
Genome-Wide Identification and Expression Pattern of the NAC Gene Family in Panax notoginseng

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of the NAC Gene Family Members in P. notoginseng
2.2. Prediction and Analysis of the Physicochemical Properties of P. notoginseng NAC Proteins
2.3. Phylogenetic Analysis of the P. notoginseng NAC Gene Family
2.4. Gene Structure Analysis of the P. notoginseng NAC Gene Family
2.5. Promoter Analysis of the P. notoginseng NAC Gene Family
2.6. Motif Analysis of the P. notoginseng NAC Gene Family
2.7. Transcriptome Data Processing
3. Results and Analysis
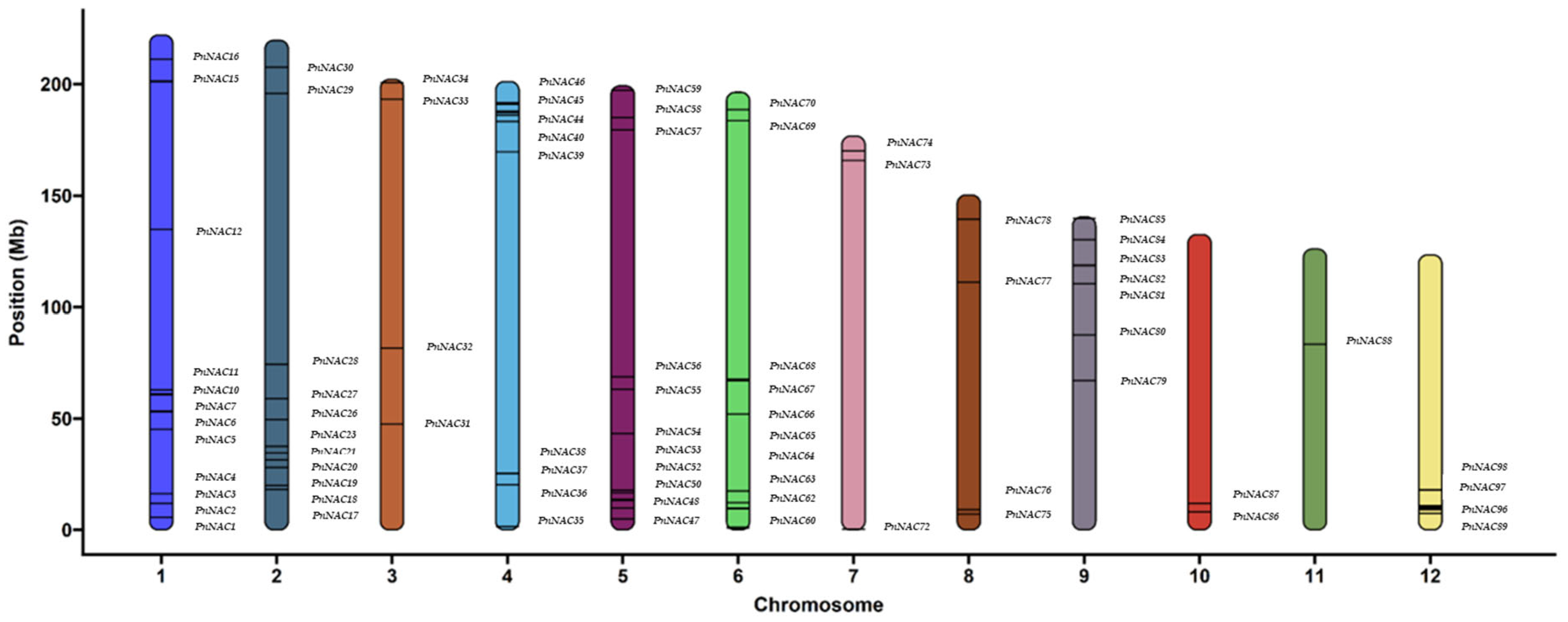
3.1. Identification of the P. notoginseng NAC Gene Family
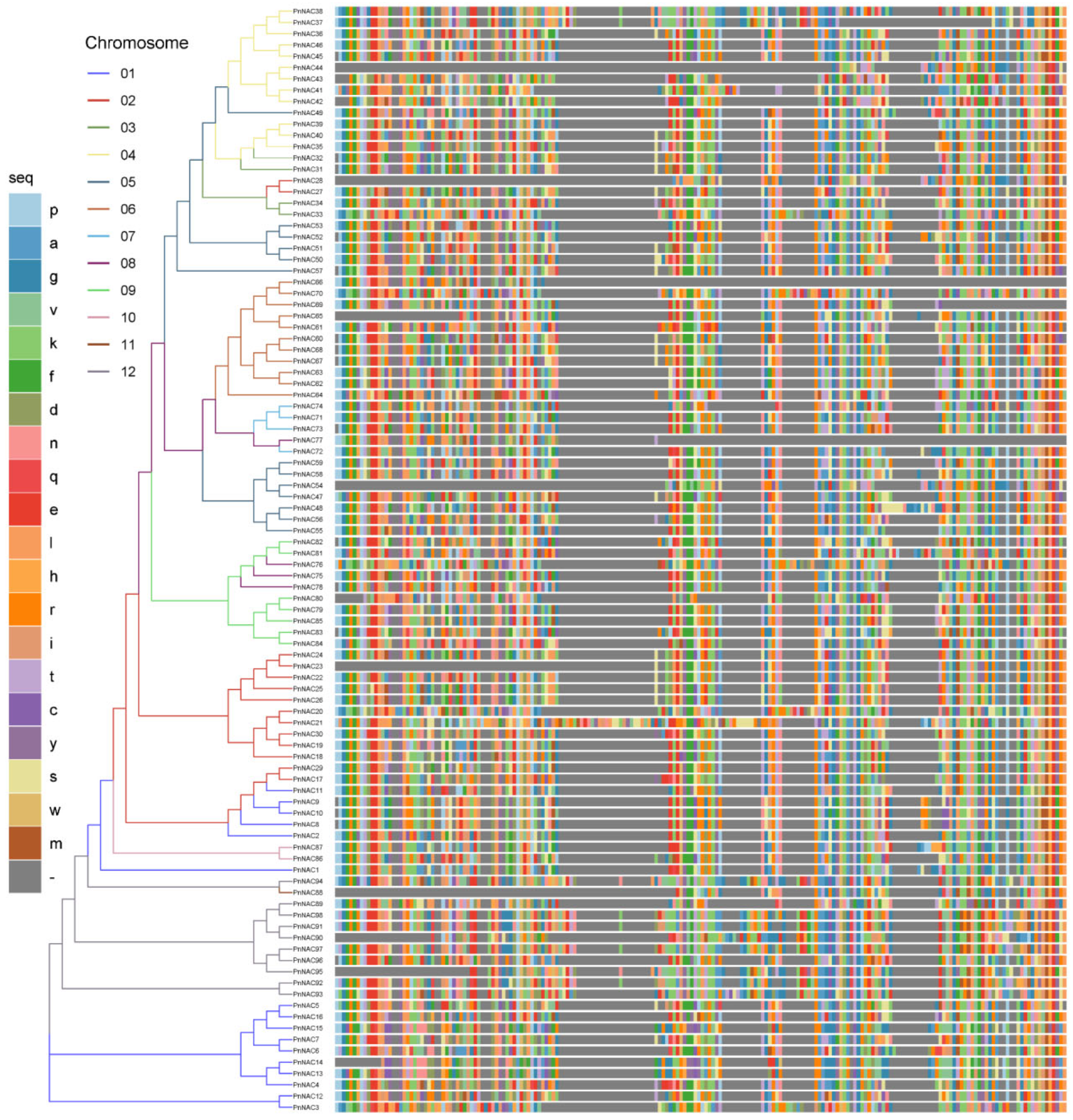
3.2. Phylogenetic Analysis of the P. notoginseng NAC Gene Family
3.3. Gene Structure Analysis of the P. notoginseng NAC Gene Family
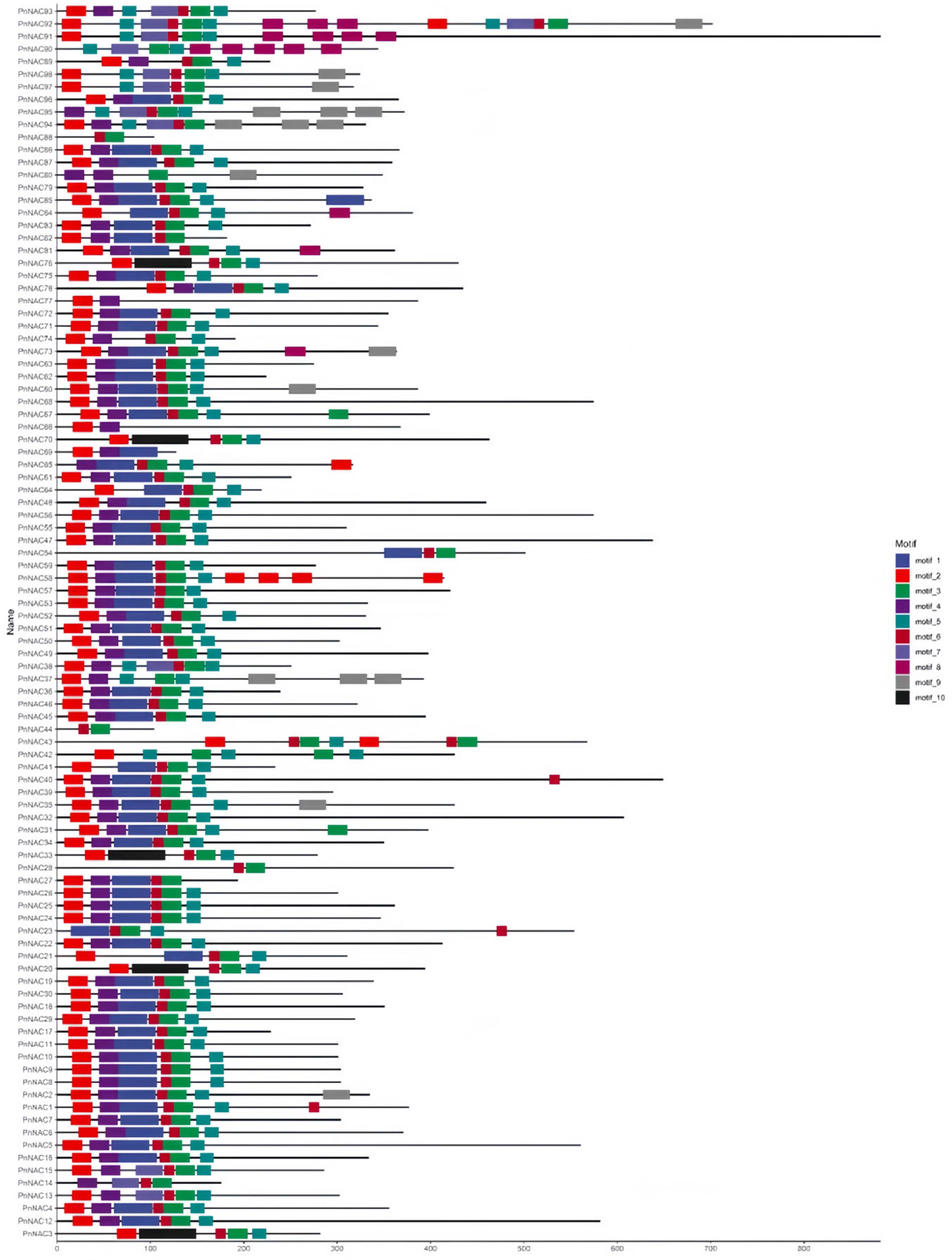
3.4. Motif Analysis of the P. notoginseng NAC Gene Family
3.5. Analysis of Cis-Acting Elements in the Promoters of the P. notoginseng NAC Gene Family
3.6. Expression Pattern Analysis of the P. notoginseng NAC Gene Family
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Briskin, D.P. Medicinal plants and phytomedicines. Linking plant biochemistry and physiology to human health. Plant Physiol. 2000, 124, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B. Pharmacological activity of sanchi ginseng (Panax notoginseng). J. Pharm. Pharmacol. 2006, 58, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; He, X.; Cui, H.; Cheng, Y.; Zhou, M.; Cui, X.; Zhang, T. Gut microbiota: A new key of understanding for Panax notoginseng against multiple disorders and biotransformation. J. Ethnopharmacol. 2025, 341, 119306. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Yu, M.; Guo, H.; Zeng, M.; Shao, Y.; Deng, W.; Qin, Q.; Li, Y.; Zhang, S. Panax notoginseng saponins inhibits oxidative stress-induced human nucleus pulposus cell apoptosis and delays disc degeneration in vivo and in vitro. J. Ethnopharmacol. 2024, 319, 117166. [Google Scholar] [CrossRef]
- Pan, Y.W.; Wu, D.P.; Liang, H.F.; Tang, G.Y.; Fan, C.L.; Shi, L.; Ye, W.C.; Li, M.M. Total saponins of Panax notoginseng activate Akt/mTOR pathway and exhibit neuroprotection in vitro and in vivo against ischemic damage. Chin. J. Integr. Med. 2022, 28, 410–418. [Google Scholar] [CrossRef]
- Liu, Y.; Li, S.; Pu, M.; Qin, H.; Wang, H.; Zhao, Y.; Chen, T. Structural characterization of polysaccharides isolated from Panax notoginseng medicinal residue and its protective effect on myelosuppression induced by cyclophosphamide. Chem. Biodivers. 2022, 19, e202100681. [Google Scholar] [CrossRef]
- Mao, Z.S.; Wei, F.G.; Chen, Z.J.; Yang, J.Y.; Wang, B.Y.; Wang, Y. Field investigation of round spot on sanqi (Panax notoginseng) in Yunnan province. J. Wenshan Univ. 2017, 30, 1–5. (In Chinese) [Google Scholar]
- He, C.; Wang, H.L.; Jin, X.; Jin, B.H.; Su, S.; Duan, Y.N.; He, X.H. Identification of Alternaria species acccosiated with black spot disease on Panax notoginseng in Yunnan and Guangxi. Plant Pathol. 2020, 50, 246–250. (In Chinese) [Google Scholar] [CrossRef]
- Chen, K.K.; Weng, F.; Qiang, Y. Control effect of two kinds of medicament on powdery mildew of sedum aizoon linn. J. Shaanxi Agric. Sci. 2014, 60, 24–25. (In Chinese) [Google Scholar]
- Li, Y.B.; Liu, Y.X.; Zhu, S.S.; Luo, L.X.; Li, J.Q. Grading system for Panax notoginseng root rot disease. Plant Pathol. 2020, 50, 450–461. (In Chinese) [Google Scholar] [CrossRef]
- Yang, L.F.; Yang, J.; Gao, L.L.; Guo, J.W.; Hong, L.; Cheng, J.X.; Kong, Q.; Tian, X.J. Screening and preliminary identification of soil bacteria antagonistic to phytophthora of Panax notoginseng. J. Honghe Univ. 2016, 14, 24–26+36. (In Chinese) [Google Scholar] [CrossRef]
- Shao, H.B.; Wang, H.Y.; Tang, X.L. NAC transcription factors in plant multiple abiotic stress responses: Progress and prospects. Front. Plant Sci. 2015, 6, 902. [Google Scholar] [CrossRef]
- Qu, Y.T.; Duan, M.; Zhang, Z.Q.; Dong, J.L.; Wang, T. Overexpression of the Medicago falcata NAC transcription factor MfNAC3 enhances cold tolerance in Medicago truncatula. Environ. Exp. Bot. 2016, 129, 67–76. [Google Scholar] [CrossRef]
- Evans, O. Functional Analysis of GhNAC18 and GhNAC20 Genes in Upland Cotton Leaf Senescence and Stress Response; Chinese Academy of Agricultural Sciences: Beijing, China, 2016. (In Chinese) [Google Scholar]
- Mao, X.G.; Chen, S.S.; Li, A.; Zhai, C.C.; Jing, R.L. Novel NAC transcription factor TaNAC67 confers enhanced multi-abiotic stress tolerances in Arabidopsis. PLoS ONE 2014, 9, e84359. [Google Scholar] [CrossRef]
- Chen, N.; Wu, S.H.; Fu, J.L.; Cao, B.H.; Lei, J.J.; Chen, C.M.; Jiang, J. Overexpression of the eggplant (Solanum melongena) NAC family transcription factor SmNAC suppresses resistance to bacterial wilt. Sci. Rep. 2016, 6, 31568. [Google Scholar] [CrossRef]
- Shen, J.B.; LV, B.; Luo, L.Q.; He, J.M.; Mao, C.J.; Xi, D.D.; Ming, F. The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 11, 40641. [Google Scholar] [CrossRef]
- Liu, Q.; Sun, C.Y.; Han, J.Z.; Li, L.; Wang, K.Y.; Wang, Y.F.; Chen, J.; Zhao, M.Z.; Wang, Y.; Zhang, M.P. Identification, characterization and functional differentiation of the NAC gene family and its roles in response to cold stress in ginseng, Panax ginseng C.A. Meyer. PLoS ONE 2020, 15, e0234423. [Google Scholar] [CrossRef]
- Yang, Z.J.; Liu, G.Z.; Zhang, G.H.; Yan, J.; Dong, Y.; Lu, Y.; Fan, W.; Hao, B.; Lin, Y.; Li, Y.; et al. The chromosome-scale high-quality genome assembly of Panax notoginseng provides insight into dencichine biosynthesis. Plant Biotechnol. J. 2021, 19, 869–871. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. Hmmer web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.Z.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.C.; David, K.S.; Zhu, H.C.; Guan, Y.; Tommy, T.-Y.L. Ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Bailey, T.L.; Nadya, W.; Chris, M.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Simon, A.; Theodor, P.P.; Wolfgang, H. HTSeq—A Python framework to work with high-throughput sequencing data. Bioimformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Wang, C.Y.; Zhang, Q. Research progress on the function of NAC transcription factors in plants. Biotechnol. Bull. 2018, 34, 8–14. (In Chinese) [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Chen, S.; Lin, X.; Zhang, D.; Li, Q.; Zhao, X.Y.; Su, C. Genome-wide analysis of NAC gene family in Betula pendula. Forests 2019, 10, 741. [Google Scholar] [CrossRef]
- Diao, W.P.; Snyder, J.C.; Wang, S.B.; Liu, J.B.; Pan, B.G.; Guo, G.J.; Ge, W.; Dawood, M.H.S.A. Genome-Wide Analyses of the NAC Transcription Factor Gene Family in Pepper (Capsicum annuum L.): Chromosome Location, Phylogeny, Structure, Expression Patterns, Cis-Elements in the Promoter, and Interaction Network. Int. J. Mol. Sci. 2018, 19, 1028. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, V.; Pal, A.K.; Acharya, V.; Ahuja, P.S. Genome-wide organization and expression profiling of the NAC transcription factor family in potato (Solanum tuberosum L.). DNA Res. 2013, 20, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.B.; Xu, X.Z.; Meng, Z.G.; Zhang, G.H.; Yang, S.C.; Chen, Z.J.; Wei, F.G.; Chen, J.W. Effects of light transmittance on plant growth and root ginsenoside content of Panax notoginseng. Chin. J. Appl. Environ. Biol. 2015, 21, 279–286. Available online: http://s.dic.cool/S/iRaXlslu (accessed on 3 March 2025). (In Chinese).
- Zhang, J.Y.; Zhang, Q.H.; Shang, S.P.; Cun, Z.; Wu, H.M.; Chen, J.W. The Responses of light reaction of photosynthesis to dynamic sun flecks in a typically shade-tolerant species Panax notoginseng. Front. Plant Sci. 2021, 12, 718981. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Max | Median | Mean | Max | |
|---|---|---|---|---|
| Length (AA) | 104.00 | 344.00 | 357.00 | 882.00 |
| Molecular weight (kDa) | 11.78 | 38.73 | 40.70 | 100.20 |
| IS | 4.12 | 6.52 | 6.54 | 9.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, B.; Hu, X.; Li, N.; Li, X.; Chen, Z.; Zhao, X.; Wu, X. Genome-Wide Identification and Expression Pattern of the NAC Gene Family in Panax notoginseng. Genes 2025, 16, 320. https://doi.org/10.3390/genes16030320
Jin B, Hu X, Li N, Li X, Chen Z, Zhao X, Wu X. Genome-Wide Identification and Expression Pattern of the NAC Gene Family in Panax notoginseng. Genes. 2025; 16(3):320. https://doi.org/10.3390/genes16030320
Chicago/Turabian StyleJin, Baihui, Xiaolong Hu, Na Li, Xiaohua Li, Zebin Chen, Xinyu Zhao, and Xiaoni Wu. 2025. "Genome-Wide Identification and Expression Pattern of the NAC Gene Family in Panax notoginseng" Genes 16, no. 3: 320. https://doi.org/10.3390/genes16030320
APA StyleJin, B., Hu, X., Li, N., Li, X., Chen, Z., Zhao, X., & Wu, X. (2025). Genome-Wide Identification and Expression Pattern of the NAC Gene Family in Panax notoginseng. Genes, 16(3), 320. https://doi.org/10.3390/genes16030320