
Bioinformatic Analysis of Autism-Related miRNAs and Their PoTential as Biomarkers for Autism Epigenetic Inheritance

Abstract
1. Introduction
2. Materials and Methods
2.1. Compilation of Autism-Related miRNAs
2.2. Identification of miRNAs Expressed in Peripheral Blood Mononuclear Cells (PBMCs), Sperm, and Neurons
2.3. Analysis of Genes Targeted by the Autism-Related miRNAs
2.4. Pathway Enrichment Analysis
2.4.1. Metascape
2.4.2. gProfiler
2.5. miRNAs Enrichment Analysis
3. Results
3.1. Autism-Related miRNAs: miRNA-to-Gene Analysis
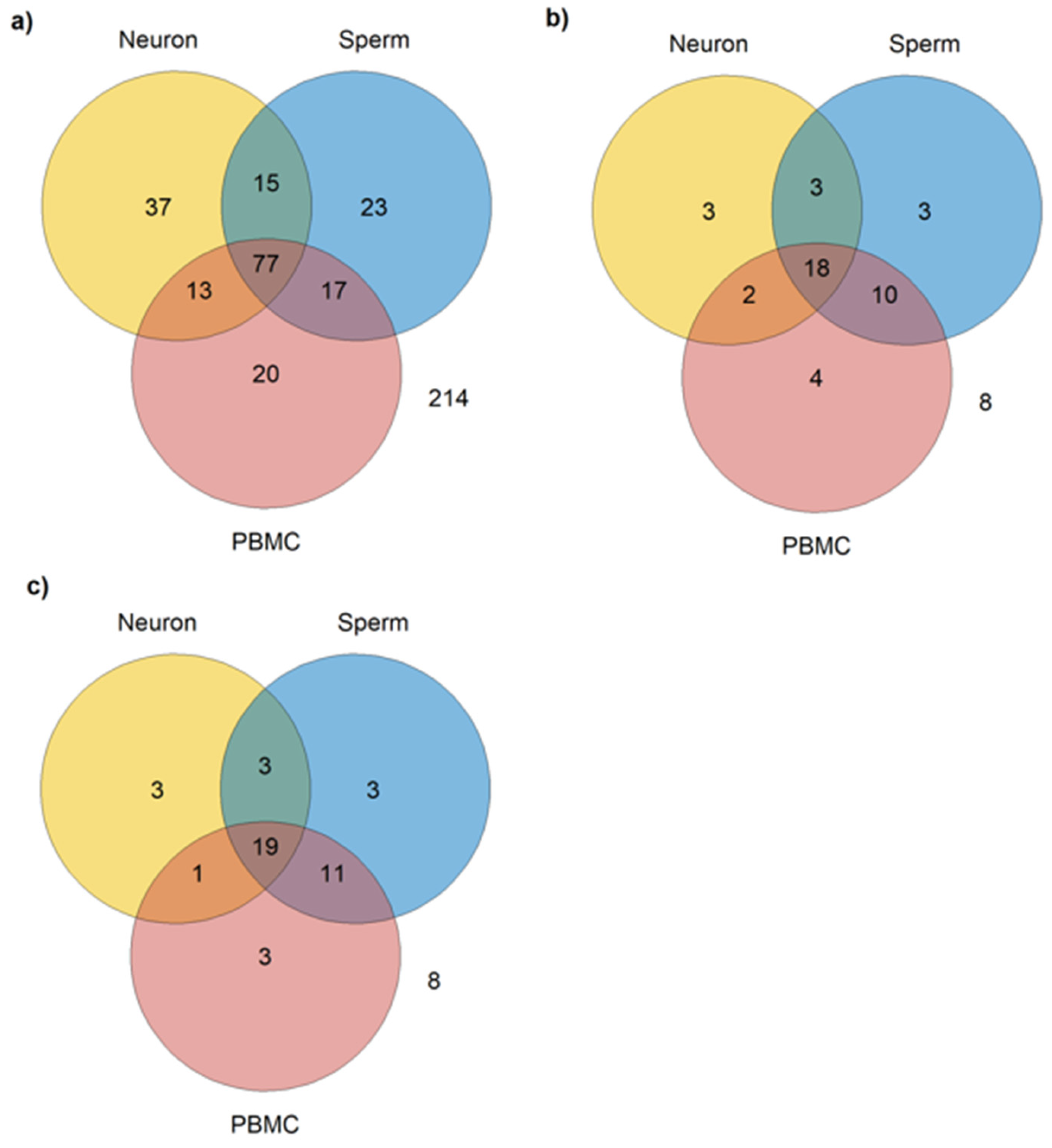
3.2. miRNAs Expressed in PBMCs, Neurons, and Sperm
3.3. Identification of Genes Targeted by Candidate Autism-Related miRNA
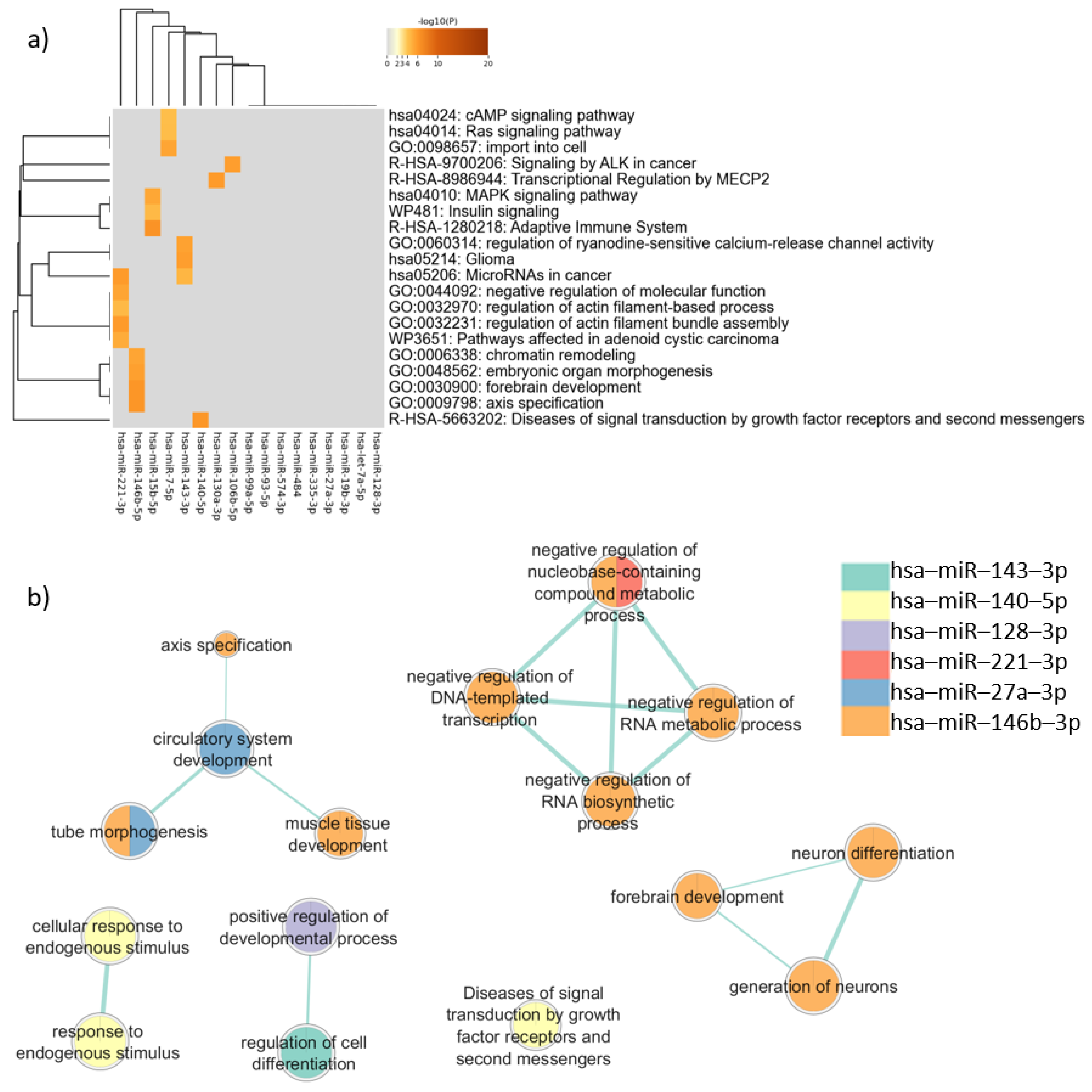
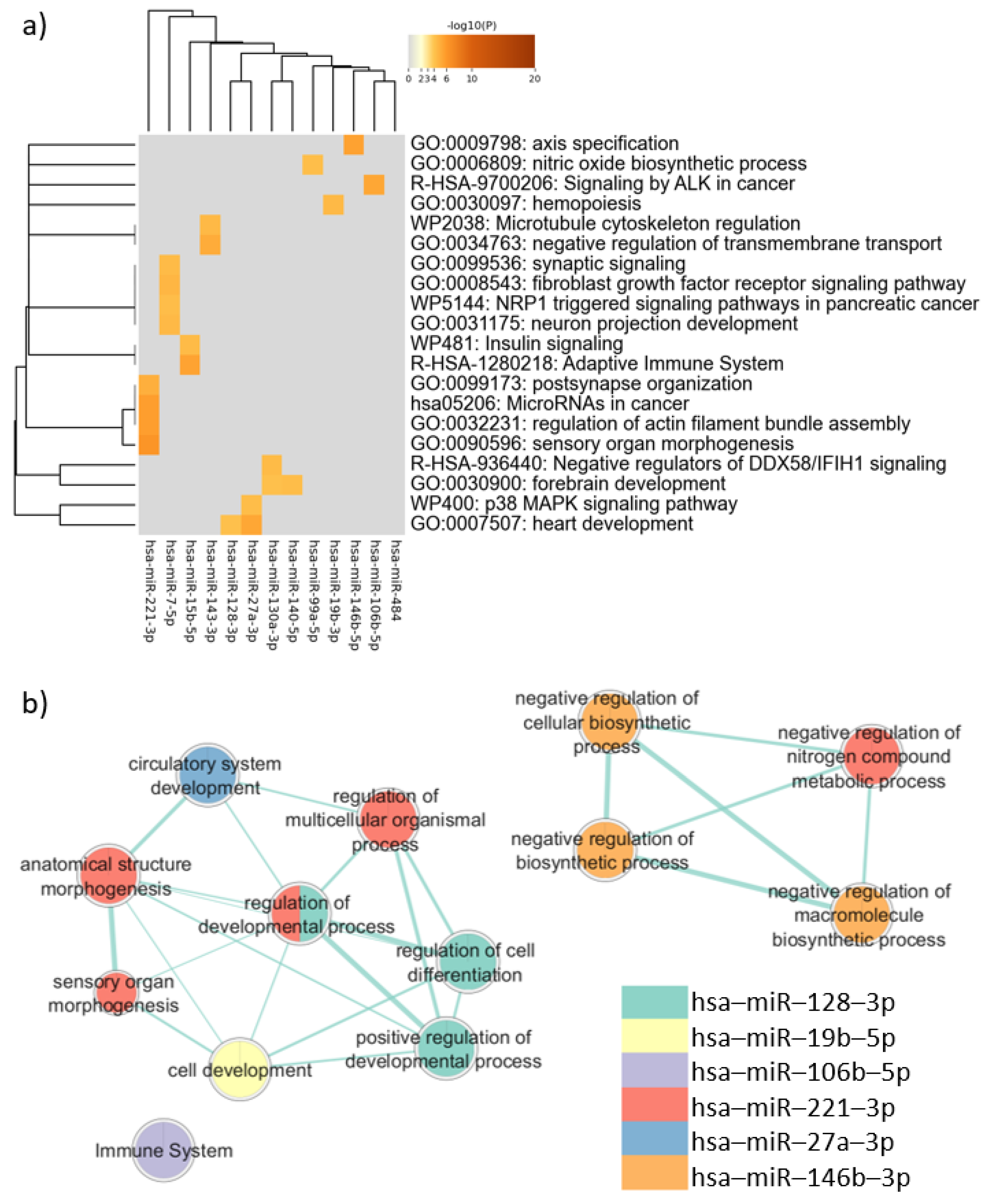
3.4. Enrichment Analysis of All the Genes Targeted by the 18 Candidate Autism-Related miRNAs
3.5. Enrichment Analysis of Target Genes Common to Neurons and Sperm
3.6. Enrichment Analysis of Target Genes Common to PBMCs, Neurons, and Sperm
3.7. Autism-Related miRNAs: Gene-to-miRNA Analysis (miRNA Enrichment)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADHD | Attention deficit hyperactivity disorder |
| ASD | Autism spectrum disorder |
| E | Enrichment value |
| eQTL | Expression quantitative trait loci |
| FDR | False discovery rate |
| GO | Gene Ontology |
| GWAS | Genome wide association studies |
| miRNA | microRNA |
| Padj | Ajusted p-value |
| PBMC | Peripheral blood mononuclear cells |
| PCR | Polymerase chain reaction |
| RPM | Reads per million |
| SFARI | Simons Foundation Autism Research Initiative |
| sncRNA | small non-coding RNA |
| SNP | Single nucleotide polymorphisms |
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Aldinger, K.A.; Lane, C.J.; Veenstra-VanderWeele, J.; Levitt, P. Patterns of Risk for Multiple Co-Occurring Medical Conditions Replicate Across Distinct Cohorts of Children with Autism Spectrum Disorder. Autism Res. 2015, 8, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Bougeard, C.; Picarel-Blanchot, F.; Schmid, R.; Campbell, R.; Buitelaar, J. Prevalence of Autism Spectrum Disorder and Co-morbidities in Children and Adolescents: A Systematic Literature Review. Front. Psychiatry 2021, 12, 744709. [Google Scholar] [CrossRef] [PubMed]
- Leader, G.; Abberton, C.; Cunningham, S.; Gilmartin, K.; Grudzien, M.; Higgins, E.; Joshi, L.; Whelen, S.; Mannion, A. Gastrointestinal Symptoms in Autism Spectrum Disorder: A Systematic Review. Nutrients 2022, 14, 1471. [Google Scholar] [CrossRef]
- McElhanon, B.O.; McCracken, C.; Karpen, S.; Sharp, W.G. Gastrointestinal Symptoms in Autism Spectrum Disorder: A Meta-analysis. Pediatrics 2014, 133, 872–883. [Google Scholar] [CrossRef]
- Viscidi, E.W.; Triche, E.W.; Pescosolido, M.F.; McLean, R.L.; Joseph, R.M.; Spence, S.J.; Morrow, E.M. Clinical Characteristics of Children with Autism Spectrum Disorder and Co-Occurring Epilepsy. PLoS ONE 2013, 8, e67797. [Google Scholar] [CrossRef]
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental risk factors for autism: An evidence-based review of systematic reviews and meta-analyses. Mol. Autism 2017, 8, 13. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Rajman, M.; Schratt, G. MicroRNAs in neural development: From master regulators to fine-tuners. Development 2017, 144, 2310–2322. [Google Scholar] [CrossRef]
- Garrido-Torres, N.; Guzmán-Torres, K.; García-Cerro, S.; Bermúdez, G.P.; Cruz-Baquero, C.; Ochoa, H.; García-González, D.; Canal-Rivero, M.; Crespo-Facorro, B.; Ruiz-Veguilla, M. miRNAs as biomarkers of autism spectrum disorder: A systematic review and meta-analysis. Eur. Child Adolesc. Psychiatry 2023, 33, 2957–2990. [Google Scholar] [CrossRef]
- Stott, J.; Wright, T.; Holmes, J.; Wilson, J.; Griffiths-Jones, S.; Foster, D.; Wright, B. A systematic review of non-coding RNA genes with differential expression profiles associated with autism spectrum disorders. PLoS ONE 2023, 18, e0287131. [Google Scholar] [CrossRef]
- Ozkul, Y.; Taheri, S.; Bayram, K.K.; Sener, E.F.; Mehmetbeyoglu, E.; Öztop, D.B.; Aybuga, F.; Tufan, E.; Bayram, A.; Dolu, N.; et al. A heritable profile of six miRNAs in autistic patients and mouse models. Sci. Rep. 2020, 10, 9011. [Google Scholar] [CrossRef] [PubMed]
- DeRosa, B.A.; Van Baaren, J.M.; Dubey, G.K.; Lee, J.M.; Cuccaro, M.L.; Vance, J.M.; Pericak-Vance, M.A.; Dykxhoorn, D.M. Derivation of autism spectrum disorder-specific induced pluripotent stem cells from peripheral blood mononuclear cells. Neurosci. Lett. 2012, 516, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Ben-Reuven, L.; Reiner, O. Modeling the autistic cell: iPSCs recapitulate developmental principles of syndromic and nonsyndromic ASD. Dev. Growth Differ. 2016, 58, 481–491. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Gao, Y.; Stanley, E.; Fanjul-Fernández, M.; Brown, N.J.; Pope, K.; Green, C.C.; Vlahos, K.; Sourris, K.; Bahlo, M.; et al. Generation of seven iPSC lines from peripheral blood mononuclear cells suitable to investigate Autism Spectrum Disorder. Stem Cell Res. 2019, 39, 101516. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, Y.; Yang, X.; Zhang, H.; Wang, B.; Guan, J.; Gao, J.; Ma, X.; Liu, Y. Generation and characterization of PBMCs-derived human induced pluripotent stem cell (iPSC) line SDQLCHi051-A from an autism spectrum disorder patient with compound CHD8 gene mutations. Stem Cell Res. 2023, 69, 103114. [Google Scholar] [CrossRef]
- Frye, R.E.; Lionnard, L.; Singh, I.; Karim, M.A.; Chajra, H.; Frechet, M.; Kissa, K.; Racine, V.; Ammanamanchi, A.; McCarty, P.J.; et al. Mitochondrial morphology is associated with respiratory chain uncoupling in autism spectrum disorder. Transl. Psychiatry 2021, 11, 527. [Google Scholar] [CrossRef]
- Liu, H.; Zimmerman, A.W.; Singh, K.; Connors, S.L.; Diggins, E.; Stephenson, K.K.; Dinkova-Kostova, A.T.; Fahey, J.W. Biomarker Exploration in Human Peripheral Blood Mononuclear Cells for Monitoring Sulforaphane Treatment Responses in Autism Spectrum Disorder. Sci. Rep. 2020, 10, 5822. [Google Scholar] [CrossRef]
- Shen, L.; Feng, C.; Zhang, K.; Chen, Y.; Gao, Y.; Ke, J.; Chen, X.; Lin, J.; Li, C.; Iqbal, J.; et al. Proteomics Study of Peripheral Blood Mononuclear Cells (PBMCs) in Autistic Children. Front. Cell Neurosci. 2019, 13, 105. [Google Scholar] [CrossRef]
- Nestor, M.W.; Phillips, A.W.; Artimovich, E.; Nestor, J.E.; Hussman, J.P.; Blatt, G.J. Human Inducible Pluripotent Stem Cells and Autism Spectrum Disorder: Emerging Technologies. Autism Res. 2016, 9, 513–535. [Google Scholar] [CrossRef]
- Pecorelli, A.; Cervellati, F.; Belmonte, G.; Montagner, G.; Waldon, P.; Hayek, J.; Gambari, R.; Valacchi, G. Cytokines profile and peripheral blood mononuclear cells morphology in Rett and autistic patients. Cytokine 2016, 77, 180–188. [Google Scholar] [CrossRef]
- Naik, U.S.; Gangadharan, C.; Abbagani, K.; Nagalla, B.; Dasari, N.; Manna, S.K. A Study of Nuclear Transcription Factor-Kappa B in Childhood Autism. PLoS ONE 2011, 6, e19488. [Google Scholar] [CrossRef] [PubMed]
- Talebizadeh, Z.; Butler, M.G.; Theodoro, M.F. Feasibility and relevance of examining lymphoblastoid cell lines to study role of microRNAs in autism. Autism Res. 2008, 1, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Sarachana, T.; Zhou, R.; Chen, G.; Manji, H.K.; Hu, V.W. Investigation of post-transcriptional gene regulatory networks associated with autism spectrum disorders by microRNA expression profiling of lymphoblastoid cell lines. Genome Med. 2010, 2, 23. [Google Scholar] [CrossRef]
- Huang, F.; Long, Z.; Chen, Z.; Li, J.; Hu, Z.; Qiu, R.; Zhuang, W.; Tang, B.; Xia, K.; Jiang, H. Investigation of Gene Regulatory Networks Associated with Autism Spectrum Disorder Based on MiRNA Expression in China. PLoS ONE 2015, 10, e0129052. [Google Scholar] [CrossRef]
- Nguyen, L.S.; Lepleux, M.; Makhlouf, M.; Martin, C.; Fregeac, J.; Siquier-Pernet, K.; Philippe, A.; Feron, F.; Gepner, B.; Rougeulle, C.; et al. Profiling olfactory stem cells from living patients identifies miRNAs relevant for autism pathophysiology. Mol. Autism 2016, 7, 1. [Google Scholar] [CrossRef]
- Jyonouchi, H.; Geng, L.; Streck, D.L.; Dermody, J.J.; Toruner, G.A. MicroRNA expression changes in association with changes in interleukin-1ß/interleukin10 ratios produced by monocytes in autism spectrum disorders: Their association with neuropsychiatric symptoms and comorbid conditions (observational study). J. Neuroinflammation 2017, 14, 229. [Google Scholar] [CrossRef]
- Nakata, M.; Kimura, R.; Funabiki, Y.; Awaya, T.; Murai, T.; Hagiwara, M. MicroRNA profiling in adults with high-functioning autism spectrum disorder. Mol. Brain 2019, 12, 82. [Google Scholar] [CrossRef]
- Atwan, H.; Assarehzadegan, M.-A.; Shekarabi, M.; Jazayeri, S.M.; Barfi, S.; Shoormasti, R.S.; Chimeh, N.; Pouretemad, H.R.; Tayebi, B. Assessment of miR-181b-5p, miR-23a-3p, BCL-2, and IL-6 in Peripheral Blood Mononuclear Cells of Autistic Patients; Likelihood of Reliable Biomarkers. Iran. J. Allergy Asthma Immunol. 2020, 19, 74–83. [Google Scholar] [CrossRef]
- Grove, J.; Ripke, S.; Als, T.D.; Mattheisen, M.; Walters, R.K.; Won, H.; Pallesen, J.; Agerbo, E.; Andreassen, O.A.; Anney, R.; et al. Identification of common genetic risk variants for autism spectrum disorder. Nat. Genet. 2019, 51, 431–444. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Z.-P.; Hu, H.; Lei, J.; Zhou, Z.; Yao, B.; Chen, L.; Liang, G.; Zhan, S.; Zhu, X.; et al. Sperm microRNAs confer depression susceptibility to offspring. Sci. Adv. 2021, 7, eabd7605. [Google Scholar] [CrossRef]
- Tyebji, S.; Hannan, A.J.; Tonkin, C.J. Pathogenic Infection in Male Mice Changes Sperm Small RNA Profiles and Transgenerationally Alters Offspring Behavior. Cell Rep. 2020, 31, 107573. [Google Scholar] [CrossRef] [PubMed]
- Yeshurun, S.; Hannan, A.J. Transgenerational epigenetic influences of paternal environmental exposures on brain function and predisposition to psychiatric disorders. Mol. Psychiatry 2019, 24, 536–548. [Google Scholar] [CrossRef]
- Gapp, K.; Jawaid, A.; Sarkies, P.; Bohacek, J.; Pelczar, P.; Prados, J.; Farinelli, L.; Miska, E.; Mansuy, I.M. Implication of sperm RNAs in transgenerational inheritance of the effects of early trauma in mice. Nat. Neurosci. 2014, 17, 667–669. [Google Scholar] [CrossRef] [PubMed]
- Marczylo, E.L.; Amoako, A.A.; Konje, J.C.; Gant, T.W.; Marczylo, T.H. Smoking induces differential miRNA expression in human spermatozoa: A potential transgenerational epigenetic concern? Epigenetics 2012, 7, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Dickson, D.A.; Paulus, J.K.; Mensah, V.; Lem, J.; Saavedra-Rodriguez, L.; Gentry, A.; Pagidas, K.; Feig, L.A. Reduced levels of miRNAs 449 and 34 in sperm of mice and men exposed to early life stress. Transl. Psychiatry 2018, 8, 101. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. miRNet 2.0: Network-based visual analytics for miRNA functional analysis and systems biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef]
- Chen, J.; Lin, J.; Hu, Y.; Ye, M.; Yao, L.; Wu, L.; Zhang, W.; Wang, M.; Deng, T.; Guo, F.; et al. RNADisease v4.0: An updated resource of RNA-associated diseases, providing RNA-disease analysis, enrichment and prediction. Nucleic Acids Res. 2023, 51, D1397–D1404. [Google Scholar] [CrossRef]
- Kavakiotis, I.; Alexiou, A.; Tastsoglou, S.; Vlachos, I.S.; Hatzigeorgiou, A.G. DIANA-miTED: A microRNA tissue expression database. Nucleic Acids Res. 2022, 50, D1055–D1061. [Google Scholar] [CrossRef]
- Panwar, B.; Omenn, G.S.; Guan, Y. miRmine: A database of human miRNA expression profiles. Bioinformatics 2017, 33, 1554–1560. [Google Scholar] [CrossRef]
- Schuster, A.; Tang, C.; Xie, Y.; Ortogero, N.; Yuan, S.; Yan, W. SpermBase: A Database for Sperm-Borne RNA Contents. Biol. Reprod. 2016, 95, 99. [Google Scholar] [CrossRef]
- McCall, M.N.; Kim, M.-S.; Adil, M.; Patil, A.H.; Lu, Y.; Mitchell, C.J.; Leal-Rojas, P.; Xu, J.; Kumar, M.; Dawson, V.L.; et al. Toward the human cellular microRNAome. Genome Res. 2017, 27, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed]
- Tokar, T.; Pastrello, C.; Rossos, A.E.M.; Abovsky, M.; Hauschild, A.-C.; Tsay, M.; Lu, R.; Jurisica, I. mirDIP 4.1—Integrative database of human microRNA target predictions. Nucleic Acids Res. 2018, 46, D360–D370. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Lin, Y.-C.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. miRTarBase update 2022: An informative resource for experimentally validated miRNA–target interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Kolberg, L.; Raudvere, U.; Kuzmin, I.; Adler, P.; Vilo, J.; Peterson, H. g:Profiler—Interoperable web service for functional enrichment analysis and gene identifier mapping (2023 update). Nucleic Acids Res. 2023, 51, W207–W212. [Google Scholar] [CrossRef]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment Map: A Network-Based Method for Gene-Set Enrichment Visualization and Interpretation. PLoS ONE 2010, 5, e13984. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Abrahams, B.S.; Arking, D.E.; Campbell, D.B.; Mefford, H.C.; Morrow, E.M.; Weiss, L.A.; Menashe, I.; Wadkins, T.; Banerjee-Basu, S.; Packer, A. SFARI Gene 2.0: A community-driven knowledgebase for the autism spectrum disorders (ASDs). Mol. Autism 2013, 4, 36. [Google Scholar] [CrossRef]
- Sollis, E.; Mosaku, A.; Abid, A.; Buniello, A.; Cerezo, M.; Gil, L.; Groza, T.; Güneş, O.; Hall, P.; Hayhurst, J.; et al. The NHGRI-EBI GWAS Catalog: Knowledgebase and deposition resource. Nucleic Acids Res. 2023, 51, D977–D985. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Germain, P.; Soutschek, M. enrichMiR: miRNA Target Enrichment Analysis. R Package Version 0.99.32. 2023. Available online: https://github.com/ETHZ-INS/enrichMiR (accessed on 10 April 2024).
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Su, N.; Liu, L.; Zhang, J.; Wang, H.; Zhang, W.; Gui, J.; Yu, K.; Li, J.; Le, T.D. miRBaseConverter: An R/Bioconductor package for converting and retrieving miRNA name, accession, sequence and family information in different versions of miRBase. BMC Bioinform. 2018, 19, 514. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R. Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 9 February 2023).
- Wickham, H.; Bryan, J. readxl: Read Excel Files. R Package Version 1.4.3. 2023. Available online: https://CRAN.R-project.org/package=readxl (accessed on 9 February 2023).
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. dplyr: A Grammar of Data Manipulation. R Package Version 1.1.4. 2023. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 9 February 2023).
- Wickham, H.; Vaughan, D.; Girlich, M. tidyr: Tidy Messy Data. R Package Version 1.3.0. 2023. Available online: https://CRAN.R-project.org/package=tidyr (accessed on 9 February 2023).
- Signorell, A. DescTools: Tools for Descriptive Statistics. R Package Version 0.99.52. 2023. Available online: https://CRAN.R-project.org/package=DescTools (accessed on 9 February 2023).
- Yan, L. ggvenn: Draw Venn Diagram by ‘ggplot2’. R Package Version 0.1.10. 2023. Available online: https://CRAN.R-project.org/package=ggvenn (accessed on 9 February 2023).
- Kolberg, L.; Raudvere, U.; Kuzmin, I.; Vilo, J.; Peterson, H. gprofiler2—An R package for gene list functional enrichment analysis and namespace conversion toolset g:Profiler. F1000Res 2020, 9, 709. [Google Scholar] [CrossRef]
- Timmons, J.A.; Szkop, K.J.; Gallagher, I.J. Multiple sources of bias confound functional enrichment analysis of global -omics data. Genome Biol. 2015, 16, 186. [Google Scholar] [CrossRef]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Primers 2021, 1, 59. [Google Scholar] [CrossRef]
- Martin, F.J.; Amode, M.R.; Aneja, A.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Becker, A.; Bennett, R.; Berry, A.; Bhai, J.; et al. Ensembl 2023. Nucleic Acids Res. 2023, 51, D933–D941. [Google Scholar] [CrossRef]
- Mor, M.; Nardone, S.; Sams, D.S.; Elliott, E. Hypomethylation of miR-142 promoter and upregulation of microRNAs that target the oxytocin receptor gene in the autism prefrontal cortex. Mol. Autism 2015, 6, 46. [Google Scholar] [CrossRef]
- Hicks, S.D.; Carpenter, R.L.; Wagner, K.E.; Pauley, R.; Barros, M.; Tierney-Aves, C.; Barns, S.; Greene, C.D.; Middleton, F.A. Saliva MicroRNA Differentiates Children with Autism from Peers with Typical and Atypical Development. J. Am. Acad. Child Adolesc. Psychiatry 2020, 59, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elneel, K.; Liu, T.; Gazzaniga, F.S.; Nishimura, Y.; Wall, D.P.; Geschwind, D.H.; Lao, K.; Kosik, K.S. Heterogeneous dysregulation of microRNAs across the autism spectrum. Neurogenetics 2008, 9, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Jyonouchi, H.; Geng, L.; Toruner, G.A.; Rose, S.; Bennuri, S.C.; Frye, R.E. Serum microRNAs in ASD: Association with Monocyte Cytokine Profiles and Mitochondrial Respiration. Front. Psychiatry 2019, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- Vasu, M.M.; Anitha, A.; Thanseem, I.; Suzuki, K.; Yamada, K.; Takahashi, T.; Wakuda, T.; Iwata, K.; Tsujii, M.; Sugiyama, T.; et al. Serum microRNA profiles in children with autism. Mol. Autism 2014, 5, 40. [Google Scholar] [CrossRef]
- Hicks, S.D.; Ignacio, C.; Gentile, K.; Middleton, F.A. Salivary miRNA profiles identify children with autism spectrum disorder, correlate with adaptive behavior, and implicate ASD candidate genes involved in neurodevelopment. BMC Pediatr. 2016, 16, 52. [Google Scholar] [CrossRef]
- Vaccaro, T.D.S.; Sorrentino, J.M.; Salvador, S.; Veit, T.; Souza, D.O.; De Almeida, R.F. Alterations in the MicroRNA of the Blood of Autism Spectrum Disorder Patients: Effects on Epigenetic Regulation and Potential Biomarkers. Behav. Sci. 2018, 8, 75. [Google Scholar] [CrossRef]
- Hicks, S.D.; Rajan, A.T.; Wagner, K.E.; Barns, S.; Carpenter, R.L.; Middleton, F.A. Validation of a Salivary RNA Test for Childhood Autism Spectrum Disorder. Front. Genet. 2018, 9, 534. [Google Scholar] [CrossRef]
- Sun, J.-J.; Chen, B.; Yu, T. Construction of an immune-related ceRNA network to screen for potential diagnostic markers for autism spectrum disorder. Front. Genet. 2022, 13, 1025813. [Google Scholar] [CrossRef]
- Kalemaj, Z.; Marino, M.M.; Santini, A.C.; Tomaselli, G.; Auti, A.; Cagetti, M.G.; Borsello, T.; Costantino, A.; Inchingolo, F.; Boccellino, M.; et al. Salivary microRNA profiling dysregulation in autism spectrum disorder: A pilot study. Front. Neurosci. 2022, 16, 945278. [Google Scholar] [CrossRef]
- Wu, Y.E.; Parikshak, N.N.; Belgard, T.G.; Geschwind, D.H. Genome-wide, integrative analysis implicates microRNA dysregulation in autism spectrum disorder. Nat. Neurosci. 2016, 19, 1463–1476. [Google Scholar] [CrossRef]
- Sehovic, E.; Spahic, L.; Smajlovic-Skenderagic, L.; Pistoljevic, N.; Dzanko, E.; Hajdarpasic, A. Identification of developmental disorders including autism spectrum disorder using salivary miRNAs in children from Bosnia and Herzegovina. PLoS ONE 2020, 15, e0232351. [Google Scholar] [CrossRef] [PubMed]
- Ramaswami, G.; Won, H.; Gandal, M.J.; Haney, J.; Wang, J.C.; Wong, C.C.Y.; Sun, W.; Prabhakar, S.; Mill, J.; Geschwind, D.H. Integrative genomics identifies a convergent molecular subtype that links epigenomic with transcriptomic differences in autism. Nat. Commun. 2020, 11, 4873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Pang, Y.; Feng, W.; Jin, Y.; Chen, S.; Ding, S.; Wang, Z.; Zou, Y.; Li, Y.; Wang, T.; et al. miR-124 regulates early isolation-induced social abnormalities via inhibiting myelinogenesis in the medial prefrontal cortex. Cell Mol. Life Sci. 2022, 79, 507. [Google Scholar] [CrossRef]
- Salloum-Asfar, S.; Elsayed, A.K.; Elhag, S.F.; Abdulla, S.A. Circulating Non-Coding RNAs as a Signature of Autism Spectrum Disorder Symptomatology. Int. J. Mol. Sci. 2021, 22, 6549. [Google Scholar] [CrossRef]
- Hosokawa, R.; Yoshino, Y.; Funahashi, Y.; Horiuchi, F.; Iga, J.-I.; Ueno, S.-I. MiR-15b-5p Expression in the Peripheral Blood: A Potential Diagnostic Biomarker of Autism Spectrum Disorder. Brain Sci. 2022, 13, 27. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, T.; Li, X.; Jiang, M.; Jia, M.; Liu, J.; Zhang, D.; Li, J.; Wang, L. Altered Expression of Brain-specific Autism-Associated miRNAs in the Han Chinese Population. Front. Genet. 2022, 13, 865881. [Google Scholar] [CrossRef]
- Cui, L.; Du, W.; Xu, N.; Dong, J.; Xia, B.; Ma, J.; Yan, R.; Wang, L.; Feng, F. Impact of MicroRNAs in Interaction with Environmental Factors on Autism Spectrum Disorder: An Exploratory Pilot Study. Front. Psychiatry 2021, 12, 715481. [Google Scholar] [CrossRef]
- Kichukova, T.M.; Popov, N.T.; Ivanov, I.S.; Vachev, T.I. Profiling of Circulating Serum MicroRNAs in Children with Autism Spectrum Disorder using Stem-loop qRT-PCR Assay. Folia Medica 2017, 59, 43–52. [Google Scholar] [CrossRef]
- Yu, D.; Jiao, X.; Cao, T.; Huang, F. Serum miRNA expression profiling reveals miR-486-3p may play a significant role in the development of autism by targeting ARID1B. NeuroReport 2018, 29, 1431–1436. [Google Scholar] [CrossRef]
- Abdelrahman, A.H.; Eid, O.M.; Ibrahim, M.H.; El-Fattah, S.N.A.; Eid, M.M.; Meguid, N.A. Evaluation of circulating miRNAs and mRNAs expression patterns in autism spectrum disorder. Egypt. J. Med. Hum. Genet. 2021, 22, 81. [Google Scholar] [CrossRef]
- Ragusa, M.; Santagati, M.; Mirabella, F.; Lauretta, G.; Cirnigliaro, M.; Brex, D.; Barbagallo, C.; Domini, C.N.; Gulisano, M.; Barone, R.; et al. Potential Associations Among Alteration of Salivary miRNAs, Saliva Microbiome Structure, and Cognitive Impairments in Autistic Children. Int. J. Mol. Sci. 2020, 21, 6203. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.S.; Fregeac, J.; Bole-Feysot, C.; Cagnard, N.; Iyer, A.; Anink, J.; Aronica, E.; Alibeu, O.; Nitschke, P.; Colleaux, L. Role of miR-146a in neural stem cell differentiation and neural lineage determination: Relevance for neurodevelopmental disorders. Mol. Autism 2018, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Bellido-Cuéllar, S.; de la Fuente, R.P.; Lezana-Rosales, J.M.; Sánchez-Calvín, M.T.; Saiz-Díaz, R.A.; de la Aleja, J.G. Epilepsy and Autism spectrum disorder caused by a pathogenic variant in TNRC6B. Seizure Eur. J. Epilepsy 2023, 110, 117–118. [Google Scholar] [CrossRef] [PubMed]
- Granadillo, J.L.; Stegmann, A.P.; Guo, H.; Xia, K.; Angle, B.; Bontempo, K.; Ranells, J.D.; Newkirk, P.; Costin, C.; Viront, J.; et al. Pathogenic variants in TNRC6B cause a genetic disorder characterised by developmental delay/intellectual disability and a spectrum of neurobehavioural phenotypes including autism and ADHD. J. Med. Genet. 2020, 57, 717. [Google Scholar] [CrossRef]
- Peterson, S.M.; Thompson, J.A.; Ufkin, M.L.; Sathyanarayana, P.; Liaw, L.; Congdon, C.B. Common features of microRNA target prediction tools. Front. Genet. 2014, 5, 23. [Google Scholar] [CrossRef]
- Sukranli, Z.Y.; Bayram, K.K.; Mehmetbeyoglu, E.; Doganyigit, Z.; Beyaz, F.; Sener, E.F.; Taheri, S.; Ozkul, Y.; Rassoulzadegan, M. Trans Species RNA Activity: Sperm RNA of the Father of an Autistic Child Programs Glial Cells and Behavioral Disorders in Mice. Biomolecules 2024, 14, 201. [Google Scholar] [CrossRef]
- Fridrich, A.; Hazan, Y.; Moran, Y. Too Many False Targets for MicroRNAs: Challenges and Pitfalls in Prediction of miRNA Targets and Their Gene Ontology in Model and Non-model Organisms. BioEssays 2019, 41, 1800169. [Google Scholar] [CrossRef]
- Godard, P.; van Eyll, J. Pathway analysis from lists of microRNAs: Common pitfalls and alternative strategy. Nucleic Acids Res. 2015, 43, 3490–3497. [Google Scholar] [CrossRef]
- Wen, Y.; Alshikho, M.J.; Herbert, M.R. Pathway Network Analyses for Autism Reveal Multisystem Involvement, Major Overlaps with Other Diseases and Convergence upon MAPK and Calcium Signaling. PLoS ONE 2016, 11, e0153329. [Google Scholar] [CrossRef]
- Gabrielli, A.P.; Manzardo, A.M.; Butler, M.G. GeneAnalytics Pathways and Profiling of Shared Autism and Cancer Genes. Int. J. Mol. Sci. 2019, 20, 1166. [Google Scholar] [CrossRef]
- Huang, Z.-X.; Chen, Y.; Guo, H.-R.; Chen, G.-F. Systematic Review and Bioinformatic Analysis of microRNA Expression in Autism Spectrum Disorder Identifies Pathways Associated with Cancer, Metabolism, Cell Signaling, and Cell Adhesion. Front. Psychiatry 2021, 12, 630876. [Google Scholar] [CrossRef] [PubMed]
- Forés-Martos, J.; Catalá-López, F.; Sánchez-Valle, J.; Ibáñez, K.; Tejero, H.; Palma-Gudiel, H.; Climent, J.; Pancaldi, V.; Fañanás, L.; Arango, C.; et al. Transcriptomic metaanalyses of autistic brains reveals shared gene expression and biological pathway abnormalities with cancer. Mol. Autism 2019, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Vorstman, J.A.; Scherer, S.W. Contemplating syndromic autism. Genet. Med. 2023, 25, 100919. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enrichment Analysis | Biological Pathways Enrichment Map | Human Phenotypes Ontology Enrichment Map |
|---|---|---|
| OC = 0.5 | JC = 0.3 |
| OC = 0.5 | OC = 0.5 |
| OC = 0.5 | OC = 0.5 |
| Sperm | Neurons | PBMCs |
|---|---|---|
| hsa-miR-424-5p | hsa-miR-432-5p | hsa-miR-451a |
| hsa-miR-619-5p | hsa-miR-196a-5p | hsa-miR-320a-3p |
| hsa-miR-193b-3p | hsa-miR-494-3p | hsa-miR-144-3p |
| hsa-miR-664a-3p |
| Sperm and Neurons | Sperm and PBMCs | Neurons and PBMCs |
|---|---|---|
| hsa-miR-106a-5p | hsa-miR-223-3p | hsa-miR-874-3p |
| hsa-miR-379-5p | hsa-miR-142-3p | hsa-miR-199a-5p |
| hsa-miR-148a-5p | hsa-miR-142-5p | |
| hsa-miR-146a-5p | ||
| hsa-miR-15a-5p | ||
| hsa-miR-29a-3p | ||
| hsa-miR-29b-3p | ||
| hsa- miR-29c-3p, | ||
| hsa-miR-19a-3p | ||
| hsa-miR-155-5p |
| miRNA | |
|---|---|
| hsa-miR-106a-5p | hsa-miR-7-5p |
| hsa-miR-223-3p | hsa-let-7a-5p |
| hsa-miR-142-3p | hsa-miR-128-3p |
| hsa-miR-146a-5p | hsa-miR-99a-5p |
| hsa-miR-155-5p | hsa-miR-15b-5p |
| hsa-miR-15a-5p | hsa-miR-484 |
| hsa-miR-29b-3p | hsa-miR-19b-3p |
| hsa-miR-29c-3p | hsa-miR-106b-5p |
| hsa-miR-19a-3p | hsa-miR-221-3p |
| hsa-miR-29a-3p | hsa-miR-574-3p |
| hsa-miR-424-5p | hsa-miR-93-5p |
| hsa-miR-619-5p | hsa-miR-27a-3p |
| hsa-miR-143-3p | hsa-miR-335-3p |
| hsa-miR-140-5p | hsa-miR-130a-3p |
| hsa-miR-23a-3p | hsa-miR-146b-5p |
| miRNA | References |
|---|---|
| hsa-let-7a-5p | [25] (blood); [68] (brain); [69] (saliva) |
| hsa-miR-93-5p | [70] (cerebellar cortex); [27] (monocytes); [71] (serum) |
| hsa-miR-27a-3p | [70] (cerebellar cortex); [72] (serum); [73] (saliva); [74] (blood) |
| hsa-miR-146b-5p | [70] (cerebellar cortex); [75] (saliva); [76] (brain) |
| hsa-miR-140-5p | [70] (cerebellar cortex); [76] (brain); [77] (saliva) |
| hsa-miR-23a-3p | [70] (cerebellar cortex); [73] (saliva); [78] (brain); [79] (saliva); [80] (brain); [81] (serum) |
| hsa-miR-7-5p | [70] (cerebellar cortex); [68] (brain); [73] (saliva); [71] (serum); [79] (saliva); [82] (plasma) |
| hsa-miR-15b-5p | [70] (cerebellar cortex); [25] (blood); [83] (blood) |
| hsa-miR-484 | [70] (cerebellar cortex); [78] (brain); [84] (plasma) |
| hsa-miR-19b-3p | [72] (serum); [25] (blood); [68] (brain); [85] (serum) |
| hsa-miR-106b-5p | [70] (cerebellar cortex); [72] (serum); [86] (serum); [87] (serum); [82] (plasma) |
| hsa-miR-221-3p | [78] (brain); [80] (brain); [82] (plasma) |
| hsa-miR-574-3p | [25] (blood); [71] (serum); [76] (brain) |
| hsa-miR-130a-3p | [72] (serum); [76] (brain) |
| hsa-miR-335-3p | [73] (saliva); [78] (brain) |
| hsa-miR-143-3p | [80] (brain); [77] (saliva) |
| hsa-miR-128-3p | [70] (cerebellar cortex); [88] (serum) |
| hsa-miR-99a-5p | [80] (brain); [82] (plasma) |
| miRNA | Total | Sperm and Neurons | Sperm, Neurons, and PBMCs | Target Genes Related to Autism | ||
|---|---|---|---|---|---|---|
| Total | Sperm and Neurons | Sperm, Neurons, and PBMCs | ||||
| hsa-miR-335-3p | 2549 | 822 | 488 | 292 | 120 | 54 |
| hsa-miR-93-5p | 2017 | 754 | 476 | 227 | 110 | 57 |
| hsa-miR-106b-5p | 2002 | 720 | 456 | 211 | 97 | 49 |
| hsa-miR-15b-5p | 1898 | 715 | 451 | 219 | 100 | 59 |
| hsa-miR-484 | 1861 | 749 | 534 | 196 | 102 | 72 |
| hsa-miR-27a-3p | 1827 | 655 | 390 | 210 | 102 | 52 |
| hsa-miR-128-3p | 1807 | 636 | 379 | 201 | 104 | 52 |
| hsa-miR-23a-3p | 1726 | 574 | 359 | 191 | 92 | 48 |
| hsa-miR-7-5p | 1694 | 623 | 393 | 186 | 96 | 54 |
| hsa-miR-19b-3p | 1675 | 630 | 392 | 243 | 125 | 61 |
| hsa-let-7a-5p | 1585 | 539 | 338 | 152 | 75 | 37 |
| hsa-miR-143-3p | 1478 | 557 | 334 | 185 | 97 | 49 |
| hsa-miR-221-3p | 1443 | 559 | 373 | 186 | 92 | 53 |
| hsa-miR-130a-3p | 1351 | 501 | 316 | 169 | 73 | 39 |
| hsa-miR-146b-5p | 1331 | 489 | 299 | 161 | 80 | 39 |
| hsa-miR-140-5p | 1229 | 487 | 308 | 181 | 99 | 50 |
| hsa-miR-99a-5p | 1037 | 405 | 263 | 103 | 60 | 30 |
| hsa-miR-574-3p | 954 | 345 | 213 | 100 | 49 | 23 |
| miRNA | Metascape | gProfiler and EnrichmentMap |
|---|---|---|
| hsa-miR-335-3p | 2 | 5 |
| hsa-miR-93-5p | 1 | 3 |
| hsa-miR-106b-5p | 3 | 3 |
| hsa-miR-15b-5p | 3 | 4 |
| hsa-miR-484 | 1 | 1 |
| hsa-miR-27a-3p | 2 | 3 |
| hsa-miR-128-3p | 3 | 3 |
| hsa-miR-23a-3p | 0 | 3 |
| hsa-miR-7-5p | 2 | 4 |
| hsa-miR-19b-3p | 1 | 2 |
| hsa-let-7a-5p | 1 | 1 |
| hsa-miR-143-3p | 2 | 2 |
| hsa-miR-221-3p | 3 | 1 |
| hsa-miR-130a-3p | 2 | 2 |
| hsa-miR-146b-5p | 5 | 4 |
| hsa-miR-140-5p | 3 | 4 |
| hsa-miR-99a-5p | 0 | 0 |
| hsa-miR-574-3p | 0 | 1 |
| miRNA | FDR | Enrichment |
|---|---|---|
| has-miR-19b-3p | 4.96 × 10−8 | 0.97 |
| has-miR-15b-5p | 0.00035 | 0.727 |
| hsa-miR-21-5p | 0.000353 | 0.787 |
| hsa-miR-16-5p | 0.00724 | 0.447 |
| hsa-miR-744-5p | 0.0126 | 0.77 |
| hsa-miR-181a-5p | 0.0149 | 0.678 |
| hsa-miR-92a-3p | 0.0165 | 0.432 |
| has-miR-221-3p | 0.0337 | 0.749 |
| hsa-miR-181b-5p | 0.0385 | 0.741 |
| miRNA | Overlap | Enrichment | FDR |
|---|---|---|---|
| Brain development (GO:0007420) | |||
| hsa-miR-223-3p | 19 | 1.839 | 0.001298270972 |
| hsa-miR-126-3p | 13 | 2.075 | 0.003057508899 |
| hsa-miR-138-5p | 18 | 1.617 | 0.007269372947 |
| hsa-miR-6740-5p | 11 | 2.103 | 0.007269372947 |
| hsa-miR-34a-3p | 14 | 1.734 | 0.0116666655 |
| hsa-miR-6806-5p | 10 | 2.018 | 0.01729235795 |
| hsa-let-7c-3p | 14 | 1.634 | 0.02040906023 |
| hsa-miR-31-3p | 12 | 1.756 | 0.02282151648 |
| hsa-miR-9-3p | 14 | 1.593 | 0.0245245798 |
| hsa-miR-133b | 12 | 1.639 | 0.03731966297 |
| hsa-miR-145-3p | 11 | 1.668 | 0.04186509355 |
| hsa-miR-4260 | 11 | 1.686 | 0.04186509355 |
| hsa-miR-4699-3p | 12 | 1.591 | 0.04186509355 |
| Forebrain development (GO:0030900) | |||
| hsa-miR-200c-3p | 22 | 1.96 | 7.98 × 10−5 |
| hsa-miR-34a-3p | 12 | 2.253 | 0.004230976391 |
| hsa-miR-138-5p | 13 | 1.909 | 0.009118847038 |
| hsa-miR-145-3p | 10 | 2.259 | 0.009118847038 |
| hsa-miR-200b-3p | 16 | 1.696 | 0.009118847038 |
| hsa-miR-31-3p | 10 | 2.226 | 0.009118847038 |
| hsa-miR-152-3p | 14 | 1.795 | 0.01067551065 |
| hsa-miR-126-3p | 9 | 2.273 | 0.01121765579 |
| hsa-miR-9-3p | 11 | 1.995 | 0.01339422516 |
| hsa-miR-429 | 13 | 1.69 | 0.02892920941 |
| hsa-miR-22-3p | 14 | 1.604 | 0.03007451519 |
| hsa-miR-223-3p | 11 | 1.814 | 0.0332969535 |
| hsa-let-7c-3p | 10 | 1.9 | 0.03376666551 |
| Neurogenesis (GO: 0022008) | |||
| hsa-miR-223-3p | 30 | 1.641 | 8.01 × 10−6 |
| hsa-miR-1179 | 17 | 1.737 | 0.001535995179 |
| hsa-miR-126-3p | 16 | 1.542 | 0.007377225259 |
| hsa-miR-127-3p | 9 | 1.791 | 0.02587966401 |
| hsa-miR-4632-3p | 9 | 1.627 | 0.04208567062 |
| Generation of neurons (GO: 0048699) | |||
| hsa-miR-223-3p | 28 | 1.756 | 1.27 × 10−5 |
| hsa-miR-1179 | 15 | 1.767 | 0.003310869485 |
| hsa-miR-31-3p | 16 | 1.54 | 0.01157149572 |
| Nervous system development (R-HSA-9675108) | |||
| hsa-miR-100-5p | 28 | 1.599 | 0.0001058059054 |
| hsa-miR-652-3p | 17 | 1.569 | 0.01202419931 |
| hsa-miR-452-5p | 11 | 1.945 | 0.01686604689 |
| Synaptic signaling pathways associated with autism spectrum disorder (WP4539) | |||
| hsa-miR-126-3p | 6 | 3.78 | 0.0001311122431 |
| hsa-miR-451a | 4 | 3.458 | 0.01312953894 |
| hsa-miR-487a-3p | 4 | 3.325 | 0.02304742794 |
| hsa-miR-19a-3p | 9 | 2.228 | 0.02807879676 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acerbi da Silva, L.N.; Stumpp, T. Bioinformatic Analysis of Autism-Related miRNAs and Their PoTential as Biomarkers for Autism Epigenetic Inheritance. Genes 2025, 16, 418. https://doi.org/10.3390/genes16040418
Acerbi da Silva LN, Stumpp T. Bioinformatic Analysis of Autism-Related miRNAs and Their PoTential as Biomarkers for Autism Epigenetic Inheritance. Genes. 2025; 16(4):418. https://doi.org/10.3390/genes16040418
Chicago/Turabian StyleAcerbi da Silva, Larissa Naísa, and Taiza Stumpp. 2025. "Bioinformatic Analysis of Autism-Related miRNAs and Their PoTential as Biomarkers for Autism Epigenetic Inheritance" Genes 16, no. 4: 418. https://doi.org/10.3390/genes16040418
APA StyleAcerbi da Silva, L. N., & Stumpp, T. (2025). Bioinformatic Analysis of Autism-Related miRNAs and Their PoTential as Biomarkers for Autism Epigenetic Inheritance. Genes, 16(4), 418. https://doi.org/10.3390/genes16040418