Abstract
Background/Objectives: The eastern periphery of the Slavic expansion (the Volga-Oka region) is the most promising region for reconstructing interactions between Slavic and pre-Slavic populations of the East European Plain. Unlike most pre-Slavic tribes, its autochthonous population practiced inhumation instead of cremation, leaving us with some ancient DNA for analysis. Methods: The region’s modern and ancient Y-chromosome gene pools are dominated by the haplogroup R1a: its frequency reaches 56% in Ryazan Russians (n = 302) and 44% in the Finnic peoples of Mordovia (n = 633). This encouraged us to analyze its Y-SNPs and Y-STRs. Results: Using 2 independent methods of phylogeny analysis, we identified 10 informative Y-STR clusters within R1a, dating back 1600–2900 YBP. The clusters included 48% of modern Ryazan Russians, 40% of Mordovia’s Finnic populations, and ancient DNA samples from the Ryazan-Oka culture (6–7th centuries), Suzdal (12–13th centuries) and Vladimir (13th century). Such a unique combination and pre-Slavic TMRCA indicate that the informative clusters represent pre-Slavic Y lineages. The presence of ancient samples from Vladimir and Suzdal in the clusters suggests that the autochthonous tribes contributed to shaping the urban population of the Vladimir-Suzdal Rus. Some of the informative clusters are associated with the ancient population of the Baltics (2000–4000 YBP). Conclusions: About half of Russian R1a carriers in the Volga-Oka region are descended from a pre-Slavic population, suggesting that the Slavs did not fully replace the autochthonous population but rather mostly culturally assimilated the Meshchyora documented in the Russian chronicles and other local tribes.
1. Introduction
Interactions between Slavic and pre-Slavic populations of the East European Plain have been studied by many disciplines. According to archaeological and toponymic evidence, Slavs had active contacts with the autochthonous tribes while expanding across the East European Plain. But due to methodological constraints, archaeology and linguistics cannot accurately assess the intensity of such contacts: the spread of a language or material culture does not always mark a change in a population’s makeup and can be associated with intertribal trade and linguistic infiltration. Biology provides almost no information about the physical appearance of the local pre-Slavic population or the rate of its change under the pressure of the Slavic expansion because cremation was the primary funeral custom in this region before Christianity. This is why genetic data is so important for reconstructing interactions between Slavic and pre-Slavic populations. The eastern periphery of the Slavic expansion is the most promising region for such reconstruction because, unlike most pre-Slavic populations, its inhabitants practiced in-ground burials, leaving us with some ancient DNA for analysis.
With no natural barriers, the East European Plain was a crossroads of multidirectional migrations; at the outset of the Slavic expansion in the 1st millennium CE, its population was very heterogenous due to the expansion of Baltic-speaking tribes from the West and Finno-Ugric tribes from the East (Appendix A). The arriving Slavs were heterogenous too: they carried traces of past interactions with Celtic and Germanic tribes [1]. In the 4–5th centuries CE, the Early Medieval Pessimum [2] pushed Slavs to migrate to the East European Plain; by the beginning of the 2nd millennium CE, the Slavic expansion had reached its easternmost frontier.
The autochthonous population from the eastern periphery of the Slavic expansion had been long absorbing migrations. The Gorodets culture that evolved from the Finno-Ugric Netted Ware culture and was influenced by the Proto-Balts existed in the Volga-Oka region until the 2nd century CE [3]. Notably, ancestors of Mordovia’s Erzya and Moksha emerged during the final stages of the Gorodets culture and within its geographical boundaries [4,5]. This makes the indigenous population of Mordovia a good model for reconstructing the pre-Slavic gene pool of the eastern periphery of the Slavic expansion. In the 2nd century CE, interactions between the descendants of the Gorodets culture, Balts and Sarmatians shaped the Ryazan-Oka culture (Appendix A) [6,7]. In the middle of the 1st millennium CE, it was heavily influenced by the Baltic tribes [1,4] displaced by early Slavic newcomers [1,4,8,9]. In the 7th century CE, the influx of steppe nomads split the Ryazan-Oka culture. Its descendants dispersed into enclaves that gave rise to the Meshchyora, other early medieval tribes and the gene pools of Mordovia’s Erzya and Moksha [10,11].
By the late 12th century, the Slavs had assimilated most of the autochthonous tribes in the Volga-Oka interfluve and founded the principalities of Ryazan and Vladimir-Suzdal [1,12]. The rate and character of the Slavic expansion varied by region. For example, the culture of the Finnic-speaking Muroma became extinct in the 12th century, but Finnic-speaking Meshchyora settlements in the historic Meshchyora region persisted until the 15–16th centuries because its woodlands and wetlands were taken up by the Slavs less intensively (Appendix A) [13,14].
By and large, the gene pool of the autochthonous population from the eastern periphery of the Slavic expansion was formed by Baltic and Finnic tribes, with some influence from steppe and Cis-Uralic tribes. Unlike most pre-Slavic populations, the pre-Slavic tribes of the Volga-Oka region (Muroma, Meshchyora, Proto-Mordvins) practiced inhumation in the 1st millennium CE, providing us with ancient DNA for analysis. This makes the Volga-Oka region the most promising model for studying interactions between Slavic and pre-Slavic populations of the East European Plain.
Previously, our team participated in an interdisciplinary study of linguistic data and autosomal and Y-chromosome gene pools [15] that demonstrated that all Slavic gene pools had a strong pre-Slavic substrate. The dramatic impact of pre-Slavic gene pools on the modern Slavic gene pool is indicated by 2 factors: (1) differences between East, West and South Slavs are determined by the differences between their pre-Slavic substrates; (2) the DNA markers are better correlated with geography than language.
Of all East Slavs, Russians have the most abundant and heterogenous gene pool. The diversity of Russian populations [16,17] is associated not only with the vastness of their habitat but also with the heterogeneity of the pre-Slavic substrate encountered by the Slavs on the East European Plain. It is the contribution of the autochthonous Baltic and Finnic populations that makes the gene pool of the Russian North so unique [18,19]. The pre-Slavic component represented by the Y haplogroups N3a3 and N3a4 constitutes a sizable proportion of the gene pool of Novgorod Russians [20]. The Y-chromosome gene pool of Yaroslavl Russians has a pre-Slavic substrate that might date back to the Merya people documented in historical chronicles [21].
Another study of autosomal gene pools confirms the important contribution of the pre-Slavic population to the genomic landscape of the East European Plain [22]. The ancestral ADMIXTURE components of Mordovia’s autosomal genomes (Erzya, Moksha and Shoksha) occur in most populations of the East European Plain regardless of their linguistic affiliation. The greatest contribution of the pre-Slavic ancestral ADMIXTURE component is observed in Ryazan Russians. This raises hope that Y-chromosome gene pools of indigenous Mordovian and Ryazan populations will provide evidence of interaction between Slavic and pre-Slavic populations.
Unfortunately, information about ancient DNA of the East European Plain is scarce. The stage of Russian ethnogenesis that preceded transition to the centralized Russian State in the 15th century is yet to be explored. The study of 32 samples from Suzdal Opolie dated to the 3rd–18th centuries pointed to the intensive assimilation of the autochthonous population by the Slavs [23]. When modeling the ancient genome of Alexander Nevsky’s son [24], the Shekshovo-9 genome from Suzdal Opolie [23] was interpreted as a result of interaction between Slavic and pre-Slavic components, and the ‘pure’ Slavic genome was modeled from a medieval Vladimir sample [25].
The aim of this study was to reconstruct the history of the gene pool at the eastern frontier of historical Russia by conducting a search for a pre-Slavic substrate in the modern population of the Volga-Oka interfluve. The reconstruction was based on the comparison of the gene pools of Ryazan Russians and Mordovia’s Finnic-speaking ethnicities using phylogenetic analysis, Y-SNP and Y-STR dating.
2. Materials and Methods
Samples (n = 935) were collected from the indigenous population of the Volga-Oka region during field expeditions in 2010–2023. Of them, 633 represented Mordovia (Ardatovsky, Chamzinsky, Ichalkovsky, Insarsky, Krasnoslobodsky, Lukoyanovsky, Ruzaevsky, Tengushevsky, Torbeyevsky districts) and 302 represented the Ryazan region (Kadomsky, Kasimovsky, Mikhaylovsky, Sapozhkovsky, Saraevsky, Shilovsky, Spassky districts). The samples were collected from unrelated men who gave voluntary written informed consent and whose ancestors from 2 previous generations represented the studied population. The samples were anonymized. The study was approved by the Ethics Committee of the Research Centre for Medical Genetics.
The gene pools of all Russian populations representing the Ryazan region were very similar (χ2) and hence analyzed as a single entity. Mordovian gene pools (Erzya, Moksha and Shoksha) differed significantly from each other (χ2) and were analyzed separately. Note that ‘Shoksha’ is an arbitrary name for the Erzyan population from Tengushevsky district with a distinct gene pool, language and culture [22,26].
DNA was extracted on magnetic beads using a Qiasymphony workstation (Qiagen, Hilden, Germany). Y-SNP genotyping included detailed R1a genotyping with Taqman Open Array custom plates on a QuantStudio 12 Flex real-time PCR platform (Thermo Fisher Scientific, Waltham, MA, USA). Samples of 401 R1a-Z280 carriers were analyzed for 37 Y-STR markers by fragment analysis (Supplementary Materials Table S1) using a Nanofor-05 sequencing system (LLC Syntol, Moscow, Russia), commercial Yfiler Plus (Thermo Fisher Scientific, Waltham, MA, USA) and PowerPlex Y23 (Promega Corp., Fitchburg, WI, USA) kits and separate kits for DYS504, DYS525, DYS552, DYS505, DYS537, DYS445, Y-GATA-A10, and GGAAT1B07. Phylogenetic networks were constructed using a median-joining algorithm [27] implemented in Network v.10.2.0.0 and visualized in Network Publisher v.2.1.2.5. The weight and epsilon parameters were set to 10 and 0, respectively. TMRCA (time to the most recent common ancestor) estimates were calculated from Y-STR haplotypes of the modern samples using two independent methods: ASD and the rho-statistic with heuristic parsimony [19,28,29,30]. The average mutation rate for 37 Y-STRs was assumed to be 0.0039 per locus per generation [31,32]; the generation interval was assumed to be 31.5 years [33].
Distribution maps for the haplogroups R1a-Z92 and R1a-CTS1211 were built in GeneGeo v.2.8 [34] using the average weighted interpolation procedure with a search radius of 400 km and a weight function of 2. Composite portraits of Ryazan and Mordovian venous blood donors were created by superimposing and aligning the photos by reference points; the principle of equal contribution was observed for each photo [35,36].
Of 15 male aDNA samples representing the ancient Volga-Oka population, 5 belonged to R1a carriers. The sample aDNA-1 represented the Ryazan-Oka culture (Undrikh, the 6–8th centuries, the burial site of an upper-class warrior) [37,38]; aDNA-2 and aDNA-4 were from Vasilievsky burial site in Suzdal (the 12–13th centuries); aDNA-4 was from an earlier burial ground than aDNA-2. The samples aDNA-3 and aDNA-5 were from a ‘sanitary’ burial ground in the city of Vladimir, associated with the 1238 AD Mongol invasion.
3. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3.1. Genetic Portraits of Modern Population at Eastern Periphery of Slavic Expansion (Y Haplogroups)
The genetic similarity between the gene pools of modern Slavic- and Finnic-speaking populations of the Volga-Oka region is manifest most visibly in the dominance of the major haplogroup R1a (48%, Table 1), making it the focus of this study. R1a, including its most common R1a-CTS1211 branch, prevails in all 3 Mordovia’s populations at 30–55% frequencies. R1a occurs at a higher frequency (56%) in Ryazan Russians, but is represented in this group by the R1a-Z92 branch more often than in Mordovians. R1a-CTS1211 and R1a-Z92 are the dominant R1a branches in all Volga-Oka populations, representing 86% of the R1a carriers.

Table 1.
Frequencies of Y-chromosome haplogroups (%) in the indigenous populations of the Volga-Oka region (Russians from the east of the Ryazan region and Mordovia’s Erzya, Shoksha and Moksha).
Y haplogroup patterns differ significantly not only between the gene pools of Mordovia and the Ryazan region but also between 3 Mordovia’s populations (Table 1). The gene pools of the Erzya, Moksha and Shoksha are so remarkably different that they should be studied separately instead of being grouped together as the ‘Mordvin’. The Erzya are characterized by the prevalence of R1a (55%) and I1 (9%); the Moksha, by high E-M96 (17%) and J2-M172 (15%) frequencies; the Shokshan gene pool is dominated by N3a (45%). High R1a frequencies also occur outside the Volga-Oka region. R1a is a major, dominant haplogroup in East Slavs, constituting almost half (47%) of their gene pool, followed by I2 (14%) and N-M178 (12%). The gene pool of Russians follows the same pattern. The only region with 2 major haplogroups is the Russian North, where R1a occurs at 26% and N-M178 at 30% frequencies. In other Russian populations, R1a (53%) is strikingly more common than N-M178 (14%); other haplogroups occur at 1–7% frequencies.
Likewise, R1a prevails in the ancient population of the eastern periphery of the Slavic expansion: of 15 aDNA samples from the Undrikh burial site, Suzdal and Vladimir (the 6–13th centuries), 5 (33%) represent R1a carriers. The dominant position of R1a in the gene pool of the Vladimir-Suzdal Rus is confirmed by [23]: of 8 aDNA samples, 3 (38%) represented R1a. So, it would be reasonable to first look for the genetic traces of pre-Slavs within R1a, which has prevailed in the region since antiquity.
The distribution map of the most widespread R1a branches (R1a-CTS1211 and R1a-Z92) shows the position of Mordovia’s and Ryazan’s gene pools among the indigenous gene pools of the entire East European Plain (Figure 1; the boundaries of the regions are shown in white). R1a-CTS1211 occurs at high frequences in the Volga-Oka interfluve (Figure 1A), reaching its peak (47%) in the Ryazan Russians of Kasimovsky district (No. 32 on the map). High R1a-CTS1211 frequencies are typical for South Russians, declining eastward to the Cis-Urals and the Russian North. R1a-Z92 (Figure 1B) is less common and has a peak frequency in the west of the East European Plain among Tver, Yaroslavl and Orel Russians. Its frequency predictably declines eastward, reaching zero values in the Cis- Urals. The same “west-to-east” pattern is observed for R1a-Z92 in the Volga-Oka region, where it has the highest frequency in the westernmost Russian populations and the lowest frequency in the easternmost populations of Mordovia.
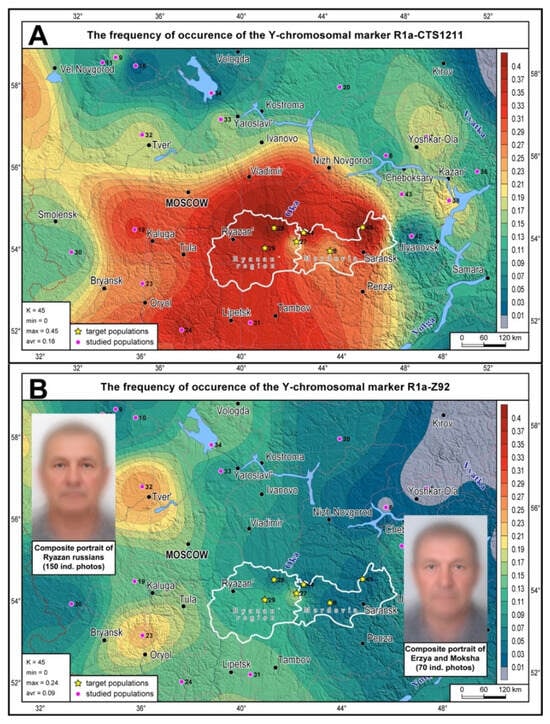
Figure 1.
The distribution of haplogroups R1a-CTS1211 (A) and R1a-Z92 (B) across indigenous populations of the East European Pain. The studied populations are marked with pink dots; yellow stars represent target populations of the Ryazan region and Mordovia (their administrative boundaries are highlighted in white). The color bar legend for haplogroup frequencies is provided in the right panel. The figure also shows composite portraits of (1) Ryazan Russians and (2) Erzyans and Mokshans (B).
3.2. Search for Pre-Slavic Genetic Substrate Using R1a Y-STR Data
It would be reasonable to start searching for a pre-Slavic substrate among those R1a branches that are the most typical for the Volga-Oka region, i.e., R1a-CTS1211 and R1a-Z92. If we find such Y-STR haplotypes that emerged before the arrival of Slavs (TMRCA > 1000 years) and occur simultaneously in Ryazan Russians and Mordovia’s Finnic-speaking peoples, then such Y-STR haplotypes will indicate the presence of a pre-Slavic substrate in the modern Russian gene pool.
Phylogenetic networks of Y-STR haplotypes and their phylogenetic trees for R1a-CTS1211 and R1a-Z92 were constructed on the basis of 401 Y-STR haplotypes representing the modern indigenous population of the Volga-Oka region: 154 Russian samples from 7 districts of the Ryazan region and 247 samples from Mordovia (148 Erzyans, 72 Mokshans, 27 Shokhans). The identified Y-STR clusters corresponded well with Y-SNP variants of the haplogroup R1a (Supplementary Materials Table S1, Figures S1–S5).
Five major mutations Y3910, Y33 (xY1390), Y35 (xY33), CTS1211 (xY35), and Z92 divided the Y-STR haplotypes into 5 more or less equally sized groups, for which 5 ‘search’ phylogenetic networks (Supplementary Materials Figures S1–S5) and 24 ‘search’ phylogenetic trees (Supplementary Materials Figures S6–S15) were constructed. They were searched for such informative clusters of Y-STR haplotypes that simultaneously met two requirements: (1) they included individuals from modern Slavic (Ryazan Russians) and Finnic-speaking (Mordovians) populations; (2) they dated back before the Slavic colonization (before the 10th century). If any of the requirements was not met, the cluster was considered non-informative and was excluded from the analysis.
Ten informative clusters of pre-Slavic Y-STR haplotypes were identified (Supplementary Materials Figures S6–S15). Importantly, these clusters included not only Russians but also individuals from 3 Mordovia’s populations: 46% of Mokshans, 44% of Shokshans, 32% of Erzyans. This suggests that the pre-Slavic substrate is ‘universal’ for the Volga-Oka region. Moreover, 10 informative clusters containing pre-Slavic haplotypes included half (74 of 154!) of all Russian R1a carriers. In other words, the proportion of descendants from the pre-Slavic population among today’s Russian R1a carriers is 48% at the eastern periphery of the Slavic expansion. Since R1a frequency in Ryazan Russians is 56% (Table 1), over one-fourth (27%) of indigenous Russians in the Ryazan region are descended from a pre-Slavic population. Note that this paper analyzes the haplogroup R1a only; the search for a pre-Slavic substrate among other haplogroups deserves a separate publication.
The phylogenetic networks and trees for 3 most widespread R1a subbranches with pre-Slavic Y-STR haplotypes included ancient DNA samples (Figure 2, Figure 3 and Figure 4).
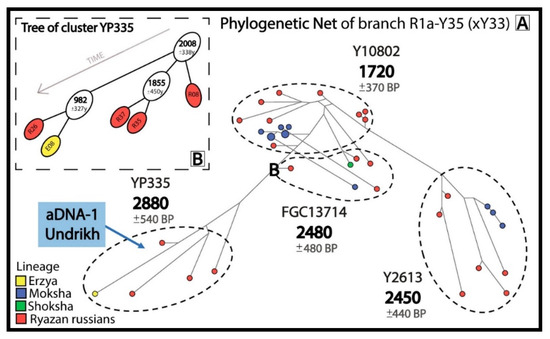
Figure 2.
Informative clusters of probable descendants of the pre-Slavic population in the phylogenetic network for R1a-Y35(xY33) (A) and a phylogenetic tree for cluster R1a-YP335 (B).
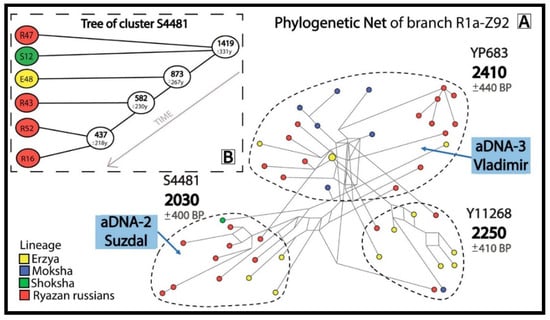
Figure 3.
Informative clusters of probable descendants of the pre-Slavic population in the phylogenetic network for R1a-Z92 (A) and a phylogenetic tree for cluster R1a-S4481 (B).
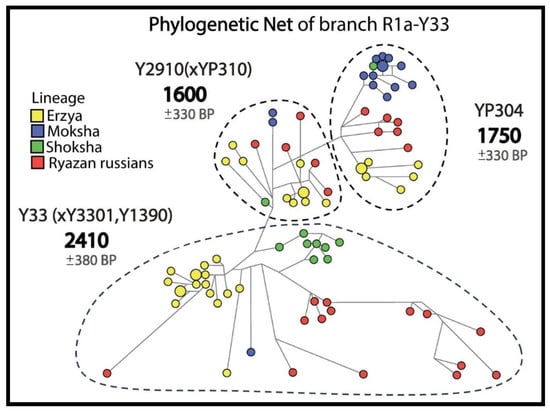
Figure 4.
Informative clusters of probable descendants of the pre-Slavic population in the phylogenetic network for R1a-Y33.
The sample aDNA-1, which dates back before the Slavic expansion (the 6–7th centuries) and was recovered from the Undrikh burial site in the Ryazan region, appears in the R1a-Y35(xY33) network (Figure 2A), confirming the pre-Slavic origin of lineages in this network. The R1a-Y35(xY33) network demonstrates that 23 Russians from 7 Ryazan districts and 13 Erzyans, Mokshans and Shokshans from Mordovia have common pre-Slavic ancestors (1720–2880 YBP). The Erzyan and Russian branches in the R1a-YP335 cluster (Figure 2B) diverged 1000 years ago.
The R1a-Z92 network (Figure 3A) contains aDNA samples from Suzdal (aDNA-2) and Vladimir (aDNA-3), indicating the presence of pre-Slavic Y-chromosome lineages in the urban population of the Vladimir-Suzdal Rus (12–13th centuries). The R1a-Z92 network shows that 26 Russians from 7 Ryazan districts and 21 Erzyans, Mokshans and Shokshans from Mordovia have common pre-Slavic ancestors (2030–2410 YBP).
In the R1a-Y33 network (Figure 4), 25 Russians from 5 Ryazan districts and 59 Erzyans, Mokshans and Shokshans from Mordovia have common pre-Slavic ancestors (1600–2410 YBP).
Thus, all informative clusters dated to 1600–2880 YBP comprise Russians (n = 74) from all the districts of the Ryazan region and individuals from all 3 Finnic-speaking populations of Mordovia (n = 93). Indeed, TMRCA estimates were calculated from the modern Y-STR haplotypes. But the fact that ancient samples (aDNA-1, aDNA-2, aDNA-3) are found within three of the phylogenetically defined clusters validates TMRCA estimates from modern genomes. The oldest aDNA-1 sample from the Undrikh burial (Ryazan-Oka culture, the 6–7th centuries CE) is direct evidence of the pre-Slavic origin of cluster YP335 (Figure 2). The samples aDNA-2 from Suzdal (pre-Mongol Rus, 12–13th centuries CE) and aDNA-3 from Vladimir (the first third of the 13th century) show that the autochthonous pre-Slavic population was incorporated into the gene pool of the urban population of the Vladimir-Suzdal Rus (Figure 3); this finding is supported by archaeological [39] and autosomal genome [23] data. Indeed, a larger ancient DNA sample size is needed to estimate the amount of such incorporation. The samples DNA-4 (Suzdal, the 12–13th centuries) and aDNA-5 (Vladimir, the first third of the 13th century) lie outside the informative clusters.
4. Discussion
Our findings are the first proof of a significant pre-Slavic input into the modern Russian gene pool of the Volga-Oka region. About half (48%) of our R1a Russian carriers represent those R1a lineages that were spread in the region before the Slavic expansion. Considering the high frequency of R1a, at least one-fourth (27%) of its Russian carriers in the region are descended from a pre-Slavic population. This means that the Slavs did not fully replace the autochthonous population but rather culturally assimilated the Meshchyora, Muroma and other pre-Slavic tribes. This is why the pre-Slavic contribution to the modern Russian gene pool is so substantial.
The significant pre-Slavic input into the modern gene pool that formed at the eastern periphery of the Slavic expansion is confirmed by the high frequency of the same R1a lineages in the indigenous population of Mordovia: about 40% of this population share common pre-Slavic ancestors with Ryazan Russians. This confirms that Russians and Mordovians have common pre-Slavic ancestors.
Another important factor is the spread of pre-Slavic R1a branches in the region. First, there are 10 such branches, and this tells against the hypothesis that the high proportion of Russians descended from pre-Slavic populations might be associated with strong genetic drift in the carriers of only one pre-Slavic R1a branch. Second, pre-Slavic R1a branches are uniformly distributed across all districts of the Ryazan region. The same uniformity is observed for all Mordovia’s populations (Erzya, Moksha and Shoksha), confirming that these R1a branches were widespread in the entire pre-Slavic population of the Volga-Oka region.
Evidence from humanities suggests that at the eastern periphery of their expansion Slavs encountered the early medieval Finnic-speaking Meshchyora, Muroma, Mordvin and other tribes, whose names are unknown. Their autochthonism is confirmed by historical continuity: from Netted Ware to Gorodets to Ryazan-Oka to early medieval tribes. Of all Finnic-speaking peoples, only Mordvinic tribes retained their identity. They have given rise to the modern Erzya, Moksha and Shoksha and are a link to the ancient pre-Slavic population.
The strength of our conclusions rests on the careful selection of blood donors: the samples were collected from rural populations that follow the tradition of monoethnic marriages and represented unrelated individuals whose grandfathers belonged, by birth, to the studied population and identified likewise. Evidence from ethnography, archaeology and history points to insignificant miscegenation between Mordovia’s indigenous populations and Russians. Therefore, it is very unlikely that almost half (44%) of the Mordovians are descended from Russians. This conclusion is indirectly supported by pre-Slavic TMRCA, the continuous passage of archaeological cultures and the gradual emergence of Slavic elements in the Meshchyora and Muroma cultures. Importantly, the results of Y-chromosome data analysis reflect the presence of the autochthonous male component in the general population besides the Slavic component from men who engaged in marriages with local women. Previous studies of autosomal genomes and mtDNA [15,22] also suggest a substantial pre-Slavic contribution to the gene pool of East Slavs.
The most important confirmation of the strong pre-Slavic input into the Russian gene pool comes from the dating of 10 informative clusters. Three independent dating methods produced similar results (Table 2); the lowest estimate was 1560 ± 230 YBP and the highest 2880 ± 540 YBP (YFull estimates were slightly higher due to the inclusion of lineages outside of the Volga-Oka region). All dating techniques show that all 10 informative R1a branches existed in the region long before the arrival of Slavs. If Slavs had carried the same R1a lineages before their expansion as the autochthonous population, then, considering that there are 10 informative clusters dated to 1600–2900 YBP, we would have to assume either close contacts between Slavs and the autochthonous population long before (>1000 years) Slavic colonization or a shared ancestral population. However, these assumptions are not supported by archaeological evidence.
Table 2.
Age of 10 informative R1a branches (years BP) estimated by 3 different methods.
The origins of pre-Slavic R1a branches can be reconstructed from archaeological data and aDNA. According to radiocarbon dating, R1a-Z93, typical for the representatives of the Fatyanovo culture (Bronze Age pastoralists, presumably early Indo-European speakers), dominated the Upper and Middle Volga regions 4500–4200 YBP [40,41]. R1a-Z280 emerged 500 years later (3700 YBP) [42] and gave rise to R1a-CTS1211 and R1a-Z92 common for the Volga-Oka region (Table 1 and Table 2). Between 4000 and 2000 YBP, these R1a branches were widespread in the Baltic region: of 13 informative samples, 3 represented R1a-Z92 and 10 represented R1a-CTS1211 [43,44] (Supplementary Materials Table S2). The oldest R1a-CTS1211 (~3900 years) was found in Lithuania [43]. The samples from Lithuania also carried R1a-YP617 (≈2900 YBP) [43], the ancestral branch for clusters Y11268 and YP683 that are informative for the Volga-Oka region (Supplementary Materials Table S2). The sample from Estonia (2500 YBP) [44] lies on the S4481subbranch, and so does the Volga-Oka cluster R1a-Z92 (Supplementary Materials Table S2). This gene geographic pattern is consistent with a potential Baltic trace: some of the identified informative R1a lineages may have shared a common ancestor with Proto-Baltic or other Indo-European tribes living to the west of the Volga-Oka interfluve [4]. Further ancient DNA evidence is needed to distinguish between a direct migration from the Baltic zone and a shared common ancestry from earlier times.
In any case, all R1a lineages dated to 4000–1000 YBP bear no relation to the Slavic expansion but might be associated with Proto-Baltic tribes. All age estimates for the informative Volga-Oka R1a branches fall within this interval (the oldest branch dates back 2880 ± 540 YBP, the youngest 1560 ± 230 YBP) and indicate a significant contribution of the autochthonous pre-Slavic population to the modern Russian gene pool of the Volga-Oka region.
Summing up, this comprehensive study of Y-chromosome gene pools from the eastern periphery of the Slavic expansion has revealed genetic traces of the autochthonous population in the modern Russian population of the Volga-Oka region and in the ancient urban population of Vladimir and Suzdal. Inclusion of the indigenous population in the dataset and the careful choice of Mordovian populations as a model of a pre-Slavic population ensures the reliability of the obtained results. Our findings suggest that the Slavic population of this region was formed through mostly cultural assimilation (Slavicisation) of the local Finnic-speaking tribes (Meshchyora and their early medieval kin tribes) but not through their complete replacement.
5. Conclusions
1. R1a dominates the Y-chromosome gene pools (n = 935) of the modern population of the Volga-Oka region: it is found in 56% of Ryazan Russians and 44% of Mordovia’s indigenous populations. The Y-chromosome gene pools of the Erzya and Moksha are so different from each other that they should be analyzed separately instead of being grouped together as the Mordvin.
2. Using two independent methods for phylogeny reconstruction, we were able to detect 10 haplogroup R1a sublineages of possible pre-Slavic origin descendants in the modern population of the Volga-Oka region dated from 2880 ± 540 YBP to 1560 ± 230 YBP.
3. The frequency of the pre-Slavic substrate lineages is high: about half of the Russian R1a carriers (i.e., one-fourth of the Russian population) are descended from the pre-Slavic population of the region. This suggests that the Russian population of the Volga-Oka interfluve was largely formed by the cultural assimilation rather than by the complete replacement of the Meshchyora and other extinct autochthonous tribes by the Slavs.
4. The analysis of ancient DNA samples from the Ryazan-Oka culture (the 6–7th centuries CE), Suzdal (late 12–early 13th centuries) and Vladimir (the first third of the 13th century) shows that the autochthonous pre-Slavic population was incorporated into the gene pool of the urban population of the Vladimir-Suzdal Rus. A larger ancient DNA sample size is needed to estimate the amount of such incorporation.
5. Many pre-Slavic R1a branches date back over 2500 years and are phylogenetically related to the ancient population of the Baltic region (2000–4000 YBP).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes16101149/s1. Supplementary Table S1: Detailed genotyping data; Supplementary Table S2: Ancient R1a samples; Supplementary Figures S1–S5: Phylogenetic search networks; Supplementary Figures S6–S15: Phylogenetic search trees.
Author Contributions
Analysis of Y-STR and Y-SNP genotyping data, bioinformatic and phylogenetic analysis, construction of phylogenetic networks, manuscript preparation, D.A.; construction of phylogenetic trees, cluster dating, A.S.; creation of composite portraits, manuscript editing, A.P.; analysis of Y-SNP genotyping data, M.V. and I.G.; Y-STR genotyping and its analysis, cartographic analysis G.P.; archaeological fieldwork, preparation of ancient DNA samples D.K., L.C. and A.G.; auxiliary utility software programming for phylogenetic trees D.R.; cartographic analysis, E.P. (Elvira Pocheshkhova); analysis of ancient DNA genomes K.Z. and E.P. (Egor Prokhortchouk); archaeological context, description of the Slavic expansion, N.G.; study design, manuscript preparation E.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 25-28-01594. Ancient DNA analysis was not part of the RSF Project and was carried out under the State Assignment from Russia’s Ministry of Science and Higher Education (Topic ID 124060400040-3, Topic ID 122032300335-0).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of the Research Centre for Medical Genetics (№1 2020-06-29).
Informed Consent Statement
Written informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data and materials are available in Supplementary Materials.
Acknowledgments
We thank the Biobank of North Eurasia that kindly provided DNA samples of Slavic and Finnic-speaking populations from the Ryazan region and Mordovia. We also thank the representatives of Ryazan and Mordovian populations who cooperated with the team in our field work.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| SNP | Single Nucleotide Polymorphism |
| STR | Short Tandem Repeat |
| YBP | Years Before Present |
| TMRCA | Time to the Most Recent Common Ancestor |
| PCR | Polymerase Chain Reaction |
| ASD | Average Squared Distance |
| aDNA | Ancient DNA |
Appendix A. The Archaeological Context of Interactions Between Pre-Slavic and Slavic Populations in the Volga-Oka Region
Archaeological cultures that existed in the early Iron Age on the East European Plain had different origins (Figure A1). This region was inhabited by Baltic tribes in the north-west and west and by the bearers of the Netted Ware (Dyakovo and Gorodets) culture in the east; the latter are thought to be the linguistic ancestors not only of the Baltic Finnic peoples and the Sami but also of the Finnic-speaking tribes of the Volga-Oka interfluve, i.e., the Moksha and Erzya of today’s Mordovia [45,46,47].
The Gorodets culture emerged in the Volga-Oka region in the 6th century BC. It is believed to have been influenced by the Bondarikha culture that extended from the Dnieper to the Upper Oka [48] and was associated with the Balts [3], Iranian-speaking Sarmatians [49] and Finnic tribes [50]. The decline of the Gorodets culture might have occurred at the turn of the Common Era due to migrations from both the West [7] and the East (the Finnic-speaking tribes of the Pyany Bor culture from the South Ural region). The migrants introduced inhumation, which is crucial for reconstructing the gene pool of the pre-Slavic population at the eastern periphery of the Slavic expansion [37,38]. The final stages of the Gorodets culture coincided in time and geographically with the formation of Proto-Mordvinic tribes [4,5].
At the turn of the Common Era, multiple expansions of Iranian Sarmatians into the woodland caused a wave of cultural changes along the border of the forest-steppe zone [51]. This wave was vividly manifest in the adoption of in-ground burial rituals, which were atypical for the cultures associated with the final stages of the early Iron age [52]. Its legacy is reflected in the burials of the Andreevka mound that comprises 4 cultural layers: Gorodets, Pyany Bor, Sarmatian, and ancient Mordvinic [6,52,53]. The effects of that cul-tural shift can be traced to the early stages of the Ryazan-Oka culture that shaped the an-cient population of the Volga-Oka interfluve in the 2nd–8th centuries CE. The arrival of new tribes destroyed the cultural and geographical unity of the Dyakovo-Gorodets cul-tures and separated the Baltic-Finnic and the Oka-Sura tribes [8].
The arrival of the Volga-Kama and Uralic (Pyany Bor and Kara-Abyz) cultures in the Volga-Oka region is associated with the global changes of the first half of the 1st millen-nium CE [8], migrations of Huns across the Eurasian steppe, migrations of Goths to the South, and the fall of the Western Roman Empire. The tribes that settled farther to the north of the Eurasian steppe went on the move too. This led to the influx of a new population associated with the East Baltic tribes to the Oka river basin in the middle of the 1st millennium CE [8] and the emergence of the Moshchiny culture that extended to Suzdal Opolie in the north [54,55]. Bearers of the Slavic Romny culture arrived in the west of the Volga-Oka region in the late 10th and early 11th centuries.
Slavs encountered 3 centers of pre-Slavic cultures in the Oka basin. The first two in the Sura and the Volga-Tyosha regions later gave rise to the Moksha and Erzya of today’s Mordovia [5]. The third was the culture of the Ryazan-Oka burial grounds (the 2nd–8th centuries CE) that evolved from the Gorodets and Pyany Bor cultures [52] and was influ-enced by the Baltic and Sarmatian tribes. The Ryazan-Oka culture was split by nomads in the 7th century. Its descendants dispersed into enclaves [10]; the biggest enclave was on the territory of today’s Kasimovsky district of the Ryazan region. Later, Slavs and the pop-ulations of the Ryazan-Oka culture formed an ethnographic group known as the Russian Meshchyora [10,56].

Figure A1.
Schematic geography of Eastern European archaeological cultures (early Iron Age–Middle Ages). Legends and estimated dates: Early Baltic cultures: Bondarikha culture (11–9th centuries BCE), Moshchiny culture (4–6th centuries CE); Proto-Finnic netted ware cultures: Gorodets culture (6th century BCE–2nd century CE), Dyakovo culture (5th century BCE–5th century CE); Presumably proto-Finnic Ryazan-Oka culture (2nd–6th centuries CE); Slavic Romny culture (9–11th centuries CE); Volga-Kama and Urals cultures of hunters and herders (2nd century BCE–3rd century CE).

Thus, the archaeological context shows that the pre-Slavic population of Eastern Europe and, specifically, the Volga-Oka region was formed by close interactions between the autochthonous Baltic and Finnic tribes. Migrations of steppe populations from the South and the arrival of the Kama and South Ural cultures from the East also contributed to its genetic makeup.
References
- Sedov, V.V. The development of the East European Plain by the Slavs. In Eastern Slavs: Anthropology and Ethnic History, 2nd ed.; Scientific World: Moscow, Russia, 2002; pp. 153–160. (In Russian) [Google Scholar]
- Barford, P.M. Slavs beyond Justinian’s frontiers. Stud. Slavica Balc. Petropolitana 2008, 4, 21–32. [Google Scholar]
- Gimbutas, M. Bronze Age Cultures in Central and Eastern Europe; De Gruyter: Berlin, Germany, 1965; 681p. [Google Scholar]
- Vikhlyaev, V.I. The Origin of Ancient Mordovian Culture; Mordovian Region Publishing: Saransk, Russia, 2000; 132p. (In Russian) [Google Scholar]
- Grishakov, V.V. Pottery of Finno-Ugric Tribes of the Volga Right Bank in the Early Middle Ages; Mari State University Press: Yoshkar-Ola, Russia, 1993; 195p. (In Russian) [Google Scholar]
- Akhmedov, I.R.; Belotserkovskaya, I.V. On the initial date of Ryazan-Oka burial grounds. Proc. State Hist. Mus. 1998, 96, 32–33. (In Russian) [Google Scholar]
- Stavitsky, V.V. Western component in the materials of Andreevsky Kurgan. Bull. Res. Inst. Humanit. Under Gov. Repub. Mordovia 2013, 3, 126–140. (In Russian) [Google Scholar]
- Stavitsky, V.V. At the origins of ancient Mordvin ethnogenesis. Genes. Hist. Res. 2014, 4, 1–13. (In Russian) [Google Scholar]
- Gavrilov, A.P.; Stavitsky, V.V. Nomads at the southern borders of the Oka Finns. Cent. Peripher. 2022, 1, 24–31. (In Russian) [Google Scholar]
- Akhmedov, I.R. The problem of the ‘final’ period of Ryazan-Oka Finns culture: (current state of research). In Archaeology of Eastern Europe in the 1st Millennium AD. Problems and Materials; IA RAS: Moscow, Russia, 2010; pp. 7–34. (In Russian) [Google Scholar]
- Akhmedov, I.R. Ryazan Finns at the end of the 1st—Beginning of the 2nd millennium AD: New data. In Slavs of Eastern Europe on the Eve of the Formation of the Old Russian State: Materials of the International Conference in Honor of I.I. Lyapushkin; Solo: St. Petersburg, Russia, 2012; pp. 244–255. (In Russian) [Google Scholar]
- Golubeva, L.A.; Mogilnikov, V.A.; Sedov, V.V.; Rosenfeldt, R.L. Finno-Ugrians and Balts in the Middle Ages; Archaeology of the USSR from Ancient Times to the Middle Ages in 20 Volumes; Nauka: Moscow, Russia, 1987; 512p. (In Russian) [Google Scholar]
- Ryabinin, E.A. Finno-Ugric Tribes Within Ancient Rus’: On the History of Slavic-Finnic Ethnocultural Connections: Historical and Archaeological Essays; St. Petersburg University Press: St. Petersburg, Russia, 1997; 260p. (In Russian) [Google Scholar]
- Mongait, A.L. The Ryazan Land; USSR Academy of Sciences Publishing: Moscow, Russia, 1961; 400p. (In Russian) [Google Scholar]
- Kushniarevich, A.; Utevska, O.; Chuhryaeva, M.; Agdzhoyan, A.; Dibirova, K.; Uktveryte, I.; Möls, M.; Mulahasanovic, L.; Pshenichnov, A.; Frolova, S.; et al. Genetic heritage of the Balto-Slavic speaking populations: A synthesis of autosomal, mitochondrial and Y-chromosomal data. PLoS ONE 2015, 10, e0135820. [Google Scholar] [CrossRef] [PubMed]
- Balanovsky, O.; Rootsi, S.; Pshenichnov, A.; Kivisild, T.; Churnosov, M.; Evseeva, I.; Pocheshkhova, E.; Boldyreva, M.; Yankovsky, N.; Balanovska, E.; et al. Two sources of the Russian patrilineal heritage in their Eurasian context. Am. J. Hum. Genet. 2008, 82, 236–250. [Google Scholar] [CrossRef]
- Balanovskaya, E.V.; Balanovsky, O.P. The Russian Gene Pool on the Russian Plain; Luch: Moscow, Russia, 2007; 416p. (In Russian) [Google Scholar]
- Okovantsev, V.S.; Ponomarev, G.Y.; Agdzhoyan, A.T.; Agdzhoyan, A.T.; Pylev, V.Y.; Balanovskaya, E.V. Peculiarity of Pomors of Onega Peninsula and Winter Coast in the genetic context of Northern Europe. Bull. Russ. State Med. Univ. 2022, 5, 5–14. (In Russian) [Google Scholar] [CrossRef]
- Balanovskaya, E.V.; Pezhemsky, D.V.; Romanov, A.G.; Baranova, E.E.; Romashkina, M.V.; Agdzhonyan, A.T.; Balagansky, A.G.; Evseeva, I.V.; Villems, E.R.; Balanovsky, O.P. The gene pool of the Russian North: Slavs? Finns? Paleo-Europeans? Mosc. Univ. Anthropol. Bull. 2011, 3, 27–58. (In Russian) [Google Scholar]
- Balanovskaya, E.V.; Agdzhoyan, A.T.; Skhalyakho, R.A.; Balaganskaya, O.A.; Freidin, G.S.; Chernevsky, K.G.; Chernevskii, D.K.; Stepanov, G.D.; Kagazezheva, Z.A.; Zaporozhchenko, V.V.; et al. Gene pool of the Novgorod population: Between the north and the south. Russ. J. Genet. 2017, 53, 1259–1271. (In Russian) [Google Scholar] [CrossRef]
- Chukhryaeva, M.I.; Pavlova, E.S.; Napolskikh, V.V.; Garin, E.V.; Klopov, A.S.; Temnyatkin, S.N.; Zaporozhchenko, V.V.; Romanov, A.G.; Agdzhoyan, A.T.; Utevska, O.M.; et al. Is there a Finno-Ugric component in the gene pool of Russians from Yaroslavl oblast? Evidence from Y-chromosome. Russ. J. Genet. 2017, 53, 388–399. (In Russian) [Google Scholar] [CrossRef]
- Balanovskaya, E.V.; Gorin, I.O.; Ponomarev, G.Y.; Pylev, V.Y.; Petrushenko, V.S.; Markina, N.V.; Mamaeva, A.D.; Agdzhoyan, A.T.; Larin, A.K. Footprints of interaction among Finnic-speaking, Slavic, and Turkic-speaking populations in modern gene pool and their reflection in pharmacogenetics. Bull. Russ. State Med. Univ. 2022, 2, 18–27. (In Russian) [Google Scholar] [CrossRef]
- Peltola, S.; Majander, K.; Makarov, N.; Dobrovolskaya, M.; Nordqvist, K.; Salmela, E.; Onkamo, P. Genetic admixture and language shift in the medieval Volga-Oka interfluve. Curr. Biol. 2023, 33, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Zhur, K.V.; Sharko, F.S.; Sedov, V.V.; Dobrovolskaya, M.V.; Volkov, V.G.; Maximov, N.G.; Seslavine, A.N.; Makarov, N.A.; Prokhortchouk, E.B. The Rurikids: The first experience of reconstructing the genetic portrait of the ruling family of medieval Rus’ based on paleogenomic data. Acta Naturae 2023, 15, 50–65. [Google Scholar] [CrossRef]
- Sikora, M.; Seguin-Orlando, A.; Sousa, V.C.; Albrechtsen, A.; Korneliussen, T.; Ko, A.; Rasmussen, S.; Dupanloup, I.; Nigst, P.R.; Bosch, M.D.; et al. Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers. Science 2017, 358, 659–662. [Google Scholar] [CrossRef]
- Ezhova, V.P. On some ethnographic features in the culture of the Mordovian population of Tengushevsky district of Mordovian ASSR. In Questions of Ethnic History of the Mordovian People: Proceedings of the Mordovian Ethnographic Expedition; Academy of Sciences Publishing: Moscow, Russia, 1960; pp. 210–226. (In Russian) [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Zhivotovsky, L.A.; Underhill, P.A.; Cinnioglu, C.; Kayser, M.; Morar, B.; Kivisild, T.; Scozzari, R.; Cruciani, F.; Destro-Bisol, G.; Spedini, G.; et al. The effective mutation rate at Y chromosome short tandem repeats, with application to human population-divergence time. Am. J. Hum. Genet. 2004, 74, 50–61. [Google Scholar] [CrossRef][Green Version]
- Haak, W.; Balanovsky, O.; Sanchez, J.; Koshel, S.; Zaporozhchenko, V.; Adler, C.J.; Der Sarkissian, C.S.I.; Brandt, G.; Schwarz, C.; Nicklisch, N.; et al. Ancient DNA from European early Neolithic farmers reveals their Near Eastern affinities. PLoS Biol. 2010, 8, e1000536. [Google Scholar] [CrossRef] [PubMed]
- Murka Software [Internet]. Available online: https://phylomurka.sf.net (accessed on 29 May 2025).
- Willuweit, S.; Roewer, L. The new Y chromosome haplotype reference database. Forensic Sci. Int. Genet. 2015, 15, 43–48. [Google Scholar] [CrossRef]
- Ballantyne, K.N.; Goedbloed, M.; Fang, R.; Schaap, O.; Lao, O.; Wollstein, A.; Choi, Y.; van Duijn, K.; Vermeulen, M.; Brauer, S.; et al. Mutability of Y-chromosomal microsatellites: Rates, characteristics, molecular bases, and forensic implications. Am. J. Hum. Genet. 2010, 87, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Fenner, J.N. Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies. Am. J. Phys. Anthropol. 2005, 128, 415–423. [Google Scholar] [CrossRef]
- Koshel, S.M. Geoinformation technologies in genogeography. In Modern Geographical Cartography; Data+: Moscow, Russia, 2012; pp. 158–166. (In Russian) [Google Scholar]
- Belikov, A.V.; Goncharov, I.A.; Goncharova, N.N. The methods of using digitized images in creating composite photograph. Mosc. Univ. Anthropol. Bull. 2014, 1, 74–83. (In Russian) [Google Scholar]
- Perevozchikov, I.V.; Maurer, A.M. Composite photoportraits: History, methods, results. Mosc. Univ. Anthropol. Bull. 2009, 1, 35–44. (In Russian) [Google Scholar]
- Akhmedov, I.R.; Belotserkovskaya, I.V. Ryazan-Oka burial grounds culture. In Eastern Europe in the Mid-1st Millennium AD; Archaeology of Slavs and Their Neighbors; IA RAS: Moscow, Russia, 2007; pp. 133–212. (In Russian) [Google Scholar]
- Gavrilov, A.P. “Silver Age” of the Middle Oka region: On social differentiation and location of Ryazan-Oka archaeological culture at the crossroads of river and land trade routes in the 1st millennium AD. Stratum Plus 2024, 4, 123–134. (In Russian) [Google Scholar] [CrossRef]
- Sedova, M.V. Suzdal in the 10th–15th Centuries; Russian World Publishing: Moscow, Russia, 1997; 320p. (In Russian) [Google Scholar]
- Saag, L.; Vasilyev, S.V.; Varul, L.; Kosorukova, N.V.; Gerasimov, D.V.; Oshibkina, S.V.; Griffith, S.J.; Solnik, A.; Saag, L.; D’aTanasio, E.; et al. Genetic ancestry changes in Stone to Bronze Age transition in the East European plain. Sci. Adv. 2021, 7, eabd6535. [Google Scholar] [CrossRef]
- Engovatova, A.; Mustafin, K.; Alborova, I.; Kanapin, A.; Samsonova, A.; Mednicova, M. New data on the Fatyanovo culture: Analysis of ancient DNA in the burials of the Volosovo-Danilovsky burial ground. In The Footsteps of My Friends Leaving: In Memory of Oleg Sharov; Stratum Publishing House: Chisinau, Moldova, 2022; pp. 115–128. (In Russian) [Google Scholar] [CrossRef]
- Krzewińska, M.; Kılınç, G.M.; Juras, A.; Koptekin, D.; Chyleński, M.; Nikitin, A.G.; Shcherbakov, N.; Shuteleva, I.; Leonova, T.; Kraeva, L.; et al. Ancient genomes suggest the eastern Pontic-Caspian steppe as the source of western Iron Age nomads. Sci. Adv. 2018, 4, eaat4457. [Google Scholar] [CrossRef]
- Mittnik, A.; Wang, C.C.; Pfrengle, S.; Daubaras, M.; Zariņa, G.; Hallgren, F.; Allmäe, R.; Khartanovich, V.; Moiseyev, V.; Tõrv, M.; et al. The genetic prehistory of the Baltic Sea region. Nat. Commun. 2018, 9, 442. [Google Scholar] [CrossRef]
- Saag, L.; Laneman, M.; Varul, L.; Malve, M.; Valk, H.; Razzak, M.; Shirobokov, I.G.; Khartanovich, V.I.; Mikhaylova, E.R.; Kushniarevich, A.; et al. The Arrival of Siberian Ancestry Connecting the Eastern Baltic to Uralic Speakers further East. Curr. Biol. 2019, 29, 1701–1711.e16. [Google Scholar] [CrossRef]
- Mironov, V.G. Gorodets culture: Current issues and prospects for study. In Archaeological Sites of the Middle Oka Region: Collection of Scientific Works; Research and Production Center for the Protection and Use of Historical and Cultural Monuments of the Ryazan Region: Ryazan, Russia, 1995; pp. 68–89. (In Russian) [Google Scholar]
- Napolskikh, V.V. Towards reconstructing the linguistic map of Central European Russia in the Early Iron Age. In Essays on Ethnic History; Kazanskaya Nedvizhimost Publishing House: Kazan, Russia, 2015; pp. 23–64. (In Russian) [Google Scholar]
- Belykh, S.K. Permian-Mari areal language connections: Historical interpretation experience. Idnakar Methods Hist.-Cult. Reconstr. 2009, 3, 5–24. (In Russian) [Google Scholar]
- Buynov, Y.V. Sites of Bondarikha culture in the Middle Oka region. Drevnosti 2016, 14, 74–89. (In Russian) [Google Scholar]
- Smirnov, K.F. Sauromatians; Nauka: Moscow, Russia, 1964; 380p. (In Russian) [Google Scholar]
- Volovik, S.I. Bondarikha Culture (Problems of Origin). PhD Thesis, Institute of Archaeology of the Russian Academy of Sciences, Moscow, Russia, 1996. (In Russian). [Google Scholar]
- Vorontsov, A.M.; Stolyarov, E.V. 1st century war at the forest zone border: Oka-Don watershed. Stratum Plus 2019, 4, 51–74. (In Russian) [Google Scholar]
- Grishakov, V.V.; Zubov, S.E. Andreevsky Kurgan in the System of Early Iron Age Archaeological Cultures of Eastern Europe; Archaeology of Eurasian Steppes; Institute of History of Academy of Sciences of Tatarstan: Kazan, Russia, 2009; p. 173. (In Russian) [Google Scholar]
- Stepanov, P.D. Andreevsky Kurgan (preliminary report). In Proceedings of MNIIYALIE; Mordovian Book Publishing House: Saransk, Russia, 1964; pp. 219–221. (In Russian) [Google Scholar]
- Vorontsov, A.M. Late period of Moshchiny culture existence. Brief Commun. Inst. Archaeol. (KSIA) 2014, 235, 311–324. (In Russian) [Google Scholar]
- Makarov, N.A.; Fedorina, A.N.; Shpolyansky, S.V. Land and city: Medieval settlements in Vladimir-on-Klyazma region. Russ. Archaeol. 2013, 4, 64–80. (In Russian) [Google Scholar]
- Akhmedov, I.R. Ryazan Finns in the lake Meshchera in the third quarter of the 1st millennium AD. In Urbi et Orbi: Collection of Scientific Works in Honor of IG Kusova’s Anniversary; Politekh: Ryazan, Russia, 2020; pp. 23–48+177–178. (In Russian) [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).