
Statistical Analysis of Reproductive Traits in Jinwu Pig and Identification of Genome-Wide Association Loci
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Animals and Phenotypic Statistical Analysis
2.2. Genotyping and Quality Control
2.3. Fixed Effects’ Statistical Modeling
2.4. Estimation of Genetic Parameters
2.5. Principal Component Analysis
2.6. Genome-Wide Association Study (GWAS)
2.7. Linkage Disequilibrium and Haplotype Analysis
2.8. Candidate Gene Annotation
3. Results
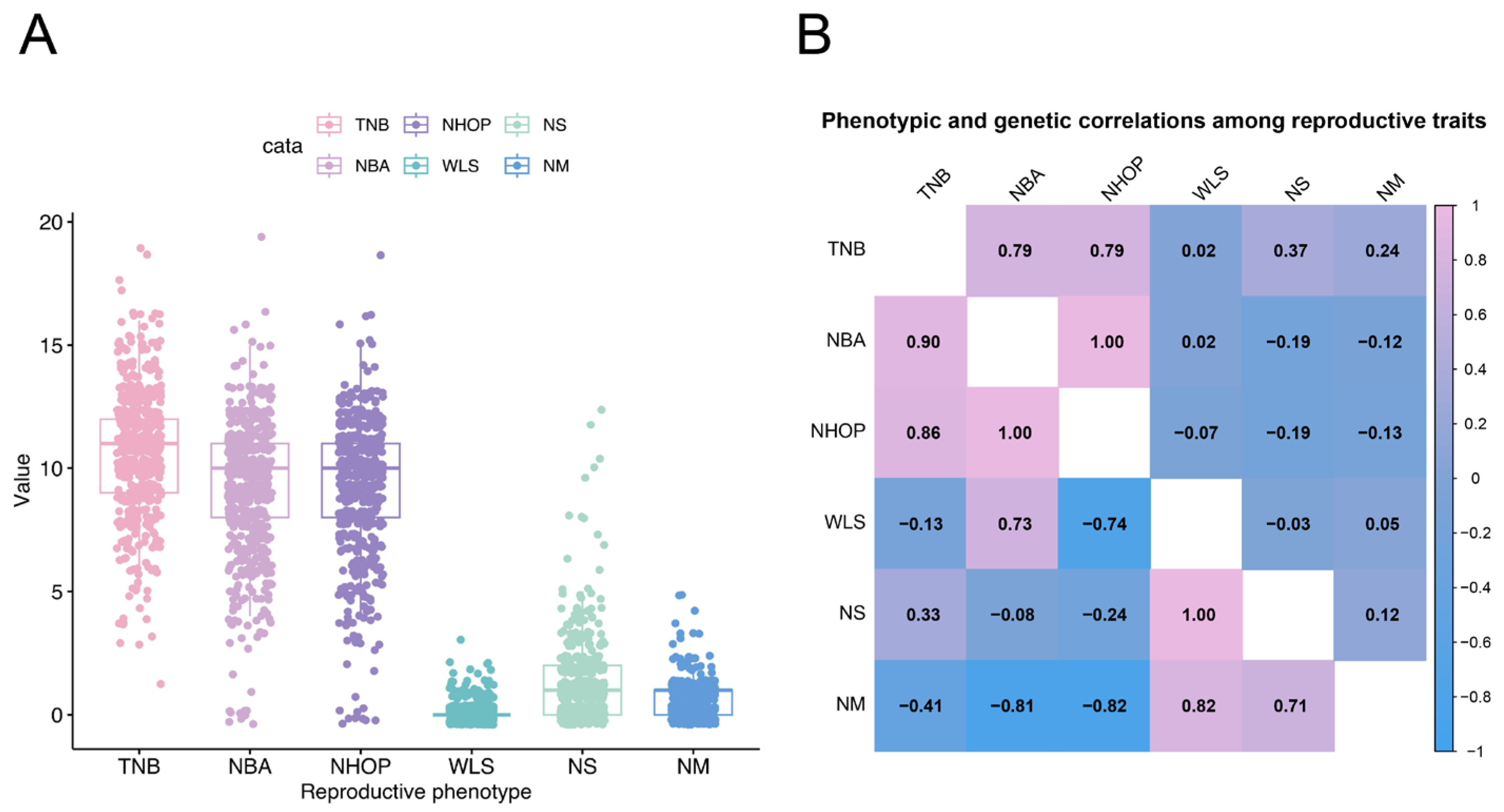
3.1. Phenotypic and Genetic Correlations
3.2. Factors Affecting the Reproductive Performance of Jinwu Sows
3.3. Estimation of Genetic Parameters for Reproductive Traits in Pigs
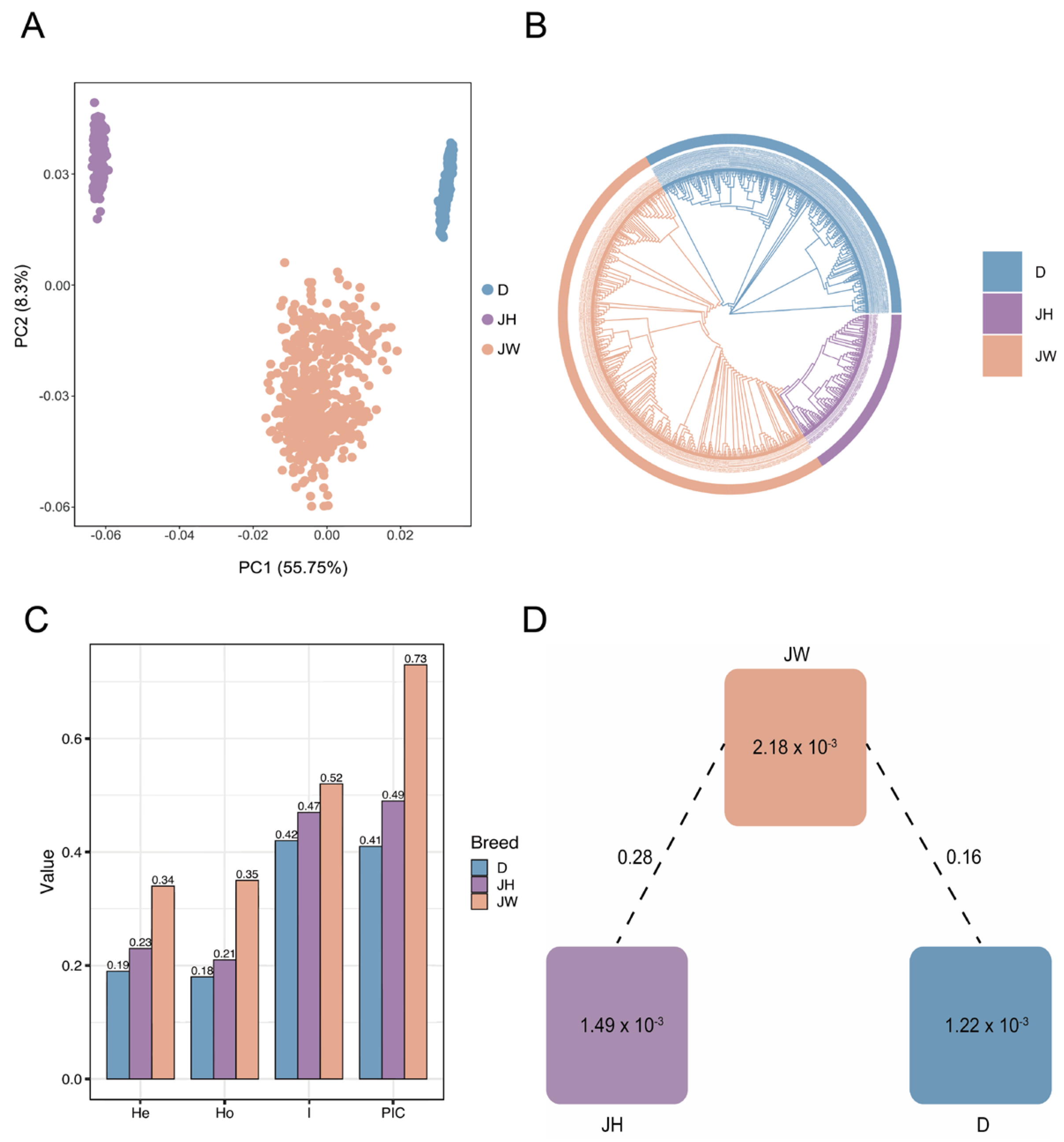
3.4. Principal Component Analysis
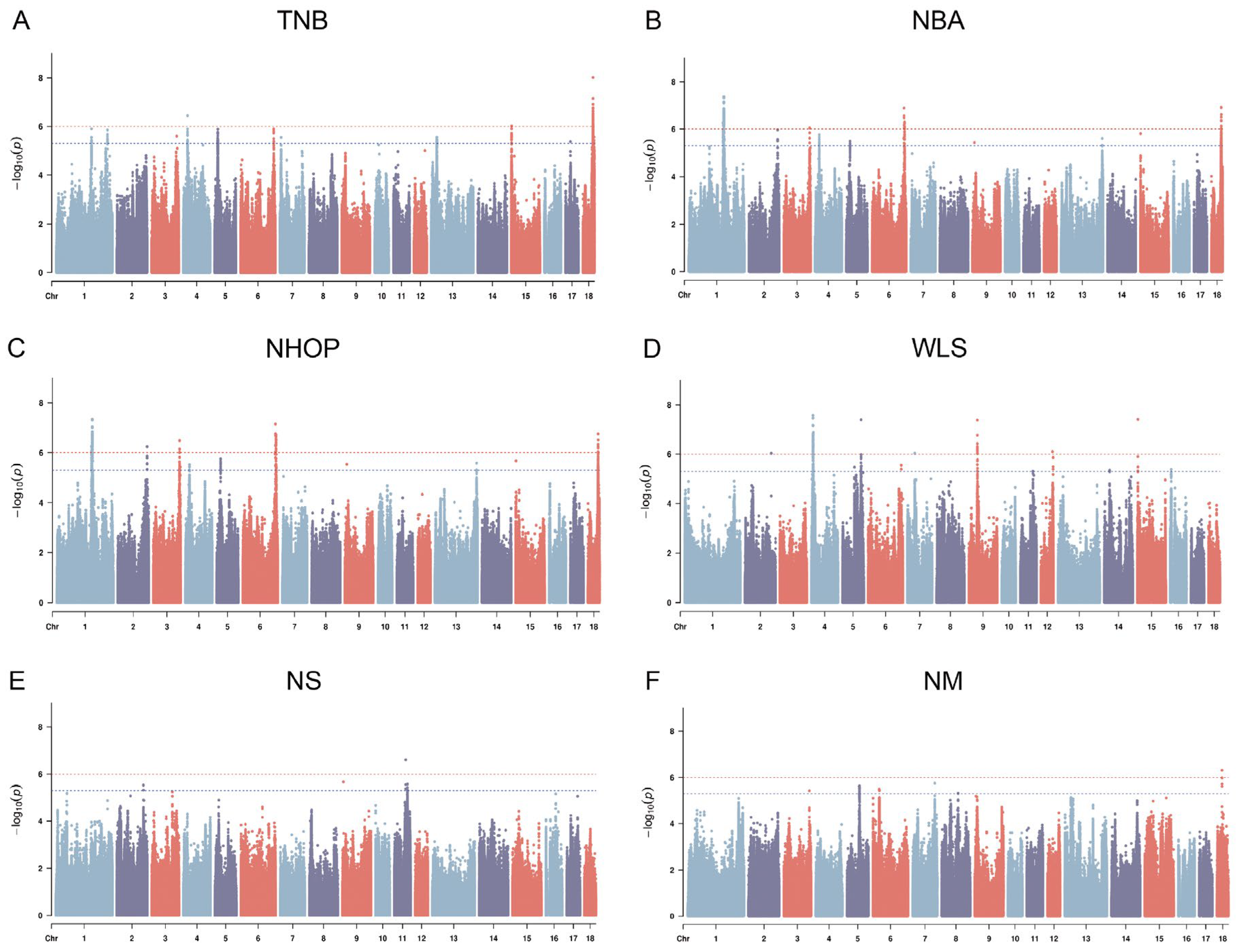
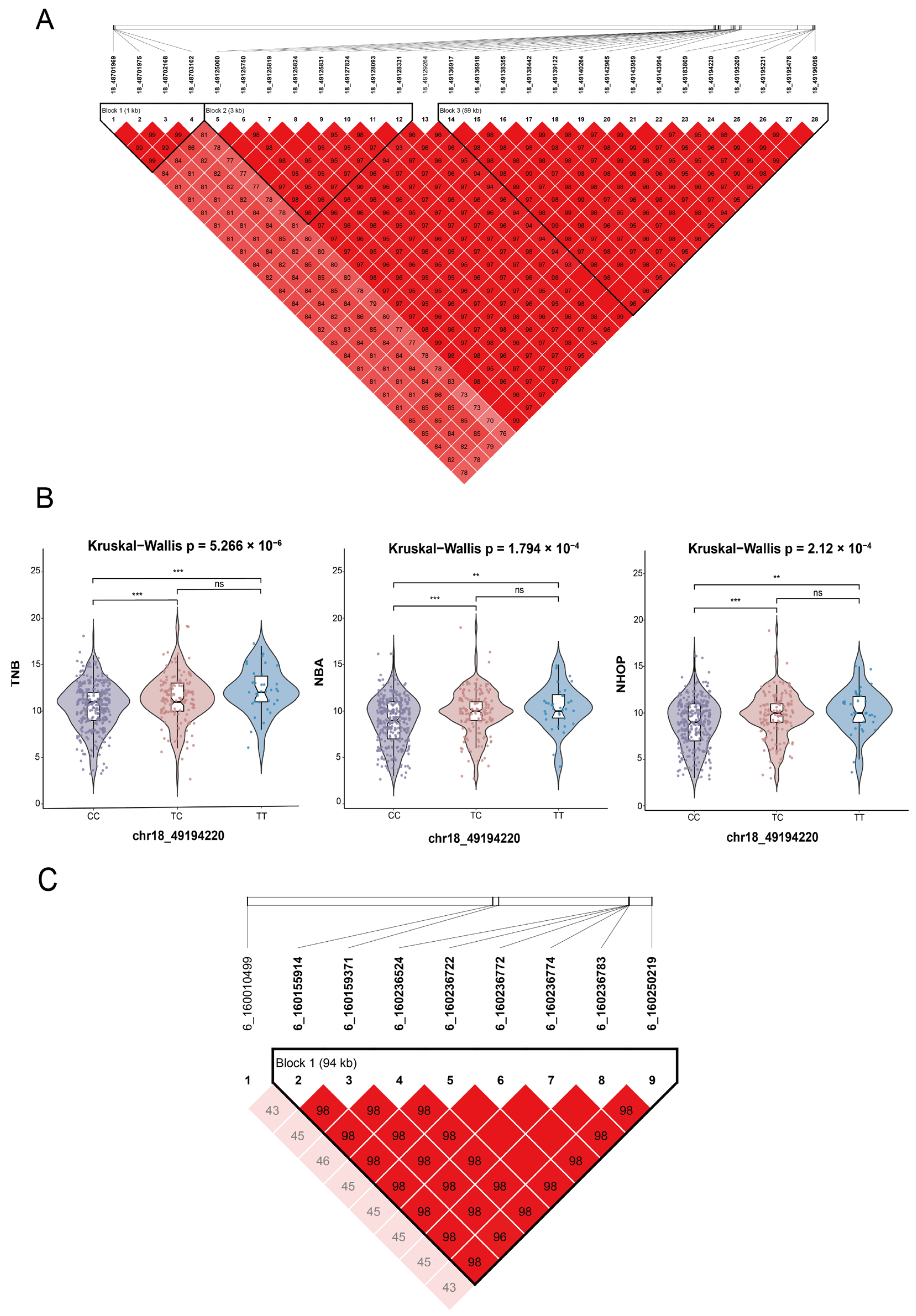
3.5. Genome-Wide Association Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AI-REML | Average Information Restricted Maximum Likelihood |
| GS | Genomic selection |
| GWAS | Genome-wide association study |
| JW | Jinwu pig |
| MAS | Marker-assisted selection |
| NBA | Number of live births |
| NHOP | Number of healthy births |
| NJ | Neighbor-joining |
| NM | Number of mummified fetuses |
| NS | Number of stillbirths |
| PCA | Principal component analysis |
| PIC | Polymorphism information content |
| QTLs | Quantitative trait loci |
| SNP | Single-nucleotide polymorphism |
| TNB | Total number of births |
| WLS | Number of weak births |
References
- Yuan, H.; Wei, W.; Zhang, Y.; Li, C.; Zhao, S.; Chao, Z.; Xia, C.; Quan, J.; Gao, C. Unveiling the Influence of Copy Number Variations on Genetic Diversity and Adaptive Evolution in China’s Native Pig Breeds via Whole-Genome Resequencing. Int. J. Mol. Sci. 2024, 25, 5843. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Gao, C.; Cai, Y.; Ge, Q.; Jiao, T.; Zhao, S. Population genetics assessment model reveals priority protection of genetic resources in native pig breeds in China. Glob. Ecol. Conserv. 2020, 21, e00829. [Google Scholar] [CrossRef]
- Luo, J.; Yang, Y.; Liao, K.; Liu, B.; Chen, Y.; Shen, L.; Chen, L.; Liu, Y.; Li, Q.; Wang, J. Genetic parameter estimation for reproductive traits in QingYu pigs and comparison of carcass and meat quality traits to Berkshire × QingYu crossbred pigs. Asian-Australas. J. Anim. Sci. 2019, 33, 1224. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, R.; Wei, Y.; Meng, X.; Wang, B.; Zhang, Z.; Wu, W.; Liu, H. Effect of MSTN mutation on growth and carcass performance in Duroc × Meishan hybrid population. Animals 2020, 10, 932. [Google Scholar] [CrossRef]
- Ye, J.; Tan, C.; Hu, X.; Wang, A.; Wu, Z. Genetic parameters for reproductive traits at different parities in Large White pigs. J. Anim. Sci. 2018, 96, 1215–1220. [Google Scholar] [CrossRef]
- Meuwissen, T.H.; Hayes, B.J.; Goddard, M. Prediction of total genetic value using genome-wide dense marker maps. genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Verardo, L.L.; Silva, F.F.; Lopes, M.S.; Madsen, O.; Bastiaansen, J.W.; Knol, E.F.; Kelly, M.; Varona, L.; Lopes, P.S.; Guimarães, S.E. Revealing new candidate genes for reproductive traits in pigs: Combining Bayesian GWAS and functional pathways. Genet. Sel. Evol. 2016, 48, 9. [Google Scholar] [CrossRef]
- Zhao, Y.; Gao, G.; Zhou, Y.; Guo, C.; Li, B.; El-Ashram, S.; Li, Z. Genome-wide association studies uncover genes associated with litter traits in the pig. Animal 2022, 16, 100672. [Google Scholar] [CrossRef]
- Uimari, P.; Sironen, A.; Sevón-Aimonen, M.-L. Whole-genome SNP association analysis of reproduction traits in the Finnish Landrace pig breed. Genet. Sel. Evol. 2011, 43, 42. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, Y.; Hou, L.; Ma, J.; Chen, C.; Ai, H.; Huang, L.; Ren, J. Genome-wide detection of genetic markers associated with growth and fatness in four pig populations using four approaches. Genet. Sel. Evol. 2017, 49, 21. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Gong, H.; Cui, L.; Zhang, W.; Ma, J.; Chen, C.; Ai, H.; Xiao, S.; Huang, L. Genetic correlation of fatty acid composition with growth, carcass, fat deposition and meat quality traits based on GWAS data in six pig populations. Meat Sci. 2019, 150, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Qiao, R.; Gao, J.; Zhang, Z.; Li, L.; Xie, X.; Fan, Y.; Cui, L.; Ma, J.; Ai, H.; Ren, J. Genome-wide association analyses reveal significant loci and strong candidate genes for growth and fatness traits in two pig populations. Genet. Sel. Evol. 2015, 47, 17. [Google Scholar] [CrossRef] [PubMed]
- Hérault, F.; Damon, M.; Cherel, P.; Le Roy, P. Combined GWAS and LDLA approaches to improve genome-wide quantitative trait loci detection affecting carcass and meat quality traits in pig. Meat Sci. 2018, 135, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, J.; Xiong, X.; Chen, C.; Xing, Y.; Duan, Y.; Xiao, S.; Yang, B.; Ma, J. An integrative analysis of transcriptome and GWAS data to identify potential candidate genes influencing meat quality traits in pigs. Front. Genet. 2021, 12, 748070. [Google Scholar] [CrossRef]
- Yin, S.; Song, G.; Gao, N.; Gao, H.; Zeng, Q.; Lu, P.; Zhang, Q.; Xu, K.; He, J. Identifying genetic architecture of carcass and meat quality traits in a Ningxiang indigenous pig population. Genes 2023, 14, 1308. [Google Scholar] [CrossRef]
- Yang, Y.; Gan, M.; Yang, X.; Zhu, P.; Luo, Y.; Liu, B.; Zhu, K.; Cheng, W.; Chen, L.; Zhao, Y. Estimation of genetic parameters of pig reproductive traits. Front. Vet. Sci. 2023, 10, 1172287. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Xing, Y.; Li, G.; Wang, Z.; Feng, B.; Song, Z.; Wu, C. GTZ: A fast compression and cloud transmission tool optimized for FASTQ files. BMC Bioinform. 2017, 18, 233–242. [Google Scholar] [CrossRef]
- Rubinacci, S.; Hofmeister, R.J.; Sousa da Mota, B.; Delaneau, O. Imputation of low-coverage sequencing data from 150,119 UK Biobank genomes. Nat. Genet. 2023, 55, 1088–1090. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, s13742-015-0047-8. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, H.; Tang, Z.; Yin, D.; Fu, Y.; Yuan, X.; Li, X.; Liu, X.; Zhao, S. HIBLUP: An integration of statistical models on the BLUP framework for efficient genetic evaluation using big genomic data. Nucleic Acids Res. 2023, 51, 3501–3512. [Google Scholar] [CrossRef] [PubMed]
- VanRaden, P.M. Efficient methods to compute genomic predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X. rMVP: A memory-efficient, visualization-enhanced, and parallel-accelerated tool for genome-wide association study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- McLaren, W.; Pritchard, B.; Rios, D.; Chen, Y.; Flicek, P.; Cunningham, F. Deriving the consequences of genomic variants with the Ensembl API and SNP Effect Predictor. Bioinformatics 2010, 26, 2069–2070. [Google Scholar] [CrossRef]
- Fonseca, P.A.; Suarez-Vega, A.; Marras, G.; Cánovas, Á. GALLO: An R package for genomic annotation and integration of multiple data sources in livestock for positional candidate loci. Gigascience 2020, 9, giaa149. [Google Scholar] [CrossRef]
- Holsinger, K.E.; Weir, B.S. Genetics in geographically structured populations: Defining, estimating and interpreting FST. Nat. Rev. Genet. 2009, 10, 639–650. [Google Scholar] [CrossRef]
- Ding, R.; Qiu, Y.; Zhuang, Z.; Ruan, D.; Wu, J.; Zhou, S.; Ye, J.; Cao, L.; Hong, L.; Xu, Z.; et al. Genome-wide association studies reveals polygenic genetic architecture of litter traits in Duroc pigs. Theriogenology 2021, 173, 269–278. [Google Scholar] [CrossRef]
- Yu, G.; Wang, C.; Wang, Y. Genetic parameter analysis of reproductive traits in Large White pigs. Anim. Biosci. 2022, 35, 1649. [Google Scholar] [CrossRef]
- Zheng, C.; Wang, Y.; Huang, X.; Wu, S.; Bao, W. A genome-wide association study of important reproduction traits in large white pigs. Gene 2022, 838, 146702. [Google Scholar]
- Piñán, J.; Alegre, B.; Kirkwood, R.N.; Soriano-Úbeda, C.; Maj, M.; Domínguez, J.C.; Manjarín, R.; Martínez-Pastor, F. Effect of season and parity on reproduction perforzmance of Iberian sows bred with Duroc semen. Animals 2021, 11, 3275. [Google Scholar] [CrossRef]
- Knecht, D.; Środoń, S.; Duziński, K. The impact of season, parity and breed on selected reproductive performance parameters of sows. Arch. Anim. Breed. 2015, 58, 49–56. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, L.-G.; Shi, H.-B.; Hua, Y.; Zhang, L.-C.; Xin, L.; Lei, P.; Liang, J.; Zhang, Y.-B.; Zhao, K.-b. Hritabilities and genetic and phenotypic correlations of litter uniformity and litter size in Large White sows. J. Integr. Agric. 2016, 15, 848–854. [Google Scholar] [CrossRef]
- Hoving, L.; Soede, N.; Graat, E.; Feitsma, H.; Kemp, B. Reproductive performance of second parity sows: Relations with subsequent reproduction. Livest. Sci. 2011, 140, 124–130. [Google Scholar] [CrossRef]
- Bertoldo, M.; Holyoake, P.; Evans, G.; Grupen, C. Seasonal variation in the ovarian function of sows. Reprod. Fertil. Dev. 2012, 24, 822–834. [Google Scholar] [CrossRef]
- Peltoniemi, O.T.; Virolainen, J.V. Seasonality of reproduction in gilts and sows. Reprod. Suppl. 2006, 62, 205–218. [Google Scholar] [CrossRef]
- Auvigne, V.; Leneveu, P.; Jehannin, C.; Peltoniemi, O.; Sallé, E. Seasonal infertility in sows: A five year field study to analyze the relative roles of heat stress and photoperiod. Theriogenology 2010, 74, 60–66. [Google Scholar] [CrossRef]
- Kim, M.-A.; Lee, E.-J.; Yang, W.; Shin, H.-Y.; Kim, Y.-H.; Kim, J.-H. Identification of a novel gene signature in second-trimester amniotic fluid for the prediction of preterm birth. Sci. Rep. 2022, 12, 3085. [Google Scholar] [CrossRef]
- Tixier, V.; Bataillé, L.; Etard, C.; Jagla, T.; Weger, M.; DaPonte, J.P.; Strähle, U.; Dickmeis, T.; Jagla, K. Glycolysis supports embryonic muscle growth by promoting myoblast fusion. Proc. Natl. Acad. Sci. USA 2013, 110, 18982–18987. [Google Scholar] [CrossRef]
- Everson, T.M.; O’Shea, T.M.; Burt, A.; Hermetz, K.; Carter, B.S.; Helderman, J.; Hofheimer, J.A.; McGowan, E.C.; Neal, C.R.; Pastyrnak, S.L. Serious neonatal morbidities are associated with differences in DNA methylation among very preterm infants. Clin. Epigenet. 2020, 12, 151. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, H.Q.; Tang, M.; Guo, C.; Liu, S.; Li, J.; Wang, Y.; Xiao, L.; Yang, G. Abnormal TNS3 gene methylation in patients with congenital scoliosis. BMC Musculoskelet. Disord. 2022, 23, 797. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, Y.; Jiang, X.; Wu, Q.; Lin, R.; Chen, H.; Zhang, M.; Zeng, T.; Tian, Y.; Xu, E.; et al. Genome-wide association study identified candidate genes for egg production traits in the Longyan Shan-ma duck. Poult. Sci. 2024, 103, 104032. [Google Scholar] [CrossRef]
- Maerz, L.D.; Casar Tena, T.; Gerhards, J.; Donow, C.; Jeggo, P.A.; Philipp, M. Analysis of cilia dysfunction phenotypes in zebrafish embryos depleted of Origin recognition complex factors. Eur. J. Hum. Genet. 2019, 27, 772–782. [Google Scholar] [CrossRef]
- Zhong, T.; Hou, D.; Zhao, Q.; Zhan, S.; Wang, L.; Li, L.; Zhang, H.; Zhao, W.; Yang, S.; Niu, L. Comparative whole-genome resequencing to uncover selection signatures linked to litter size in Hu Sheep and five other breeds. Bmc Genom. 2024, 25, 480. [Google Scholar] [CrossRef]
- Li, X.; Ye, J.; Han, X.; Qiao, R.; Li, X.; Lv, G.; Wang, K. Whole-genome sequencing identifies potential candidate genes for reproductive traits in pigs. Genomics 2020, 112, 199–206. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, H.; Shao, F.; Zhang, Y.; Nie, H.; Zhang, J.; Li, C.; Hou, Z.; Chen, Z.-J.; Wang, J.; et al. Remodeling of maternal mRNA through poly (A) tail orchestrates human oocyte-to-embryo transition. Nat. Struct. Mol. Biol. 2023, 30, 200–215. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, C.; Huo, J.; Zhao, S. Expression and genomic imprinting of DCN, PON2 and PEG3 genes in porcine placenta. Anim. Reprod. Sci. 2011, 123, 70–74. [Google Scholar] [CrossRef]
- Eckler, M.J.; Chen, B. Fez family transcription factors: Controlling neurogenesis and cell fate in the developing mammalian nervous system. BioEssays 2014, 36, 788–797. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait 1 | Litters 2 | CV (%) 3 | Mean | SD | Max | Min |
|---|---|---|---|---|---|---|
| TNB | 2831 | 21.49 | 11.65 | 2.5 | 24 | 3 |
| NBA | 2809 | 21.49 | 10.15 | 2.18 | 19 | 3 |
| NHOP | 2808 | 21.58 | 10.12 | 2.18 | 19 | 3 |
| WLS | 2831 | 709.56 | 0.03 | 0.21 | 3 | 0 |
| NS | 2831 | 119.16 | 1.08 | 1.29 | 12 | 0 |
| NM | 2831 | 152.68 | 0.48 | 0.74 | 7 | 0 |
| Trait 1 | Boar 2 | Season | Parity | |||
|---|---|---|---|---|---|---|
| df | F | df | F | df | F | |
| TNB | 40 | 1.39 | 3 | 4 ** | 6 | 14.14 *** |
| NBA | 40 | 2.06 *** | 3 | 3.81 ** | 6 | 13.87 *** |
| NHOP | 40 | 2.23 *** | 3 | 3.88 ** | 6 | 16.1 *** |
| WLS | 40 | 1.42 * | 3 | 11.09 *** | 6 | 11.48 *** |
| NS | 40 | 1.19 | 3 | 22.84 *** | 6 | 7.38 *** |
| NM | 40 | 1.45 * | 3 | 3.19 * | 6 | 7.05 *** |
| Trait 1 | Spring | Summer | Autumn | Winter |
|---|---|---|---|---|
| TNB | 11.78 ± 2.6 a | 11.76 ± 2.41 a | 11.69 ± 2.46 ab | 11.34 ± 2.52 b |
| NBA | 9.8 ± 2.52 a | 10.3 ± 2.13 a | 10.28 ± 2.24 b | 9.92 ± 2.38 b |
| NHOP | 9.77 ± 2.52 a | 10.27 ± 2.12 a | 10.28 ± 2.25 b | 9.84 ± 2.4 b |
| WLS | 0.02 ± 0.18 a | 0.03 ± 0.25 b | 0 ± 0 bc | 0.07 ± 0.3 c |
| NS | 1.4 ± 1.59 a | 1.01 ± 1.16 b | 0.93 ± 1.11 b | 0.96 ± 1.17 b |
| NM | 0.56 ± 0.76 a | 0.45 ± 0.78 ab | 0.46 ± 0.78 b | 0.45 ± 0.6 b |
| Parity | |||||||
|---|---|---|---|---|---|---|---|
| Trait 1 Litters 2 | 1st 516 | 2nd 494 | 3rd 442 | 4th 357 | 5th 319 | 6th 289 | 7th 245 |
| TNB | 10.85 ± 2.54 a | 11.31 ± 2.64 a | 11.72 ± 2.42 a | 12.07 ± 2.56 a | 11.91 ± 2.29 ab | 12.18 ± 2.29 bc | 12.16 ± 2.38 c |
| NBA | 9.34 ± 2.53 a | 10.19 ± 2.37 a | 10.36 ± 2.07 a | 10.39 ± 2.18 a | 10.32 ± 1.77 a | 10.45 ± 1.8 a | 10.38 ± 1.9 b |
| NHOP | 9.24 ± 2.53 a | 10.18 ± 2.36 a | 10.36 ± 2.07 a | 10.35 ± 2.17 a | 10.29 ± 1.76 a | 10.45 ± 1.8 a | 10.34 ± 1.9 b |
| WLS | 0.1 ± 0.36 a | 0.01 ± 0.12 b | 0 ± 0 b | 0.03 ± 0.25 b | 0.02 ± 0.2 b | 0 ± 0b | 0.04 ± 0.25 b |
| NS | 1.11 ± 1.68 a | 0.8 ± 1.08 a | 1 ± 1.19 a | 1.24 ± 1.13 a | 1.13 ± 1.16 a | 1.2 ± 1.09 ab | 1.19 ± 1.34 b |
| NM | 0.6 ± 0.71 a | 0.4 ± 0.74 a | 0.38 ± 0.76 ab | 0.47 ± 0.65 abc | 0.45 ± 0.7 abc | 0.55 ± 0.84 bc | 0.61 ± 0.81 c |
| Traits 1 | 2 | 3 | 4 | h2 (SE) 5 |
|---|---|---|---|---|
| TNB | 0.5457 | 0.2733 | 5.2795 | 0.0894 (0.0256) |
| NBA | 0.4866 | 0.1206 | 4.6074 | 0.0918 (0.0240) |
| NHOP | 0.4755 | 0.1183 | 4.5939 | 0.0895 (0.0237) |
| WLS | 0.0003 | <0.01 | 0.0402 | 0.0085 (0.0082) |
| NS | 0.0046 | 0.0883 | 1.5071 | 0.0029 (0.0068) |
| NM | 0.0073 | < 0.01 | 0.5295 | 0.0136 (0.0102) |
| SSC 1 | Related Trait 2 | SNP 3 | Position | Corresponding p_Wald | Candidate Gene |
|---|---|---|---|---|---|
| 18 | TNB/NBA/NHOP | chr18_49194220 (rs326174997) | 49,194,220 | 9.57 × 10−9 2.42 × 10−7/3.05 × 10−7 | TNS3 |
| 18 | TNB/NBA/NHOP | chr18_48701969 (rs81233849) | 48,701,969 | 3.56 × 10−7 1.25 × 10−7/1.76 × 10−7 | Vopp1/PGAM2 |
| 1 | NBA/NHOP | chr1_172136167 (rs80793150) | 172,136,167 | 4.28 × 10−8/4.53 × 10−8 | LRFN5 |
| 6 | NBA/NHOP | chr6_160159371 (rs332416322) | 160,159,371 | 5.17 × 10−7 7.10 × 10−8 | TUT4/ORC1/CC2D1B/ZFYVE9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Zhao, A.; Pan, J.; Tan, K.; Zhu, Z.; Zhang, L.; Yu, F.; Liu, R.; Zhong, L.; Huang, J. Statistical Analysis of Reproductive Traits in Jinwu Pig and Identification of Genome-Wide Association Loci. Genes 2025, 16, 550. https://doi.org/10.3390/genes16050550
Chen W, Zhao A, Pan J, Tan K, Zhu Z, Zhang L, Yu F, Liu R, Zhong L, Huang J. Statistical Analysis of Reproductive Traits in Jinwu Pig and Identification of Genome-Wide Association Loci. Genes. 2025; 16(5):550. https://doi.org/10.3390/genes16050550
Chicago/Turabian StyleChen, Wenduo, Ayong Zhao, Jianzhi Pan, Kai Tan, Zhiwei Zhu, Liang Zhang, Fuxian Yu, Renhu Liu, Liepeng Zhong, and Jing Huang. 2025. "Statistical Analysis of Reproductive Traits in Jinwu Pig and Identification of Genome-Wide Association Loci" Genes 16, no. 5: 550. https://doi.org/10.3390/genes16050550
APA StyleChen, W., Zhao, A., Pan, J., Tan, K., Zhu, Z., Zhang, L., Yu, F., Liu, R., Zhong, L., & Huang, J. (2025). Statistical Analysis of Reproductive Traits in Jinwu Pig and Identification of Genome-Wide Association Loci. Genes, 16(5), 550. https://doi.org/10.3390/genes16050550