
Genetic Contributions to Aggressive Behaviour in Pigs: A Comprehensive Review
 ,
,  , , and
, , and
Abstract
1. Introduction
2. The Issue of Pig Aggression and Its Repercussions on the Efficacy of Agricultural Production
Definition, Detection, and Classification of Pig Aggressiveness
- (1)
- Social aggression is defined as a form of aggression that occurs within a social framework. It is frequently associated with the establishment of a dominance hierarchy between individuals. It has been particularly prevalent in the context of pig interactions, particularly within group housing systems. The manifestations of social aggression may include physical confrontations such as fighting, biting, or pushing.
- (2)
- Maternal aggression is considered as the display of behaviour directed toward the protection of offspring from potential threats. This behaviour is critical for the survival of the piglets and may include aggressive postures, loud cries, and physical attacks on other individuals or humans.
- (3)
- The phenomenon of fear-induced aggression has been observed in pigs, with evidence indicating that these animals exhibit aggressive behaviour in conditions and situations associated with fear caused by threats from other individuals or humans or from stressors such as loud noises and the presence of unfamiliar animals.
- (4)
- Aggressive behaviour has been reported in young piglets during non-stressful interactions, such as play, where imitations of confrontations and pursuits have been noted. The significance of this behaviour lies in its role in the development of social skills and the understanding of social interactions.
- (5)
- Redirected aggression is defined as a variant of aggressive behaviour, in which a pig fails to express aggression towards an intended target and instead redirects it towards another pig or object. This phenomenon has been observed in contexts involving confined animals or those exposed to stressful situations.
3. Physiological Markers Associated with the Manifestation of Aggression
Environmental Factors (Addition of Novel Animal Groups, Limited Farm Space)
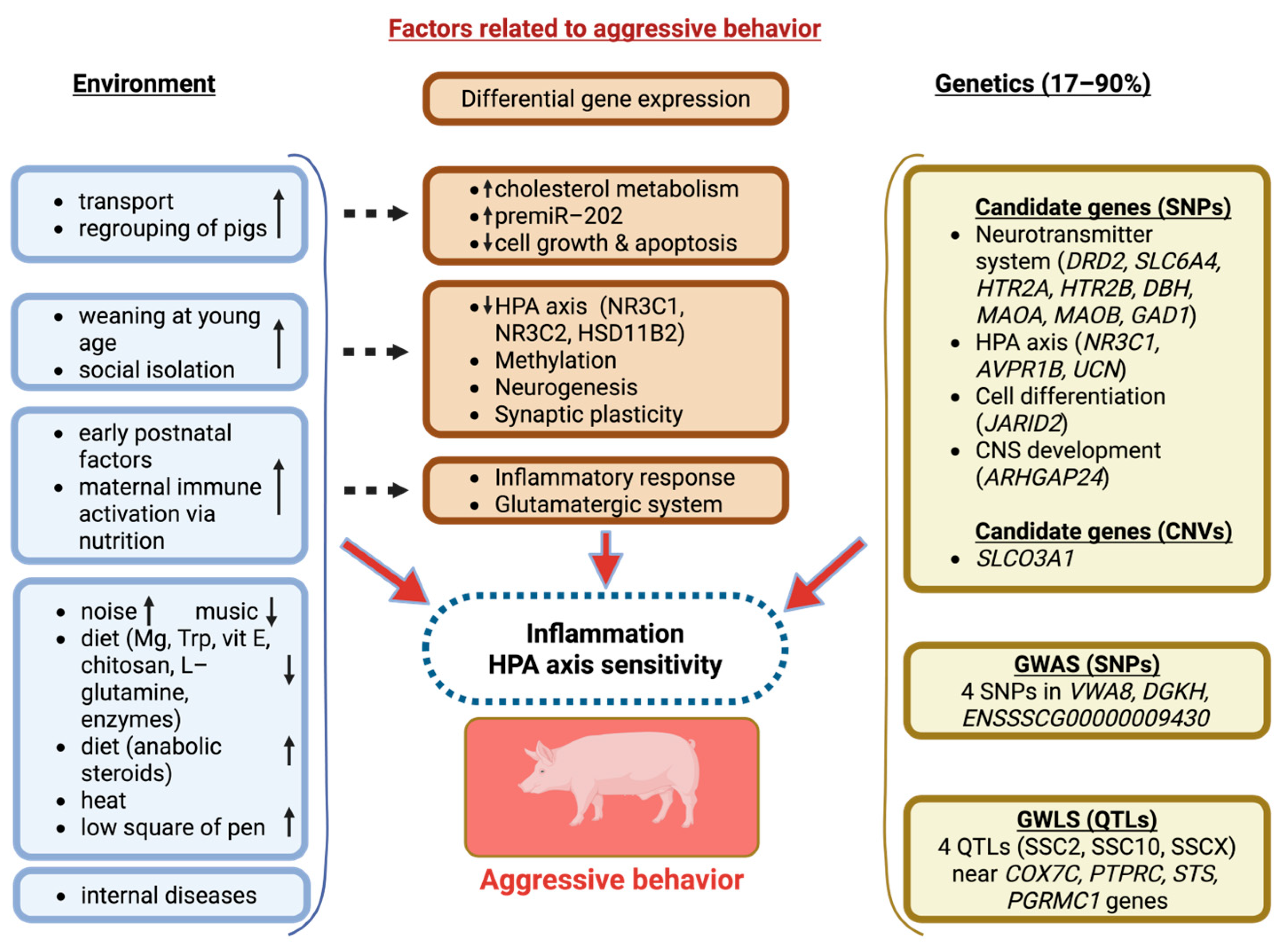
4. Genetic Predisposition and Genetic Markers of Aggression
4.1. Candidate Gene Studies of Aggression in Pigs
4.2. Genome-Wide Linkage Studies of Aggression in Pigs
4.3. Genome-Wide Association Studies of Aggression in Pigs
4.4. Gene Transcription as a Marker of Aggressive Behaviour in Pigs
5. The Link Between Environmental-Induced Epigenetic Changes and Aggressive Behaviour
6. Problems of Strategy Implementation
Future Prospects
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations—With Major Processing by Our World in Data. 2023. Available online: https://ourworldindata.org/grapher/beef-and-buffalo-meat-production-tonnes (accessed on 2 February 2025).
- Li, J.; Topaz, M.; Xun, W.; Li, W.; Wang, X.; Liu, H.; Yuan, Y.; Chen, S.; Li, Y.; Li, X. New swine model of infected soft tissue blast injury. J. Trauma Acute Care Surg. 2012, 73, 908–913. [Google Scholar] [CrossRef]
- Li, K.; Wagner, L.; Moctezuma-Ramirez, A.; Vela, D.; Perin, E. A Robust Percutaneous Myocardial Infarction Model in Pigs and Its Effect on Left Ventricular Function. J. Cardiovasc. Transl. Res. 2021, 14, 1075–1084. [Google Scholar] [CrossRef]
- Hildebrand, F.; Andruszkow, H.; Huber-Lang, M.; Pape, H.C.; van Griensven, M. Combined hemorrhage/trauma models in pigs-current state and future perspectives. Shock 2013, 40, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Jaishankar, R.; Teichmann, D.; Hayward, A.; Holsapple, J.W.; Heldt, T. Open cranium model for the study of cerebrovascular dynamics in intracranial hypertension. J. Neurosci. Methods 2024, 409, 110196. [Google Scholar] [CrossRef] [PubMed]
- Racewicz, P.; Ludwiczak, A.; Skrzypczak, E.; Składanowska-Baryza, J.; Biesiada, H.; Nowak, T.; Nowaczewski, S.; Zaborowicz, M.; Stanisz, M.; Ślósarz, P. Welfare Health and Productivity in Commercial Pig Herds. Animals 2021, 11, 1176. [Google Scholar] [CrossRef] [PubMed]
- Camerlink, I.; Chou, J.Y.; Turner, S.P. Intra-Group Lethal Gang Aggression in Domestic Pigs (Sus scrofa domesticus). Animals 2020, 10, 1287. [Google Scholar] [CrossRef]
- Peden, R.S.E.; Turner, S.P.; Camerlink, I.; Akaichi, F. An estimation of the financial consequences of reducing pig aggression. PLoS ONE 2021, 16, e0250556. [Google Scholar] [CrossRef]
- Jensen, P. The Ethology of Domestic Animals: An Introductory Text, 3rd ed.; Behavior of pigs; CABI Publishing: Linkoping, Sweden, 2002; pp. 159–172. ISBN 978-1-78639-165-0. [Google Scholar]
- Schaefer, A.L.; Salomons, M.O.; Tong, A.K.W.; Soather, A.P.; Lepage, P. The effect of environment enrichment on aggression in newly weaned pigs. Appl. Anim. Behav. Sci. 1990, 27, 41–52. [Google Scholar] [CrossRef]
- Camerlink, I.; Turner, S.P.; Ursinus, W.W.; Reimert, I.; Bolhuis, J.E. Aggression and affiliation during social conflict in pigs. PLoS ONE 2014, 9, e113502. [Google Scholar] [CrossRef]
- Fielding, D. Human–Livestock Interactions—The Stockperson and the Productivity and Welfare of Intensively Farmed Animals. P.H. Hemsworth and G.J. Coleman. Vet. Res. Commun. 1999, 23, 264. [Google Scholar] [CrossRef]
- Van de Weerd, H.A.; Docking, C.M.; Day, J.E.L.; Edwards, S.A. The development of harmful social behaviour in pigs with intact tails and different enrichment backgrounds in two housing systems. Anim. Sci. 2005, 80, 289–298. [Google Scholar] [CrossRef]
- Available online: https://www.merckvetmanual.com/behavior/normal-social-behavior-and-behavioral-problems-of-domestic-animals/behavioral-problems-of-swine (accessed on 2 February 2025).
- Marchant, J.N.; Marchant-Forde, R.M. Methods to reduce aggression at mixing in swine. Pig News Inf. 2005, 26, 63–73. [Google Scholar]
- Henry, M.; Jansen, H.; Amezcua, M.D.R.; O’Sullivan, T.L.; Niel, L.; Shoveller, A.K.; Friendship, R.M. Tail-Biting in Pigs: A Scoping Review. Animals 2021, 11, 2002. [Google Scholar] [CrossRef]
- Wurtz, K.; Camerlink, I.; D’Eath, R.B.; Fernandez, A.P.; Norton, T.; Steibel, J.; Siegford, J. Recording behaviour of indoor-housed farm animals automatically using machine vision technology: A systematic review. PLoS ONE 2019, 14, e0226669. [Google Scholar] [CrossRef]
- Ji, H.; Teng, G.; Yu, J.; Wen, Y.; Deng, H.; Zhuang, Y. Efficient Aggressive Behavior Recognition of Pigs Based on Temporal Shift Module. Animals 2023, 13, 2078. [Google Scholar] [CrossRef] [PubMed]
- Oczak, M.; Viazzi, S.; Ismayilova, G.; Sonoda, L.T.; Roulston, N.; Fels, M.; Bahr, C.; Hartung, J.; Guarino, M.; Berckmans, D.; et al. Classification of aggressive behaviour in pigs by activity index and multilayer feed forward neural network. Biosyst. Eng. 2014, 119, 89–97. [Google Scholar] [CrossRef]
- Wang, Z.; Xiao, Z.; Li, H.; Shi, Z.; Pu, S. Behavioral Transition Path and Pivotal Nodes Regulating Attack in Initial Encounters between Unfamiliar Pigs. Animals 2024, 14, 2595. [Google Scholar] [CrossRef]
- Taylor, N.R.; Main, D.C.J.; Mendl, M.; Edwards, S.A. Tail-biting: A new perspective. TVJ 2010, 186, 137–147. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Z.; Guo, H.; Qin, J.; Zhang, X. Changes in Aggressive Behavior, Cortisol and Brain Monoamines during the Formation of Social Hierarchy in Black Rockfish (Sebastes schlegelii). Animals 2020, 10, 2357. [Google Scholar] [CrossRef]
- Knezevic, E.; Nenic, K.; Milanovic, V.; Knezevic, N.N. The Role of Cortisol in Chronic Stress, Neurodegenerative Diseases, and Psychological Disorders. Cells 2023, 12, 2726. [Google Scholar] [CrossRef]
- D’Eath, R.B.; Turner, S.P.; Kurt, E.; Evans, G.; Thölking, L.; Looft, H.; Wimmers, K.; Murani, E.; Klont, R.; Foury, A.; et al. Pigs’ aggressive temperament affects pre-slaughter mixing aggression, stress and meat quality. Animal 2010, 4, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Papatsiros, V.G.; Maragkakis, G.; Papakonstantinou, G.I. Stress Biomarkers in Pigs: Current Insights and Clinical Application. Vet. Sci. 2024, 11, 640. [Google Scholar] [CrossRef] [PubMed]
- Ursinus, W.W.; Van Reenen, C.G.; Reimert, I.; Bolhuis, J.E. Tail biting in pigs: Blood serotonin and fearfulness as pieces of the puzzle. PLoS ONE 2014, 9, e107040. [Google Scholar] [CrossRef]
- Lay, D.C.; Enneking, S.A.; Anderson, N.C.; Richert, B.T.; Sapkota, A. Effect of short-term high tryptophan diet fed to sows on their subsequent piglet behavior. Transl. Anim. Sci. 2021, 5, txab127. [Google Scholar] [CrossRef] [PubMed]
- Skripkin, V.S.; Kvochko, A.N.; Sidelnikov, A.; Pisarenko, N.A.; Krivoruchko, A. Morphological and Functional Activity Dynamics of Blood Lymphocytes in Large White Breed Pigs in Postnatal Ontogenesis and during Pregnancy. IOP Conf. Ser. Earth Environ. Sci. 2021, 852, 012100. [Google Scholar] [CrossRef]
- Urban, G.; Krotova, O.; Polozyuk, O.; Zabelina, M.; Katusov, D.; Manzhikova, A. Morpho-biochemical parameters of blood and antioxidant protection of the body of repair pigs using natural metabolites. BIO Web Conf. 2024, 113, 10.1051. [Google Scholar] [CrossRef]
- Hari, J.J.; Pliska, V. Hypothalamo-pituitary-adrenocortical axis in stress-susceptible and stress-resistant pigs: Endocrine responses to corticotrophin-releasing factor and vasopressin. J. Anim. Breed. Genet. 2005, 122 (Suppl. 1), 87–96. [Google Scholar] [CrossRef]
- Nian, H.; Ding, S.; Feng, Y.; Liu, H.; Li, J.; Li, X.; Zhang, R.; Bao, J. Effect of Noise and Music on Neurotransmitters in the Amygdala: The Role Auditory Stimuli Play in Emotion Regulation. Metabolites 2023, 13, 928. [Google Scholar] [CrossRef]
- Alvarez-Hernandez, N.; Vallejo-Timarán, D.; de Jesús Rodriguez, B. Adapted Original Music as an Environmental Enrichment in an Intensive Pig Production System Reduced Aggression in Weaned Pigs during Regrouping. Animals 2023, 13, 3599. [Google Scholar] [CrossRef]
- Ciborowska, P.; Michalczuk, M.; Bień, D. The Effect of Music on Livestock: Cattle, Poultry and Pigs. Animals 2021, 11, 3572. [Google Scholar] [CrossRef]
- Zapata Cardona, J.; Ceballos, M.C.; Tarazona Morales, A.M.; David Jaramillo, E.; Rodríguez, B.J. Music modulates emotional responses in growing pigs. Sci. Rep. 2022, 12, 3382. [Google Scholar] [CrossRef] [PubMed]
- Burrows, M.S.; Moss, B.W.; Beattie, V.E. The role of dietary manipulation in the control of aggression in pigs. Biochem. Soc. Trans. 1998, 26, S58. [Google Scholar] [CrossRef]
- Chen, F.; Yang, L.; Zhe, L.; Jlali, M.; Zhuo, Y.; Jiang, X.; Huang, L.; Wu, F.; Zhang, R.; Xu, S.; et al. Supplementation of a Multi-Carbohydrase and Phytase Complex in Diets Regardless of Nutritional Levels, Improved Nutrients Digestibility, Growth Performance, and Bone Mineralization of Growing-Finishing Pigs. Animals 2023, 13, 1557. [Google Scholar] [CrossRef] [PubMed]
- Parois, S.P.; Duttlinger, A.W.; Richert, B.T.; Lindemann, S.R.; Johnson, J.S.; Marchant-Forde, J.N. Effects of Three Distinct 2-Week Long Diet Strategies After Transport on Weaned Pigs’ Short and Long-Term Welfare Markers, Behaviors, and Microbiota. Front. Vet. Sci. 2020, 7, 140. [Google Scholar] [CrossRef]
- Peeters, E.; Neyt, A.; Beckers, F.; De Smet, S.; Aubert, A.E.; Geers, R. Influence of supplemental magnesium, tryptophan, vitamin C, and vitamin E on stress responses of pigs to vibration. J. Anim. Sci. 2005, 83, 1568–1580. [Google Scholar] [CrossRef]
- O’Driscoll, K.; O’Gorman, D.M.; Taylor, S.; Boyle, L.A. The influence of a magnesium-rich marine extract on behaviour, salivary cortisol levels and skin lesions in growing pigs. Animal 2013, 7, 1017–1027. [Google Scholar] [CrossRef]
- Li, Y.Z.; Baidoo, S.K.; Johnston, L.J.; Anderson, J.E. Effects of tryptophan supplementation on aggression among group-housed gestating sows. J. Anim. Sci. 2011, 89, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Hu, X.M.; Zhang, T.; Wei, H.K.; Zhou, Y.F.; Zhou, Z.X.; Peng, J. Effects of dietary oregano essential oil and vitamin E supplementation on meat quality, stress response and intestinal morphology in pigs following transport stress. J. Vet. Med. Sci. 2017, 79, 328–335. [Google Scholar] [CrossRef]
- Skoupá, K.; Šťastný, K.; Sládek, Z. Anabolic Steroids in Fattening Food-Producing Animals—A Review. Animals 2022, 12, 2115. [Google Scholar] [CrossRef]
- Abuajamieh, M.; Kvidera, S.K.; Mayorga, E.J.; Kaiser, A.; Lei, S.; Seibert, J.T.; Horst, E.A.; Sanz Fernandez, M.V.; Ross, J.W.; Selsby, J.T.; et al. The effect of recovery from heat stress on circulating bioenergetics and inflammatory biomarkers. J. Anim. Sci. 2018, 96, 4599–4610. [Google Scholar] [CrossRef]
- Whittaker, A.L.; van Wettere, W.H.; Paul, E. Hughes Space Requirements to Optimize Welfare and Performance in Group Housed Pigs—A Review. Am. J. Anim. Vet. Sci. 2012, 7, 48–54. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, K.S.; Kim, J.E.; Kim, D.W.; Seol, K.H.; Lee, S.H.; Chae, B.J.; Kim, Y.H. The effect of optimal space allowance on growth performance and physiological responses of pigs at different stages of growth. Animal 2017, 11, 478–485. [Google Scholar] [CrossRef]
- Song, D.C.; Lee, J.H.; Yun, W.; Chang, S.Y.; Park, S.H.; Jeon, K.H.; Kim, H.; Cho, J.H. Effects of Stocking Density and Illuminance in Lairage of Fattening Pigs in Different Temperatures. Animals 2024, 14, 2145. [Google Scholar] [CrossRef] [PubMed]
- Boyle, L.A.; Edwards, S.A.; Bolhuis, J.E.; Pol, F.; Šemrov, M.Z.; Schütze, S.; Nordgreen, J.; Bozakova, N.; Sossidou, E.N.; Valros, A. The Evidence for a Causal Link Between Disease and Damaging Behavior in Pigs. Front. Vet. Sci. 2021, 8, 771682. [Google Scholar] [CrossRef]
- Lovendahl, P.; Damgaard, L.H.; Nielsen, B.L.; Thodberg, K.; Su, G.; Rydhmer, L. Aggressive behaviour of sows at mixing and maternal behaviour are heritable and genetically correlated traits. Livest. Prod. Sci. 2005, 93, 73–85. [Google Scholar] [CrossRef]
- Turner, S.P.; D’Eath, R.B.; Roehe, R.; Lawrence, A.B. Selection against aggressiveness in pigs at re-grouping: Practical application and implications for long-term behavioural patterns. Anim. Welf. 2010, 19, 123–132. [Google Scholar] [CrossRef]
- Turner, S.P.; Roehe, R.; Mekkawy, W.; Farnworth, M.J.; Knap, P.W.; Lawrence, A.B. Bayesian analysis of genetic associations of skin lesions and behavioural traits to identify genetic components of individual aggressiveness in pigs. Behav. Genet. 2008, 38, 67–75. [Google Scholar] [CrossRef]
- Knap, P.W.; Merks, J.W.M. A note on the genetics of aggressiveness of primiparous sows towards their piglets. Livest. Prod. Sci. 1987, 17, 161–167. [Google Scholar] [CrossRef]
- Larzul, C. How to Improve Meat Quality and Welfare in Entire Male Pigs by Genetics. Animals 2021, 11, 699. [Google Scholar] [CrossRef]
- Peres, L.M.; Bridi, A.M.; da Silva, C.A.; Andreo, N.; Tarsitano, M.A.; Stivaletti, E.L.T. Effect of low or high stress in pre-slaughter handling on pig carcass and meat quality. R. Bras. Zootec. 2014, 43, 363–368. [Google Scholar] [CrossRef]
- Popovski, Z.T.; Tanaskovska, B.; Miskoska-Milevska, E.; Andonov, S.; Domazetovska, S. Associations of biochemical changes and maternal traits with mutation 1843 (C>T) in the RYR1 gene as a common cause for porcine stress syndrome. Balkan J. Med. Genet. 2016, 19, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Acosta, D.B.; Español, L.Á.; Figueroa, C.E.; Marini, S.J.; Mac Allister, M.E.; Carpinetti, B.N.; Fernández, G.P.; Merino, M.L. Wild pigs (Sus scrofa) population as reservoirs for deleterious mutations in the RYR1 gene associated with Porcine Stress Syndrome. Vet. Anim. Sci. 2021, 11, 100160. [Google Scholar] [CrossRef]
- da Silva, A.N.; Ibelli, A.M.G.; Savoldi, I.R.; Cantão, M.E.; Zanella, E.L.; Marques, M.G.; da Silva, M.V.G.B.; de Peixoto, J.O.; Ledur, M.C.; Lopes, J.S.; et al. Whole-Exome Sequencing Indicated New Candidate Genes Associated with Unilateral Cryptorchidism in Pigs. Sex. Dev. 2023, 17, 56–66. [Google Scholar] [CrossRef]
- Zhao, J.; Gao, S.; Guo, Y.; Xu, Q.; Liu, M.; Zhang, C.; Cheng, M.; Zhao, X.; Schinckel, A.P.; Zhou, B. Functionally Antagonistic Transcription Factors IRF1 and IRF2 Regulate the Transcription of the Dopamine Receptor D2 Gene Associated with Aggressive Behavior of Weaned Pigs. Biology 2022, 11, 135. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, M.; Abdalla, B.A.; Zhang, Z.; Xu, Z.; Ye, Q.; Xu, H.; Luo, W.; Nie, Q.; Zhang, X. Genome-wide association study of aggressive behaviour in chicken. Sci. Rep. 2016, 6, 30981. [Google Scholar] [CrossRef] [PubMed]
- Kazantseva, A.V.; Gaysina, D.A.; Faskhutdinova, G.G.; Noskova, T.; Malykh, S.B.; Khusnutdinova, E.K. Polymorphisms of the serotonin transporter gene (5-HTTLPR, A/G SNP in 5-HTTLPR, and STin2 VNTR) and their relation to personality traits in healthy individuals from Russia. Psychiatr. Genet. 2008, 18, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Mbiydzenyuy, N.E.; Joanna Hemmings, S.M.; Shabangu, T.W.; Qulu-Appiah, L. Exploring the influence of stress on aggressive behavior and sexual function: Role of neuromodulator pathways and epigenetics. Heliyon 2024, 10, e27501. [Google Scholar] [CrossRef]
- Kazantseva, A.; Davydova, Y.; Enikeeva, R.; Mustafin, R.; Malykh, S.; Lobaskova, M.; Kanapin, A.; Prokopenko, I.; Khusnutdinova, E. A Combined Effect of Polygenic Scores and Environmental Factors on Individual Differences in Depression Level. Genes 2023, 14, 1355. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, J.; Xu, Q.; Gao, S.; Liu, M.; Zhang, C.; Schinckel, A.P.; Zhou, B. Identification of functional single nucleotide polymorphisms in the porcine SLC6A4 gene associated with aggressive behavior in weaned pigs after mixing. J. Anim. Sci. 2022, 100, skac131. [Google Scholar] [CrossRef]
- Magwai, T.; Xulu, K.R. Physiological Genomics Plays a Crucial Role in Response to Stressful Life Events, the Development of Aggressive Behaviours, and Post-Traumatic Stress Disorder (PTSD). Genes 2022, 13, 300. [Google Scholar] [CrossRef]
- Davydova, Y.D.; Enikeeva, R.F.; Kazantseva, A.V.; Mustafin, R.N.; Romanova, A.R. Genetic basis of depressive disorders. Vavilovskii Zh Genet. Sel. 2019, 23, 465–472. [Google Scholar] [CrossRef]
- Piwecka, M.; Glažar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357, eaam8526. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.; Murphy, D.L.; Crawley, J.N. Reduced aggression in mice lacking the serotonin transporter. Psychopharmacology 2002, 161, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Liang, T.; Fu, L.; Li, H.; Zhou, B. Behavioural genetic differences between Chinese and European pigs. J. Genet. 2017, 96, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Alia-Klein, N.; Goldstein, R.Z.; Kriplani, A.; Logan, J.; Tomasi, D.; Williams, B.; Telang, F.; Shumay, E.; Biegon, A.; Craig, I.W.; et al. Brain monoamine oxidase A activity predicts trait aggression. J. Neurosci. 2008, 28, 5099–5104. [Google Scholar] [CrossRef]
- Chen, R.; Chu, Q.; Shen, C.; Tong, X.; Gao, S.; Liu, X.; Zhou, B.; Schinckel, A.P. Identification of Single Nucleotide Polymorphisms in Porcine MAOA Gene Associated with Aggressive Behavior of Weaned Pigs after Group Mixing. Animals 2019, 9, 952. [Google Scholar] [CrossRef]
- Cases, O.; Seif, I.; Grimsby, J.; Gaspar, P.; Chen, K.; Pournin, S.; Müller, U.; Aguet, M.; Babinet, C.; Shih, J.C. Aggressive behavior and altered amounts of brain serotonin and norepinephrine in mice lacking MAOA. Science 1995, 268, 1763–1766. [Google Scholar] [CrossRef]
- Brunner, H.G.; Nelen, M.; Breakefield, X.O.; Ropers, H.H.; van Oost, B.A. Abnormal behavior associated with a point mutation in the structural gene for monoamine oxidase A. Science 1993, 262, 578–580. [Google Scholar] [CrossRef]
- Terenina, E.; Babigumira, B.M.; Le Mignon, G.; Bazovkina, D.; Rousseau, S.; Salin, F.; Bendixen, C.; Mormede, P. Association study of molecular polymorphisms in candidate genes related to stress responses with production and meat quality traits in pigs. Domest. Anim. Endocrinol. 2013, 44, 81–97. [Google Scholar] [CrossRef]
- Velie, B.D.; Maltecca, C.; Cassady, J.P. Genetic relationships among pig behavior, growth, backfat, and loin muscle area. J. Anim. Sci. 2009, 87, 2767–2773. [Google Scholar] [CrossRef]
- Muráni, E.; Ponsuksili, S.; D’Eath, R.B.; Turner, S.P.; Kurt, E.; Evans, G.; Thölking, L.; Klont, R.; Foury, A.; Mormède, P.; et al. Association of HPA axis-related genetic variation with stress reactivity and aggressive behaviour in pigs. BMC Genet. 2010, 11, 74. [Google Scholar] [CrossRef]
- Li, J.; Tang, L.; Wang, Y.; Li, F.; Bao, M.; Xiang, J.; Lei, D.; Tang, B. Genetic Associations and Interactions Between the NR3C1 (GR) and NR3C2 (MR) Genes and Aggressive Behavior in a Central South Chinese Han Population. Genet. Test. Mol. Biomarkers 2017, 21, 497–505. [Google Scholar] [CrossRef]
- Kazantseva, A.V.; Davydova, Y.D.; Enikeeva, R.F.; Valinurov, R.G.; Gareeva, A.E.; Khusnutdinova, N.N.; Khusnutdinova, E.K. The association study of polymorphic variants of hypothalamic-pituitary-adrenal system genes (AVPR1B, OXTR) and aggressive behavior manifestation: A focus on social environment. Res. Results Biomed. 2021, 7, 232–244. [Google Scholar] [CrossRef]
- Weaver, S.A.; Dixon, W.T.; Schaefer, A.L. The effects of mutated skeletal ryanodine receptors on hypothalamic-pituitary-adrenal axis function in boars. J. Anim. Sci. 2000, 78, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, C.; Chao, X.; Zhao, J.; Liu, M.; Chen, J.; Liu, S.; Wang, T.; Muhammad, A.; Schinckel, A.P.; et al. A Functional Single Nucleotide Polymorphism in the 3’ Untranslated Region of the Porcine JARID2 Gene Is Associated with Aggressive Behavior of Weaned Pigs after Mixing. Int. J. Mol. Sci. 2023, 25, 27. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T. A gene trap approach to identify genes that control development. Dev. Growth Differ. 1997, 39, 127–134. [Google Scholar] [CrossRef]
- Liu, X.; Shimada, T.; Otowa, T.; Wu, Y.Y.; Kawamura, Y.; Tochigi, M.; Iwata, Y.; Umekage, T.; Toyota, T.; Maekawa, M.; et al. Genome-wide Association Study of Autism Spectrum Disorder in the East Asian Populations. Autism Res. 2016, 9, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.S.; Jolly, L.; Shoubridge, C.; Chan, W.K.; Huang, L.; Laumonnier, F.; Raynaud, M.; Hackett, A.; Field, M.; Rodriguez, J.; et al. Transcriptome profiling of UPF3B/NMD-deficient lymphoblastoid cells from patients with various forms of intellectual disability. Mol. Psychiatry 2012, 17, 1103–1115. [Google Scholar] [CrossRef]
- Antoine-Bertrand, J.; Fu, M.; Lamarche-Vane, N. Direct measurement of oscillatory RhoA activity in embryonic cortical neurons stimulated with the axon guidance cue netrin-1 using fluorescence resonance energy transfer. Biol. Cell 2016, 108, 115–126. [Google Scholar] [CrossRef]
- Zhang-James, Y.; Fernàndez-Castillo, N.; Hess, J.L.; Malki, K.; Glatt, S.J.; Cormand, B.; Faraone, S.V. An integrated analysis of genes and functional pathways for aggression in human and rodent models. Mol. Psychiatry 2019, 24, 1655–1667. [Google Scholar] [CrossRef]
- Meng, Q.; Wang, K.; Liu, X.; Zhou, H.; Xu, L.; Wang, Z.; Fang, M. Identification of growth trait related genes in a Yorkshire purebred pig population by genome-wide association studies. Asian-Australas. J. Anim. Sci. 2017, 30, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhao, J.; Guo, Y.; Liu, M.; Schinckel, A.P.; Zhou, B. A Single-Nucleotide Polymorphism in the Promoter of Porcine ARHGAP24 Gene Regulates Aggressive Behavior of Weaned Pigs After Mixing by Affecting the Binding of Transcription Factor p53. Front. Cell Dev. Biol. 2022, 10, 839583. [Google Scholar] [CrossRef]
- Quilter, C.R.; Blott, S.C.; Wilson, A.E.; Bagga, M.R.; Sargent, C.A.; Oliver, G.L.; Southwood, O.I.; Gilbert, C.L.; Mileham, A.; Affara, N.A. Porcine maternal infanticide as a model for puerperal psychosis. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144B, 862–868. [Google Scholar] [CrossRef]
- Bacanu, S.A.; Bulik, C.M.; Klump, K.L.; Fichter, M.M.; Halmi, K.A.; Keel, P.; Kaplan, A.S.; Mitchell, J.E.; Rotondo, A.; Strober, M.; et al. Linkage analysis of anorexia and bulimia nervosa cohorts using selected behavioral phenotypes as quantitative traits or covariates. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 139B, 61–68. [Google Scholar] [CrossRef]
- Rincic, M.; Rados, M.; Krsnik, Z.; Gotovac, K.; Borovecki, F.; Liehr, T.; Brecevic, L. Complex intrachromosomal rearrangement in 1q leading to 1q32.2 microdeletion: A potential role of SRGAP2 in the gyrification of cerebral cortex. Mol. Cytogenet. 2016, 9, 19. [Google Scholar] [CrossRef]
- Bai, M.; Ye, D.; Guo, X.; Xi, J.; Liu, N.; Wu, Y.; Jia, W.; Wang, G.; Chen, W.; Li, G.; et al. Critical regulation of a NDIME/MEF2C axis in embryonic stem cell neural differentiation and autism. EMBO Rep. 2020, 21, e50283. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Rodriguez, J.; de-la-Rosa Fernandez, E.; Loizate Sarrionandia, I.; Benitez Garcia, E.; Herrero Moyano, M.; Morales Moreno, H.J.; Suarez Hernández, J. 5q14.3 Microdeletion Syndrome With Simultaneous Involvement of MEF2C and RASA1. Clinical Case and Review of the Literature. Pediatr. Dermatol. 2025, 42, 158–165. [Google Scholar] [CrossRef]
- Ahmadiyeh, N.; Churchill, G.A.; Shimomura, K.; Solberg, L.C.; Takahashi, J.S.; Redei, E.E. X-linked and lineage-dependent inheritance of coping responses to stress. Mamm. Genome 2003, 14, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Le Roy, I.; Mortaud, S.; Tordjman, S.; Donsez-Darcel, E.; Carlier, M.; Degrelle, H.; Roubertoux, P.L. Genetic correlation between steroid sulfatase concentration and initiation of attack behavior in mice. Behav. Genet. 1999, 29, 131–136. [Google Scholar] [CrossRef]
- Meffre, D.; Delespierre, B.; Gouézou, M.; Leclerc, P.; Vinson, G.P.; Schumacher, M.; Stein, D.G.; Guennoun, R. The membrane-associated progesterone-binding protein 25-Dx is expressed in brain regions involved in water homeostasis and is up-regulated after traumatic brain injury. J. Neurochem. 2005, 93, 1314–1326. [Google Scholar] [CrossRef]
- Pavlova, I.V.; Broshevitskaya, N.D.; Potekhina, A.A.; Shvadchenko, A.M. The Effect of Chronic Overcrowding on Social Behavior and Expression of Neuroinflammation-Associated Genes in Rats. Biochemistry 2024, 89, 1582–1594. [Google Scholar] [CrossRef]
- Yoshimoto, N.; Nakamura, Y.; Hisaoka-Nakashima, K.; Morioka, N. Mitochondrial dysfunction and type I interferon signaling induce anxiodepressive-like behaviors in mice with neuropathic pain. Exp. Neurol. 2023, 367, 114470. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, K.E.; Siegford, J.M.; Ernst, C.W.; Raney, N.E.; Bates, R.O.; Steibel, J.P. Genome-wide association analyses of lesion counts in group-housed pigs. Anim. Genet. 2018, 49, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Weißflog, L.; Becker, N.; Bossert, N.; Freudenberg, F.; Kittel-Schneider, S.; Reif, A. Expressional profile of the diacylglycerol kinase eta gene DGKH. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 445–454. [Google Scholar] [CrossRef]
- Iafrate, A.J.; Feuk, L.; Rivera, M.N.; Listewnik, M.L.; Donahoe, P.K.; Qi, Y.; Scherer, S.W.; Lee, C. Detection of large-scale variation in the human genome. Nat. Genet. 2004, 36, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, H.; Xu, Q.; Liu, M.; Chao, X.; Chen, J.; Zhou, B.; Liu, Y. Comprehensive Genome and Transcriptome Analysis Identifies SLCO3A1 Associated with Aggressive Behavior in Pigs. Biomolecules 2023, 13, 1381. [Google Scholar] [CrossRef]
- Lauterbach, E.C. Psychotropic drug effects on gene transcriptomics relevant to Parkinson’s disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 38, 107–115. [Google Scholar] [CrossRef]
- Huber, R.D.; Gao, B.; Sidler Pfändler, M.A.; Zhang-Fu, W.; Leuthold, S.; Hagenbuch, B.; Folkers, G.; Meier, P.J.; Stieger, B. Characterization of two splice variants of human organic anion transporting polypeptide 3A1 isolated from human brain. Am. J. Physiol. Cell Physiol. 2007, 292, C795–C806. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, Z.; Sun, Y.; Wang, H.; Wang, C.; Yu, S.; Liu, J.; Zhang, Y.; Fan, B.; Li, K.; et al. Analysis of genome-wide copy number variations in Chinese indigenous and western pig breeds by 60 K SNP genotyping arrays. PLoS ONE 2014, 9, e106780. [Google Scholar] [CrossRef]
- Cucchi, T.; Neaux, D.; Féral, L.; Goussard, F.; Adriensen, H.; Elleboudt, F.; Sansalone, G.; Schafberg, R. How domestication, feralization and experience-dependent plasticity affect brain size variation in Sus scrofa. R. Soc. Open Sci. 2024, 11, 240951. [Google Scholar] [CrossRef]
- Long, K.; Mao, K.; Che, T.; Zhang, J.; Qiu, W.; Wang, Y.; Tang, Q.; Ma, J.; Li, M.; Li, X. Transcriptome differences in frontal cortex between wild boar and domesticated pig. Anim. Sci. J. 2018, 89, 848–857. [Google Scholar] [CrossRef]
- Albert, F.W.; Somel, M.; Carneiro, M.; Aximu-Petri, A.; Halbwax, M.; Thalmann, O.; Blanco-Aguiar, J.A.; Plyusnina, I.Z.; Trut, L.; Villafuerte, R.; et al. A comparison of brain gene expression levels in domesticated and wild animals. PLoS Genet. 2012, 8, e1002962. [Google Scholar] [CrossRef] [PubMed]
- Honsova, E. What a pathologist needs to know about disorders of the complement system. Cesk Patol. 2024, 60, 144–149. [Google Scholar]
- Suchankova, P.; Henningsson, S.; Baghaei, F.; Rosmond, R.; Holm, G.; Ekman, A. Genetic variability within the innate immune system influences personality traits in women. Genes. Brain Behav. 2009, 8, 212–217. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Ojha, S.K.; Kartawy, M.; Hamoudi, W.; Choudhary, A.; Stern, S.; Aran, A.; Amal, H. The NO Answer for Autism Spectrum Disorder. Adv. Sci. 2023, 10, e2205783. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Yao, Y.; Yin, H.; Cai, Z.; Wang, Y.; Bai, L.; Kern, C.; Halstead, M.; Chanthavixay, G.; Trakooljul, N.; et al. Pig genome functional annotation enhances the biological interpretation of complex traits and human disease. Nat. Commun. 2021, 12, 5848. [Google Scholar] [CrossRef]
- Li, H.; Jin, L.; Wang, Y.; Hu, S.; Long, K.; Li, M. Identification and analysis of circRNAs in the prefrontal cortices of wild boar and domestic pig. Anim. Biotechnol. 2023, 34, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Poletto, R.; Cheng, H.W.; Meisel, R.L.; Richert, B.T.; Marchant, J.N. Gene expression of serotonin and dopamine receptors and monoamine oxidase-A in the brain of dominant and subordinate pubertal domestic pigs (Sus scrofa) fed a β-adrenoreceptor agonist. Brain Res. 2011, 1381, 11–20. [Google Scholar] [CrossRef]
- Murani, E.; Ponsuksili, S.; D’Eath, R.B.; Turner, S.P.; Evans, G.; Tholking, L.; Kurt, E.; Klont, R.; Foury, A.; Mormede, P.; et al. Differential mRNA expression of genes in the porcine adrenal gland associated with psychosocial stress. J. Mol. Endocrinol. 2011, 46, 165–174. [Google Scholar] [CrossRef]
- MacArthur Clark, J. The 3Rs in research: A contemporary approach to replacement, reduction and refinement. Br. J. Nutr. 2018, 120, S1–S7. [Google Scholar] [CrossRef]
- Mustafina, R.N.; Kazantseva, A.V.; Enikeeva, R.F.; Davydova, Y.D.; Karunas, A.S.; Malykhd, S.B.; Khusnutdinovaa, E.K. Epigenetics of Aggressive Behavior. Russ. J. Genet. 2019, 55, 987–997. [Google Scholar] [CrossRef]
- Tatemoto, P.; Pértille, F.; Bernardino, T.; Zanella, R.; Guerrero-Bosagna, C.; Zanella, A.J. An enriched maternal environment and stereotypies of sows differentially affect the neuro-epigenome of brain regions related to emotionality in their piglets. Epigenetics 2023, 18, 2196656. [Google Scholar] [CrossRef]
- Tatemoto, P.; Bernardino, T.; Morrone, B.; Queiroz, M.R.; Zanella, A.J. Stereotypic Behavior in Sows Is Related to Emotionality Changes in the Offspring. Front. Vet. Sci. 2020, 7, 79. [Google Scholar] [CrossRef]
- Palma-Gudiel, H.; Fañanás, L. An integrative review of methylation at the serotonin transporter gene and its dialogue with environmental risk factors, psychopathology and 5-HTTLPR. Neurosci. Biobehav. Rev. 2017, 72, 190–209. [Google Scholar] [CrossRef] [PubMed]
- Keever-Keigher, M.R.; Zhang, P.; Bolt, C.R.; Rymut, H.E.; Antonson, A.M.; Caputo, M.P.; Houser, A.K.; Hernandez, A.G.; Southey, B.R.; Rund, L.A.; et al. Interacting impact of maternal inflammatory response and stress on the amygdala transcriptome of pigs. G3 2021, 11, jkab113. [Google Scholar] [CrossRef] [PubMed]
- Khaledi, F.; Dehkordi, H.T.; Zarean, E.; Shahrani, M.; Amini-Khoei, H. Possible role of NO/NMDA pathway in the autistic-like behaviors induced by maternal separation stress in mice. PLoS ONE 2023, 18, e0292631. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, L.; Ma, W.; Zheng, X.; He, E.; Zhang, B.; Vashisth, M.K.; Gong, Z. Maternal separation affects anxiety-like behavior beginning in adolescence and continuing through adulthood and related to Dnmt3a expression. J. Neurophysiol. 2022, 128, 611–618. [Google Scholar] [CrossRef]
- Poletto, R.; Steibel, J.P.; Siegford, J.M.; Zanella, A.J. Effects of early weaning and social isolation on the expression of glucocorticoid and mineralocorticoid receptor and 11beta-hydroxysteroid dehydrogenase 1 and 2 mRNAs in the frontal cortex and hippocampus of piglets. Brain Res. 2006, 1067, 36–42. [Google Scholar] [CrossRef]
- Kanitz, E.; Manteuffel, G.; Otten, W. Effects of weaning and restraint stress on glucocorticoid receptor binding capacity in limbic areas of domestic pigs. Brain Res. 1998, 804, 311–315. [Google Scholar] [CrossRef]
- Mizoguchi, K.; Ishige, A.; Aburada, M.; Tabira, T. Chronic stress attenuates glucocorticoid negative feedback: Involvement of the prefrontal cortex and hippocampus. Neuroscience 2003, 119, 887–897. [Google Scholar] [CrossRef]
- Corbett, R.J.; Luttman, A.M.; Wurtz, K.E.; Siegford, J.M.; Raney, N.E.; Ford, L.M.; Ernst, C.W. Weaning Induces Stress-Dependent DNA Methylation and Transcriptional Changes in Piglet PBMCs. Front. Genet. 2021, 12, 633564. [Google Scholar] [CrossRef]
- Lee, A.L.; Ogle, W.O.; Sapolsky, R.M. Stress and depression: Possible links to neuron death in the hippocampus. Bipolar Disord. 2002, 4, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Kanitz, E.; Tuchscherer, M.; Puppe, B.; Tuchscherer, A.; Stabenow, B. Consequences of repeated early isolation in domestic piglets (Sus scrofa) on their behavioural, neuroendocrine, and immunological responses. Brain Behav. Immun. 2004, 18, 35–45. [Google Scholar] [CrossRef]
- Lapiz, M.D.; Fulford, A.; Muchimapura, S.; Mason, R.; Parker, T.; Marsden, C.A. Influence of postweaning social isolation in the rat on brain development, conditioned behavior, and neurotransmission. Neurosci. Behav. Physiol. 2003, 33, 13–29. [Google Scholar] [CrossRef]
- Keever, M.R.; Zhang, P.; Bolt, C.R.; Antonson, A.M.; Rymut, H.E.; Caputo, M.P.; Houser, A.K.; Hernandez, A.G.; Southey, B.R.; Rund, L.A.; et al. Lasting and Sex-Dependent Impact of Maternal Immune Activation on Molecular Pathways of the Amygdala. Front. Neurosci. 2020, 14, 774. [Google Scholar] [CrossRef] [PubMed]
- Pie, S.; Lalles, J.P.; Blazy, F.; Laffitte, J.; Seve, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef]
- Agha, S.; Foister, S.; Roehe, R.; Turner, S.P.; Doeschl-Wilson, A. Genetic Analysis of Novel Behaviour Traits in Pigs Derived from Social Network Analysis. Genes 2022, 13, 561. [Google Scholar] [CrossRef]
- Peden, R.S.E.; Akaichi, F.; Camerlink, I.; Boyle, L.A.; Turner, S.P. Pig farmers’ willingness to pay for management strategies to reduce aggression between pigs. PLoS ONE 2019, 14, e0224924. [Google Scholar] [CrossRef] [PubMed]
- Camerlink, I.; Ursinus, W.W.; Bijma, P.; Kemp, B.; Bolhuis, J.E. Indirect genetic effects for growth rate in domestic pigs alter aggressive and manipulative biting behaviour. Behav. Genet. 2015, 45, 117–126. [Google Scholar] [CrossRef]
- Martelli, G.; Sardi, L.; Stancampiano, L.; Govoni, N.; Zannoni, A.; Nannoni, E.; Forni, M.; Bacci, M.L. A study on some welfare-related parameters of hDAF transgenic pigs when compared with their conventional close relatives. Animal 2014, 8, 810–816. [Google Scholar] [CrossRef]

{kind=link}
| Severity of the Damage Caused | Variant of Aggression | Description |
|---|---|---|
| Medium | Head-to-head impact | The act of striking the cranium, muzzle, or neck of another animal without employing the mouth is referred to as “striking the head/muzzle”. |
| Head-to-body impact | Impact of a blow to the area posterior to the ears of another animal. | |
| Parallel pressing | Animals exhibit a lateral-to-lateral comparison of their bodies, characterized by the application of pressure through their shoulder girdle regions, accompanied by their craniums being positioned posterior to one another’s necks or crania. | |
| Reverse parallel pressing | The animals adopt a position opposite one another followed by a force with their shoulder girdle, causing their heads to move behind each other’s neck or head. | |
| Heavy | Neck bite | The neck bite was characterized by its aggressive nature, without a concomitant head thrust. |
| Body bite | Aggressive bite to the body. | |
| Ear bite | Aggressive biting of another pig’s neck. | |
| Tail bite/tail bite off | The range of interactions with the tail encompasses a variety of severity levels, from minor nicks to severe damage to the sacrum [21]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazantseva, A.; Bilyalov, A.; Filatov, N.; Perepechenov, S.; Gusev, O. Genetic Contributions to Aggressive Behaviour in Pigs: A Comprehensive Review. Genes 2025, 16, 534. https://doi.org/10.3390/genes16050534
Kazantseva A, Bilyalov A, Filatov N, Perepechenov S, Gusev O. Genetic Contributions to Aggressive Behaviour in Pigs: A Comprehensive Review. Genes. 2025; 16(5):534. https://doi.org/10.3390/genes16050534
Chicago/Turabian StyleKazantseva, Anastasiya, Airat Bilyalov, Nikita Filatov, Stepan Perepechenov, and Oleg Gusev. 2025. "Genetic Contributions to Aggressive Behaviour in Pigs: A Comprehensive Review" Genes 16, no. 5: 534. https://doi.org/10.3390/genes16050534
APA StyleKazantseva, A., Bilyalov, A., Filatov, N., Perepechenov, S., & Gusev, O. (2025). Genetic Contributions to Aggressive Behaviour in Pigs: A Comprehensive Review. Genes, 16(5), 534. https://doi.org/10.3390/genes16050534