
A Double-Gene Metabarcoding Approach for the Authentication of Shrimp Surimi-Based Products

 ,
,  , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
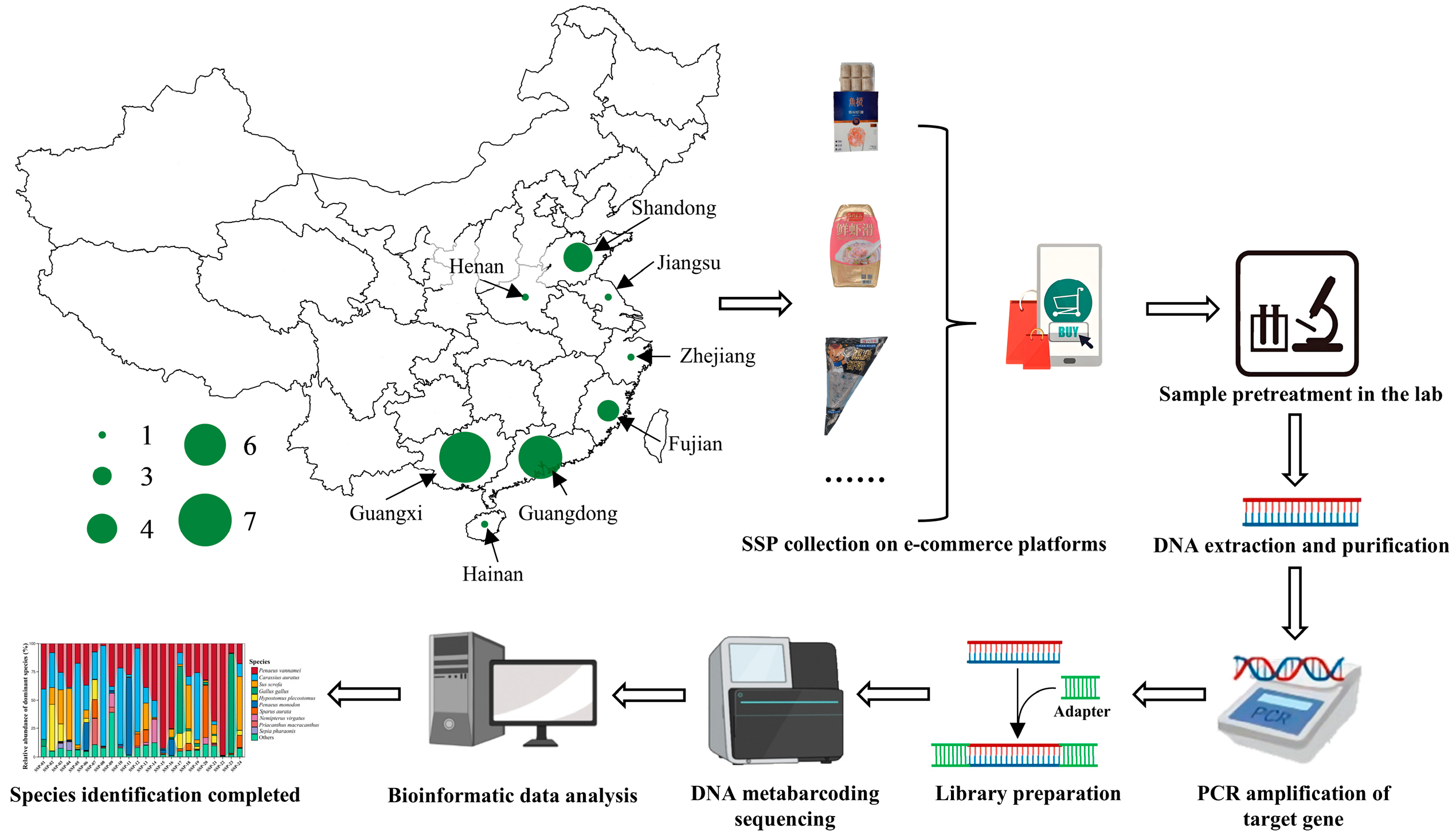

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Production Area (City or County, Province) | List of Ingredients | Declared Shrimp Species/Chinese Commercial Name | Other Declared Meat Species | Price (USD/kg) |
|---|---|---|---|---|---|
| SSP-01 | Weifang City, Shandong Province | Shrimp (N.W. ≥ 95%), fish surimi, vegetable oil, starch, water, egg white, chicken, trehalose, salt, sodium glutamate, sugar, chicken essence seasoning, compound phosphate | — | — | 23.98 |
| SSP-02 | Chengmai County, Hainan Province | Oriental river prawn (N.W. ≥ 65%), fish surimi, potato starch, egg white, salt, sugar, food additive | M. nipponense/青虾 | — | 13.07 |
| SSP-03 | Zhanjiang City, Guangdong Province | Shrimp (N.W. ≥ 95%), starch, salt, sodium glutamate, food additive | — | — | 18.43 |
| SSP-04 | Dongxing City, Guangxi Zhuang Autonomous Region | Shrimp (N.W. ≥ 80%), cuttlefish (N.W. ≥ 10%), pig fat, fish surimi, water, salt, sugar, starch, sesame oil, spices, acetate starch, tapioca starch, compound phosphate, food seasoning | — | — | 24.07 |
| SSP-05 | Anyang City, Henan Province | Oriental river prawn, fish surimi, starch, salt, egg white, sugar, sodium glutamate, vegetable oil, compound phosphate | M. nipponense/青虾 | — | 12.74 |
| SSP-06 | Zhangzhou City, Fujian Province | Shrimp (giant tiger prawn, Pacific white shrimp, N.W. ≥ 95%), starch, water, egg white, vegetable oil, salt, sugar, sodium glutamate, chicken essence seasoning, food additives | P. monodon/黑虎虾, P. vannamei/南美白对虾 | — | 40.39 |
| SSP-07 | Beihai City, Guangxi Zhuang Autonomous Region | Shrimp (N.W. ≥ 80%), fish surimi, starch, egg white, vegetable oil, trehalose, salt, sodium glutamate, compound phosphate | — | — | 19.26 |
| SSP-08 | Zhangzhou City, Fujian Province | Shrimp, frozen fish surimi, drinking water, starch, trehalose, vegetable oil, salt, sodium glutamate, chicken powder seasoning, yeast extract, food additives | — | — | 19.35 |
| SSP-09 | Rizhao City, Shandong Province | Shrimp, fish surimi (shrimp + fish surimi N.W. ≥ 95%), food additives, trehalose, water, vegetable oil, protein powder, salt, sodium glutamate, edible flavor | — | — | 18.43 |
| SSP-10 | Zhangzhou City, Fujian Province | Shrimp, frozen fish surimi, starch, water, vegetable oil, egg white, salt, sodium glutamate, chicken powder seasoning, yeast extract, food additives | — | — | 23.89 |
| SSP-11 | Beihai City, Guangxi Zhuang Autonomous Region | Giant tiger prawn (N.W. ≥ 95%), starch, egg white, vegetable oil, salt, water, sugar, chicken powder seasoning | P. monodon/黑虎虾 | — | 27.69 |
| SSP-12 | Shantou City, Guangdong Province | Shrimp (N.W. ≥ 85%), fish surimi, drinking water, starch, trehalose, sugar, compound acidity regulator, salt, sodium glutamate, egg white powder, frozen egg white, sodium tripolyphosphate, sodium hexametaphosphate, sodium pyrophosphate | — | — | 25.83 |
| SSP-13 | Zhanjiang City, Guangdong Province | Oriental river prawn (N.W. ≥ 95%), water, egg white, pork fat, chicken powder seasoning, salt, trehalose, sodium citrate, sodium D-isoascorbate, moisture retainer, acetate starch, hydroxypropyl distarch phosphate | M. nipponense/青虾 | — | 24.35 |
| SSP-14 | Taizhou City, Jiangsu Province | Shrimp, fish surimi, edible starch, sugar, salt, sodium glutamate, egg protein, food additives | — | — | 23.03 |
| SSP-15 | Beihai City, Guangxi Zhuang Autonomous Region | Shrimp (N.W. ≥ 90%), acetate starch, egg white, sugar, trehalose, salt, vegetable oil, sodium glutamate, compound water retention agent | — | — | 34.17 |
| SSP-16 | Zhanjiang City, Guangdong Province | Giant tiger prawn (N.W. ≥ 95%), water, protein liquid, pork fat, chicken powder seasoning, salt, trehalose, sodium citrate, sodium D-isoascorbate, moisture retainer, acetate starch, hydroxypropyl distarch phosphate | P. monodon/黑虎虾 | — | 27.91 |
| SSP-17 | Beihai City, Guangxi Zhuang Autonomous Region | Oriental river prawn, fish surimi, squid, starch, egg white, salt, sugar, monosodium glutamate, vegetable oil, food additives | M. nipponense/青虾 | — | 18.43 |
| SSP-18 | Beihai City, Guangxi Zhuang Autonomous Region | Shrimp, pork, starch, water, egg white liquid, sugar, salt, chicken powder seasoning, food additives | — | — | 22.57 |
| SSP-19 | Beihai City, Guangxi Zhuang Autonomous Region | Shrimp (N.W. ≥ 95%), fish surimi, potato starch, salt, frozen egg white, white sugar, chicken powder seasoning, compound water retaining agent | — | — | 30.83 |
| SSP-20 | Shantou City, Guangdong Province | Shrimp, fish surimi, edible starch, egg white, trehalose, salt, sugar, sodium glutamate, vegetable oil, compound water retention agent | — | — | 24.91 |
| SSP-21 | Weihai City, Shandong Province | Shrimp (N.W. ≥ 95%), fish surimi, water, starch, egg white liquid, sugar, salt, sodium glutamate, chicken powder seasoning, chicken broth seasoning, food additives | — | — | 22.22 |
| SSP-22 | Jiaxing City, Zhejiang Province | Shrimp (N.W. ≥ 80%), frozen egg white, starch, vegetable oil, flying fish roe (N.W. ≥ 2%), salt, sugar, sodium glutamate, egg white powder, trehalose, compound moisture retainer, curdlan, edible flavors and spices | — | Exocoetidae | 32.81 |
| SSP-23 | Weihai City, Shandong Province | Oriental river prawn (N.W. ≥ 95%), egg white, salt, peanut oil, starch, sodium glutamate | M. nipponense/青虾 | — | 13.70 |
| SSP-24 | Zhanjiang City, Guangdong Province | Oriental river prawn, water, water chestnuts, egg white liquid, pig fat, chicken powder seasoning, salt, trehalose, acetate starch, hydroxypropyl distarch phosphate, sodium citrate, sodium tripolyphosphate, sodium pyrophosphate | M. nipponense/青虾 | — | 11.94 |
2.2. DNA Extraction and PCR Amplification
2.3. Library Preparation and Sequencing
2.4. Processing of Sequencing Data and Sequence Taxonomic Assignment
2.5. Mislabeling Assessment and Evaluation of Environmental Impact
3. Results and Discussion
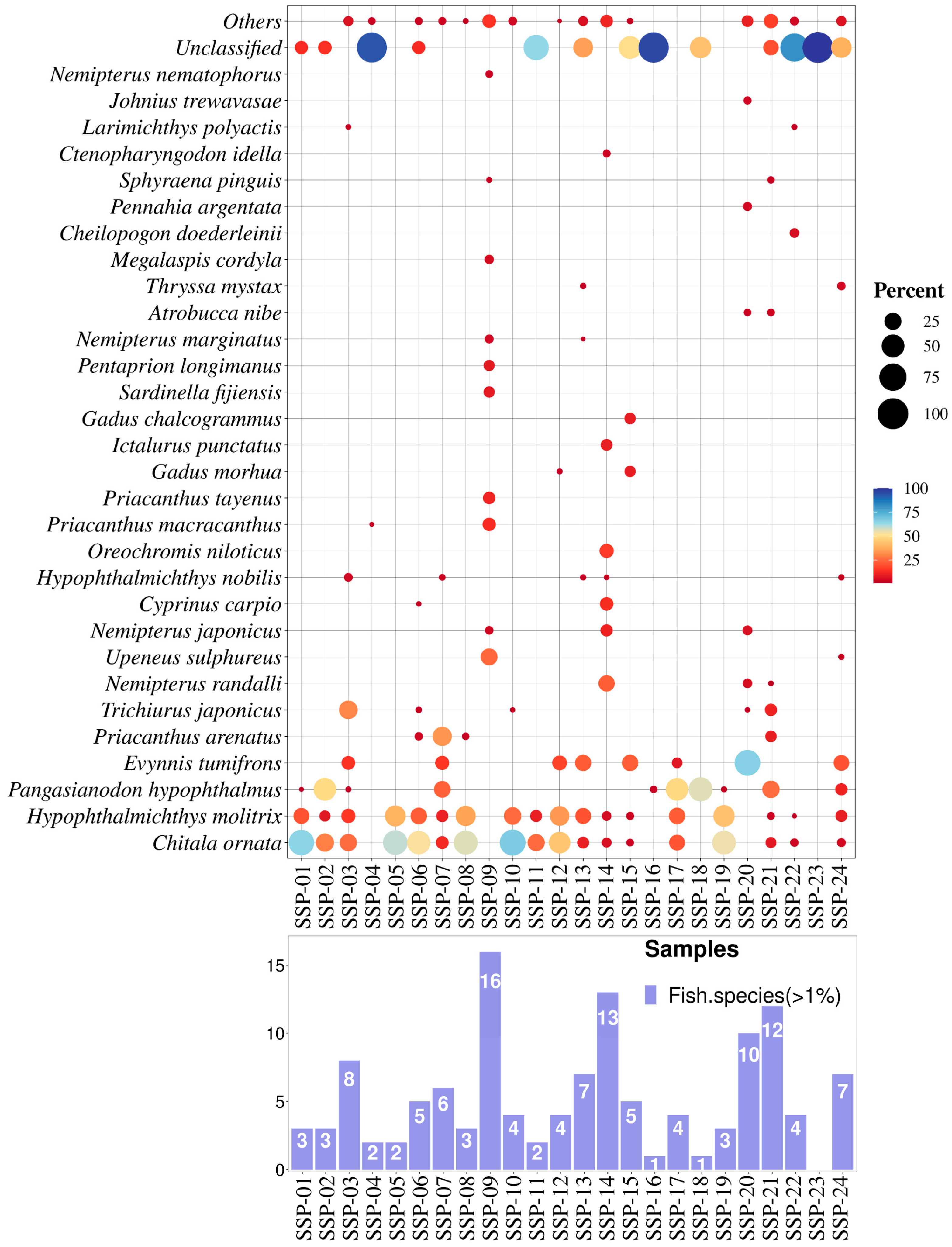
3.1. Sampling and SSP Declared Composition
3.2. Molecular Analysis and Taxonomic Assignment
3.3. Mislabeling Assessment and Evaluation of Environmental Impact
3.3.1. Mislabeling Assessment
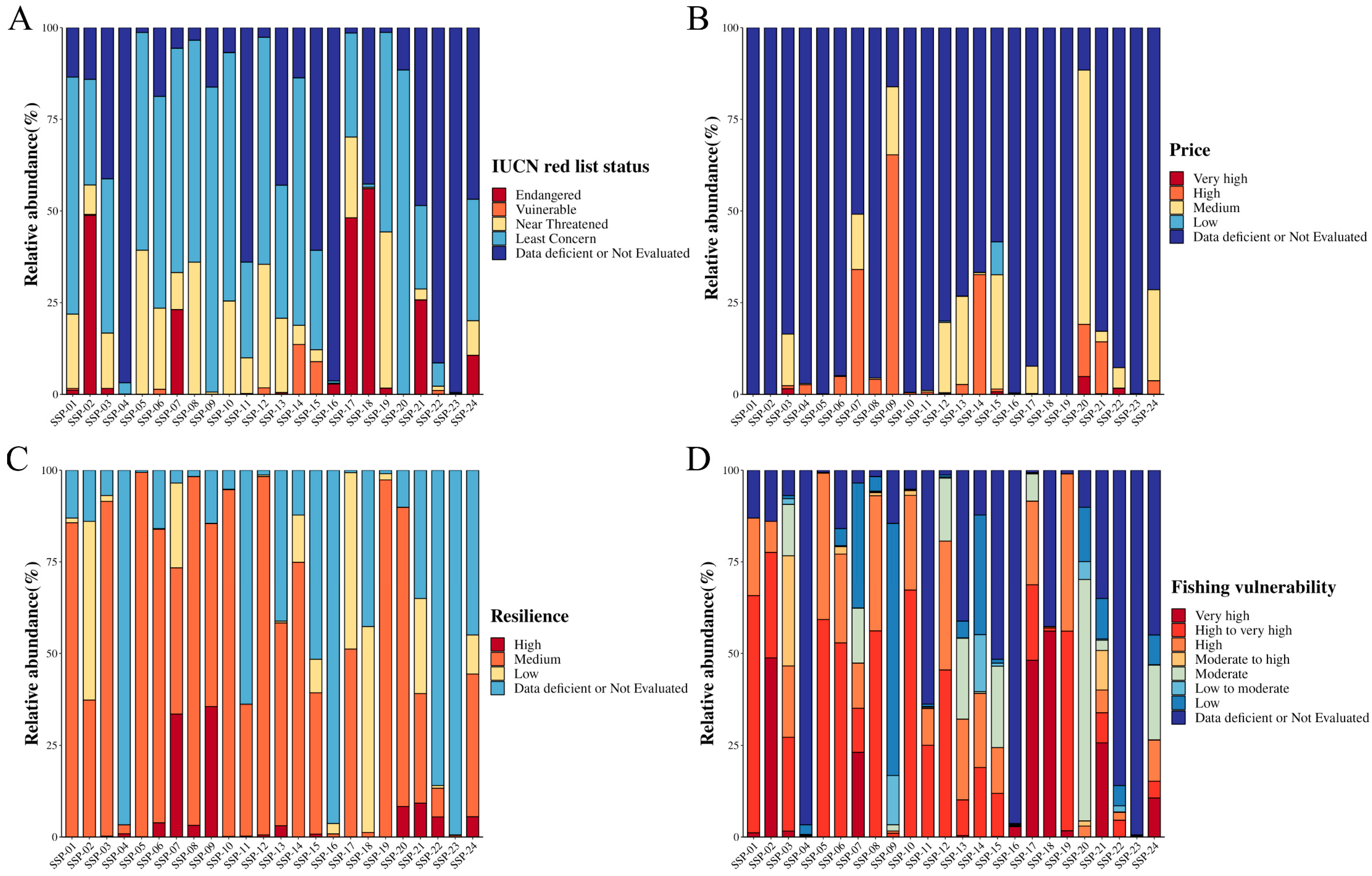
3.3.2. Fisheries Protection, Price, Resilience, and Vulnerability of Fish Species Used in SSPs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lawrence, S.; Elliott, C.; Huisman, W.; Dean, M.; van Ruth, S. Food fraud threats in UK post-harvest seafood supply chains; an assessment of current vulnerabilities. npj Sci. Food 2024, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2024—Blue Transformation in Action; FAO: Rome, Italy, 2024. [Google Scholar]
- BFFA. 2024 China Fishery Statistical Yearbook; China Agriculture Press Co., Ltd.: Beijing, China, 2024; pp. 1–144. (In Chinese) [Google Scholar]
- Quan, T.H.; Benjakul, S. Impact of salted duck egg albumen powder on proteolysis and gelling properties of sardine surimi. J. Texture Stud. 2019, 50, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Shui, S.-S.; Qi, H.; Shaimaa, H.; Aubourg, S.P.; Zhang, B. Kappa-carrageenan and its oligosaccharides maintain the physicochemical properties of myofibrillar proteins in shrimp mud (Xia-Hua) during frozen storage. J. Food Sci. 2021, 86, 140–148. [Google Scholar] [CrossRef]
- Xiao, H.; Yang, Y.; Yu, J.; Hu, M.; Xue, Y.; Xue, C. Alaska pollock surimi addition affects Pacific white shrimp (Litopenaeus vannamei) surimi gel properties. Rheol. Acta 2021, 60, 741–749. [Google Scholar] [CrossRef]
- Zhang, X.; Giusti, A.; Sun, Z.; Li, Y.; Guo, J.; Deng, W.; Chen, Y.; He, A.; Peng, H.; Tinacci, L.; et al. Corrigendum to “Molecular authentication of surimi-based products (fish cakes, 鱼糕) sold on the Chinese e-commerce: Traditional (DNA barcoding) and innovative techniques (metabarcoding) to tackle seafood fraud” [Food Control 155 (2024) 110110]. Food Control 2024, 160, 110332. [Google Scholar] [CrossRef]
- Xiong, X.; Yuan, F.; Huang, M.; Cao, M.; Xiong, X. Comparative Evaluation of Web Page and Label Presentation for Imported Seafood Products Sold on Chinese E-Commerce Platform and Molecular Identification Using DNA Barcoding. J. Food Prot. 2020, 83, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Giusti, A.; Malloggi, C.; Magagna, G.; Filipello, V.; Armani, A. Is the metabarcoding ripe enough to be applied to the authentication of foodstuff of animal origin? A systematic review. Compr. Rev. Food Sci. F 2024, 23, e13256. [Google Scholar] [CrossRef]
- Joenperä, J.; Vainio, A.; Lundén, J. External and internal food fraud prevention in Finnish food businesses. Food Control 2024, 164, 110496. [Google Scholar] [CrossRef]
- Robson, K.; Dean, M.; Haughey, S.; Elliott, C. A comprehensive review of food fraud terminologies and food fraud mitigation guides. Food Control 2021, 120, 107516. [Google Scholar] [CrossRef]
- Chin Chin, T.; Adibah, A.B.; Danial Hariz, Z.A.; Siti Azizah, M.N. Detection of mislabelled seafood products in Malaysia by DNA barcoding: Improving transparency in food market. Food Control 2016, 64, 247–256. [Google Scholar] [CrossRef]
- Galal-Khallaf, A.; Ardura, A.; Borrell, Y.J.; Garcia-Vazquez, E. Towards more sustainable surimi? PCR-cloning approach for DNA barcoding reveals the use of species of low trophic level and aquaculture in Asian surimi. Food Control 2016, 61, 62–69. [Google Scholar] [CrossRef]
- Giusti, A.; Armani, A.; Sotelo, C.G. Advances in the analysis of complex food matrices: Species identification in surimi-based products using Next Generation Sequencing technologies. PLoS ONE 2017, 12, e0185586. [Google Scholar] [CrossRef] [PubMed]
- Ooi, Z.S.; Jahari, P.N.S.; Sim, K.S.; Foo, S.X.; Mohd Zawai, N.N.; Mohd Salleh, F. DNA barcoding of commercial fish products using dual mitochondrial markers exposes evidence for mislabelling and trade of endangered species. IOP Conf. Ser. Earth Environ. Sci. 2021, 736, 012052. [Google Scholar] [CrossRef]
- Pascoal, A.; Barros-Velázquez, J.; Cepeda, A.; Gallardo, J.M.; Calo-Mata, P. Survey of the authenticity of prawn and shrimp species in commercial food products by PCR-RFLP analysis of a 16S rRNA/tRNAVal mitochondrial region. Food Chem. 2008, 109, 638–646. [Google Scholar] [CrossRef]
- Kim, K.H.; Kang, T.S. Development of advanced PCR-based methods for accurate identification and authentication of commercial shrimp products. Food Control 2024, 159, 110288. [Google Scholar] [CrossRef]
- GB/T43650-2024; National Standard of the People’s Republic of China-Technical Procedures for DNA Species Identification of Wild Animals and Their Products. National Standardization Administration of China: Beijing, China, 2024; pp. 1–20. Available online: https://openstd.samr.gov.cn/bzgk/gb/newGbInfo?hcno=9FCDBFFF10E9ADA1C9685FFB6E552F68 (accessed on 23 September 2024).
- Handy, S.M.; Deeds, J.R.; Ivanova, N.V.; Hebert, P.D.N.; Hanner, R.H.; Ormos, A.; Weigt, L.A.; Moore, M.M.; Yancy, H.F. A Single-Laboratory Validated Method for the Generation of DNA Barcodes for the Identification of Fish for Regulatory Compliance. J. Aoac. Int. 2011, 94, 201–210. [Google Scholar] [CrossRef] [PubMed]
- EU. Regulation (EU) No 1379/2013 of the European Parliament and of the Council of 11 December 2013 on the common organisation of the markets in fishery and aquaculture products, amending Council Regulations (EC) No 1184/2006 and (EC) No 1224/2009 and repealing Council Regulation (EC) No 104/2000. Off. J. Eur. Union 2013, 354, 1–21. Available online: http://data.europa.eu/eli/reg/2013/1379/oj (accessed on 15 November 2024).
- Giusti, A.; Malloggi, C.; Lonzi, V.; Forzano, R.; Meneghetti, B.; Solimeo, A.; Tinacci, L.; Armani, A. Metabarcoding for the authentication of complex seafood products: The fish burger case. J. Food Compos. Anal. 2023, 123, 105559. [Google Scholar] [CrossRef]
- Cristescu, M.E. From barcoding single individuals to metabarcoding biological communities: Towards an integrative approach to the study of global biodiversity. Trends Ecol. Evol. 2014, 29, 566–571. [Google Scholar] [CrossRef]
- Piredda, R.; Mottola, A.; Cipriano, G.; Carlucci, R.; Ciccarese, G.; Di Pinto, A. Next Generation Sequencing (NGS) approach applied to species identification in mixed processed seafood products. Food Control 2022, 133, 108590. [Google Scholar] [CrossRef]
- Fonseca, V.G.; Kirse, A.; Giebner, H.; Vause, B.J.; Drago, T.; Power, D.M.; Peck, L.S.; Clark, M.S. Metabarcoding the Antarctic Peninsula biodiversity using a multi-gene approach. ISME Commun. 2022, 2, 37. [Google Scholar] [CrossRef]
- Marquina, D.; Andersson, A.F.; Ronquist, F. New mitochondrial primers for metabarcoding of insects, designed and evaluated using in silico methods. Mol. Ecol. Resour. 2019, 19, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.K.; Chain, F.J.J.; Abbott, C.L.; Cristescu, M.E. Metabarcoding using multiplexed markers increases species detection in complex zooplankton communities. Evol. Appl. 2018, 11, 1901–1914. [Google Scholar] [CrossRef]
- Lorusso, L.; Shum, P.; Piredda, R.; Mottola, A.; Maiello, G.; Cartledge, E.L.; Neave, E.F.; Di Pinto, A.; Mariani, S. Mismanagement and poor transparency in the European processed seafood supply revealed by DNA metabarcoding. Food Res. Int. 2024, 194, 114901. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.L.; Kelly, R.P.; Lowell, N.C.; Port, J.A. Indexed PCR Primers Induce Template-Specific Bias in Large-Scale DNA Sequencing Studies. PLoS ONE 2016, 11, e0148698. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, X.; Liang, D.; Wang, Q.; Zhang, L.; Zhang, P. VertU: Universal multilocus primer sets for eDNA metabarcoding of vertebrate diversity, evaluated by both artificial and natural cases. Front. Ecol. Evol. 2023, 11, 1164206. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Detcharoen, M.; Khrueakaew, P.; Sukkapat, P.; Benjakul, S.; Saetang, J. Metabarcoding for authentication of fish species in surimi-based products by Nanopore sequencing. Food Biosci. 2024, 61, 104628. [Google Scholar] [CrossRef]
- Park, J.W.; Nozaki, H.; Suzuki, T.; Beliveau, J.-L. Historical review of surimi technology and market developments. Surimi Surimi Seaf. 2013, 1, 1. [Google Scholar]
- Tian, C.; Luan, W.; You, D.; Su, M.; Jin, X. Seafood availability and geographical distance: Evidence from Chinese seafood restaurants. Ocean. Coast. Manag. 2022, 225, 106219. [Google Scholar] [CrossRef]
- GB7718-2011; Chinese National Standard for Food Safety—General Standard for the Labeling of Prepackaged Foods. National Standardization Administration of China: Beijing, China, 2011; pp. 1–9, English Version GAIN Report Number CH 13001. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/2021-02/gb-7718-2011-part-i.pdf (accessed on 7 October 2024).
- Steenhuis, I.H.M.; Waterlander, W.E.; de Mul, A. Consumer food choices: The role of price and pricing strategies. Public Health Nutr. 2011, 14, 2220–2226. [Google Scholar] [CrossRef]
- Ho, J.K.I.; Puniamoorthy, J.; Srivathsan, A.; Meier, R. MinION sequencing of seafood in Singapore reveals creatively labelled flatfishes, confused roe, pig DNA in squid balls, and phantom crustaceans. Food Control 2020, 112, 107144. [Google Scholar] [CrossRef]
- Asmild, M.; Hukom, V.; Nielsen, R.; Nielsen, M. Is economies of scale driving the development in shrimp farming from Penaeus monodon to Litopenaeus vannamei? The case of Indonesia. Aquaculture 2024, 579, 740178. [Google Scholar] [CrossRef]
- Hannan, M.A.; Habib, K.A.; Shahabuddin, A.M.; Haque, M.A.; Munir, M.B. General Discussion. In Post-Harvest Processing, Packaging and Inspection of Frozen Shrimp: A Practical Guide; Hannan, M.A., Habib, K.A., Shahabuddin, A.M., Haque, M.A., Bodrul Munir, M., Eds.; Springer Nature: Singapore, 2022; pp. 1–11. [Google Scholar]
- Kroetz, K.; Luque, G.M.; Gephart, J.A.; Jardine, S.L.; Lee, P.; Chicojay Moore, K.; Cole, C.; Steinkruger, A.; Donlan, C.J. Consequences of seafood mislabeling for marine populations and fisheries management. Proc. Natl. Acad. Sci. USA 2020, 117, 30318–30323. [Google Scholar] [CrossRef] [PubMed]
- Günther, B.; Raupach, M.J.; Knebelsberger, T. Full-length and mini-length DNA barcoding for the identification of seafood commercially traded in Germany. Food Control 2017, 73, 922–929. [Google Scholar] [CrossRef]
- Lee, H.-T.; Liao, C.-H.; Hsu, T.-H. DNA metabarcoding unveils the hidden species composition in fish surimi: Implications for the management of unlabeled and mixed seafood products. Heliyon 2024, 10, e36287. [Google Scholar] [CrossRef] [PubMed]
- Al-Kahtani, H.A.; Ismail, E.A.; Asif Ahmed, M. Pork detection in binary meat mixtures and some commercial food products using conventional and real-time PCR techniques. Food Chem. 2017, 219, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Mottola, A.; Piredda, R.; Lorusso, L.; Armani, A.; Di Pinto, A. Preliminary study on species authentication in poultry meat products by next-generation sequencing. Food Control 2023, 145, 109459. [Google Scholar] [CrossRef]
- Zhang, H.; Xiong, S.; Yu, X.; An, Y. Fishy odorants in pre-processed fish fillet and surimi products made from freshwater fish: Formation mechanism and control methods. Trends Food Sci. Technol. 2023, 142, 104212. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Sinthusamran, S.; Kishimura, H. Gelatin from clown featherback skin: Extraction conditions. LWT-Food Sci. Technol. 2016, 66, 186–192. [Google Scholar] [CrossRef]
- Vaishali; Mandal, A.; Holeyappa, S.A.; Khairnar, S.O.; Barik, S.; Tyagi, A.; Surasani, V.K.R. Growth performance, health status and flesh quality of striped catfish (Pangasianodon hypophthalmus) reared in variable stocking densities in biofloc system. Aquaculture 2024, 590, 741047. [Google Scholar] [CrossRef]
- Haslawati, B.; Amatul-Samahah, M.A.; Rizman-Idid, M.; Muniandy, S. DNA Barcoding and Phylogenetics Relationship of Pangasiid Catfishes in Peninsular Malaysia Revealed the Impacts of Aquaculture on the Native Species Conservation. Hydrobiology 2023, 2, 431–445. [Google Scholar] [CrossRef]
- Torsabo, D.; Ishak, S.D.; Noordin, N.M.; Waiho, K.; Koh, I.C.C.; Yazed, M.A.; Abol-Munafi, A.B. Optimizing reproductive performance in pangasius catfish broodstock: A review of dietary and molecular strategies. Vet. Anim. Sci. 2024, 25, 100375. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Yang, H.; Li, X.; Cao, H.; Zhang, N.; Yan, B.; Hu, B.; Huang, J.; Zhao, J.; Zhang, H.; et al. Green and sustainable microwave processing of surimi seafood: A review of protein component interactions, mechanisms, and industrial applications. Trends Food Sci. Technol. 2024, 143, 104266. [Google Scholar] [CrossRef]
- Luo, H.; Sheng, Z.; Guo, C.; Jia, R.; Yang, W. Quality attributes enhancement of ready-to-eat hairtail fish balls by high-pressure processing. LWT-Food Sci. Technol. 2021, 147, 111658. [Google Scholar] [CrossRef]
- Choi, Y.M.; Choe, J.H.; Cho, D.K.; Kim, B.C. Practical use of surimi-like material made from porcine longissimus dorsi muscle for the production of low-fat pork patties. Meat Sci. 2012, 90, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Kirsten, H.; Seib-Pfeifer, L.-E.; Lüth, C.A.; Rosenfeld, D.L. Validation and application of a German version of the Dietarian Identity Questionnaire: Revealing differences between omnivores, vegetarians, and vegans. Food Qual. Prefer. 2020, 86, 103988. [Google Scholar] [CrossRef]
- Pillai, N.K. A miscellaneous collection of copepods parasitic on South Indian fishes. J. Mar. Biol. Assoc. India 1964, 6, 61–83. [Google Scholar]
- Pedrosa, M.; Boyano-Martínez, T.; García-Ara, C.; Quirce, S. Shellfish Allergy: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2015, 49, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Gil, F.; Beroiz, B.; Ballesteros, I.; Horreo, J.L. Can consumers avoid mislabelling? Genetic species identification provides recommendations for shrimp/prawn products. J. Sci. Food Agr. 2024, 104, 9486–9493. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Gu, X.; Zeng, Q.; Mao, Z.; Chen, H.; Yang, H. Risk screening of non-native freshwater fishes in Yunnan Province, China. Manag. Biol. Invasions 2024, 15, 73–90. [Google Scholar] [CrossRef]
- Karim, A.; Mudassar Shahzad, M.; Kamal, K.; Khwaja, S.; Ijaz, A.; Imtiaz, S. Efficacy of clove oil and rosewood oil as anesthetics on goldfish (Carassius auratus). Egypt. J. Aquat. Res. 2024, 50, 408–413. [Google Scholar] [CrossRef]
- Gilman, E.L. Bycatch governance and best practice mitigation technology in global tuna fisheries. Mar. Policy 2011, 35, 590–609. [Google Scholar] [CrossRef]
- Jacquet, J.; Hocevar, J.; Lai, S.; Majluf, P.; Pelletier, N.; Pitcher, T.; Sala, E.; Sumaila, R.; Pauly, D. Conserving wild fish in a sea of market-based efforts. Oryx 2010, 44, 45–56. [Google Scholar] [CrossRef]
- FAO. FAO Fish Stat J Database: 2019 Dataset. 2019. Available online: https://www.fao.org/fishery/en/statistics/software/fishstatj (accessed on 21 November 2024).
- Ritonga, I.R.; Bureekul, S.; Ubonyaem, T.; Chanrachkij, I.; Sompongchaiyakul, P. Mercury content and consumption risk of 8 species threadfin bream (Nemipterus spp.) caught along the Gulf of Thailand. Mar. Pollut. Bull. 2022, 175, 113363. [Google Scholar] [CrossRef] [PubMed]


| Sample | Ingredients | 16S rRNA | Other Not Declared (16S rRNA) | Declared Shrimp Species | 16S rRNA | Other Not Declared (16S rRNA) | Mislabeling |
|---|---|---|---|---|---|---|---|
| SSP-01 | Shrimp (≥95%) | Yes (45.41%) | No | ||||
| Fish | Yes (44.32%) | ||||||
| Egg white/chicken | Yes (6.16%) | ||||||
| SSP-02 | Shrimp (≥65%) | Yes (7.81%) | Pork (14.66%) | M. nipponense | No | P. vannamei (7.81%) | Yes |
| Fish | Yes (72.40%) | ||||||
| Egg white/chicken | No * | ||||||
| SSP-03 | Shrimp (≥95%) | Yes (25.63%) | Cephalopod (6.11%) | Yes | |||
| Fish (30.51%) | |||||||
| Pork (30.24%) | |||||||
| SSP-04 | Shrimp (≥80%) | Yes (38.88%) | No | ||||
| Cephalopod (≥10%) | Yes (10.85%) | ||||||
| Pork | Yes (45.20%) | ||||||
| Fish | Yes (1%) | ||||||
| SSP-05 | Shrimp [a] | Yes (17.71%) | Pork (2.77%) | M. nipponense | No | P. vannamei (17.71%) | Yes |
| Fish | Yes (72.97%) | ||||||
| Egg white/chicken | No | ||||||
| SSP-06 | Shrimp (≥95%) | Yes (61.40%) | Fish (25.92%) | P. monodon | Yes (36.93%) | Yes | |
| Egg white/chicken | No | Pork (7.09%) | P. vannamei | Yes (24.47%) | |||
| SSP-07 | Shrimp (≥80%) | Yes (7.24%) | Sea lice (4.01%) | Yes | |||
| Fish | Yes (81.39%) | ||||||
| Egg white/chicken | No | ||||||
| SSP-08 | Shrimp | Yes (1.87%) | Yes | ||||
| Fish | Yes (88.73%) | ||||||
| SSP-09 | Shrimp (shrimp + fish ≥ 95%) | Yes (39.48%) | Yes | ||||
| Fish (shrimp + fish ≥ 95%) | Yes (53.76%) | ||||||
| Chicken | No | ||||||
| SSP-10 | Shrimp [a] | Yes (23.61%) | Yes | ||||
| Fish | Yes (63.35%) | ||||||
| Egg white/chicken | No | ||||||
| SSP-11 | Shrimp (≥95%) | Yes (95.65%) | Fish (1.94%) | P. monodon | Yes (68.10%) | P. vannamei (27.55%) | Yes |
| Egg white/chicken | No | ||||||
| SSP-12 | Shrimp (≥85%) | Yes (4.16%) | Pork (1.76%) | Yes | |||
| Fish | Yes (85.95%) | ||||||
| Egg white/chicken | No * | ||||||
| SSP-13 | Shrimp (≥95%) | Yes (38.94%) | Fish (27.97%) | M. nipponense | No | P. vannamei (38.94%) | Yes |
| Egg white/chicken | No * | ||||||
| Pork | Yes (23.74%) | ||||||
| SSP-14 | Shrimp [a] | Yes (50.29%) | Yes | ||||
| Fish | Yes (43.25%) | ||||||
| Egg white/chicken | No | ||||||
| SSP-15 | Shrimp (≥90%) | Yes (95.34%) | Fish (1.08%) | Yes | |||
| Egg white/chicken | No | ||||||
| SSP-16 | Shrimp (≥95%) | Yes (89.71%) | P. monodon | Yes (14.25%) | P. vannamei (75.46%) | Yes | |
| Egg white/chicken | Yes (1.24%) | ||||||
| Pork | Yes (6.85%) | ||||||
| SSP-17 | Shrimp [a] | Yes (7.76%) | Pork (1.81%) | M. nipponense | No | P. vannamei (7.76%) | Yes |
| Fish | Yes (26.41%) | ||||||
| Cephalopod | Yes (1.91%) | ||||||
| Egg white/chicken | Yes (58.96%) | ||||||
| SSP-18 | Shrimp [a] | Yes (29.14%) | Fish (24.62%) | Yes | |||
| Pork | Yes (38.71%) | ||||||
| Egg white/chicken | Yes (1.83%) | ||||||
| SSP-19 | Shrimp (≥95%) | Yes (25.82%) | Pork (3.18%) | Yes | |||
| Fish | Yes (61.63.%) | ||||||
| Egg white/chicken | Yes (2.63%) | ||||||
| SSP-20 | Shrimp [a] | Yes (32.20%) | Pork (1.55%) | Yes | |||
| Fish | Yes (55.54%) | ||||||
| Egg white/chicken | Yes (2.24%) | ||||||
| SSP-21 | Shrimp (≥95%) | Yes (68.24%) | Pork (8.21%) | Yes | |||
| Fish | Yes (15.40%) | ||||||
| Egg white/chicken | Yes (1.20%) | ||||||
| SSP-22 | Shrimp (≥80%) | Yes (98.57%) | No [b] | ||||
| Egg white/chicken | No * | ||||||
| Fish (≥2%) | No * | ||||||
| SSP-23 | Shrimp (≥95%) | Yes (8.14%) | Fish (1.00%) | M. nipponense | No | P. vannamei (8.14%) | Yes |
| Egg white/chicken | Yes (88.14%) | ||||||
| Pork | No * | ||||||
| SSP-24 | Shrimp (≥95%) | Yes (18.11%) | Fish (26.08%) | M. nipponense | No | P. vannamei (18.11%) | Yes |
| Egg white/chicken | No * | ||||||
| Pork | Yes (47.63%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Giusti, A.; Zhang, J.; Tinacci, L.; Zhao, C.; Ying, X.; Armani, A.; Guidi, A.; Deng, S. A Double-Gene Metabarcoding Approach for the Authentication of Shrimp Surimi-Based Products. Genes 2025, 16, 144. https://doi.org/10.3390/genes16020144
Hu J, Giusti A, Zhang J, Tinacci L, Zhao C, Ying X, Armani A, Guidi A, Deng S. A Double-Gene Metabarcoding Approach for the Authentication of Shrimp Surimi-Based Products. Genes. 2025; 16(2):144. https://doi.org/10.3390/genes16020144
Chicago/Turabian StyleHu, Jiajie, Alice Giusti, Jixiang Zhang, Lara Tinacci, Chenyang Zhao, Xiaoguo Ying, Andrea Armani, Alessandra Guidi, and Shanggui Deng. 2025. "A Double-Gene Metabarcoding Approach for the Authentication of Shrimp Surimi-Based Products" Genes 16, no. 2: 144. https://doi.org/10.3390/genes16020144
APA StyleHu, J., Giusti, A., Zhang, J., Tinacci, L., Zhao, C., Ying, X., Armani, A., Guidi, A., & Deng, S. (2025). A Double-Gene Metabarcoding Approach for the Authentication of Shrimp Surimi-Based Products. Genes, 16(2), 144. https://doi.org/10.3390/genes16020144