

Genome-Wide Identification and Characterization of the Pirin Gene Family in Nicotiana benthamiana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of Pirin Genes in N. benthamiana
2.2. Characterization of NbPirins
2.3. Multiple Sequence Alignments and Phylogenetic Analysis
2.4. Gene Structural Domain, Gene Structure, and Motif Analysis of NbPirin
2.5. Analysis of Chromosomal Location and Duplication
2.6. The Calculation of the Ka/Ks Values
2.7. Cis-Acting Regulatory Elements Analysis
2.8. Tissue-Specific Expression of NbPirin
2.9. Plant Growth, Abiotic Stress, and Biotic Stress Treatment
2.10. RNA Extraction and Gene Expression Analysis by qRT-PCR
3. Results
3.1. Genome-Wide Identification and Characterization of the Pirin Gene Family in N. benthamiana
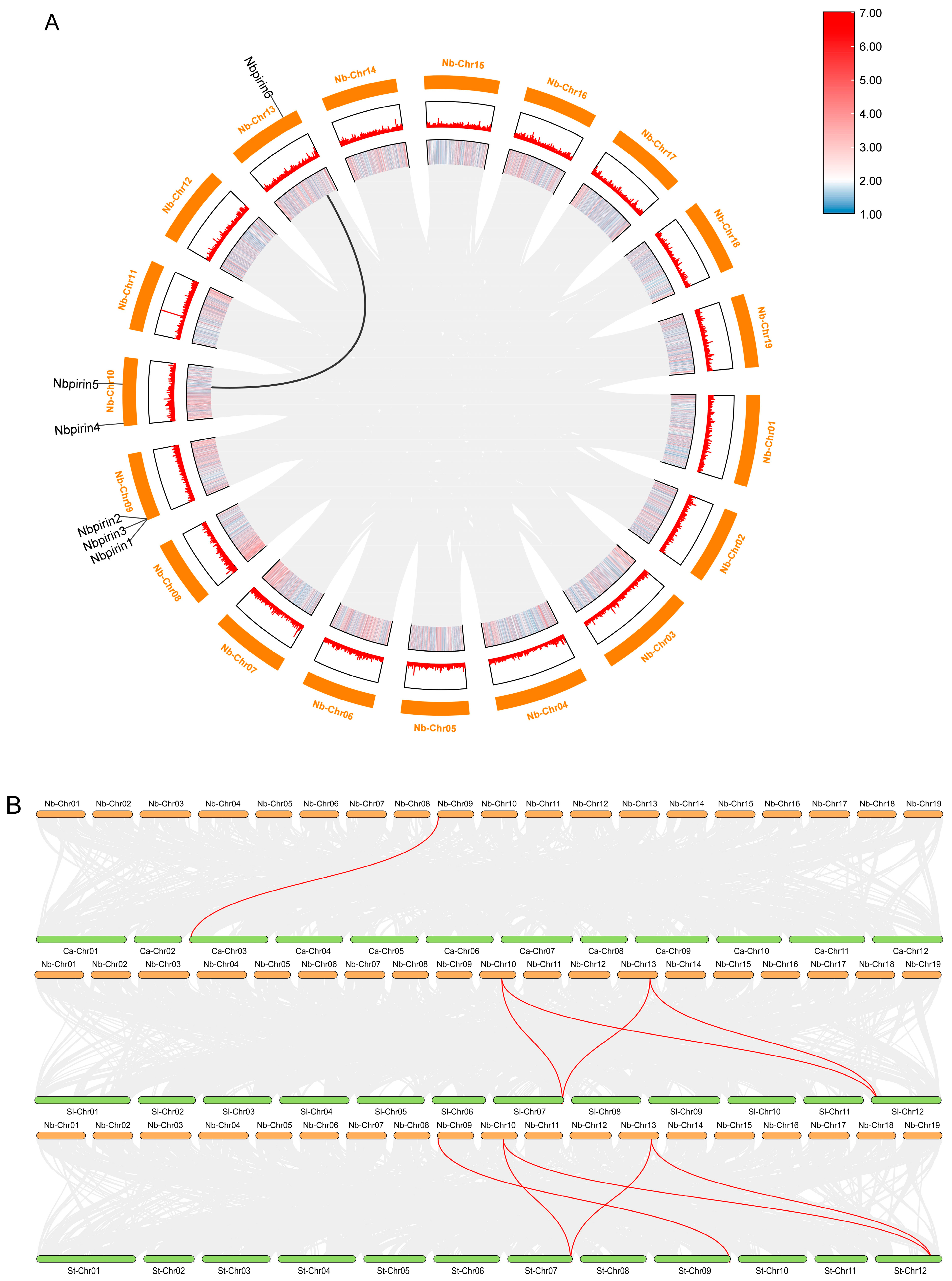
3.2. Determination of the Chromosomal Location and Duplication of NbPirins
3.3. Conserved Motif and Gene Structure Analyses
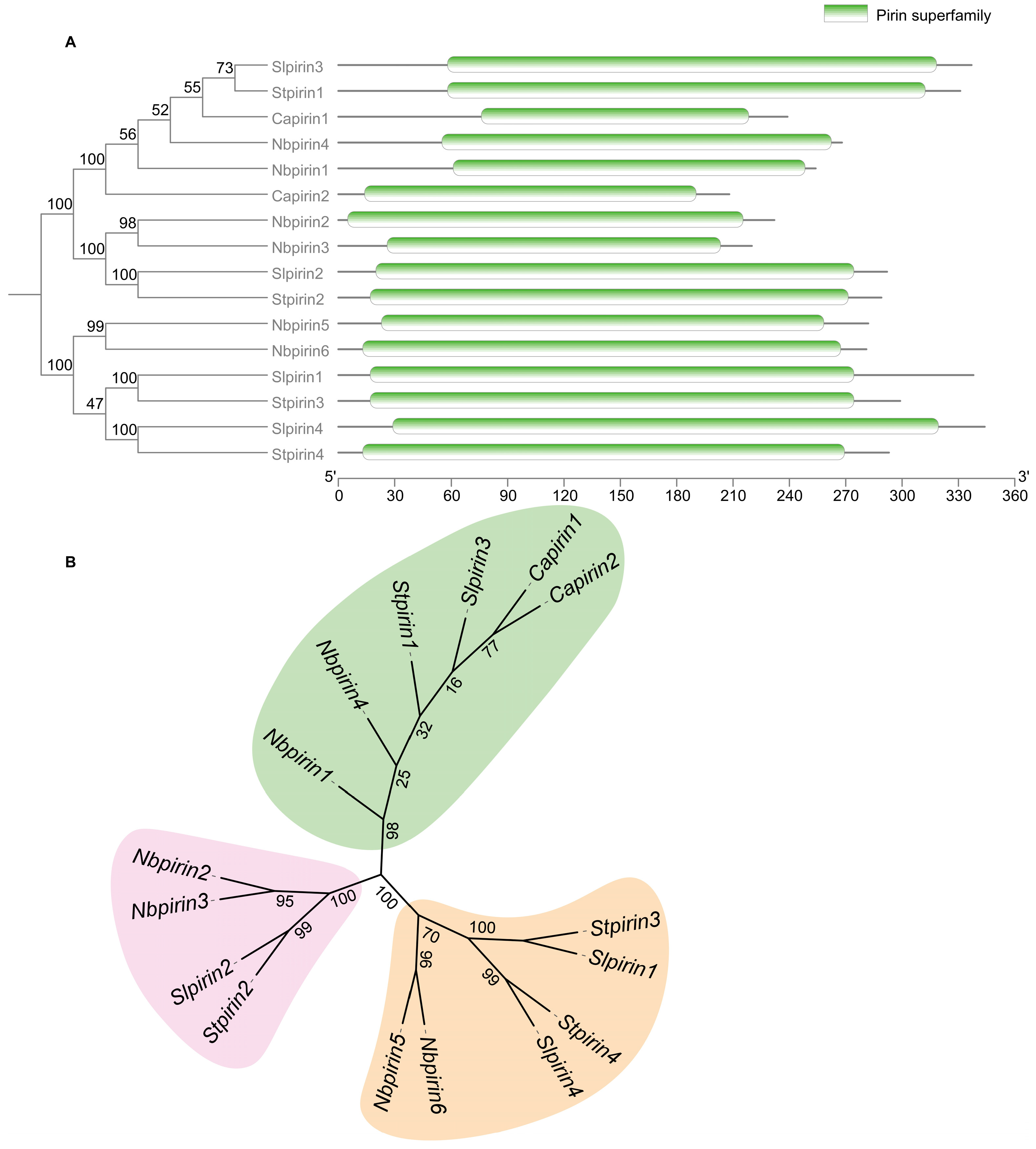
3.4. Evolutionary and Divergence Patterns
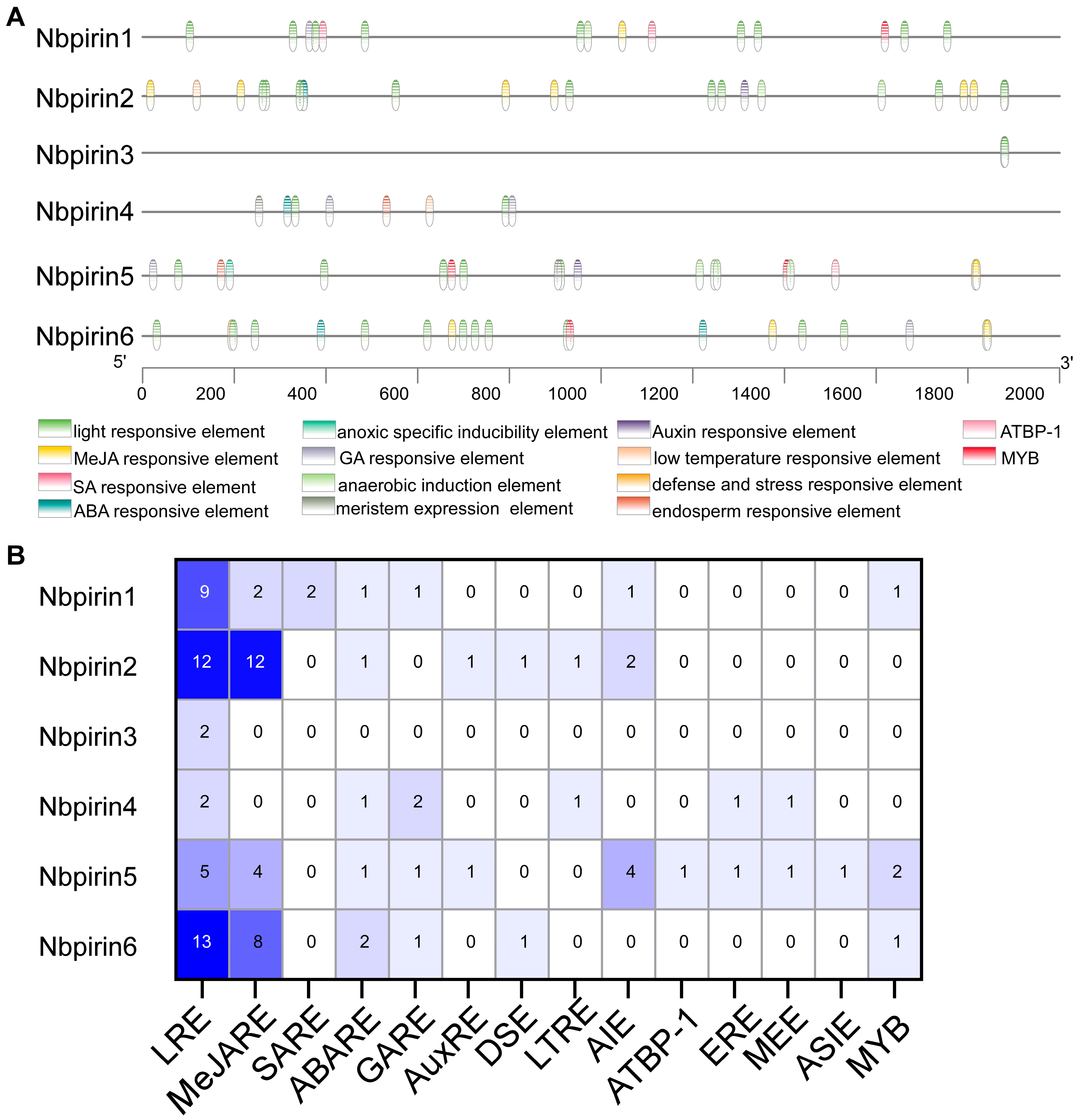
3.5. Prediction of Cis-Acting Regulatory Elements of NbPirins
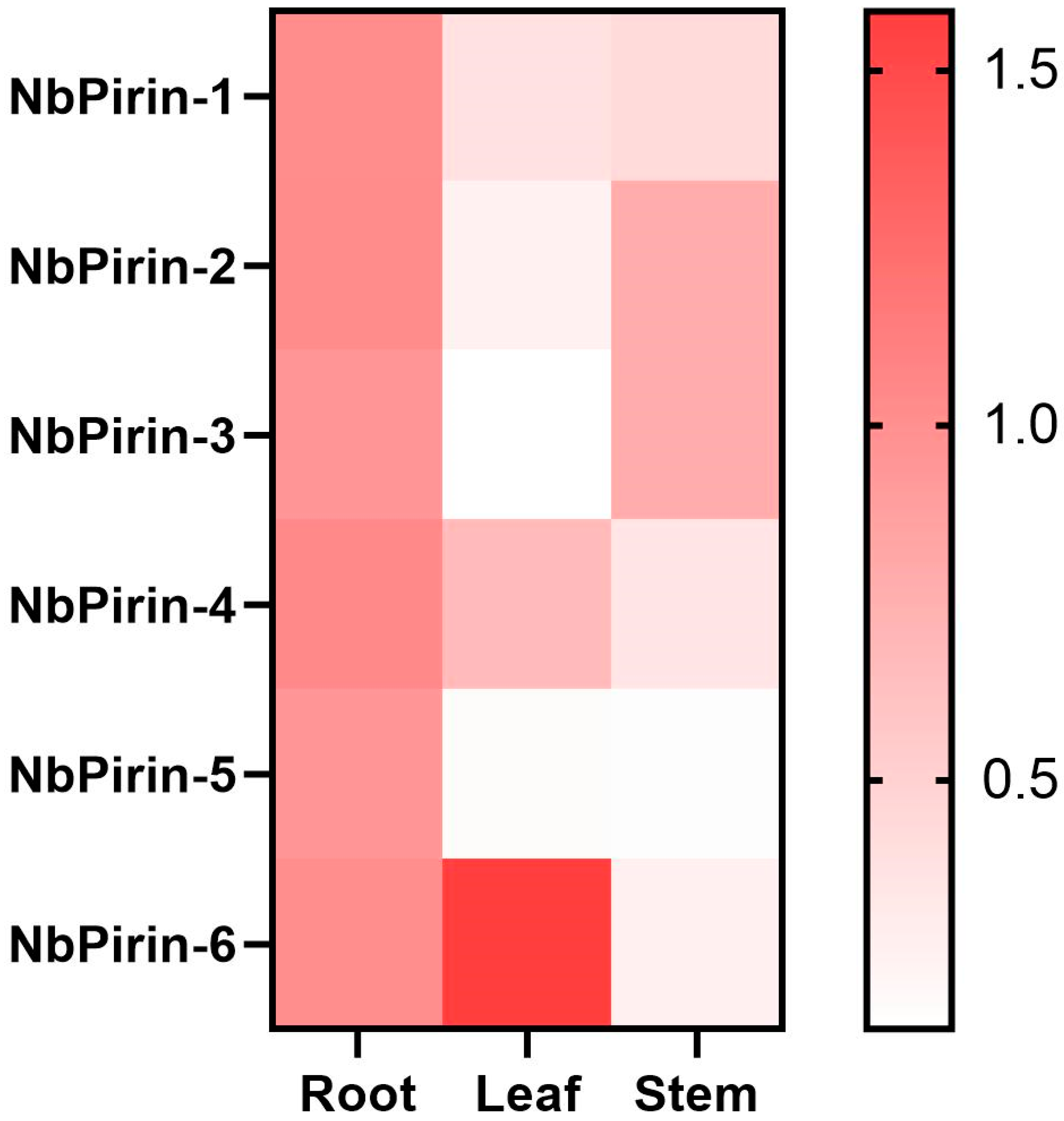
3.6. Tissue-Specific Expression of NbPirins
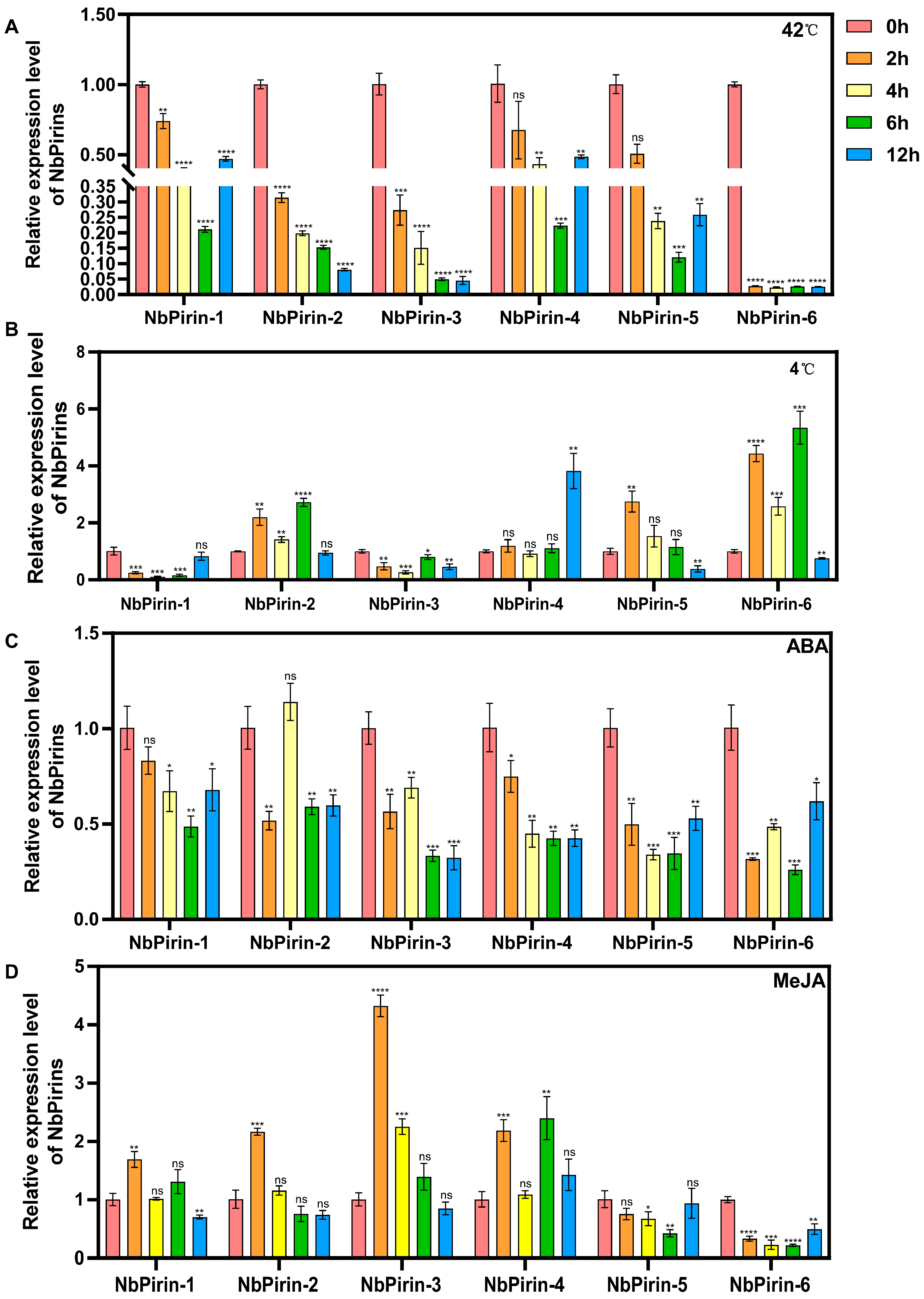
3.7. Expression of NbPirins After Abiotic Stress Treatment
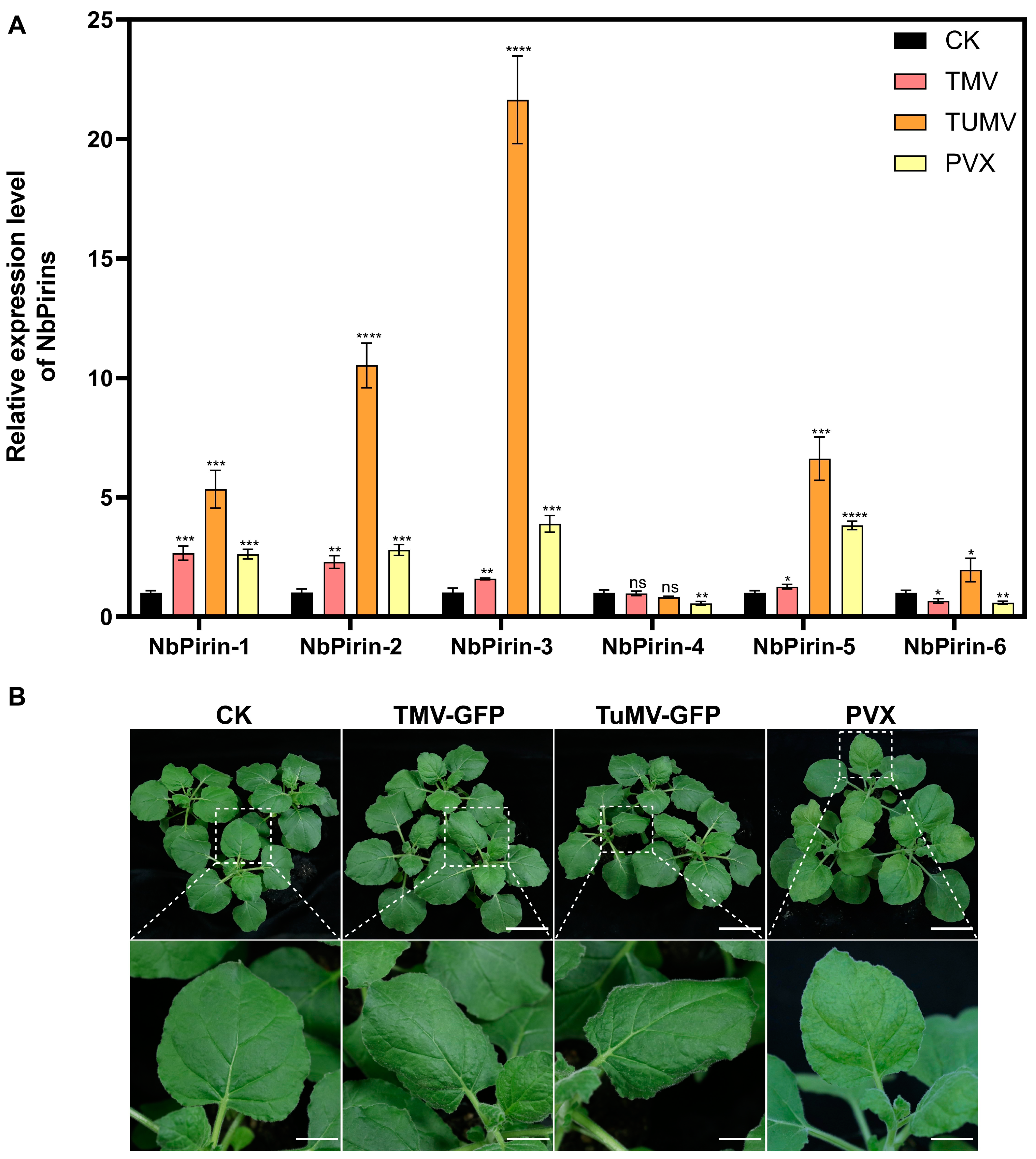
3.8. Expression of NbPirins During TMV, TuMV, or PVX Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dunwell, J.M.; Purvis, A.; Khuri, S. Cupins: The most functionally diverse protein superfamily? Phytochemistry 2004, 65, 7–17. [Google Scholar] [CrossRef]
- Dunwell, J.M. Cupins: A new superfamily of functionally diverse proteins that include germins and plant storage proteins. Biotechnol. Genet. Eng. Rev. 1998, 15, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Bartlam, M.; Zeng, Q.; Miyatake, H.; Hisano, T.; Miki, K.; Wong, L.L.; Gao, G.F.; Rao, Z. Crystal structure of human Pirin: An iron-binding nuclear protein and transcription cofactor. J. Biol. Chem. 2004, 279, 1491–1498. [Google Scholar] [CrossRef] [PubMed]
- Soo, P.-C.; Horng, Y.-T.; Lai, M.-J.; Wei, J.-R.; Hsieh, S.-C.; Chang, Y.-L.; Tsai, Y.-H.; Lai, H.-C. Pirin regulates pyruvate catabolism by interacting with the pyruvate dehydrogenase E1 subunit and modulating pyruvate dehydrogenase activity. J. Bacteriol. 2007, 189, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Wendler, W.M.; Kremmer, E.; Förster, R.; Winnacker, E.L. Identification of Pirin, a novel highly conserved nuclear protein. J. Biol. Chem. 1997, 272, 8482–8489. [Google Scholar] [CrossRef]
- Lapik, Y.R.; Kaufman, L.S. The Arabidopsis cupin domain protein AtPirin1 interacts with the G protein α-subunit GPA1 and regulates seed germination and early seedling development. Plant Cell 2003, 15, 1578–1590. [Google Scholar] [CrossRef]
- Brunetti, S.C.; Arseneault, M.K.M.; Gulick, P.J. Characterization and expression of the Pirin gene family in Triticum aestivum. Genome 2022, 65, 349–362. [Google Scholar] [CrossRef]
- Dechend, R.; Hirano, F.; Lehmann, K.; Heissmeyer, V.; Ansieau, S.; Wulczyn, F.G.; Scheidereit, C.; Leutz, A. The Bcl-3 oncoprotein acts as a bridging factor between NF-kappaB/Rel and nuclear co-regulators. Oncogene 1999, 18, 3316–3323. [Google Scholar] [CrossRef]
- Liu, F.; Rehmani, I.; Esaki, S.; Fu, R.; Chen, L.; de Serrano, V.; Liu, A. Pirin is an iron-dependent redox regulator of NF-κB. Proc. Natl. Acad. Sci. USA 2013, 110, 9722–9727. [Google Scholar] [CrossRef]
- Cao, L.; Zhu, Z.; Qin, H.; Xia, Z.; Xie, J.; Li, X.; Rang, J.; Hu, S.; Sun, Y.; Xia, L. Effects of a Pirin-like protein on strain growth and spinosad biosynthesis in Saccharopolyspora spinosa. Appl. Microbiol. Biotechnol. 2023, 107, 5439–5451. [Google Scholar] [CrossRef]
- Gurmu, D.; Lu, J.; Johnson, K.A.; Nordlund, P.; Holmgren, A.; Erlandsen, H. The crystal structure of the protein YhaK from Escherichia coli reveals a new subclass of redox sensitive enterobacterial bicupins. Proteins 2009, 74, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Muday, G.; Orozco-Nunnelly, D.A.; Muhammad, D.; Mezzich, R.; Lee, B.-S.; Jayathilaka, L.; Kaufman, L.S.; Warpeha, K.M. Pirin1 (PRN1) is a multifunctional protein that regulates quercetin, and impacts specific light and UV responses in the seed-to-seedling transition of Arabidopsis thaliana. PLoS ONE 2014, 9, e93371. [Google Scholar] [CrossRef]
- Zhang, B.; Sztojka, B.; Escamez, S.; Vanholme, R.; Hedenström, M.; Wang, Y.; Turumtay, H.; Gorzsás, A.; Boerjan, W.; Tuominen, H. PIRIN2 suppresses S-type lignin accumulation in a noncell-autonomous manner in Arabidopsis xylem elements. New Phytol. 2020, 225, 1923–1935. [Google Scholar] [CrossRef]
- Bandaranayake, P.C.; Tomilov, A.; Tomilova, N.B.; Ngo, Q.A.; Wickett, N.; dePamphilis, C.W.; Yoder, J.I. The TvPirin gene is necessary for haustorium development in the parasitic plant Triphysaria versicolor. Plant Physiol. 2012, 158, 1046–1053. [Google Scholar] [CrossRef]
- Orzaez, D.; de Jong, A.J.; Woltering, E.J. A tomato homologue of the human protein PIRIN is induced during programmed cell death. Plant Mol. Biol. 2001, 46, 459–468. [Google Scholar] [CrossRef]
- Boddu, J.; Cho, S.; Muehlbauer, G.J. Transcriptome analysis of trichothecene-induced gene expression in barley. Mol. Plant-Microbe Interact. 2007, 20, 1364–1375. [Google Scholar] [CrossRef]
- Nishiuchi, T.; Masuda, D.; Nakashita, H.; Ichimura, K.; Shinozaki, K.; Yoshida, S.; Kimura, M.; Yamaguchi, I.; Yamaguchi, K. Fusarium phytotoxin trichothecenes have an elicitor-like activity in Arabidopsis thaliana, but the activity differed significantly among their molecular species. Mol. Plant-Microbe Interact. 2006, 19, 512–520. [Google Scholar] [CrossRef]
- Zhang, B.; Tremousaygue, D.; Denancé, N.; van Esse, H.P.; Hörger, A.C.; Dabos, P.; Goffner, D.; Thomma, B.P.H.J.; van der Hoorn, R.A.L.; Tuominen, H. PIRIN2 stabilizes cysteine protease XCP2 and increases susceptibility to the vascular pathogen Ralstonia solanacearum in Arabidopsis. Plant J. 2014, 79, 1009–1019. [Google Scholar] [CrossRef]
- Ze, W.; Ting, L.; Liping, D.; Chengpeng, W.; Renda, T.; Sujuan, X.; Xing, C.; Nianjun, T. Lily LlHSFC2 coordinates with HSFAs to balance heat stress response and improve thermotolerance. New Phytol. 2024, 241, 2124–2142. [Google Scholar] [CrossRef]
- Yan, Y.; Zeyang, B.; Qihang, Z.; Wei, W.; Wei, C.; Zhenfeng, Y.; Li, W.; Xuewen, L.; Shifeng, C.; Liyu, S. EjWRKY6 is involved in the ABA-induced carotenoid biosynthesis in loquat fruit during ripening. Foods 2024, 13, 2829. [Google Scholar] [CrossRef]
- Zhengyu, J.; Tao, Z.; Jiajia, C.; Chaoting, L.; Qingqing, Z.; Jin, Q.; Haibo, L.; Jianrong, L.; Xiaofang, Z. Genome-wide identification and expression analysis of the BZR gene family in Zanthoxylum armatum DC and functional analysis of ZaBZR1 in drought tolerance. Planta 2024, 260, 41. [Google Scholar] [CrossRef]
- Chen, X.; Wu, X.; Qiu, S.; Zheng, H.; Lu, Y.; Peng, J.; Wu, G.; Chen, J.; Rao, S.; Yan, F. genome-wide identification and expression profiling of the BZR transcription factor gene family in Nicotiana benthamiana. Int. J. Mol. Sci. 2021, 22, 10379. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiao, M.; Li, Y.; Zhong, Y.; Li, X.; Chen, Z.; Chen, S.; Wang, J.; Vissenberg, K. Penicillium chrysogenum polypeptide extract protects tobacco plants from tobacco mosaic virus infection through modulation of ABA biosynthesis and callose priming. J. Exp. Bot. 2021, 72, 3526–3539. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Tu, A.; Feng, H.; Guo, Y.; Xu, G.; Shi, J.; Chen, J.; Yang, J.; Zhong, K. genome-wide identification and analysis of the ABCF gene family in Triticum aestivum. Int. J. Mol. Sci. 2023, 24, 16478. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Francisco, R.M.; Regalado, A.; Ageorges, A.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.; Chaves, M.M.; et al. ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-Glucosides. Plant Cell 2013, 25, 1840–1854. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Badone, F.C.; Cassani, E.; Landoni, M.; Doria, E.; Panzeri, D.; Lago, C.; Mesiti, F.; Nielsen, E.; Pilu, R. The low phytic acid1-241 (lpa1-241) maize mutation alters the accumulation of anthocyanin pigment in the kernel. Planta 2010, 231, 1189–1199. [Google Scholar] [CrossRef]
- Butler, J.E.F.; Kadonaga, J.T. The RNA polymerase II core promoter: A key component in the regulation of gene expression. Genes Dev. 2002, 16, 2583–2592. [Google Scholar] [CrossRef]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. ABA control of plant macroelement membrane transport systems in response to water deficit and high salinity. New Phytol. 2014, 202, 35–49. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van Loon, L. NPR1: The spider in the web of induced resistance signaling pathways. Curr. Opin. Plant Biol. 2004, 7, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yuan, C.; Feng, S.; Zhong, S.; Li, H.; Zhong, J.; Shen, C.; Liu, J. Genome-wide analysis and characterization of Aux/IAA family genes related to fruit ripening in papaya (Carica papaya L.). BMC Genom. 2017, 18, 351. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Kurilla, A.; Toth, T.; Dorgai, L.; Darula, Z.; Lakatos, T.; Silhavy, D.; Kerenyi, Z.; Dallmann, G. Nectar- and stigma exudate-specific expression of an acidic chitinase could partially protect certain apple cultivars against fire blight disease. Planta 2019, 251, 20. [Google Scholar] [CrossRef]
- Shan, T.; Rong, W.; Xu, H.; Du, L.; Liu, X.; Zhang, Z. The wheat R2R3-MYB transcription factor TaRIM1 participates in resistance response against the pathogen Rhizoctonia cerealis infection through regulating defense genes. Sci. Rep. 2016, 6, 28777. [Google Scholar] [CrossRef]
- Yu, Y.; Guo, D.; Li, G.; Yang, Y.; Zhang, G.; Li, S.; Liang, Z. The grapevine R2R3-type MYB transcription factor VdMYB1 positively regulates defense responses by activating the stilbene synthase gene 2 (VdSTS2). BMC Plant Biol. 2019, 19, 478. [Google Scholar] [CrossRef]
- Islam, W.; Naveed, H.; Zaynab, M.; Huang, Z.; Chen, H.Y.H. Plant defense against virus diseases; growth hormones in highlights. Plant Signal. Behav. 2019, 14, 1596719. [Google Scholar] [CrossRef]
- Wang, X.; Xu, C.; Cai, X.; Wang, Q.; Dai, S. Heat-responsive photosynthetic and signaling pathways in plants: Insight from proteomics. Int. J. Mol. Sci. 2017, 18, 2191. [Google Scholar] [CrossRef]
- Warpeha, K.M.; Upadhyay, S.; Yeh, J.; Adamiak, J.; Hawkins, S.I.; Lapik, Y.R.; Anderson, M.B.; Kaufman, L.S. The GCR1, GPA1, PRN1, NF-Y signal chain mediates both blue light and abscisic acid responses in Arabidopsis. Plant Physiol. 2007, 143, 1590–1600. [Google Scholar] [CrossRef]
- Howard, A.R.; Heppler, M.L.; Ju, H.J.; Krishnamurthy, K.; Payton, M.E.; Verchot-Lubicz, J. Potato virus X TGBp1 induces plasmodesmata gating and moves between cells in several host species whereas CP moves only in N. benthamian leaves. Virology 2004, 328, 185–197. [Google Scholar] [CrossRef]
- Tu, C.W.; Huang, Y.W.; Lee, C.W.; Kuo, S.Y.; Lin, N.S.; Hsu, Y.H.; Hu, C.C. Argonaute 5-mediated antiviral defense and viral counter-defense in Nicotiana benthamiana. Virus Res. 2023, 334, 199179. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Zhang, C.Q.; Dittman, J.D.; Whitham, S.A. Differential requirement of ribosomal protein S6 by plant RNA viruses with different translation initiation strategies. Virology 2009, 390, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Guo, Z.; Zhou, X.; Zhou, L.; Wang, D.; Bo, K.; Zhu, P. Genome-wide identification and characterization of the WRKY gene family in cucurbita maxima. Genes 2023, 14, 2030. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, G.; Shi, J.; Qiao, J.; Liao, P.; Yong, B.; Zhong, K. Genome-Wide Identification and Characterization of the Pirin Gene Family in Nicotiana benthamiana. Genes 2025, 16, 121. https://doi.org/10.3390/genes16020121
Xu G, Shi J, Qiao J, Liao P, Yong B, Zhong K. Genome-Wide Identification and Characterization of the Pirin Gene Family in Nicotiana benthamiana. Genes. 2025; 16(2):121. https://doi.org/10.3390/genes16020121
Chicago/Turabian StyleXu, Gecheng, Jingjing Shi, Jiliang Qiao, Pingan Liao, Bin Yong, and Kaili Zhong. 2025. "Genome-Wide Identification and Characterization of the Pirin Gene Family in Nicotiana benthamiana" Genes 16, no. 2: 121. https://doi.org/10.3390/genes16020121
APA StyleXu, G., Shi, J., Qiao, J., Liao, P., Yong, B., & Zhong, K. (2025). Genome-Wide Identification and Characterization of the Pirin Gene Family in Nicotiana benthamiana. Genes, 16(2), 121. https://doi.org/10.3390/genes16020121