
miR395e from Manihot esculenta Decreases Expression of PD-L1 in Renal Cancer: A Preliminary Study

 , , , , ,
, , , , ,  , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
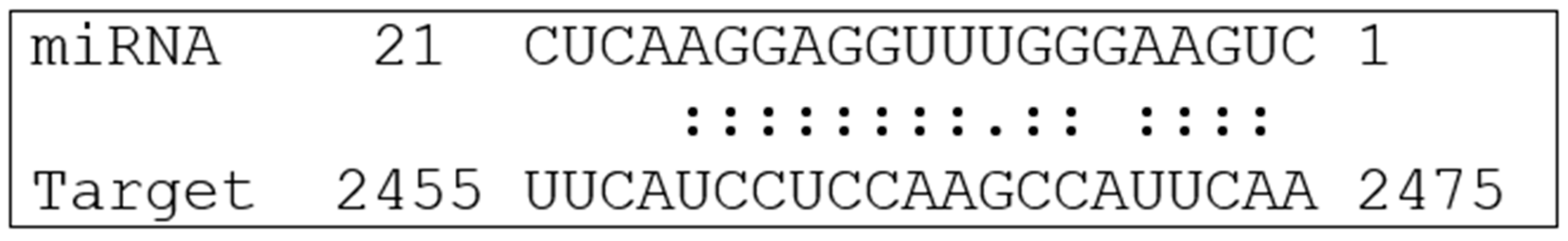
3.1. Bioinformatic Analysis Predicts Plant miRNAs That Have the Potential to Regulate PD-L1 Expression
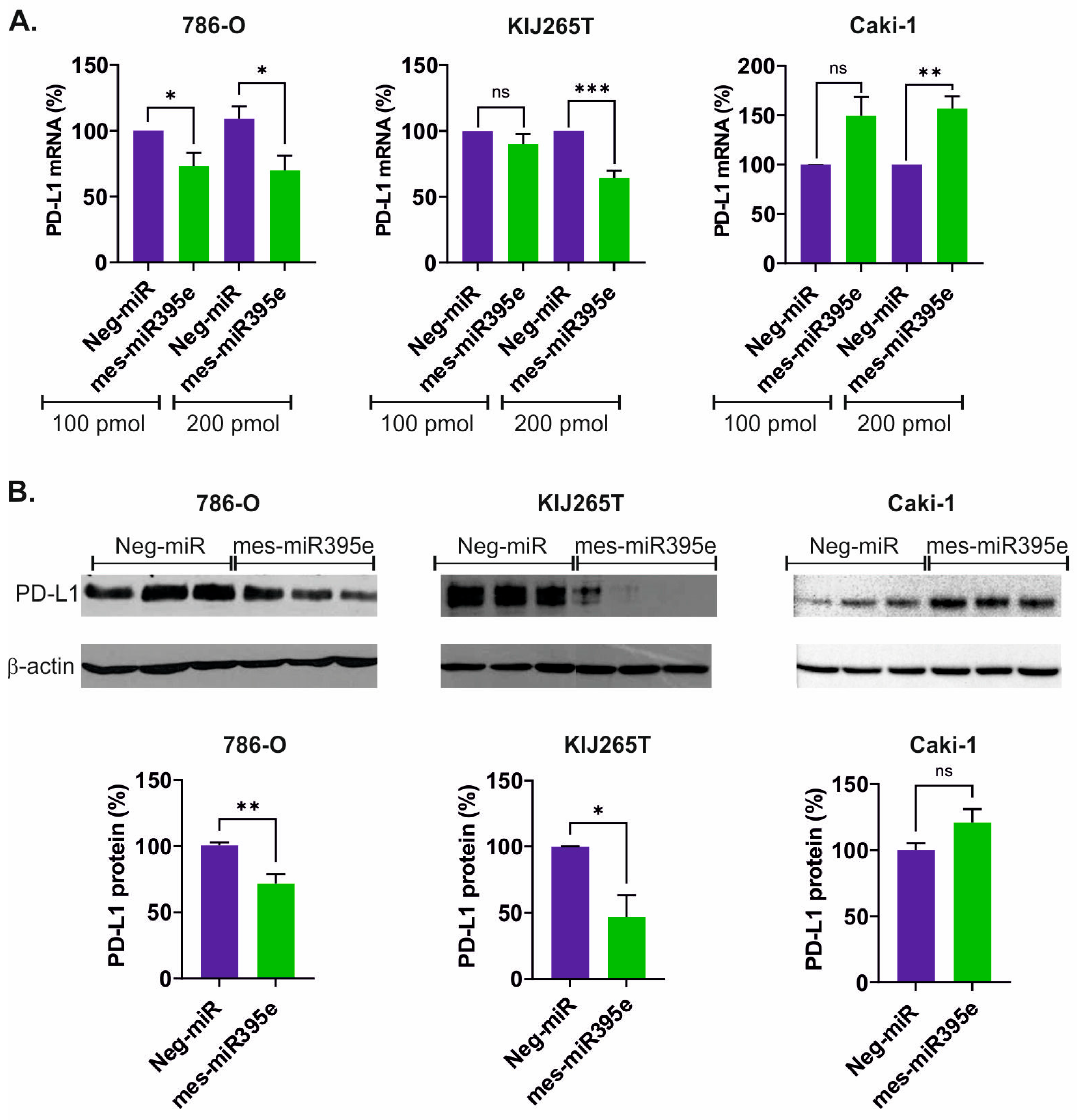
3.2. Mes-miR395e Attenuates PD-L1 Expression in Renal Cancer Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, Z.; Jin, Y.; Zhou, J.; Chen, F.; Chen, M.; Gao, Z.; Hu, L.; Xuan, J.; Li, X.; Song, Z.; et al. PD1/PD-L1 blockade in clear cell renal cell carcinoma: Mechanistic insights, clinical efficacy, and future perspectives. Mol. Cancer 2024, 23, 146. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, M.; Yazdani, O.; Kahrizi, M.S.; Soltanzadeh, S.; Javididashtbayaz, H.; Mivefroshan, A.; Ilkhani, S.; Esbati, R. Clinical potential of PD-1/PD-L1 blockade therapy for renal cell carcinoma (RCC): A rapidly evolving strategy. Cancer Cell Int. 2022, 22, 401. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Li, G.; Zheng, Z.; Zhang, X.; Wang, S.; Jiang, B.; Li, X.; Wang, X.; Wu, G. The molecular code of kidney cancer: A path of discovery for gene mutation and precision therapy. Mol. Asp. Med. 2025, 101, 101335. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Bochtler, T.; Pauli, C.; Baciarello, G.; Delorme, S.; Hemminki, K.; Mileshkin, L.; Moch, H.; Oien, K.; Olivier, T.; et al. Cancer of unknown primary: ESMO Clinical Practice Guideline for diagnosis, treatment and follow-up. Ann. Oncol. 2023, 34, 228–246. [Google Scholar] [CrossRef]
- Bellin, M.F.; Valente, C.; Bekdache, O.; Maxwell, F.; Balasa, C.; Savignac, A.; Meyrignac, O. Update on Renal Cell Carcinoma Diagnosis with Novel Imaging Approaches. Cancers 2024, 16, 1926. [Google Scholar] [CrossRef]
- Najem, E.J.; Shaikh, M.J.S.; Shinagare, A.B.; Krajewski, K.M. Navigating advanced renal cell carcinoma in the era of artificial intelligence. Cancer Imaging 2025, 25, 16. [Google Scholar] [CrossRef] [PubMed]
- Boussios, S.; Devo, P.; Goodall, I.C.A.; Sirlantzis, K.; Ghose, A.; Shinde, S.D.; Papadopoulos, V.; Sanchez, E.; Rassy, E.; Ovsepian, S.V. Exosomes in the Diagnosis and Treatment of Renal Cell Cancer. Int. J. Mol. Sci. 2023, 24, 14356. [Google Scholar] [CrossRef]
- Brown, J.E.; Royle, K.L.; Gregory, W.; Ralph, C.; Maraveyas, A.; Din, O.; Eisen, T.; Nathan, P.; Powles, T.; Griffiths, R.; et al. Temporary treatment cessation versus continuation of first-line tyrosine kinase inhibitor in patients with advanced clear cell renal cell carcinoma (STAR): An open-label, non-inferiority, randomised, controlled, phase 2/3 trial. Lancet Oncol. 2023, 24, 213–227. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Powles, T.; Peltola, K.; de Velasco, G.; Burotto, M.; Suarez, C.; Ghatalia, P.; Iacovelli, R.; Lam, E.T.; Verzoni, E.; et al. Belzutifan versus Everolimus for Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2024, 391, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Boguslawska, J.; Kryst, P.; Poletajew, S.; Piekielko-Witkowska, A. TGF-beta and microRNA Interplay in Genitourinary Cancers. Cells 2019, 8, 1619. [Google Scholar] [CrossRef]
- Alshehri, B. Plant-derived xenomiRs and cancer: Cross-kingdom gene regulation. Saudi J. Biol. Sci. 2021, 28, 2408–2422. [Google Scholar] [CrossRef]
- Yang, L.; Feng, H. Cross-kingdom regulation by plant-derived miRNAs in mammalian systems. Animal. Model. Exp. Med. 2023, 6, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, J.; Yang, Y.; Liu, J.; Li, H.; Li, R.; Cao, C.; Shi, L.; Wu, W.; He, K. A Timely Review of Cross-Kingdom Regulation of Plant-Derived MicroRNAs. Front. Genet. 2021, 12, 613197. [Google Scholar] [CrossRef]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, F.; Weng, Z.; Sui, X.; Fang, Y.; Tang, X.; Shen, X. Soybean-derived miRNAs specifically inhibit proliferation and stimulate apoptosis of human colonic Caco-2 cancer cells but not normal mucosal cells in culture. Genomics 2020, 112, 2949–2958. [Google Scholar] [CrossRef]
- Marzano, F.; Caratozzolo, M.F.; Consiglio, A.; Licciulli, F.; Liuni, S.; Sbisa, E.; D’Elia, D.; Tullo, A.; Catalano, D. Plant miRNAs Reduce Cancer Cell Proliferation by Targeting MALAT1 and NEAT1: A Beneficial Cross-Kingdom Interaction. Front. Genet. 2020, 11, 552490. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.H.; Li, Z.L.; Shih, Y.J.; Bennett, J.A.; Whang-Peng, J.; Lin, H.Y.; Davis, P.J.; Wang, K. Herbal Medicines Attenuate PD-L1 Expression to Induce Anti-Proliferation in Obesity-Related Cancers. Nutrients 2019, 11, 2979. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.Y.; Chen, Y.C.; Lyu, W.Y.; He, X.Y.; Ye, Z.H.; Huang, C.Y.; He, X.L.; Chen, X.; Chen, X.; Zhang, B.; et al. Ginsenoside Rh2 augmented anti-PD-L1 immunotherapy by reinvigorating CD8(+) T cells via increasing intratumoral CXCL10. Pharmacol. Res. 2023, 198, 106988. [Google Scholar] [CrossRef]
- Ravindran Menon, D.; Li, Y.; Yamauchi, T.; Osborne, D.G.; Vaddi, P.K.; Wempe, M.F.; Zhai, Z.; Fujita, M. EGCG Inhibits Tumor Growth in Melanoma by Targeting JAK-STAT Signaling and Its Downstream PD-L1/PD-L2-PD1 Axis in Tumors and Enhancing Cytotoxic T-Cell Responses. Pharmaceuticals 2021, 14, 1081. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Lukasik, A.; Wojcikowski, M.; Zielenkiewicz, P. Tools4miRs-one place to gather all the tools for miRNA analysis. Bioinformatics 2016, 32, 2722–2724. [Google Scholar] [CrossRef] [PubMed]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef]
- Kertesz, M.; Iovino, N.; Unnerstall, U.; Gaul, U.; Segal, E. The role of site accessibility in microRNA target recognition. Nat. Genet. 2007, 39, 1278–1284. [Google Scholar] [CrossRef]
- Wu, H.J.; Ma, Y.K.; Chen, T.; Wang, M.; Wang, X.J. PsRobot: A web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 2012, 40, W22–W28. [Google Scholar] [CrossRef] [PubMed]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.S.; Tam, W.L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, W.; Giegerich, R. GUUGle: A utility for fast exact matching under RNA complementary rules including G-U base pairing. Bioinformatics 2006, 22, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Sturm, M.; Hackenberg, M.; Langenberger, D.; Frishman, D. TargetSpy: A supervised machine learning approach for microRNA target prediction. BMC Bioinform. 2010, 11, 292. [Google Scholar] [CrossRef] [PubMed]
- Vejnar, C.E.; Blum, M.; Zdobnov, E.M. miRmap web: Comprehensive microRNA target prediction online. Nucleic Acids Res. 2013, 41, W165–W168. [Google Scholar] [CrossRef]
- Poplawski, P.; Alseekh, S.; Jankowska, U.; Skupien-Rabian, B.; Iwanicka-Nowicka, R.; Kossowska, H.; Fogtman, A.; Rybicka, B.; Boguslawska, J.; Adamiok-Ostrowska, A.; et al. Coordinated reprogramming of renal cancer transcriptome, metabolome and secretome associates with immune tumor infiltration. Cancer Cell Int. 2023, 23, 2. [Google Scholar] [CrossRef] [PubMed]
- Hanusek, K.; Rybicka, B.; Poplawski, P.; Adamiok-Ostrowska, A.; Gluchowska, K.; Piekielko-Witkowska, A.; Boguslawska, J. TGF-beta1 affects the renal cancer miRNome and regulates tumor cells proliferation. Int. J. Mol. Med. 2022, 49, 52. [Google Scholar] [CrossRef]
- Mohidin, S.; Moshawih, S.; Hermansyah, A.; Asmuni, M.I.; Shafqat, N.; Ming, L.C. Cassava (Manihot esculenta Crantz): A Systematic Review for the Pharmacological Activities, Traditional Uses, Nutritional Values, and Phytochemistry. J. Evid.-Based Integr. Med. 2023, 28, 2515690X231206227. [Google Scholar] [CrossRef]
- Kool, E.T. Hydrogen bonding, base stacking, and steric effects in dna replication. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 1–22. [Google Scholar] [CrossRef]
- Garg, A.; Heinemann, U. A novel form of RNA double helix based on G.U and C.A(+) wobble base pairing. RNA 2018, 24, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Leontis, N.B.; Stombaugh, J.; Westhof, E. The non-Watson-Crick base pairs and their associated isostericity matrices. Nucleic Acids Res. 2002, 30, 3497–3531. [Google Scholar] [CrossRef]
- Tsumbu, C.N.; Deby-Dupont, G.; Tits, M.; Angenot, L.; Franck, T.; Serteyn, D.; Mouithys-Mickalad, A. Antioxidant and antiradical activities of Manihot esculenta Crantz (Euphorbiaceae) leaves and other selected tropical green vegetables investigated on lipoperoxidation and phorbol-12-myristate-13-acetate (PMA) activated monocytes. Nutrients 2011, 3, 818–838. [Google Scholar] [CrossRef] [PubMed]
- Montagnac, J.A.; Davis, C.R.; Tanumihardjo, S.A. Processing Techniques to Reduce Toxicity and Antinutrients of Cassava for Use as a Staple Food. Compr. Rev. Food Sci. Food Saf. 2009, 8, 17–27. [Google Scholar] [CrossRef]
- Henneh, I.T.; Ahlidja, W.; Alake, J.; Mohammed, H.; Boapeah, S.O.; Kwabil, A.; Malcolm, F.; Armah, F.A. Acute toxicity profile and gastroprotective potential of ethanolic leaf extract of Manihot esculenta Crantz. Sci. Afr. 2022, 17, e01284. [Google Scholar] [CrossRef]
- Sreejith, S.; Tom, J.; Sangeetha, V.P.; Vandana, U.; Xavier, J.; Jayaprakas, C.A.; Mohanan, P.V. Antineoplastic effects of cassava-cyanide extract on human glioblastoma (LN229) cells. Toxicon 2023, 232, 107200. [Google Scholar] [CrossRef]
- Idibie, C.A.; Davids, H.; Iyuke, S.E. Cytotoxicity of purified cassava linamarin to a selected cancer cell lines. Bioprocess. Biosyst. Eng. 2007, 30, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Abotbina, W.; Sapuan, S.M.; Ilyas, R.A.; Sultan, M.T.H.; Alkbir, M.F.M.; Sulaiman, S.; Harussani, M.M.; Bayraktar, E. Recent Developments in Cassava (Manihot esculenta) Based Biocomposites and Their Potential Industrial Applications: A Comprehensive Review. Materials 2022, 15, 6992. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jiang, G.; Jing, N.; Liu, X.; Li, Q.; Liang, W.; Liu, Z. Bilberry anthocyanin extracts enhance anti-PD-L1 efficiency by modulating gut microbiota. Food Funct. 2020, 11, 3180–3190. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Sun, Q.; Liu, Q.; Li, H.; Zhang, W.; Sun, C. Focus on immune checkpoint PD-1/PD-L1 pathway: New advances of polyphenol phytochemicals in tumor immunotherapy. Biomed. Pharmacother. 2022, 154, 113618. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Han, Y.; Wang, W.; Jo, H.; Kim, H.; Kim, S.; Yang, K.M.; Kim, S.J.; Dhanasekaran, D.N.; Song, Y.S. Phytochemicals in Cancer Immune Checkpoint Inhibitor Therapy. Biomolecules 2021, 11, 1107. [Google Scholar] [CrossRef] [PubMed]
- Ri, M.H.; Ma, J.; Jin, X. Development of natural products for anti-PD-1/PD-L1 immunotherapy against cancer. J. Ethnopharmacol. 2021, 281, 114370. [Google Scholar] [CrossRef]
- Zhang, Q.; Xie, Z.; Li, Y.; Zhu, Q.; Shi, H.; Zhao, R.; Yang, X.; Tian, J.; Ma, L. The potential of Lycium barbarum miR166a in kidney cancer treatment. Exp. Cell Res. 2023, 423, 113455. [Google Scholar] [CrossRef]
- Franzin, R.; Netti, G.S.; Spadaccino, F.; Porta, C.; Gesualdo, L.; Stallone, G.; Castellano, G.; Ranieri, E. The Use of Immune Checkpoint Inhibitors in Oncology and the Occurrence of AKI: Where Do We Stand? Front. Immunol. 2020, 11, 574271. [Google Scholar] [CrossRef]
- Santovito, D.; Weber, C. Non-canonical features of microRNAs: Paradigms emerging from cardiovascular disease. Nat. Rev. Cardiol. 2022, 19, 620–638. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Lin, Z. Autophagy Regulation by Crosstalk between miRNAs and Ubiquitination System. Int. J. Mol. Sci. 2021, 22, 11912. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogusławska, J.; Rakhmetullina, A.; Grzanka, M.; Białas, A.; Rybicka, B.; Życka-Krzesińska, J.; Molcan, T.; Zielenkiewicz, P.; Pączek, L.; Piekiełko-Witkowska, A. miR395e from Manihot esculenta Decreases Expression of PD-L1 in Renal Cancer: A Preliminary Study. Genes 2025, 16, 293. https://doi.org/10.3390/genes16030293
Bogusławska J, Rakhmetullina A, Grzanka M, Białas A, Rybicka B, Życka-Krzesińska J, Molcan T, Zielenkiewicz P, Pączek L, Piekiełko-Witkowska A. miR395e from Manihot esculenta Decreases Expression of PD-L1 in Renal Cancer: A Preliminary Study. Genes. 2025; 16(3):293. https://doi.org/10.3390/genes16030293
Chicago/Turabian StyleBogusławska, Joanna, Aizhan Rakhmetullina, Małgorzata Grzanka, Alex Białas, Beata Rybicka, Joanna Życka-Krzesińska, Tomasz Molcan, Piotr Zielenkiewicz, Leszek Pączek, and Agnieszka Piekiełko-Witkowska. 2025. "miR395e from Manihot esculenta Decreases Expression of PD-L1 in Renal Cancer: A Preliminary Study" Genes 16, no. 3: 293. https://doi.org/10.3390/genes16030293
APA StyleBogusławska, J., Rakhmetullina, A., Grzanka, M., Białas, A., Rybicka, B., Życka-Krzesińska, J., Molcan, T., Zielenkiewicz, P., Pączek, L., & Piekiełko-Witkowska, A. (2025). miR395e from Manihot esculenta Decreases Expression of PD-L1 in Renal Cancer: A Preliminary Study. Genes, 16(3), 293. https://doi.org/10.3390/genes16030293