
The Analysis of Autosomal STRs Draws the Current Genetic Map and Evolutionary History of Northernmost South America
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Selection and Classification
2.2. DNA Extraction, Amplification, and Genotyping
2.3. Statistical and Population Genetic Analysis
2.4. Classifications Based on Ethnic Belonging
3. Results
3.1. A Priori Classifications Based on Genealogy
3.2. Final Population Classifications
3.3. Genetic Diversity Analysis
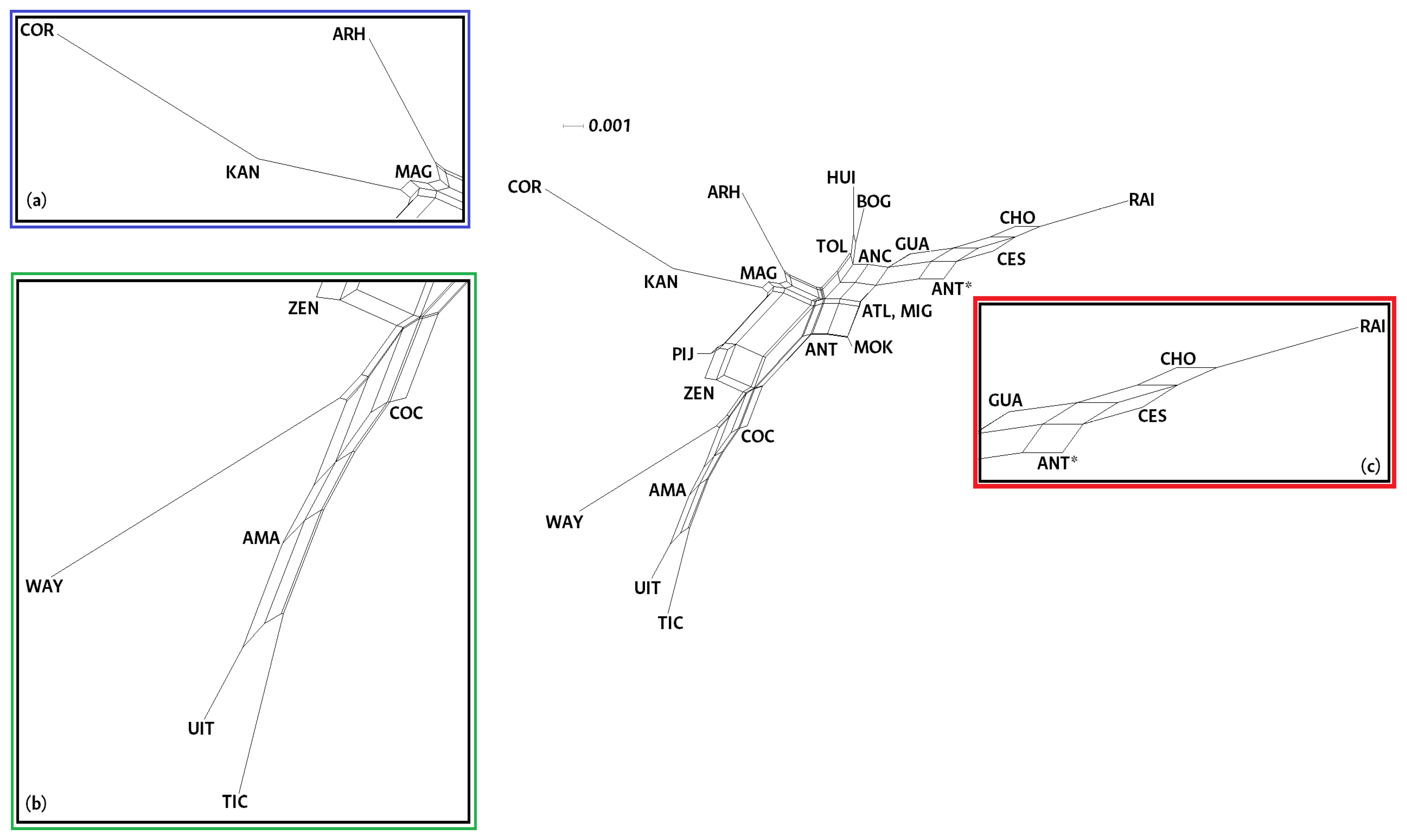
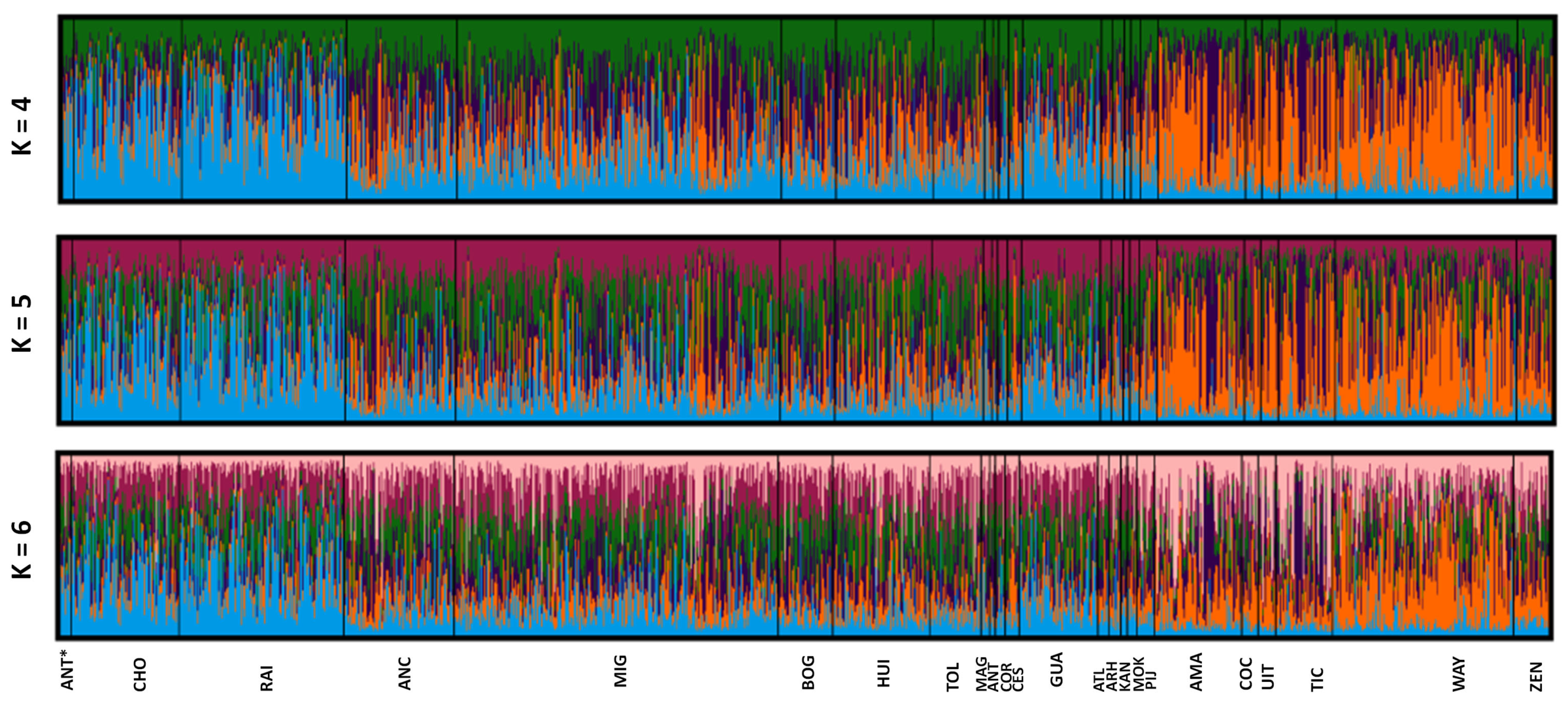
3.4. Genetic Structure Analysis
3.5. Comparative Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, S. The Journey of Man: A Genetic Odyssey; Princeton University Press: Princeton, NJ, USA, 2017; Volume 51. [Google Scholar]
- Roberts, D.F.; Cavalli-Sforza, L.L. The History and Geography of Human Genes. J. R. Anthropol. Inst. 1996, 2, 165–166. [Google Scholar] [CrossRef]
- Grillo, R. Emigración italiana a las Américas. Hisp. Escandinava 2013, 2, 66–86. [Google Scholar]
- Goossen, B.W. Mennonites in Latin America: A review of the literature. Conrad Grebel Rev. 2016, 34, 236–265. [Google Scholar]
- Homburger, J.R.; Moreno-Estrada, A.; Gignoux, C.R.; Nelson, D.; Sanchez, E.; Ortiz-Tello, P.; Pons-Estel, B.A.; Acevedo-Vasquez, E.; Miranda, P.; Langefeld, C.D.; et al. Genomic Insights into the Ancestry and Demographic History of South America. PLoS Genet. 2015, 11, 1–26. [Google Scholar] [CrossRef]
- Masterson, D.M.; Funada-Classen, S. The Japanese in Latin America; Illinois University Press: Champaine, IL, USA, 2004; Volume 132, ISBN 0-252-02869-4. [Google Scholar]
- Rothhammer, F.; Dillehay, T.D. The late pleistocene colonization of South America: An interdisciplinary perspective. Ann. Hum. Genet. 2009, 73, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Crosby, A.W. The Columbian Exchange: Biological and Cultural Consequences of 1492; Greenwood Publishing Company: Westport, CT, USA, 1972. Available online: https://books.google.com.co/books?id=YoWI_LkA8KEC (accessed on 10 December 2022).
- Fawcett, L.; Carbó, E.P. En la tierra de las oportunidades: Los sirios-libaneses en Colombia. J. BoletÍN Cult. Y BibliogrÁFico 1992, 29, 29. Available online: https://www.cervantesvirtual.com/obra/en-la-tierra-de-las-oportunidades-los-sirios-libaneses-en-colombia-917086 (accessed on 24 November 2024).
- Franky, C.E.; Zárate, C.G. Imani Mundo: Estudios en la Amazonia Colombiana; Universidad Nacional de Colombia, Editorial Unibiblos: Bogota, Colombia, 2001; Available online: https://www.researchgate.net/publication/311800596_Imani_Mundo_Estudios_en_la_Amazonia_colombiana (accessed on 17 November 2022).
- Tovar Pinzón, H. Emigración y éxodo en la historia de Colombia. Amérique Lat. Hist. Mémoire 2001. [Google Scholar] [CrossRef]
- Niño, J. Las migraciones forzadas de población, por la violencia. Colomb. una Hist. Éxodos Miedo Terror. y Pobreza. Scr. Nov. 1999, 45, 33. [Google Scholar]
- Camargo, G.; Sampayo, A.M.; Peña Galindo, A.; Escobedo, F.J.; Carriazo, F.; Feged-Rivadeneira, A. Exploring the dynamics of migration, armed conflict, urbanization, and anthropogenic change in Colombia. PLoS ONE 2020, 15, e0242266. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.E.; Ferraz, T.; Couto-Silva, C.M.; Lemes, R.B.; Nunes, K.; Comas, D.; Hünemeier, T. Population Histories and Genomic Diversity of South American Natives. Mol. Biol. Evol. 2021, 39, msab339. [Google Scholar] [CrossRef]
- Posth, C.; Nakatsuka, N.; Lazaridis, I.; Skoglund, P.; Mallick, S.; Lamnidis, T.C.; Rohland, N.; Nägele, K.; Adamski, N.; Bertolini, E.; et al. Reconstructing the Deep Population History of Central and South America. Cell 2018, 175, 1185–1197.e22. [Google Scholar] [CrossRef]
- Harcourt, A.H. Human phylogeography and diversity. Proc. Natl. Acad. Sci. USA 2016, 113, 8072–8078. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Rousset, F. Genepop version 4.6.9. March 2017. Available online: http://kimura.univ-montp2.fr/~rousset/Genepop.htm (accessed on 23 June 2022).
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of Molecular Variance Inferred From Metric Distances Among DNA Haplotypes: Application to Human Mitochondrial DNA Restriction Data. Genetics 1992, 491, 479–491. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Wen, X.; Falush, D. Documentation for structure software: Version 2.3. Univ. Chicago 2010, 6, 321–326. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kovach, W.L. MVSP-A Multivariate Statistical Package for Windows, Ver. 3.22; Kovach Comput. Serv. Pentraeth: Wales, UK, 1999; Volume 137. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2019. [Online]. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 7 November 2022).
- González-Martín, A.; Gorostiza, A.; Rangel-Villalobos, H.; Acunha, V.; Barrot, C.; Sánchez, C.; Ortega, M.; Gené, M.; Calderón, R. Analyzing the genetic structure of the Tepehua in relation to other neighbouring Mesoamerican populations. A study based on allele frequencies of STR markers. Am. J. Hum. Biol. 2008, 20, 605–613. [Google Scholar] [CrossRef]
- Ibarra-Rivera, L.; Mirabal, S.; Regueiro, M.M.; Herrera, R.J. Delineating genetic relationships among the Maya. Am. J. Phys. Anthropol. 2008, 135, 329–347. [Google Scholar] [CrossRef] [PubMed]
- Kohlrausch, F.B.; Callegari-Jacques, S.M.; Tsuneto, L.T.; Petzl-Erler, M.L.; Hill, K.; Hurtado, A.M.; Salzano, F.M.; Hutz, M.H. Geography influences microsatellite polymorphism diversity in Amerindians. Am. J. Phys. Anthropol. 2005, 126, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Bernal, L.P.; Borjas, L.; Zabala, W.; Portillo, M.G.; Fernández, E.; Delgado, W.; Tovar, F.; Lander, N.; Chiurillo, M.A.; Ramírez, J.L.; et al. Genetic variation of 15 STR autosomal loci in the Maracaibo population from Venezuela. Forensic Sci. Int. 2006, 161, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Rodrıguez, A.; Arrieta, G.; Sanóu, I.; Vargas, M.C.; Garcıa, O.; Yurrebaso, I.; Pérez, J.A.; Villalta, M.; Espinoza, M. Population genetic data for 18 STR loci in Costa Rica. Forensic Sci. Int. 2007, 168, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, C. Análisis de ADN Mitocondrial y de Polimorfismos Genéticos de los Cromosomas Autosómicos y Sexuales en la Población Mestiza de Nicaragua. Ph.D. Thesis, Universidad de Zaragoza, Zaragoza, Spain, 2011. [Google Scholar]
- Ramachandran, S.; Deshpande, O.; Roseman, C.C.; Rosenberg, N.A.; Feldman, M.W.; Cavalli-Sforza, L.L. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa. Proc. Natl. Acad. Sci. USA 2005, 102, 15942–15947. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.F.; Mourant, A.E.; Kopec, A.C.; Domaniewska-Sobczak, K. Blood Groups and Diseases: A Study of Associations of Diseases with Blood Groups and Other Polymorphisms. J. Med. Genet. 1979, 16, 241–242. [Google Scholar] [CrossRef]
- Adhikari, K.; Chacón-Duque, J.C.; Mendoza-Revilla, J.; Fuentes-Guajardo, M.; Ruiz-Linares, A. The genetic diversity of the Americas. Annu. Rev. Genomics Hum. Genet. 2017, 18, 277–296. [Google Scholar] [CrossRef]
- Ossa, H.; Aquino, J.; Pereira, R.; Ibarra, A.; Ossa, R.H.; Pérez, L.A.; Granda, J.D.; Lattig, M.C.; Groot, H.; De Carvalho, E.F.; et al. Outlining the ancestry landscape of Colombian admixed populations. PLoS ONE 2016, 11, e0164414. [Google Scholar] [CrossRef]
- Mogollón Olivares, F.; Moncada Madero, J.; Casas-Vargas, A.; Zea Montoya, S.; Suárez Medellín, D.; Gusmão, L.; Usaquén, W. Contrasting the ancestry patterns of three distinct population groups from the northernmost region of South America. Am. J. Phys. Anthropol. 2020, 173, 24130. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, M. La granjería de las perlas del Río de la Hacha: Rebelión y resistencia esclava (1570–1615). Hist. Caribe 2003, 3, 8. [Google Scholar]
- Suárez, D.; Cruz, R.; Torres, M.; Mogollón, F.; Moncada, J.; Carracedo, A.; Usaquén, W. Ancestry analysis using autosomal SNPs in northern South America, reveals interpretation differences between an AIM panel and an identification panel. Forensic Sci. Int. 2021, 326, 110934. [Google Scholar] [CrossRef] [PubMed]
- Keyeux, G.; Usaquén, W. Rutas migratorias hacia Sudamérica y poblamiento de las cuencas de los ríos Amazonas y Orinoco, deducidas a partir de estudios genéticos moleculares. In Pueblos y Paisajes Antiguos de la Selva Amazónica; Morcote, G., Mora, S., Calvo, C., Eds.; Universidad Nacional de Colombia: Bogotá, Colombia, 2006; p. 415. [Google Scholar]
- Wang, S.; Lewis, C.M.; Jakobsson, M.; Ramachandran, S.; Ray, N.; Bedoya, G.; Rojas, W.; Parra, M.V.; Molina, J.A.; Gallo, C.; et al. Genetic variation and population structure in Native Americans. PLoS Genet. 2007, 3, 2049–2067. [Google Scholar] [CrossRef] [PubMed]
- Tarazona-Santos, E.; Carvalho-Silva, D.R.; Pettener, D.; Luiselli, D.; De Stefano, G.F.; Labarga, C.M.; Rickards, O.; Tyler-Smith, C.; Pena, S.D.; Santos, F.R. Genetic differentiation in South Amerindians is related to environmental and cultural diversity: Evidence from the Y chromosome. Am. J. Hum. Genet. 2001, 68, 1485–1496. [Google Scholar] [CrossRef] [PubMed]
- Fuselli, S.; Tarazona-Santos, E.; Dupanloup, I.; Soto, A.; Luiselli, D.; Pettener, D. Mitochondrial DNA diversity in South America and the genetic history of Andean highlanders. Mol. Biol. Evol. 2003, 20, 1682–1691. [Google Scholar] [CrossRef]
- Reich, D.; Patterson, N.; Campbell, D.; Tandon, A.; Mazieres, S.; Ray, N.; Parra, M.V.; Rojas, W.; Duque, C.; Mesa, N.; et al. Reconstructing Native American population history. Nature 2012, 488, 370. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, P.; Mallick, S.; Bortolini, M.C.; Chennagiri, N.; Hünemeier, T.; Petzl-Erler, M.L.; Salzano, F.M.; Patterson, N.; Reich, D. Genetic evidence for two founding populations of the Americas. Nature 2015, 525, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, P.; Reich, D. A genomic view of the peopling of the Americas. Curr. Opin. Genet. Dev. 2016. [Google Scholar] [CrossRef]
- G.-Camargo, D.; Camacho-Mejorado, R.; Gómez-Alegría, C.; Alario, A.; H.-Tobías, E.; García, G.; M.-Ríos, M.; Gómez, R. Genetic Structure of Cartagena de Indias Population Using Hypervariable Markers of Y Chromosome. Open J. Genet. 2015, 5, 27–41. [Google Scholar] [CrossRef]
- Rojas, W.; Parra, M.V.; Campo, O.; Caro, M.A.; Lopera, J.G.; Arias, W.; Duque, C.; Naranjo, A.; García, J.; Vergara, C.; et al. Genetic make up and structure of Colombian populations by means of uniparental and biparental DNA markers. Am. J. Phys. Anthropol. 2010, 143, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Noguera, M.; Schwegler, A.; Gomes, V.; Briceño, I.; Alvarez, L.; Uricoechea, D.; Amorim, A.; Benavides, E.; Silvera, C.; Charris, M.; et al. Colombia’s racial crucible: Y chromosome evidence from six admixed communities in the Department of Bolivar. Ann. Hum. Biol. 2013, 41, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Alonso, L.A.; Usaquén, W. Y-chromosome and surname analysis of the native islanders of San Andrés and Providencia (Colombia). HOMO-J. Comp. Hum. Biol. 2013, 64, 71–84. [Google Scholar] [CrossRef]
- Trombetta, B.; Cruciani, F.; Sellitto, D.; Scozzari, R. A new topology of the human Y chromosome haplogroup E1b1 (E-P2) revealed through the use of newly characterized binary polymorphisms. PLoS ONE 2011, 6, e16073. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, M.C. Génesis y Desarrollo de la Esclavitud en Colombia Siglos XVI y XVII; Universidad del Valle: Cali, Colombia, 2005. [Google Scholar]
- Wang, S.; Ray, N.; Rojas, W.; Parra, M.V.; Bedoya, G.; Gallo, C.; Poletti, G.; Mazzotti, G.; Hill, K.; Hurtado, A.M. Geographic patterns of genome admixture in Latin American Mestizos. PLoS Genet. 2008, 4, e1000037. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Estrada, A.; Gignoux, C.R.; Fernández-López, J.C.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuña-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science (80) 2014, 344, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Chacón-Duque, J.C.; Adhikari, K.; Fuentes-Guajardo, M.; Mendoza-Revilla, J.; Acuña-Alonzo, V.; Barquera, R.; Quinto-Sánchez, M.; Gómez-Valdés, J.; Martínez, P.E.; Villamil-Ramírez, H.; et al. Latin Americans show wide-spread Converso ancestry and imprint of local Native ancestry on physical appearance. Nat. Commun. 2018. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.N.; Song, W.; Shetty, A.C.; Levano, K.S.; Cáceres, O.; Padilla, C.; Borda, V.; Tarazona, D.; Trujillo, O.; Sanchez, C.; et al. Evolutionary genomic dynamics of Peruvians before, during, and after the Inca Empire. Proc. Natl. Acad. Sci. USA 2018, 115, E6526–E6535. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
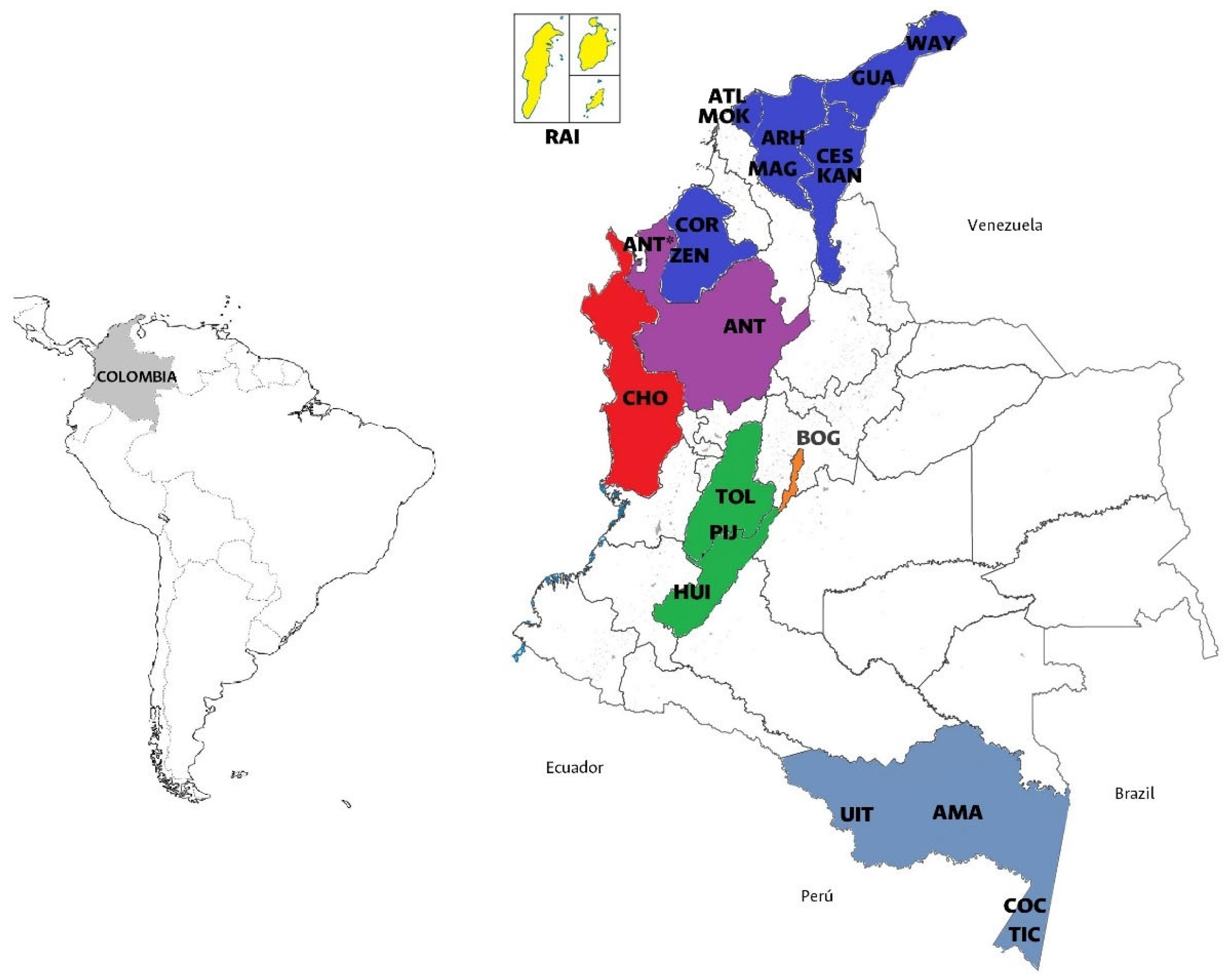
| Classification | Abbreviation | n | Criteria |
|---|---|---|---|
| Bogotá | BOG | 50 | Participants born in Bogotá whose data were obtained from filiation cases. They reported the highest paternity rates, and both their parents were also born in Bogotá. |
| Tolima | TOL | 47 | Participants who did not report belonging to any ethnic group and whose parents were also born in the given department. |
| Huila | HUI | 89 | |
| Amazonas | AMA | 80 | |
| Atlántico | ATL | 10 | |
| Antioquia | ANT | 8 | |
| Cesar | CES | 9 | |
| Córdoba | COR | 13 | |
| Guajira | GUA | 72 | |
| Magdalena | MAG | 5 | |
| Afro-Antioquia | ANT* | 10 | Participants from Antioquia who were born in the municipalities of Vigía del Fuerte, Necoclí, and Turbo who self-determined as Afro-descendants. |
| Chocó | CHO | 99 | Afro-descendants from Chocó. |
| San Andrés and Providencia | RAI | 151 | A total of 117 Raizales and 34 born there whose mother and father are also from San Andrés and Providencia |
| Wayúu | WAY | 166 | Participants who self-determined as belonging to a specific Native American ethnicity. |
| Zenú | ZEN | 32 | |
| Arhuacos | ARH | 11 | |
| Kankuamos | KAN | 6 | |
| Mokaná | MOK | 9 | |
| Pijao | PIJ | 16 | |
| Ticuna | TIC | 52 | |
| Uitoto | UIT | 16 | |
| Cocama | COC | 15 | |
| Migrants | MIG | 297 | Participants whose parents were born in a different region from themselves. |
| Multiple ancestries | ANC | 101 | Individuals whose paternal and maternal lineages differed from each other. |
| Population | Location | Code | Reference |
|---|---|---|---|
| Maya | Mexico—Native | IMAY | pop.STR—USC * |
| Pima | Mexico—Native | IPIM | pop.STR—USC |
| Tepehua | Mexico—Native | ITAP | [27] |
| Yucatán | Mexico—Native | IYUC | [28] |
| Caingang | Brazil—Native | ICAI | [29] |
| Guarani | Brazil—Native | IGUA | [29] |
| Karitiana | — | IKAR | pop.STR—USC |
| Surui | Brazil—Native | ISUR | pop.STR—USC |
| Aché | Paraguay—Native | IACH | [29] |
| Ayoreo | Paraguay—Native | IAYO | [29] |
| Venezuela | Venezuela—Admixed | MVEN | [30] |
| Costa Rica | Costa Rica—Admixed | MCR | [31] |
| Nicaragua | Nicaragua—Admixed | MNIC | [32] |
| Dominican Republic | Dominican Republic—Admixed | MDR | pop.STR—USC |
| Biaka Pygmies | Central African Republic | ABIA | pop.STR—USC |
| Mbuti Pygmies | Democratic Republic of Congo | AMBU | pop.STR—USC |
| Bantu N.E. | Kenya | AKBA | pop.STR—USC |
| Yoruba | Nigeria | ANYO | pop.STR—USC |
| Mandenka | Senegal | ASMA | pop.STR—USC |
| Somalia | Somalia | ASOM | pop.STR—USC |
| Bantu | South Africa | ASAB | pop.STR—USC |
| U.S. Europeans | United States | USEU | pop.STR—USC |
| French | France | FRA | pop.STR—USC |
| N.W. Spain | Spain | SPN | pop.STR—USC |
| Agrupación | Code | Participantes | |
|---|---|---|---|
| AFRO | Afro-Insular | AFRI | Raizales |
| Afro-Pacific | AFRP | Chocó and Antioquia Afro | |
| NATIVE AMERICAN | Wayúu | IWAY | Wayúu |
| Zenú | IZEN | Zenú and Córdoba | |
| Native Americans from the Amazon | IAMA | Ticuna, Uitoto, Cocama, and Amazonas | |
| ADMIXED | Pijaos | IPIJ | Pijaos |
| Native American admixture | MICA | Kankuamos, Arhuacos, Magdalena, Mokaná, and Atlántico | |
| Afro admixture | MACA | Guajira and Cesar | |
| European admixture | MEU | Tolima, Huila, Bogotá, Antioquia, Multiple ancestry, and Migrants |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moncada Madero, J.; Mogollón Olivares, F.; Suárez Medellín, D.; Coronel Guzmán, A.; Casas-Vargas, A.; Usaquén Martínez, W. The Analysis of Autosomal STRs Draws the Current Genetic Map and Evolutionary History of Northernmost South America. Genes 2025, 16, 574. https://doi.org/10.3390/genes16050574
Moncada Madero J, Mogollón Olivares F, Suárez Medellín D, Coronel Guzmán A, Casas-Vargas A, Usaquén Martínez W. The Analysis of Autosomal STRs Draws the Current Genetic Map and Evolutionary History of Northernmost South America. Genes. 2025; 16(5):574. https://doi.org/10.3390/genes16050574
Chicago/Turabian StyleMoncada Madero, Julie, Fernanda Mogollón Olivares, Dayana Suárez Medellín, Alejandra Coronel Guzmán, Andrea Casas-Vargas, and William Usaquén Martínez. 2025. "The Analysis of Autosomal STRs Draws the Current Genetic Map and Evolutionary History of Northernmost South America" Genes 16, no. 5: 574. https://doi.org/10.3390/genes16050574
APA StyleMoncada Madero, J., Mogollón Olivares, F., Suárez Medellín, D., Coronel Guzmán, A., Casas-Vargas, A., & Usaquén Martínez, W. (2025). The Analysis of Autosomal STRs Draws the Current Genetic Map and Evolutionary History of Northernmost South America. Genes, 16(5), 574. https://doi.org/10.3390/genes16050574