
ABA-Insensitive 5 (ABI5) Is Involved in ABA-Induced Dormancy via Activating PavCIG1/2 Expression in Sweet Cherries

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Isolation and Gene Expression Analysis
2.3. Yeast One-Hybrid Assays
2.4. Dual-Luciferase Assays
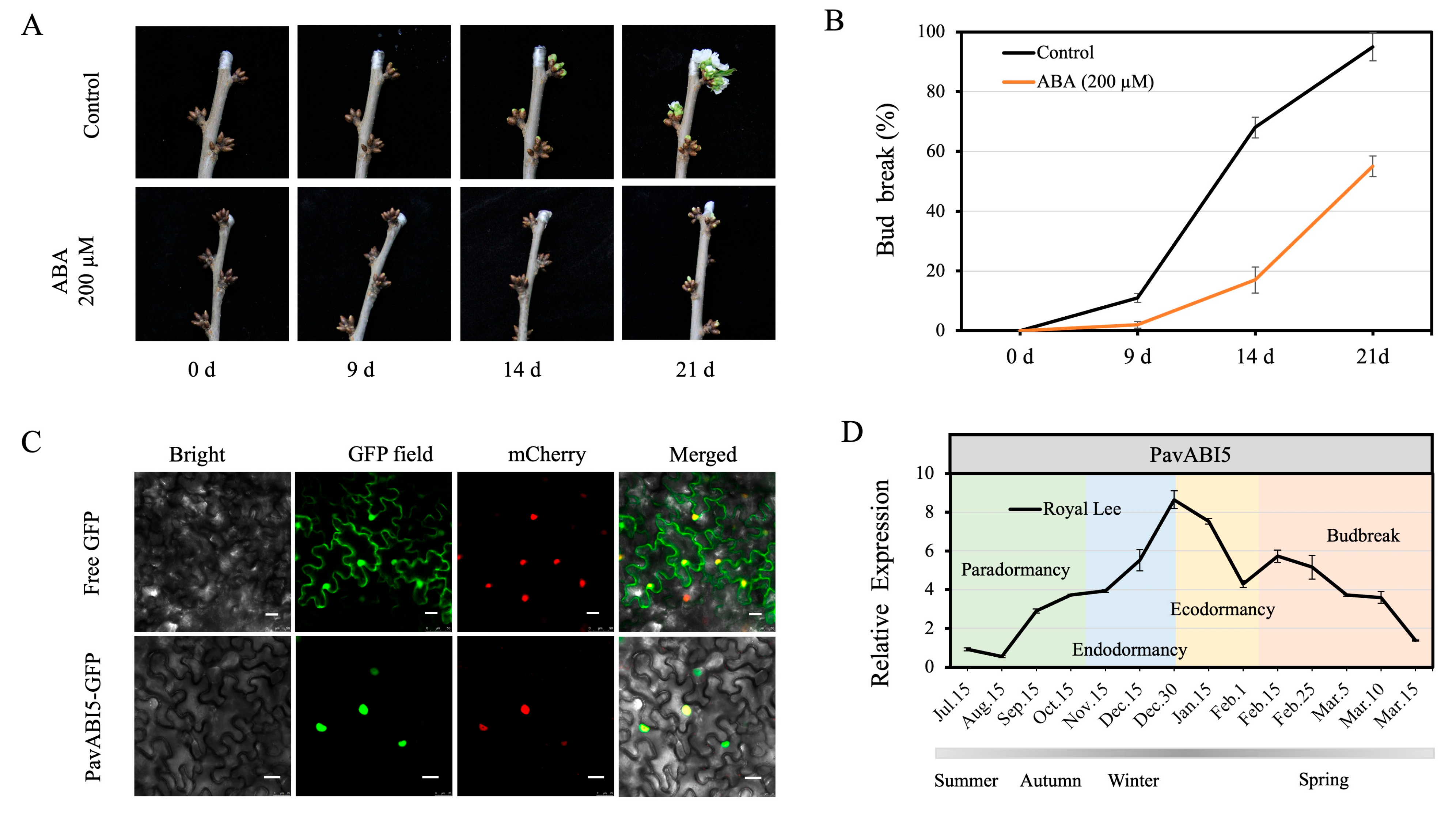
2.5. Subcellular Localization of PavABI5
2.6. ABA and Low Temperature Treatments
2.7. Protein–Protein Interaction Prediction and Expression Pattern Analysis
3. Results
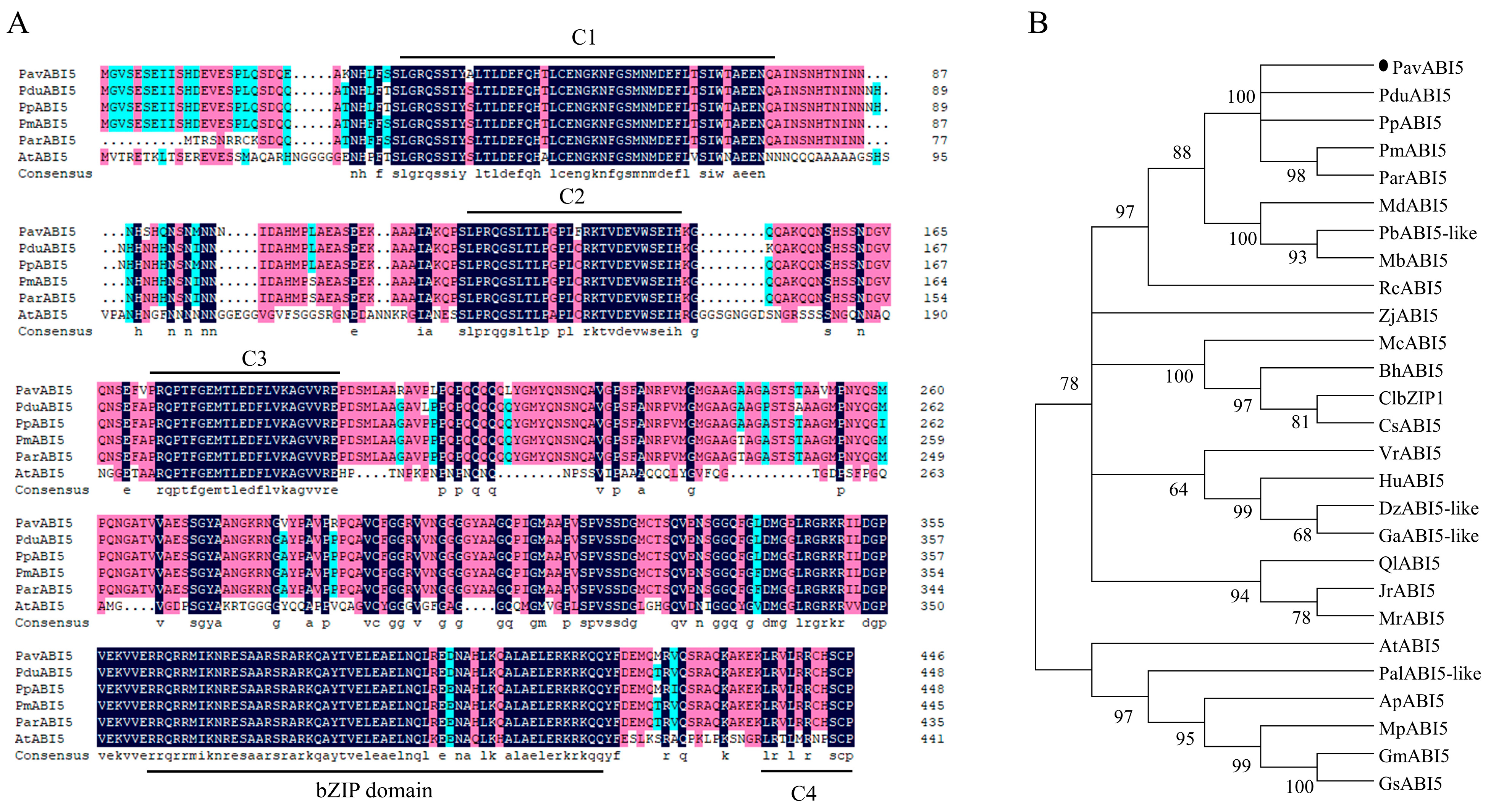
3.1. Identification of Sweet Cherry PavABI5 Gene
3.2. PavCIG1 and PavCIG2 Are Downstream of PavABI5
3.3. ABA Delayed Floral Budburst in Sweet Cherry
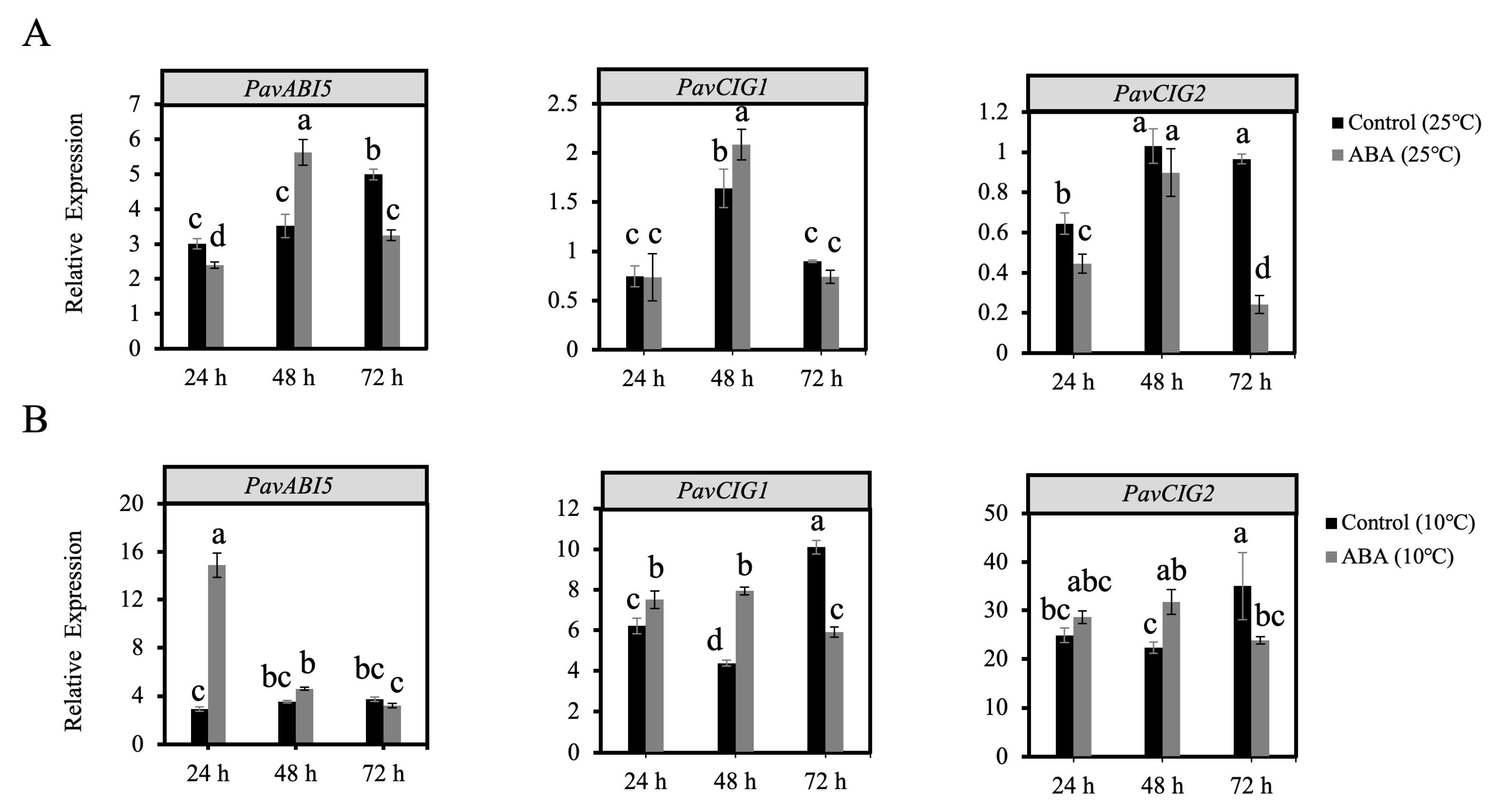
3.4. Gene Expression Analysis Through ABA and Low Temperature Treatments
3.5. Expression Analysis of Downstream Gene DAMs After ABA and Cold Treatments
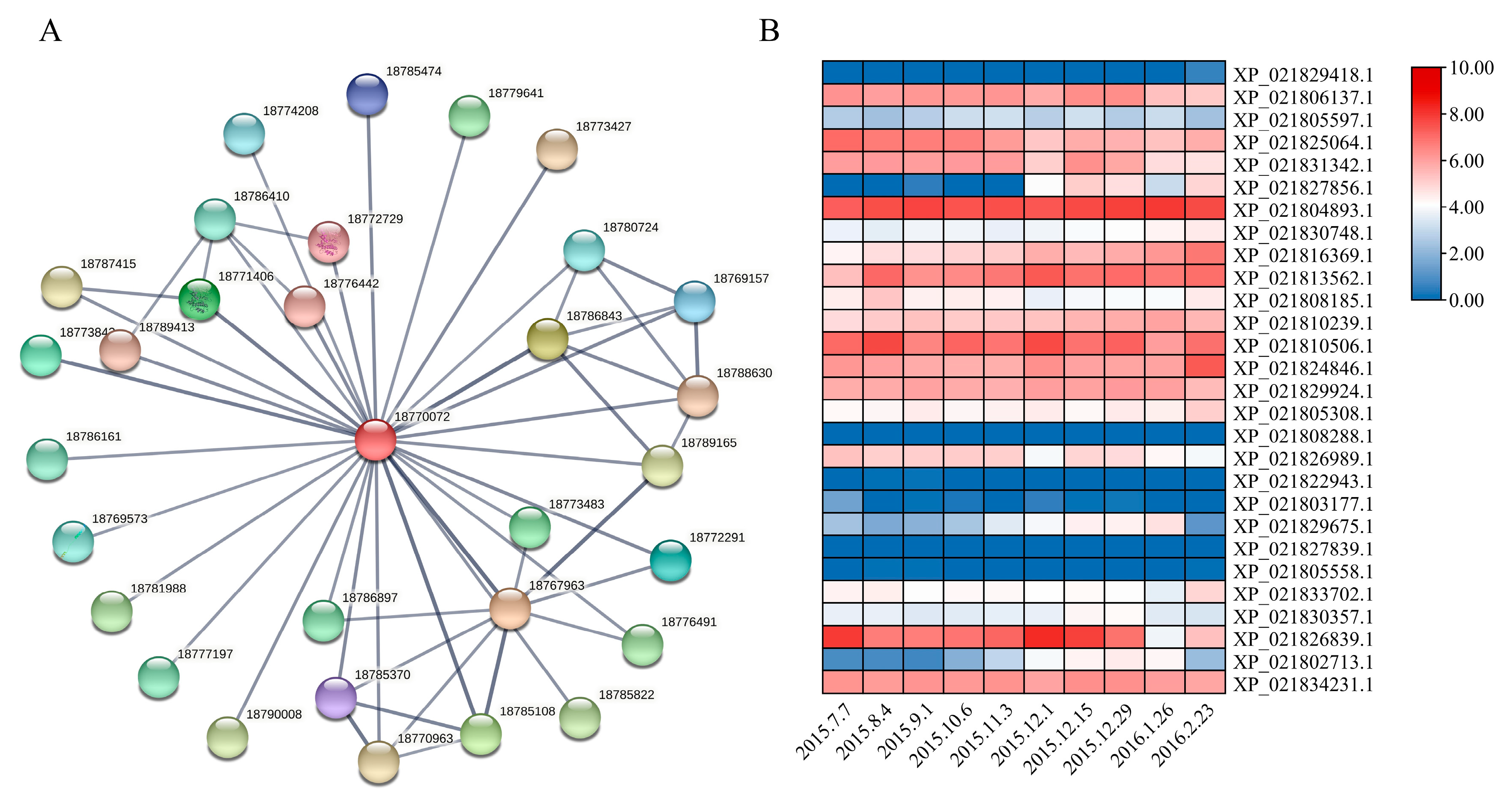
3.6. Prediction of Protein–Protein Interactions Between PavABI5 and Other Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arora, R.; Rowland, L.J.; Tanino, K. Induction and release of bud dormancy in woody perennials: A science comes of age. HortScience 2003, 38, 911–921. [Google Scholar] [CrossRef]
- Rodrigo, J. Spring frosts in deciduous fruit trees-morphological damage and flower hardiness. Sci. Hortic. 2000, 85, 155–173. [Google Scholar] [CrossRef]
- Salazar-Gutiérrez, M.R.; Chaves, B.; Hoogenboom, G. Freezing tolerance of apple flower buds. Sci. Hortic. 2016, 198, 344–351. [Google Scholar] [CrossRef]
- Kaya, O.; Kose, C.; Gecim, T. An exothermic process involved in the late spring frost injury to flower buds of some apricot cultivars (Prunus armenica L.). Sci. Hortic. 2018, 241, 322–328. [Google Scholar] [CrossRef]
- Heide, O.M.; Prestrud, A.K. Low temperature, but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiol. 2005, 25, 109–114. [Google Scholar] [CrossRef]
- Kovaleski, A.P. The potential for an increasing threat of unseasonal temperature cycles to dormant plants. New Phytol. 2024, 244, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zeng, L.; Kong, D.; Mao, Y.; Xu, Y.; Wang, M.; Zhao, Y.; Jiang, C.; Zhang, Y.; Sun, D. Abscisic acid–induced transcription factor PsMYB306 negatively regulates tree peony bud dormancy release. Plant Physiol. 2024, 194, 2449–2471. [Google Scholar] [CrossRef]
- Zheng, C.; Halaly, T.; Acheampong, A.K.; Takebayashi, Y.; Jikumaru, Y.; Kamiya, Y.; Or, E. Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism. J. Exp. Bot. 2015, 66, 1527–1542. [Google Scholar] [CrossRef]
- Alburquerque, N.; García-Montiel, F.; Carrillo, A.; Burgos, L. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ. Exp. Bot. 2008, 64, 162–170. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L.; Ma, C.; Xu, W.; Liu, Z.; Zhang, C.; Matthew, D.W.; Wang, S. Impact of chilling accumulation and hydrogen cyanamide on floral organ development of sweet cherry in a warm region. J. Integr. Agr. 2016, 15, 2529–2538. [Google Scholar] [CrossRef]
- Artlip, T.; McDermaid, A.; Ma, Q.; Wisniewski, M. Differential gene expression in non-transgenic and transgenic “M. 26” apple overexpressing a peach CBF gene during the transition from eco-dormancy to bud break. Hortic. Res. 2019, 6, 86. [Google Scholar] [CrossRef] [PubMed]
- Barros, P.M. Insights into the Role of Almond CBF Transcription Factors in the Environmental Control of Cold Acclimation and Dormancy Break. Doctoral Dissertation, Universidade NOVA de Lisboa, Lisbon, Portugal, 2012. [Google Scholar]
- Artlip, T.S.; Wisniewski, M.E.; Norelli, J.L. Field evaluation of apple overexpressing a peach CBF gene confirms its effect on cold hardiness, dormancy, and growth. Environ. Exp. Bot. 2014, 106, 79–86. [Google Scholar] [CrossRef]
- Niu, Q.; Li, J.; Cai, D.; Qian, M.; Jia, H.; Bai, S.; Hussain, S.; Liu, G.; Teng, Y.; Zheng, X. Dormancy-associated MADS-box genes and microRNAs jointly control dormancy transition in pear (Pyrus pyrifolia white pear group) flower bud. J. Exp. Bot. 2016, 67, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yan, X.; Yang, Q.; Ma, Y.; Yang, B.; Tian, J.; Teng, Y.; Bai, S. PpCBFs selectively regulate PpDAMs and contribute to the pear bud endodormancy process. Plant Mol. Biol. 2019, 99, 575–586. [Google Scholar] [CrossRef]
- Park, S.; Lee, C.M.; Doherty, C.J.; Gilmour, S.J.; Kim, Y.; Thomashow, M.F. Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network. Plant J. 2015, 82, 193–207. [Google Scholar] [CrossRef]
- Artlip, T.S.; Wisniewski, M.E.; Arora, R.; Norelli, J.L. An apple rootstock overexpressing a peach CBF gene alters growth and flowering in the scion but does not impact cold hardiness or dormancy. Hortic. Res. 2016, 3, 16006. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Sun, W.; Xu, Y.; Sabir, I.A.; Abdullah, M.; Wang, S.; Jiu, S.; Zhang, C. Cold induced genes (CIGs) regulate flower development and dormancy in Prunus avium L. Plant Sci. 2021, 313, 111061. [Google Scholar] [CrossRef]
- Zhao, K.; Zhou, Y.; Ahmad, S.; Yong, X.; Xie, X.; Han, Y.; Li, Y.; Zhang, Q. PmCBFs synthetically affect PmDAM6 by alternative promoter binding and protein complexes towards the dormancy of bud for Prunus mume. Sci. Rep. 2018, 8, 4527. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Z.; Li, H.; Jiu, S.; Qu, Y.; Wang, L.; Ma, C.; Xu, W.; Wang, S.; Zhang, C. Dormancy-associated MADS-Box (DAM) genes influence chilling requirement of sweet cherries and co-regulate flower development with SOC1 gene. Int. J. Mol. Sci. 2020, 21, 921. [Google Scholar] [CrossRef]
- Li, Z.; Reighard, G.L.; Abbott, A.G.; Bielenberg, D.G. Dormancy-associated MADS genes from the EVG locus of peach [Prunus persica (L.) Batsch] have distinct seasonal and photoperiodic expression patterns. J. Exp. Bot. 2009, 60, 3521–3530. [Google Scholar] [CrossRef]
- Hu, C.; Wang, M.; Zhu, C.; Wu, S.; Li, J.; Yu, J.; Hu, Z. A transcriptional regulation of ERF15 contributes to ABA-mediated cold tolerance in tomato. Plant Cell Environ. 2024, 47, 1334–1347. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Zareen, S.; Park, J.; Khan, H.A.; Lim, C.J.; Bader, Z.E.; Hussain, S.; Chung, W.S.; Gechev, T.; Pardo, J.M.; et al. ABA INSENSITIVE 2 promotes flowering by inhibiting OST1/ABI5-dependent FLOWERING LOCUS C transcription in Arabidopsis. J. Exp. Bot. 2024, 75, 2481–2493. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L.; Sun, W.; Wang, L.; Liu, X.; Jiu, S.; Liu, R.; Zhang, C. N6-methyladenosine RNA methylation is important for dormancy release in sweet cherry. Sci. Hortic. 2024, 338, 113725. [Google Scholar] [CrossRef]
- Vimont, N.; Schwarzenberg, A.; Domijan, M.; Donkpegan, A.S.; Beauvieux, R.; Le, D.L.; Arkoun, M.; Jamois, F.; Yvin, J.; Wigge, A.P.; et al. Fine tuning of hormonal signaling is linked to dormancy status in sweet cherry flower buds. Tree Physiol. 2021, 41, 544–561. [Google Scholar] [CrossRef]
- Zheng, C.; Acheampong, A.K.; Shi, Z.; Mugzech, A.; Halaly-Basha, T.; Shaya, F.; Sun, Y.; Colova, V.; Mosquna, A.; Ophir, R.; et al. Abscisic acid catabolism enhances dormancy release of grapevine buds. Plant Cell Environ. 2018, 41, 2490–2503. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Lynch, T.J. The Arabidopsis abscisic acid response gene ABI5 encodes a basic leucine zipper transcription factor. Plant Cell 2000, 12, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Gampala, S.S.; Ritchie, G.L.; Payton, P.; Burke, J.J.; Rock, C.D. Related to ABA-Insensitive3 (ABI3)/Viviparous1 and AtABI5 transcription factor coexpression in cotton enhances drought stress adaptation. Plant Biotechnol J. 2014, 12, 578–589. [Google Scholar]
- Chen, J.; Li, Y.; Li, F.; Wu, Q.; Jiang, Y.; Yuan, D. Banana MaABI5 is involved in ABA-induced cold tolerance through interaction with a RING E3 ubiquitin ligase; MaC3HC4-1. Sci. Hortic. 2018, 237, 239–246. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, K.; Zhou, H.; Yan, X.; Zhan, Q.; Zheng, Y.; Song, C.P. ABI5 modulates seed germination via feedback regulation of the expression of the PYR/PYL/RCAR ABA receptor genes. New Phytol. 2020, 228, 596–608. [Google Scholar] [CrossRef]
- Han, Y.; Wang, Z.; Han, B.; Zhang, Y.; Liu, J.; Yang, Y. Allelic variation of TaABI5-A4 significantly affects seed dormancy in bread wheat. Theor. Appl. Genet. 2024, 137, 240. [Google Scholar] [CrossRef]
- Carles, C.; Bies-Etheve, N.; Aspart, L.; Léon-Kloosterziel, K.M.; Koornneef, M.; Echeverria, M.; Delseny, M. Regulation of Arabidopsis thaliana Em genes: Role of ABI5. Plant J. 2002, 30, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, Y.N.; Xue, L.J.; Zou, M.J.; Liu, J.Y.; Chen, F.; Xue, H.W. Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes. Plant Physiol. 2011, 156, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Deng, W.; Wang, X.; Yang, C.; Li, Z. Maize (Zea mays L.) homologue of ABA-insensitive (ABI) 5 gene plays a negative regulatory role in abiotic stresses response. Plant Growth Regul. 2012, 68, 383–393. [Google Scholar] [CrossRef]
- Rubio, S.; Noriega, X.; Pérez, F.J. Abscisic acid (ABA) and low temperatures synergistically increase the expression of CBF/DREB1 transcription factors and cold-hardiness in grapevine dormant buds. Ann. Bot. 2019, 123, 681–689. [Google Scholar] [CrossRef]
- Yang, Q.; Yang, B.; Li, J.; Wang, Y.; Tao, R.; Yang, F.; Wu, X.; Yan, X.; Ahmad, M.; Shen, J.; et al. ABA-responsive ABRE-BINDING FACTOR3 activates DAM3 expression to promote bud dormancy in Asian pear. Plant Cell Environ. 2020, 43, 1360–1375. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein–protein interaction networks.; integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Vimont, N.; Fouché, M.; Campoy, J.A.; Tong, M.; Arkoun, M.; Yvin, J.C.; Wigge, P.A.; Dirlewanger, E.; Cortijo, S.; Wenden, B. From bud formation to flowering: Transcriptomic state defines the cherry developmental phases of sweet cherry bud dormancy. BMC Genom. 2019, 20, 974. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lefebvre, V.; North, H.; Frey, A.; Sotta, B.; Seo, M.; Okamoto, M.; Nambara, E.; Marion-Poll, A. Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy. Plant J. 2006, 45, 309–319. [Google Scholar] [CrossRef]
- Frey, A.; Effroy, D.; Lefebvre, V.; Seo, M.; Perreau, F.; Berger, A.; Sechet, J.; To, A.; North, H.M.; Marion-Poll, A. Epoxycarotenoid cleavage by NCED5 fine-tunes ABA accumulation and affects seed dormancy and drought tolerance with other NCED family members. Plant J. 2012, 70, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases.; are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol. 2006, 141, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Kendall, S.L.; Hellwege, A.; Marriot, P.; Whalley, C.; Graham, I.A.; Penfield, S. Induction of dormancy in Arabidopsis summer annuals requires parallel regulation of DOG1 and hormone metabolism by low temperature and CBF transcription factors. Plant Cell 2011, 23, 2568–2580. [Google Scholar] [CrossRef]
- Cooke, J.E.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gao, J.; Im, K.J.; Chen, K.; Bressan, R.A.; Zhu, J.K. Control of plant water use by ABA induction of senescence and dormancy: An overlooked lesson from evolution. Plant Cell Physiol. 2017, 58, 1319–1327. [Google Scholar] [CrossRef]
- Guak, S.; Fuchigami, L.H. Effects of applied ABA on growth cessation, bud dormancy, cold acclimation, leaf senescence and N mobilization in apple nursery plants. J. Hortic. Sci. Biotech 2001, 76, 459–464. [Google Scholar] [CrossRef]
- Or, E.; Belausov, E.; Popilevsky, I.; Bental, Y. Changes in endogenous ABA level in relation to the dormancy cycle in grapevines grown in a hot climate. J. Hortic. Sci. Biotech 2000, 75, 190–194. [Google Scholar] [CrossRef]
- Li, J.; Xu, Y.; Niu, Q.; He, L.; Teng, Y.; Bai, S. Abscisic acid (ABA) promotes the induction and maintenance of pear (Pyrus pyrifolia white pear group) flower bud endodormancy. Int. J. Mol. Sci. 2018, 19, 310. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Gotz, K.; Homann, T.; Huschek, G.; Rawel, H. Identification of endodormancy release for cherries (Prunus avium L.) by abscisic acid and sugars. J. Hortic. 2017, 4, 1000210. [Google Scholar]
- Park, S.Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.-F.F.; et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef]
- Vlad, F.; Rubio, S.; Rodrigues, A.; Sirichandra, C.; Belin, C.; Robert, N.; Leung, J.; Rodriguez, P.L.; Laurière, C.; Merlot, S. Protein phosphatases 2C regulate the activation of the Snf1-related kinase OST1 by abscisic acid in Arabidopsis. Plant Cell 2009, 21, 3170–3184. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Chen, S.; Yang, Y.; An, C. ABA-insensitive (ABI) 4 and ABI5 synergistically regulate DGAT1 expression in Arabidopsis seedlings under stress. FEBS Lett. 2013, 587, 3076–3082. [Google Scholar] [CrossRef] [PubMed]
- Utsugi, S.; Ashikawa, I.; Nakamura, S.; Shibasaka, M. TaABI5, a wheat homolog of Arabidopsis thaliana ABA insensitive 5, controls seed germination. J. Plant Res. 2020, 133, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Bensmihen, S.; Rippa, S.; Lambert, G.; Jublot, D.; Pautot, V.; Granier, F.; Giraudat, J.; Parcy, F. The homologous ABI5 and EEL transcription factors function antagonistically to fine-tune gene expression during late embryogenesis. Plant Cell 2002, 14, 1391–1403. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-Insensitive 5) in plant development.; abiotic stress responses and phytohormone crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef]
- Wang, Y.H.; Que, F.; Li, T.; Zhang, R.R.; Khadr, A.; Xu, Z.S.; Tian, Y.S.; Xiong, A.S. DcABF3, an ABF transcription factor from carrot.; alters stomatal density and reduces ABA sensitivity in transgenic Arabidopsis. Plant Sci. 2021, 302, 110699. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Seng, S.; Sui, J.; Vonaartis, E.; Luo, X.; Gong, B.; Liu, C.; Wu, C.; Liu, C.; Zhang, F.; et al. Gladiolus hybridus ABSCISIC ACID INSENSITIVE 5 (GhABI5) is an important transcription factor in ABA signaling that can enhance Gladiolus corm dormancy and Arabidopsis seed dormancy. Front. Plant Sci. 2015, 6, 960. [Google Scholar] [CrossRef]
- Miura, K.; Lee, J.; Jin, J.B.; Yoo, C.Y.; Miura, T.; Hasegawa, P.M. Sumoylation of ABI5 by the Arabidopsis SUMO E3 ligase SIZ1 negatively regulates abscisic acid signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 5418–5423. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Mongrand, S.; McLachlin, D.T.; Chait, B.T.; Chua, N.H. ABI5 acts downstream of ABI3 to execute an ABA-dependent growth arrest during germination. Plant J. 2002, 32, 317–328. [Google Scholar] [CrossRef]
- Wang, Y.; Tao, Z.; Wang, W.; Filiault, D.; Qiu, C.; Wang, C.; Wang, H.; Rehman, S.; Shi, J.; Zhang, Y.; et al. Molecular variation in a functionally divergent homolog of FCA regulates flowering time in Arabidopsis thaliana. Nat. Commun. 2020, 11, 5830. [Google Scholar] [CrossRef]
- Murmu, J.; Bush, M.J.; DeLong, C.; Li, S.; Xu, M.; Khan, M.; Malcolmson, C.; Fobert, P.R.; Zachgo, S.; Hepworth, S.R. Arabidopsis basic leucine-zipper transcription factors TGA9 and TGA10 interact with floral glutaredoxins ROXY1 and ROXY2 and are redundantly required for anther development. Plant Physiol. 2010, 154, 1492–1504. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Siddiqua, M.; Braybrook, S.; Nassuth, A. Three grape CBF/DREB1 genes respond to low temperature, drought and abscisic acid. Plant Cell Environ. 2006, 29, 1410–1421. [Google Scholar] [CrossRef]
- An, D.; Ma, Q.; Wang, H.; Yang, J.; Zhou, W.; Zhang, P. Cassava C-repeat binding factor 1 gene responds to low temperature and enhances cold tolerance when overexpressed in Arabidopsis and cassava. Plant Mol. Biol. 2017, 94, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Siddiqua, M.; Nassuth, A. Vitis CBF1 and Vitis CBF4 differ in their effect on Arabidopsis abiotic stress tolerance.; development and gene expression. Plant Cell Environ. 2011, 34, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, J.; Guo, H.; He, X.; Wu, W.; Du, J.; Zhang, Z.; An, X. Characterization of two highly similar CBF/DREB1-like genes, PhCBF4a and PhCBF4b, in Populus hopeiensis. Plant Physiol. Bioch. 2014, 83, 107–116. [Google Scholar] [CrossRef]
- Wisniewski, M.; Norelli, J.; Artlip, T. Overexpression of a peach CBF gene in apple: A model for understanding the integration of growth, dormancy, and cold hardiness in woody plants. Front. Plant Sci. 2015, 6, 85. [Google Scholar] [CrossRef]
- Tuan, P.A.; Bai, S.; Saito, T.; Ito, A.; Moriguchi, T. Dormancy-Associated MADS-Box (DAM) and the abscisic acid pathway regulate pear endodormancy through a feedback mechanism. Plant Cell Physiol. 2017, 58, 1378–1390. [Google Scholar] [CrossRef]
- Yamane, H.; Wada, M.; Honda, C.; Matsuura, T.; Ikeda, Y.; Hirayama, T.; Osako, Y.; Gao-Takai, M.; Kojima, M.; Sakakibara, H.; et al. Overexpression of Prunus DAM6 inhibits growth.; represses bud break competency of dormant buds and delays bud outgrowth in apple plants. PLoS ONE 2019, 14, e0214788. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID. | Protein ID Prunus persica | Peach Protein Name | Score | Protein ID Prunus avium |
|---|---|---|---|---|
| 18767963 | XP_007200956 | B3 domain-containing transcription factor ABI3 | 1.0 | XP_021829418.1 |
| 18786843 | XP_007220580 | E3 ubiquitin-protein ligase KEG | 0.99 | XP_021806137.1 |
| 18785108 | XP_007221036 | ethylene-responsive transcription factor ABI4 | 0.97 | XP_021805597.1 |
| 18771406 | XP_007202101 | serine/threonine-protein kinase SRK2I | 0.92 | XP_021825064.1 |
| 18773843 | XP_020423164 | E3 SUMO-protein ligase SIZ1 | 0.91 | XP_021831342.1 |
| 18772291 | XP_007206180 | protein SLE1-like | 0.83 | XP_021827856.1 |
| 18769157 | XP_007204373 | eukaryotic initiation factor 4A-2-like | 0.83 | XP_021804893.1 |
| 18785370 | XP_007218800 | xanthoxin dehydrogenase | 0.82 | XP_021830748.1 |
| 18772729 | XP_007205499 | serine/threonine-protein kinase SAPK1 | 0.82 | XP_021816369.1 |
| 18776442 | XP_020419823 | serine/threonine-protein kinase SAPK3 | 0.82 | XP_021813562.1 |
| 18789413 | XP_007222500 | serine/threonine-protein kinase SAPK2 | 0.82 | XP_021808185.1 |
| 18788630 | XP_007222234 | WD repeat-containing protein DWA2 | 0.81 | XP_021810239.1 |
| 18773427 | XP_007205151 | CBL-interacting serine/threonine-protein kinase 11 | 0.81 | XP_021810506.1 |
| 18770963 | XP_020423783 | zeaxanthin epoxidase | 0.79 | XP_021824846.1 |
| 18787415 | XP_020413451 | abscisic acid-insenstivie 5-like protein 2 | 0.78 | XP_021829924.1 |
| 18789165 | XP_007222944 | E3 ubiquitin-protein ligase AIP2 | 0.78 | XP_021805308.1 |
| 18790008 | XP_020419715 | flowering time control protein FCA | 0.78 | XP_021808288.1 |
| 18785822 | XP_007219735 | E3 SUMO-protein ligase SIZ1-like | 0.77 | XP_021826989.1 |
| 18781988 | XP_020416273 | B3 domain-containing transcription factor FUS3 | 0.75 | XP_021822943.1 |
| 18776491 | XP_007209687 | protein MOTHER of FT and TFL1 isoform X2 | 0.75 | XP_021803177.1 |
| 18779641 | XP_007213620 | transcription factor TGA9 | 0.74 | XP_021829675.1 |
| 18773483 | XP_007206180 | late embryogenesis abundant protein EMB564-like | 0.74 | XP_021827839.1 |
| 18786897 | XP_007219705 | em protein H5 | 0.74 | XP_021805558.1 |
| 18777197 | XP_007209879 | probable aminotransferase ACS10 | 0.74 | XP_021833702.1 |
| 18786161 | XP_007219026 | probable aminotransferase ACS12 | 0.74 | XP_021830357.1 |
| 18786410 | XP_007220143 | probable protein phosphatase 2C 75 | 0.72 | XP_021826839.1 |
| 18769573 | XP_007204568 | bZIP transcription factor TGA10 | 0.72 | XP_021802713.1 |
| 18780724 | XP_007214632 | cullin-4 | 0.72 | XP_021834231.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Wang, L.; Usman, M.; Zhu, J.; Jiu, S.; Liu, R.; Zhang, C. ABA-Insensitive 5 (ABI5) Is Involved in ABA-Induced Dormancy via Activating PavCIG1/2 Expression in Sweet Cherries. Genes 2025, 16, 596. https://doi.org/10.3390/genes16050596
Wang J, Wang L, Usman M, Zhu J, Jiu S, Liu R, Zhang C. ABA-Insensitive 5 (ABI5) Is Involved in ABA-Induced Dormancy via Activating PavCIG1/2 Expression in Sweet Cherries. Genes. 2025; 16(5):596. https://doi.org/10.3390/genes16050596
Chicago/Turabian StyleWang, Jiyuan, Li Wang, Muhammad Usman, Jie Zhu, Songtao Jiu, Ruie Liu, and Caixi Zhang. 2025. "ABA-Insensitive 5 (ABI5) Is Involved in ABA-Induced Dormancy via Activating PavCIG1/2 Expression in Sweet Cherries" Genes 16, no. 5: 596. https://doi.org/10.3390/genes16050596
APA StyleWang, J., Wang, L., Usman, M., Zhu, J., Jiu, S., Liu, R., & Zhang, C. (2025). ABA-Insensitive 5 (ABI5) Is Involved in ABA-Induced Dormancy via Activating PavCIG1/2 Expression in Sweet Cherries. Genes, 16(5), 596. https://doi.org/10.3390/genes16050596