
RiceReceptor: The Cell-Surface and Intracellular Immune Receptors of the Oryza Genus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Organization of Rice Genome Information
2.2. Identification of LRR Genes
2.3. Identification of NLR Genes
2.4. Construction of the Phylogenetic Tree for 300 Rice Genomes
2.5. Data Analysis and Visualization
3. Results
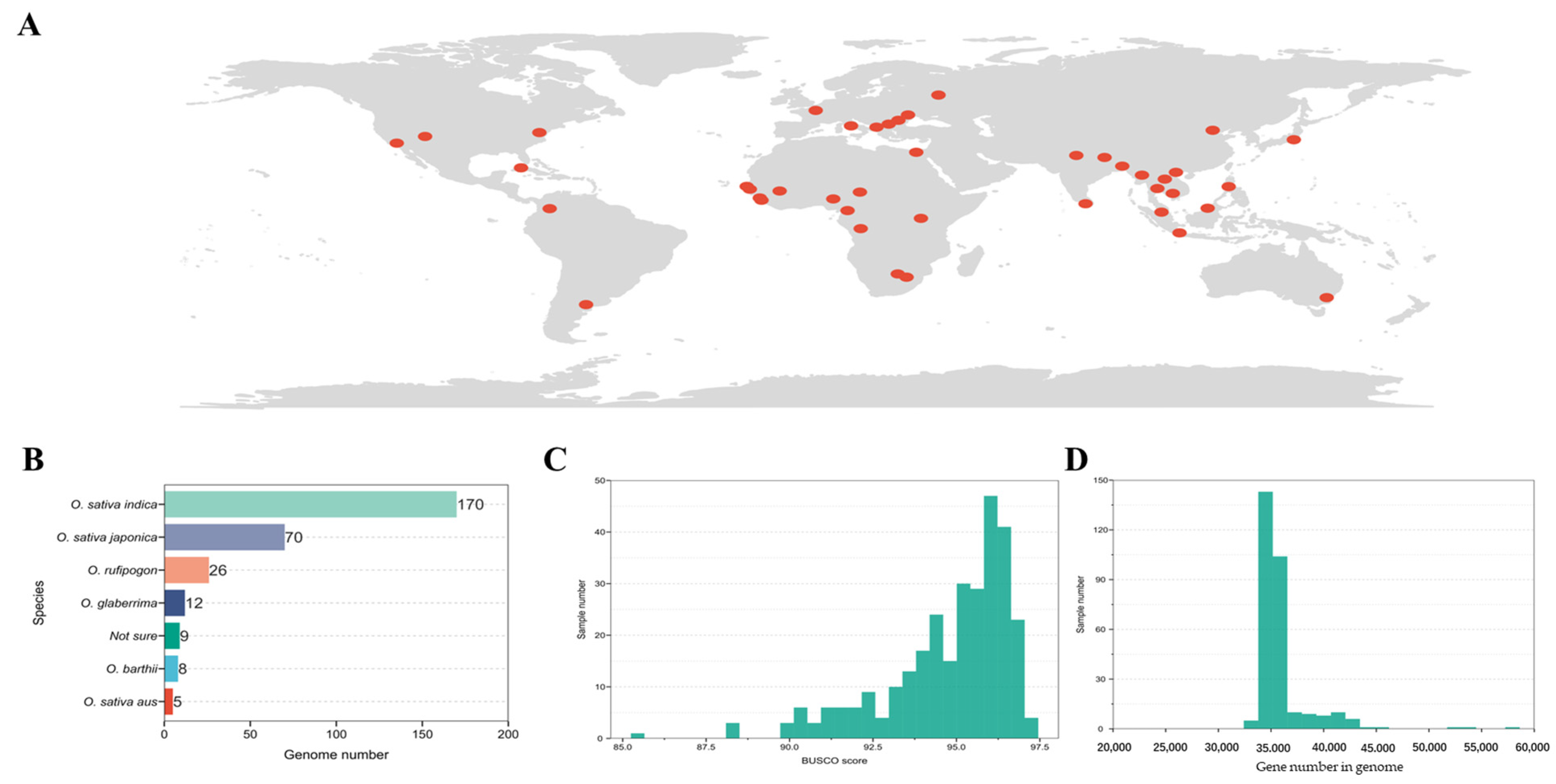
3.1. Collection and Analysis of Rice Genome Samples
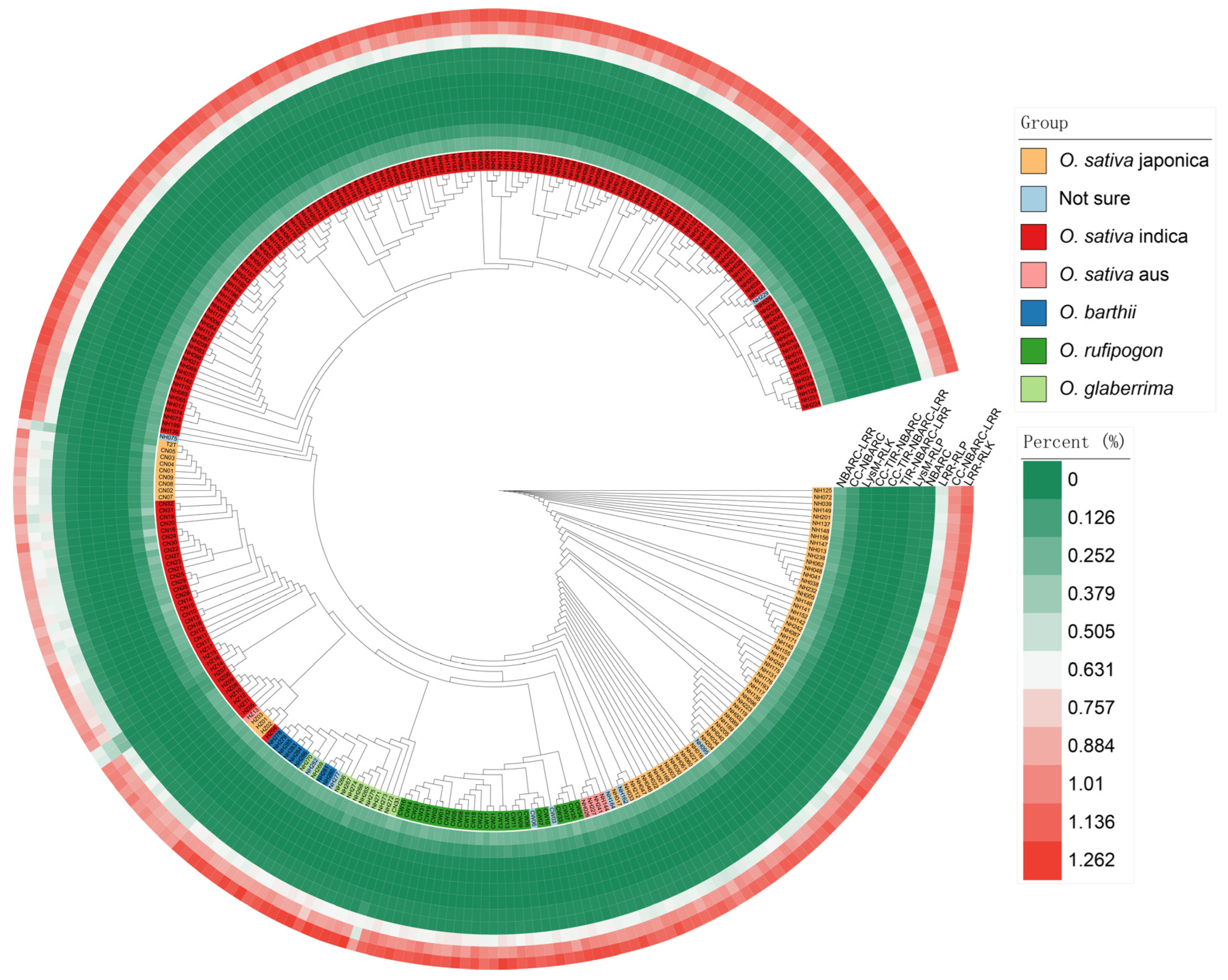
3.2. Phylogenetic Tree and the Number of Each Gene Type
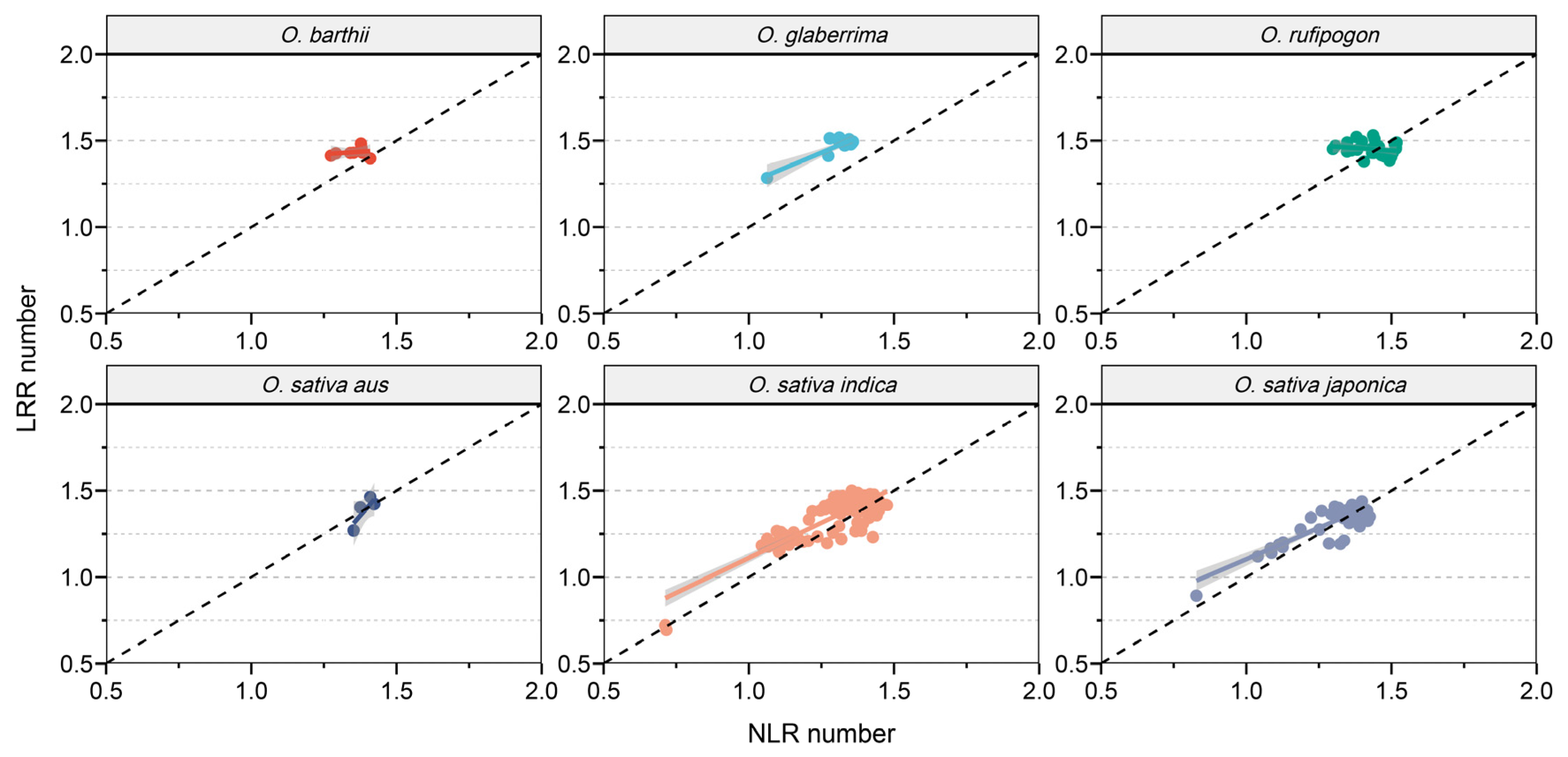
3.3. Correlation Between the Sizes of Immune Receptor Families
3.4. Potential TNL Genes in Rice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qin, P.; Lu, H.; Du, H.; Wang, H.; Chen, W.; Chen, Z.; He, Q.; Ou, S.; Zhang, H.; Li, X.; et al. Pan-genome analysis of 33 genetically diverse rice accessions reveals hidden genomic variations. Cell 2021, 184, 3542–3558.e16. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Li, X.; He, H.; Yuan, Q.; Song, Y.; Wei, Z.; Lin, H.; Hu, M.; Zhao, F.; Zhang, C.; et al. A super pan-genomic landscape of rice. Cell Res. 2022, 32, 878–896. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Chen, Y.; Zhou, Y.; Zhang, Y.; Li, M.; Ouyang, Y.; Chebotarov, D.; Mauleon, R.; Zhao, H.; Xie, W.; et al. Rice gene index: A comprehensive pan-genome database for comparative and functional genomics of Asian rice. Mol. Plant 2023, 16, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.F.; Ullah, H.; Aung, M.Z.; Tisarum, R.; Cha-Um, S.; Datta, A. Iron toxicity tolerance of rice genotypes in relation to growth, yield and physiochemical characters. Rice Sci. 2023, 30, 321–334. [Google Scholar] [CrossRef]
- Cai, J.; Ma, E.; Lin, J.; Liao, L.; Han, Y. Exploring global food security pattern from the perspective of spatio-temporal evolution. J. Geogr. Sci. 2020, 30, 179–196. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Saijo, Y.; Loo, E.P.-I. Plant immunity in signal integration between biotic and abiotic stress responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef]
- Fiyaz, R.A.; Shivani, D.; Chaithanya, K.; Mounika, K.; Chiranjeevi, M.; Laha, G.S.; Viraktamath, B.C.; Subba Rao, L.V.; Sundaram, R.M. Genetic improvement of rice for bacterial blight resistance: Present status and future pospects. Rice Sci. 2022, 29, 118–132. [Google Scholar] [CrossRef]
- He, X.; Li, Z.; Guo, S.; Zheng, X.; Liu, C.; Liu, Z.; Li, Y.; Yuan, Z.; Li, L. Epistasis-aware genome-wide association studies provide insights into theefficient breeding of high-yield and high-quality rice. J. Integr. Agric. 2024, 23, 2541–2556. [Google Scholar] [CrossRef]
- Ning, X.; Yunyu, W.; Aihong, L. Strategy for use of rice blast resistance genes in rice molecular breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar] [CrossRef]
- Kesel, J.; Gómez-Rodríguez, R.; Bonneure, E.; Mangelinckx, S.; Kyndt, T. The use of pti-marker genes to identify novel compounds that establish induced resistance in rice. Int. J. Mol. Sci. 2020, 21, 317. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. Pti-Eti crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ning, Y.; Shi, X.; He, F.; Zhang, C.; Fan, J.; Jiang, N.; Zhang, Y.; Zhang, T.; Hu, Y.; et al. Immunity to rice blast disease by suppression of effector-triggered necrosis. Curr. Biol. 2016, 26, 2399–2411. [Google Scholar] [CrossRef] [PubMed]
- Varden, F.A.; Saitoh, H.; Yoshino, K.; Franceschetti, M.; Kamoun, S.; Terauchi, R.; Banfield, M.J. Cross-reactivity of a rice NLR immune receptor to distinct effectors from the rice blast pathogen Magnaporthe oryzae provides partial disease resistance. J. Biol. Chem. 2019, 294, 13006–13016. [Google Scholar] [CrossRef]
- Su, J.; Chen, S.; Zhu, X. Research progress in molecular mechanism of broad-spectrum disease resistance in rice. Guangdong Agric. Sci. 2020, 47, 84–92. [Google Scholar] [CrossRef]
- Sun, C.; Hu, Z.; Zheng, T.; Lu, K.; Zhao, Y.; Wang, W.; Shi, J.; Wang, C.; Lu, J.; Zhang, D.; et al. RPAN: Rice pan-genome browser for ∼3000 rice genomes. Nucleic Acids Res. 2017, 45, 597–605. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3,010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef]
- Huang, C.; Chen, Z.; Liang, C. Oryza pan-genomics: A new foundation for future rice research and improvement. Crop J. 2021, 9, 622–632. [Google Scholar] [CrossRef]
- Li, J.; Meng, L.; Hou, B.; Hao, X.; Wang, D.; Qu, L.; Li, Z.; Zhang, L.; Jin, X. Detection of novel BEST1 variations in autosomal recessive bestrophinopathy using third-generation sequencing. Curr. Med. Sci. 2024, 44, 419–425. [Google Scholar] [CrossRef]
- Gamaarachchi, H.; Bayat, A.; Gaeta, B.; Parameswaran, S. Cache friendly optimisation of de bruijn graph based local re-assembly in variant calling. IEEE/ACM Trans. Comput. Biol. Bioinform. 2020, 17, 1125–1133. [Google Scholar] [CrossRef]
- Liu, W.; Wang, G. Plant innate immunity in rice: A defense against pathogen infection. Natl. Sci. Rev. 2016, 3, 295–308. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, W.; Li, L.; Francis, F.; Wang, X. Transcriptome analysis reveals different response of resistant and susceptible rice varieties to rice stripe virus infection. J. Integr. Agric. 2023, 22, 1750–1762. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, C.; Zhu, D.; He, H.; Wei, Z.; Yuan, Q.; Li, X.; Gao, X.; Zhang, B.; Gao, H.; et al. Identifying candidate genes and patterns of heat-stress response in rice using a genome-wide association study and transcriptome analyses. Crop J. 2022, 10, 1633–1643. [Google Scholar] [CrossRef]
- Qi, T.; Seong, K.; Thomazella, D.P.T.; Kim, J.R.; Pham, J.; Seo, E.; Cho, M.; Schultink, A.; Staskawicz, B.J. NRG1 functions downstream of EDS1 to regulate TIR-NLR-mediated plant immunity in Nicotiana benthamiana. Proc. Natl. Acad. Sci. USA 2018, 115, E10979–E10987. [Google Scholar] [CrossRef]
- Hong, X.; Li, S.; Cheng, X.; Zhi, H.; Yin, J.; Xu, K. Searching for plant NLR immune receptors conferring resistance to potyviruses. Crop J. 2024, 12, 28–44. [Google Scholar] [CrossRef]
- Pertea, G.; Pertea, M. GFF Utilities: Gffread and gffcompare [version 2; peer review: 3 approved]. F1000Research 2020, 9, 304. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Heal, R.; Wyler, M.; Schmid, M.W.; Jones, J.D.G. Concerted expansion and contraction of immune receptor gene repertoires in plant genomes. Nat. Plants 2022, 8, 1146–1152. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Steuernagel, B.; Witek, K.; Krattinger, S.G.; Ramirez-Gonzalez, R.H.; Schoonbeek, H.-J.; Yu, G.; Baggs, E.; Witek, A.I.; Yadav, I.; Krasileva, K.V.; et al. The NLR-annotator tool enables annotation of the intracellular immune receptor repertoire1 [OPEN]. Plant Physiol. 2020, 183, 468–482. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Xu, K.; Zhao, F.; Liu, W.; Li, L.; Hua, Z.; Zhou, X. Itol.toolkit accelerates working with iTOL (Interactive Tree of Life) by an automated generation of annotation files. Bioinformatics 2023, 39, btad339. [Google Scholar] [CrossRef] [PubMed]
- Ihaka, R.; Gentleman, R. R: A Language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Li, X.; Ma, L.; Wang, Y.; Ye, C.; Guo, C.; Li, Y.; Mei, X.; Du, F.; Huang, H. PlantNLRatlas: A comprehensive dataset of full- and partial-length NLR resistance genes across 100 chromosome-level plant genomes. Front. Plant Sci. 2023, 14, 1178069. [Google Scholar] [CrossRef]
- Tamborski, J.; Krasileva, K.V. Evolution of Plant NLRs: From Natural History to Precise Modifcations. Annu. Rev. Plant Biol. 2020, 29, 355–378. [Google Scholar] [CrossRef]
- Zhao, D.; Li, B.; Zhao, D. Identification and Analysis of U-box Gene Family in Eucommia ulmoides Oliv. Seed 2021, 40, 10–17. [Google Scholar] [CrossRef]
- Ha, K.N.; Mastersona, L.R.; Hou, Z.; Verardia, R.; Walsha, N.; Veglia, G.; Robia, S.L. Lethal Arg9Cys phospholamban mutation hindersCa2+-ATPase regulation and phosphorylationby protein kinase A. Proc. Natl. Acad. Sci. USA 2011, 108, 2735–2740. [Google Scholar] [CrossRef]
- Karkute, S.G.; Kumar, V.; Tasleem, M.; Mishra, D.C.; Chaturvedi, K.K.; Rai, A.; Sevanthi, A.M.; Gaikwad, K.; Sharma, T.R.; Solanke, A.U. Genome-Wide Analysis of von Willebrand Factor A Gene Family in Rice for Its Role in Imparting Biotic Stress Resistance with Emphasis on Rice Blast Disease. Rice Sci. 2022, 29, 375–384. [Google Scholar] [CrossRef]
- Monteiro, F.; Nishimura, M.T. Structural, Functional, and Genomic Diversity of Plant NLR Proteins: An Evolved Resource for Rational Engineering of Plant Immunity. Annu. Rev. Phytopathol. 2018, 25, 243–267. [Google Scholar] [CrossRef]
- Li, J.; Tao, X. Structural insights into the plant NLR immune receptors. J. Nanjing Agric. Univ. 2022, 45, 929–937. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, B.; Dong, J.; Hu, X.; Li, N.; Li, X.; Long, D.; Wu, X. RiceReceptor: The Cell-Surface and Intracellular Immune Receptors of the Oryza Genus. Genes 2025, 16, 597. https://doi.org/10.3390/genes16050597
Jin B, Dong J, Hu X, Li N, Li X, Long D, Wu X. RiceReceptor: The Cell-Surface and Intracellular Immune Receptors of the Oryza Genus. Genes. 2025; 16(5):597. https://doi.org/10.3390/genes16050597
Chicago/Turabian StyleJin, Baihui, Jian Dong, Xiaolong Hu, Na Li, Xiaohua Li, Dawei Long, and Xiaoni Wu. 2025. "RiceReceptor: The Cell-Surface and Intracellular Immune Receptors of the Oryza Genus" Genes 16, no. 5: 597. https://doi.org/10.3390/genes16050597
APA StyleJin, B., Dong, J., Hu, X., Li, N., Li, X., Long, D., & Wu, X. (2025). RiceReceptor: The Cell-Surface and Intracellular Immune Receptors of the Oryza Genus. Genes, 16(5), 597. https://doi.org/10.3390/genes16050597