CrossRef]
- Verbrugge, S.A.J.; Schönfelder, M.; Becker, L.; Yaghoob Nezhad, F.; Hrabě de Angelis, M.; Wackerhage, H. Genes whose gain or loss-of-function increases skeletal muscle mass in mice: A systematic literature review. Front. Physiol. 2018, 9, 553. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.N.; Raudsepp, T.; Alshanbari, F.; Gutiérrez, G.; Ponce de León, F.A. Chromosomal localization of candidate genes for fiber growth and color in alpaca (Vicugna pacos). Front. Genet. 2019, 10, 583. [Google Scholar] [CrossRef] [PubMed]
- Olsen, H.G.; Knutsen, T.M.; Kohler, A.; Svendsen, M.; Gidskehaug, L.; Grove, H.; Nome, T.; Sodeland, M.; Sundsaasen, K.K.; Kent, M.P.; et al. Genome-wide association mapping for milk fat composition and fine mapping of a QTL for de novo synthesis of milk fatty acids on bovine chromosome 13. Genet. Sel. Evol. 2017, 49, 20. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, T.M.; Olsen, H.G.; Tafintseva, V.; Svendsen, M.; Kohler, A.; Kent, M.P.; Lien, S. Unravelling genetic variation underlying de novo-synthesis of bovine milk fatty acids. Sci. Rep. 2018, 8, 2179. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | n | ROH Length | ROH Number | FROH | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SE | Min | Max | Mean ± SE | Min | Max | Mean ± SE | Min | Max | ||
| Orenburg | 19 | 78.43 ± 3.66 | 47.57 | 102.96 | 438.79 ± 16.83 | 283 | 540 | 0.032 ± 0.001 | 0.02 | 0.04 |
| Karachay | 20 | 134.13 ± 18.79 | 30.38 | 404.40 | 695.25 ± 94.20 | 189 | 2075 | 0.054 ± 0.008 | 0.01 | 0.16 |
| Breed | CHI | Start, bp | End, bp | No. of SNPs | Gene |
|---|---|---|---|---|---|
| Orenburg | 3 | 110,894,406 | 110,897,532 | 9 | |
| 11 | 14,760,399 | 15,261,320 | 1358 | YIPF4, BIRC6, TTC27 | |
| 12 | 50,342,201 | 50,432,910 | 168 | PARP4, MPHOSPH8 | |
| 12 | 50,530,724 | 50,533,436 | 10 | ||
| 12 | 50,671,450 | 50,805,766 | 459 | GJB2, GJB6, CRYL1 | |
| 12 | 50,809,376 | 50,947,946 | 425 | IFT88, IL17D, EEF1AKMT1 | |
| 12 | 50,948,756 | 50,965,731 | 50 | ||
| 12 | 50,968,611 | 51,033,025 | 133 | XPO4, LATS2 | |
| 12 | 51,035,319 | 51,155,729 | 386 | SAP18 | |
| 15 | 32,124,217 | 32,379,590 | 394 | STIM1, RRM1 | |
| 16 | 54,524,824 | 54,531,631 | 20 | RABGAP1L | |
| Karachay | 3 | 110,862,077 | 110,964,407 | 226 | SLAMF1 |
| 5 | 47,322,478 | 47,471,119 | 626 | ||
| 7 | 60,029,670 | 60,088,713 | 184 | SIL1 | |
| 7 | 60,090,256 | 60,138,223 | 128 | ||
| 12 | 50,286,354 | 50,432,910 | 292 | PARP4, MPHOSPH8 | |
| 12 | 50,530,724 | 50,537,889 | 24 | ||
| 12 | 50,543,508 | 50,573,061 | 41 | ZMYM2 | |
| 13 | 53,173,841 | 53,239,419 | 116 | PRPF6, SAMD10, ZNF512B, UCKL1 | |
| 13 | 62,528,703 | 62,629,819 | 327 | ||
| 13 | 63,614,834 | 63,737,342 | 398 | TP53INP2, NCOA6, GGT7 | |
| 15 | 32,188,813 | 32,383,957 | 400 | STIM1, RRM1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deniskova, T.E.; Dotsev, A.V.; Koshkina, O.A.; Solovieva, A.D.; Churbakova, N.A.; Petrov, S.N.; Frolov, A.N.; Platonov, S.A.; Abdelmanova, A.S.; Vladimirov, M.A.; et al. Examination of Runs of Homozygosity Distribution Patterns and Relevant Candidate Genes of Potential Economic Interest in Russian Goat Breeds Using Whole-Genome Sequencing. Genes 2025, 16, 631. https://doi.org/10.3390/genes16060631
Deniskova TE, Dotsev AV, Koshkina OA, Solovieva AD, Churbakova NA, Petrov SN, Frolov AN, Platonov SA, Abdelmanova AS, Vladimirov MA, et al. Examination of Runs of Homozygosity Distribution Patterns and Relevant Candidate Genes of Potential Economic Interest in Russian Goat Breeds Using Whole-Genome Sequencing. Genes. 2025; 16(6):631. https://doi.org/10.3390/genes16060631
Chicago/Turabian StyleDeniskova, Tatiana E., Arsen V. Dotsev, Olga A. Koshkina, Anastasia D. Solovieva, Nadezhda A. Churbakova, Sergey N. Petrov, Alexey N. Frolov, Stanislav A. Platonov, Alexandra S. Abdelmanova, Maxim A. Vladimirov, and et al. 2025. "Examination of Runs of Homozygosity Distribution Patterns and Relevant Candidate Genes of Potential Economic Interest in Russian Goat Breeds Using Whole-Genome Sequencing" Genes 16, no. 6: 631. https://doi.org/10.3390/genes16060631
APA StyleDeniskova, T. E., Dotsev, A. V., Koshkina, O. A., Solovieva, A. D., Churbakova, N. A., Petrov, S. N., Frolov, A. N., Platonov, S. A., Abdelmanova, A. S., Vladimirov, M. A., Gladyr, E. A., Gusev, I. V., Lebedev, S. V., Griffin, D. K., Romanov, M. N., & Zinovieva, N. A. (2025). Examination of Runs of Homozygosity Distribution Patterns and Relevant Candidate Genes of Potential Economic Interest in Russian Goat Breeds Using Whole-Genome Sequencing. Genes, 16(6), 631. https://doi.org/10.3390/genes16060631