
PIWI-Interacting RNAs: A Pivotal Regulator in Neurological Development and Disease

Abstract
1. Introduction
2. The Basic Information of piRNAs
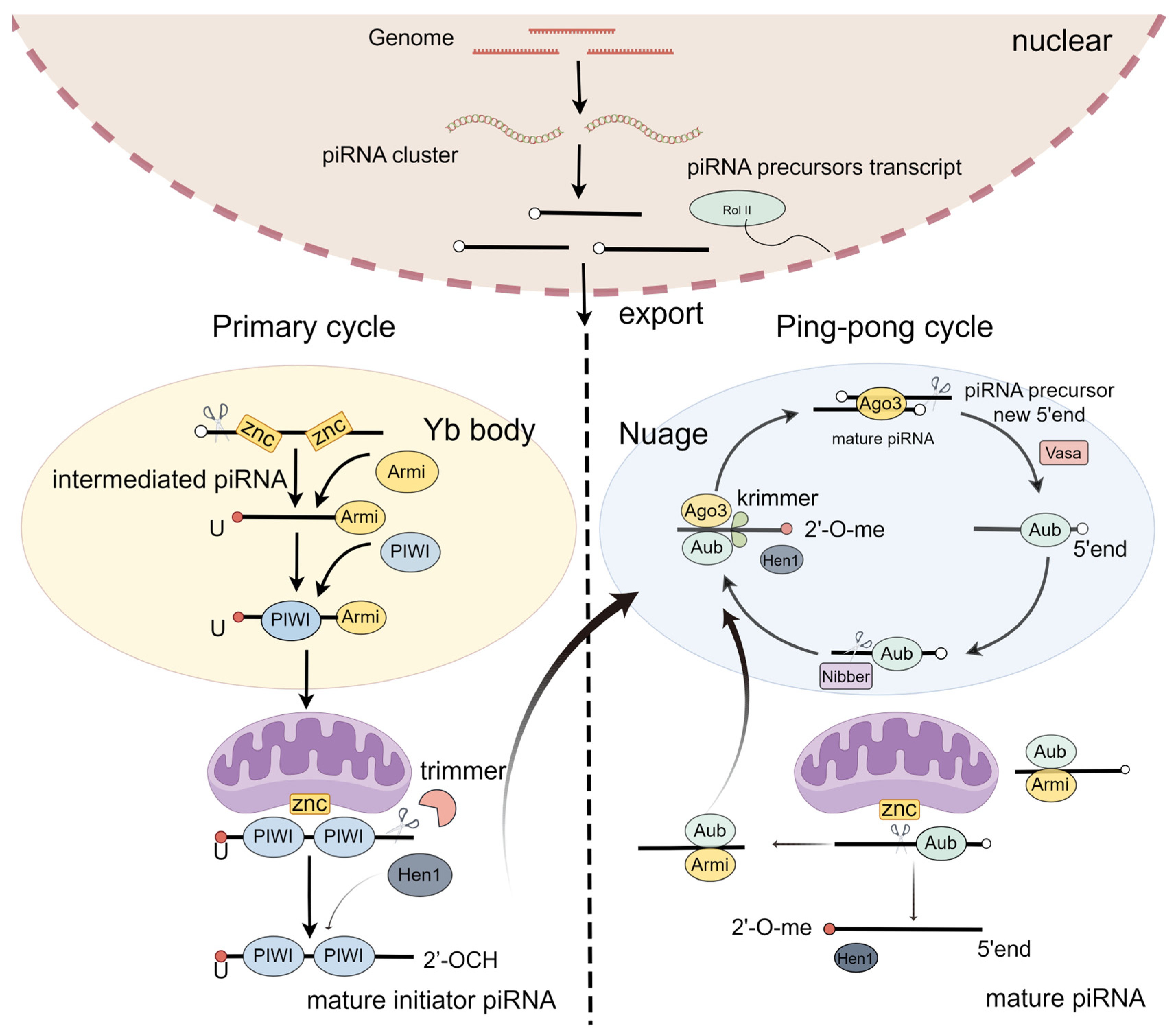
2.1. piRNA Biogenesis
2.2. The Structure, Classification, and Distribution of the piRNA/PIWI Complex
2.2.1. Structure of the piRNA/PIWI Complex
2.2.2. PiRNAs in Different Genomic Areas and Cell Types
2.3. The Databases of piRNAs
3. PiRNAs/PIWI in Neural Differentiation and Neurocognitive Behaviors
3.1. Neuronal Differentiation
3.1.1. PIWI Proteins
3.1.2. PiRNAs
3.2. The Blood–Brain Barrier (BBB)
3.3. Neurocognitive Behaviors (Learning, Memory, Anxiety, and Food Intake)
3.3.1. Learning and Memory
3.3.2. Anxiety
3.3.3. Food Intake
3.3.4. Axon Injury
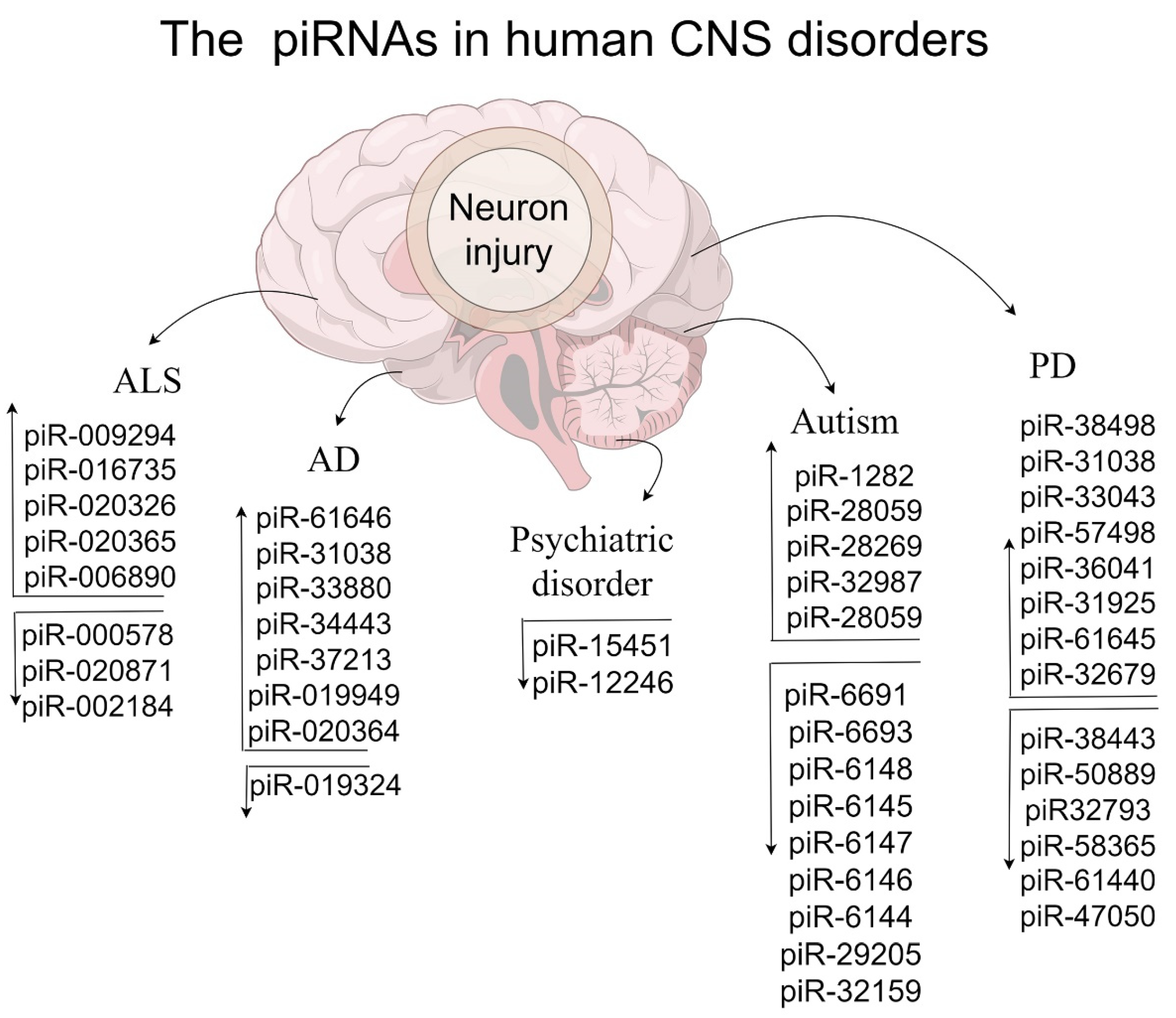
4. piRNAs/PIWI in Neurodevelopmental and Psychiatric Disorders
4.1. Stroke
4.2. Psychiatric Disorders
4.2.1. Schizophrenia (SCZ)
4.2.2. Autism Spectrum Disorder (ASD)
5. PiRNAs/PIWI in Neurodegenerative Disease
5.1. Alzheimer’s Disease (AD)
5.1.1. AD Models
5.1.2. AD Patients
5.2. Parkinson’s Disease (PD)
5.2.1. PD Models
5.2.2. PD Patients
5.3. Amyotrophic Lateral Sclerosis (ALS)
5.3.1. ALS Models
5.3.2. ALS Patients
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, D.; Taylor, D.H.; van Wolfswinkel, J.C. PIWI-mediated control of tissue-specific transposons is essential for somatic cell differentiation. Cell Rep. 2021, 37, 109776. [Google Scholar] [CrossRef] [PubMed]
- Girard, A.; Sachidanandam, R.; Hannon, G.J.; Carmell, M.A. A germline-specific class of small RNAs binds mammalian Piwi proteins. Nature 2006, 442, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Aravin, A.; Gaidatzis, D.; Pfeffer, S.; Lagos-Quintana, M.; Landgraf, P.; Iovino, N.; Morris, P.; Brownstein, M.J.; Kuramochi-Miyagawa, S.; Nakano, T.; et al. A novel class of small RNAs bind to MILI protein in mouse testes. Nature 2006, 442, 203–207. [Google Scholar] [CrossRef]
- Czech, B.; Hannon, G.J. One Loop to Rule Them All: The Ping-Pong Cycle and piRNA-Guided Silencing. Trends Biochem. Sci. 2016, 41, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dou, M.; Song, X.; Dong, Y.; Liu, S.; Liu, H.; Tao, J.; Li, W.; Yin, X.; Xu, W. The emerging role of the piRNA/piwi complex in cancer. Mol. Cancer 2019, 18, 123. [Google Scholar] [CrossRef]
- Jia, D.D.; Jiang, H.; Zhang, Y.F.; Zhang, Y.; Qian, L.L.; Zhang, Y.F. The regulatory function of piRNA/PIWI complex in cancer and other human diseases: The role of DNA methylation. Int. J. Biol. Sci. 2022, 18, 3358–3373. [Google Scholar] [CrossRef] [PubMed]
- Zuo, L.; Wang, Z.; Tan, Y.; Chen, X.; Luo, X. piRNAs and Their Functions in the Brain. Int. J. Hum. Genet. 2016, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Wen, Z.J.; Xu, H.M.; Zhang, Y.; Zhang, Y.F. Exosomal noncoding RNAs in central nervous system diseases: Biological functions and potential clinical applications. Front. Mol. Neurosci. 2022, 15, 1004221. [Google Scholar] [CrossRef]
- Gainetdinov, I.; Vega-Badillo, J.; Cecchini, K.; Bagci, A.; Colpan, C.; De, D.; Bailey, S.; Arif, A.; Wu, P.H.; MacRae, I.J.; et al. Relaxed targeting rules help PIWI proteins silence transposons. Nature 2023, 619, 394–402. [Google Scholar] [CrossRef]
- He, C.; Wang, K.; Gao, Y.; Wang, C.; Li, L.; Liao, Y.; Hu, K.; Liang, M. Roles of Noncoding RNA in Reproduction. Front. Genet. 2021, 12, 777510. [Google Scholar] [CrossRef]
- Saritas, G.; Main, A.M.; Winge, S.B.; Mørup, N.; Almstrup, K. PIWI-interacting RNAs and human testicular function. WIREs Mech. Dis. 2022, 14, e1572. [Google Scholar] [CrossRef] [PubMed]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defence. Nat. Reviews. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Siomi, M.C. The piRNA pathway in Drosophila ovarian germ and somatic cells. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2020, 96, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Onishi, R.; Yamanaka, S.; Siomi, M.C. piRNA- and siRNA-mediated transcriptional repression in Drosophila, mice, and yeast: New insights and biodiversity. EMBO Rep. 2021, 22, e53062. [Google Scholar] [CrossRef] [PubMed]
- Hasuwa, H.; Iwasaki, Y.W.; Au Yeung, W.K.; Ishino, K.; Masuda, H.; Sasaki, H.; Siomi, H. Production of functional oocytes requires maternally expressed PIWI genes and piRNAs in golden hamsters. Nat. Cell Biol. 2021, 23, 1002–1012. [Google Scholar] [CrossRef]
- Loubalova, Z.; Fulka, H.; Horvat, F.; Pasulka, J.; Malik, R.; Hirose, M.; Ogura, A.; Svoboda, P. Formation of spermatogonia and fertile oocytes in golden hamsters requires piRNAs. Nat. Cell Biol. 2021, 23, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Perillo, G.; Shibata, K.; Wu, P.H. piRNAs in sperm function and embryo viability. Reproduction 2023, 165, R91–R102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, F.; Chen, Q.; Li, M.; Lv, X.; Xiao, Y.; Zhang, Z.; Hou, L.; Lai, Y.; Zhang, Y.; et al. The piRNA pathway is essential for generating functional oocytes in golden hamsters. Nat. Cell Biol. 2021, 23, 1013–1022. [Google Scholar] [CrossRef]
- Sohn, E.J.; Jo, Y.R.; Park, H.T. Downregulation MIWI-piRNA regulates the migration of Schwann cells in peripheral nerve injury. Biochem. Biophys. Res. Commun. 2019, 519, 605–612. [Google Scholar] [CrossRef]
- Kim, K.W.; Tang, N.H.; Andrusiak, M.G.; Wu, Z.; Chisholm, A.D.; Jin, Y. A Neuronal piRNA Pathway Inhibits Axon Regeneration in C. elegans. Neuron 2018, 97, 511–519.e6. [Google Scholar] [CrossRef]
- Rajasethupathy, P.; Antonov, I.; Sheridan, R.; Frey, S.; Sander, C.; Tuschl, T.; Kandel, E.R. A role for neuronal piRNAs in the epigenetic control of memory-related synaptic plasticity. Cell 2012, 149, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Leighton, L.J.; Wei, W.; Marshall, P.R.; Ratnu, V.S.; Li, X.; Zajaczkowski, E.L.; Spadaro, P.A.; Khandelwal, N.; Kumar, A.; Bredy, T.W. Disrupting the hippocampal Piwi pathway enhances contextual fear memory in mice. Neurobiol. Learn. Mem. 2019, 161, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Landry, C.D.; Kandel, E.R.; Rajasethupathy, P. New mechanisms in memory storage: piRNAs and epigenetics. Trends Neurosci. 2013, 36, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Pierouli, K.; Papakonstantinou, E.; Papageorgiou, L.; Diakou, I.; Mitsis, T.; Dragoumani, K.; Spandidos, D.A.; Bacopoulou, F.; Chrousos, G.P.; Goulielmos, G.; et al. Role of non-coding RNAs as biomarkers and the application of omics technologies in Alzheimer’s disease (Review). Int. J. Mol. Med. 2023, 51, 5. [Google Scholar] [CrossRef]
- Zhang, T.; Wong, G. Dysregulation of Human Somatic piRNA Expression in Parkinson’s Disease Subtypes and Stages. Int. J. Mol. Sci. 2022, 23, 2469. [Google Scholar] [CrossRef] [PubMed]
- Wakisaka1, K.T.; Ima, a.Y. The dawn of piRNA research in various neuronal disorders. Front. Biosci. 2019, 24, 1440–1451. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Samimi, H.; Gamez, M.; Zare, H.; Frost, B. Pathogenic tau-induced piRNA depletion promotes neuronal death through transposable element dysregulation in neurodegenerative tauopathies. Nat. Neurosci. 2018, 21, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, C.; Zhang, T.; Li, R.; Chen, L.; Leung, K.L.; Lakso, M.; Zhou, Q.; Zhang, H.; Wong, G. PIWI-interacting RNA expression regulates pathogenesis in a Caenorhabditis elegans model of Lewy body disease. Nat. Commun. 2023, 14, 6137. [Google Scholar] [CrossRef]
- Chen, S.; Ben, S.; Xin, J.; Li, S.; Zheng, R.; Wang, H.; Fan, L.; Du, M.; Zhang, Z.; Wang, M. The biogenesis and biological function of PIWI-interacting RNA in cancer. J. Hematol. Oncol. 2021, 14, 93. [Google Scholar] [CrossRef]
- Mohn, F.; Handler, D.; Brennecke, J. Noncoding RNA. piRNA-guided slicing specifies transcripts for Zucchini-dependent, phased piRNA biogenesis. Science 2015, 348, 812–817. [Google Scholar] [CrossRef]
- Wu, Z.; Yu, X.; Zhang, S.; He, Y.; Guo, W. Novel roles of PIWI proteins and PIWI-interacting RNAs in human health and diseases. Cell Commun. Signal. 2023, 21, 343. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, S.; Liu, K. Structural insights into piRNA biogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2022, 1865, 194799. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, J.; Cho, W.C.; Zheng, Y. Argonaute proteins: Structural features, functions and emerging roles. J. Adv. Res. 2020, 24, 317–324. [Google Scholar] [CrossRef]
- Iwasaki, Y.W.; Siomi, M.C.; Siomi, H. PIWI-Interacting RNA: Its Biogenesis and Functions. Annu. Rev. Biochem. 2015, 84, 405–433. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, I.; Colpan, C.; Cecchini, K.; Arif, A.; Jouravleva, K.; Albosta, P.; Vega-Badillo, J.; Lee, Y.; Ozata, D.M.; Zamore, P.D. Terminal modification, sequence, length, and PIWI-protein identity determine piRNA stability. Mol. Cell 2021, 81, 4826–4842.e8. [Google Scholar] [CrossRef]
- Hirakata, S.; Siomi, M.C. Assembly and Function of Gonad-Specific Non-Membranous Organelles in Drosophila piRNA Biogenesis. Noncoding RNA 2019, 5, 52. [Google Scholar] [CrossRef]
- Munafo, M.; Manelli, V.; Falconio, F.A.; Sawle, A.; Kneuss, E.; Eastwood, E.L.; Seah, J.W.E.; Czech, B.; Hannon, G.J. Daedalus and Gasz recruit Armitage to mitochondria, bringing piRNA precursors to the biogenesis machinery. Genes. Dev. 2019, 33, 844–856. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, C.; Liao, S.; Zhu, Z.; Zhang, J.; Tu, X.; Yao, X.; Feng, X.; Guang, S.; Xu, C. Molecular basis for PICS-mediated piRNA biogenesis and cell division. Nat. Commun. 2021, 12, 5595. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhou, T.; Chen, Q. Exploring the expanding universe of small RNAs. Nat. Cell Biol. 2022, 24, 415–423. [Google Scholar] [CrossRef]
- Cerutti, L.; Mian, N.; Bateman, A. Domains in gene silencing and cell differentiation proteins: The novel PAZ domain and redefinition of the Piwi domain. Trends Biochem. Sci. 2000, 25, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, W.; Liu, Y.; Pi, M.; Jiang, Y.; Ainiwaer, A.; Mao, S.; Chen, H.; Ran, Y.; Sun, S.; et al. Emerging roles and potential application of PIWI-interacting RNA in urological tumors. Front. Endocrinol. 2022, 13, 1054216. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sheng, G.; Juranek, S.; Tuschl, T.; Patel, D.J. Structure of the guide-strand-containing argonaute silencing complex. Nature 2008, 456, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, T.; Gao, X.Q.; Chen, X.Z.; Wang, F.; Zhou, L.Y. Emerging functions of piwi-interacting RNAs in diseases. J. Cell Mol. Med. 2021, 25, 4893–4901. [Google Scholar] [CrossRef] [PubMed]
- Copley, K.E.; Shorter, J. Repetitive elements in aging and neurodegeneration. Trends Genet. 2023, 39, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, M.; Kounosu, A.; Murcott, B.; Dayi, M.; Pawluk, R.; Yoshida, A.; Viney, M.; Kikuchi, T.; Hunt, V.L. piRNA-like small RNAs target transposable elements in a Clade IV parasitic nematode. Sci. Rep. 2022, 12, 10156. [Google Scholar] [CrossRef] [PubMed]
- Houwing, S.; Kamminga, L.M.; Berezikov, E.; Cronembold, D.; Girard, A.; van den Elst, H.; Filippov, D.V.; Blaser, H.; Raz, E.; Moens, C.B.; et al. A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish. Cell 2007, 129, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Siomi, M.C.; Miyoshi, T.; Siomi, H. piRNA-mediated silencing in Drosophila germlines. Semin. Cell Dev. Biol. 2010, 21, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Chuma, S.; Nakano, T. piRNA and spermatogenesis in mice. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20110338. [Google Scholar] [CrossRef] [PubMed]
- Ghildiyal, M.; Seitz, H.; Horwich, M.D.; Li, C.; Du, T.; Lee, S.; Xu, J.; Kittler, E.L.; Zapp, M.L.; Weng, Z.; et al. Endogenous siRNAs derived from transposons and mRNAs in Drosophila somatic cells. Science 2008, 320, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Aravin, A.A.; Naumova, N.M.; Tulin, A.V.; Vagin, V.V.; Rozovsky, Y.M.; Gvozdev, V.A. Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr. Biol. 2001, 11, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W. PIWI Proteins and piRNAs in the Nervous System. Mol. Cells 2019, 42, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Anzelon, T.A.; Chowdhury, S.; Hughes, S.M.; Xiao, Y.; Lander, G.C.; MacRae, I.J. Structural basis for piRNA targeting. Nature 2021, 597, 285–289. [Google Scholar] [CrossRef]
- Wang, X.; Ramat, A.; Simonelig, M.; Liu, M.F. Emerging roles and functional mechanisms of PIWI-interacting RNAs. Nat. Rev. Mol. Cell Biol. 2023, 24, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.P.; Young, P.E.; McCorkindale, A.L.; Dang, T.H.; Clancy, J.L.; Humphreys, D.T.; Preiss, T.; Hutvagner, G.; Martin, D.I.; Cropley, J.E.; et al. The human Piwi protein Hiwi2 associates with tRNA-derived piRNAs in somatic cells. Nucleic Acids Res. 2014, 42, 8984–8995. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, P.; Ghosh, S.; Wang, B.; Heyns, M.; Li, D.; Mackey, J.R.; Kovalchuk, O.; Damaraju, S. Genome-wide profiling of transfer RNAs and their role as novel prognostic markers for breast cancer. Sci. Rep. 2016, 6, 32843. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Kawamura, T.; Loher, P.; Morichika, K.; Rigoutsos, I.; Kirino, Y. The biogenesis pathway of tRNA-derived piRNAs in Bombyx germ cells. Nucleic Acids Res. 2017, 45, 9108–9120. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.; Brasset, E.; Parey, E.; Roest Crollius, H.; Sharakhov, I.V.; Vaury, C. Conserved Small Nucleotidic Elements at the Origin of Concerted piRNA Biogenesis from Genes and lncRNAs. Cells 2020, 9, 1491. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.H.; Lee, B.; Li, X.Z. The birth of piRNAs: How mammalian piRNAs are produced, originated, and evolved. Mamm. Genome 2022, 33, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Feltzin, V.L.; Khaladkar, M.; Abe, M.; Parisi, M.; Hendriks, G.J.; Kim, J.; Bonini, N.M. The exonuclease Nibbler regulates age-associated traits and modulates piRNA length in Drosophila. Aging Cell 2015, 14, 443–452. [Google Scholar] [CrossRef]
- Cacchione, S.; Cenci, G.; Raffa, G.D. Silence at the End: How Drosophila Regulates Expression and Transposition of Telomeric Retroelements. J. Mol. Biol. 2020, 432, 4305–4321. [Google Scholar] [CrossRef] [PubMed]
- Gou, L.T.; Dai, P.; Yang, J.H.; Xue, Y.; Hu, Y.P.; Zhou, Y.; Kang, J.Y.; Wang, X.; Li, H.; Hua, M.M.; et al. Pachytene piRNAs instruct massive mRNA elimination during late spermiogenesis. Cell Res. 2014, 24, 680–700. [Google Scholar] [CrossRef] [PubMed]
- Goh, W.S.; Falciatori, I.; Tam, O.H.; Burgess, R.; Meikar, O.; Kotaja, N.; Hammell, M.; Hannon, G.J. piRNA-directed cleavage of meiotic transcripts regulates spermatogenesis. Genes. Dev. 2015, 29, 1032–1044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Kang, J.Y.; Gou, L.T.; Wang, J.; Xue, Y.; Skogerboe, G.; Dai, P.; Huang, D.W.; Chen, R.; Fu, X.D.; et al. MIWI and piRNA-mediated cleavage of messenger RNAs in mouse testes. Cell Res. 2015, 25, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.H.; Wang, R.H.; Du, K.; Zhu, J.; Zheng, J.; Xie, L.H.; Pereira, A.A.; Zhang, C.; Ricci, E.P.; Li, X.Z. Coupled protein synthesis and ribosome-guided piRNA processing on mRNAs. Nat. Commun. 2021, 12, 5970. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.H.; Cui, H.; Song, C.; Shen, J.T.; Zhuo, X.; Wang, R.H.; Yu, X.; Ndamba, R.; Mu, Q.; Gu, H.; et al. Amniotes co-opt intrinsic genetic instability to protect germ-line genome integrity. Nat. Commun. 2023, 14, 812. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Huang, Y.; Ding, Y.; Shi, L.; Zhong, X.; Kang, M.; Li, C. Analysis of piRNA expression spectra in a non-alcoholic fatty liver disease mouse model induced by a methionine- and choline-deficient diet. Exp. Ther. Med. 2020, 19, 3829–3839. [Google Scholar] [CrossRef] [PubMed]
- Subhramanyam, C.S.; Cao, Q.; Wang, C.; Heng, Z.S.L.; Zhou, Z.; Hu, Q. Role of PIWI-like 4 in modulating neuronal differentiation from human embryonal carcinoma cells. RNA Biol. 2020, 17, 1613–1624. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Banerjee, S.; Zhou, H.; Jammalamadaka, A.; Arcila, M.; Manjunath, B.S.; Kosik, K.S. Identification of piRNAs in the central nervous system. Rna 2011, 17, 1090–1099. [Google Scholar] [CrossRef] [PubMed]
- Galton, R.; Fejes-Toth, K.; Bronner, M.E. Co-option of the piRNA pathway to regulate neural crest specification. Sci. Adv. 2022, 8, eabn1441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Tu, S.; Stubna, M.; Wu, W.S.; Huang, W.C.; Weng, Z.; Lee, H.C. The piRNA targeting rules and the resistance to piRNA silencing in endogenous genes. Science 2018, 359, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.; Yu, K.; Sun, S.; Xie, C.; Skogerbo, G.; Miao, R.; Xiao, H.; Liao, Q.; Luo, H.; Zhao, G.; et al. NONCODE v3.0: Integrative annotation of long noncoding RNAs. Nucleic Acids Res. 2012, 40, D210–D215. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Yuan, J.; Li, H.; Li, M.; Zhao, G.; Bu, D.; Zhu, W.; Wu, W.; Chen, R.; Zhao, Y. NONCODEv4: Exploring the world of long non-coding RNA genes. Nucleic Acids Res. 2013, 42, D98–D103. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Liu, S.; Wang, J.; Xuan, J.; Zhang, X.; Qu, L.; Zheng, L.; Yang, J. deepBase v3.0: Expression atlas and interactive analysis of ncRNAs from thousands of deep-sequencing data. Nucleic Acids Res. 2021, 49, D877–D883. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Shao, P.; Zhou, H.; Chen, Y.Q.; Qu, L.H. deepBase: A database for deeply annotating and mining deep sequencing data. Nucleic Acids Res. 2010, 38, D123–D130. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, Y.; Zhou, H.; Zhang, P.; Song, T.; Ying, Z.; Yu, H.; Li, Y.; Zhao, Y.; Zeng, X.; et al. piRBase: Integrating piRNA annotation in all aspects. Nucleic Acids Res. 2022, 50, D265–D272. [Google Scholar] [CrossRef] [PubMed]
- Sai Lakshmi, S.; Agrawal, S. piRNABank: A web resource on classified and clustered Piwi-interacting RNAs. Nucleic Acids Res. 2008, 36, D173–D177. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, D.; Zischler, H.; Gebert, D. piRNAclusterDB 2.0: Update and expansion of the piRNA cluster database. Nucleic Acids Res. 2022, 50, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.S.; Brown, J.S.; Chen, T.T.; Chu, Y.H.; Huang, W.C.; Tu, S.; Lee, H.C. piRTarBase: A database of piRNA targeting sites and their roles in gene regulation. Nucleic Acids Res. 2019, 47, D181–D187. [Google Scholar] [CrossRef] [PubMed]
- Barrenada, O.; Larriba, E.; Brieno-Enriquez, M.A.; Mazo, J.D. piRNA-IPdb: A PIWI-bound piRNAs database to mining NGS sncRNA data and beyond. BMC Genom. 2021, 22, 765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Si, X.; Skogerbo, G.; Wang, J.; Cui, D.; Li, Y.; Sun, X.; Liu, L.; Sun, B.; Chen, R.; et al. piRBase: A web resource assisting piRNA functional study. Database 2014, 2014, bau110. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, P.; Lu, Y.; Li, Y.; Zheng, Y.; Kan, Y.; Chen, R.; He, S. piRBase: A comprehensive database of piRNA sequences. Nucleic Acids Res. 2019, 47, D175–D180. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.D.; Alam, W.; Tayara, H.; Chong, K.T. Identification of Functional piRNAs Using a Convolutional Neural Network. IEEE/ACM Trans. Comput. Biol. Bioinform. 2022, 19, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Hoeksema, J.; Liang, C. piRNN: Deep learning algorithm for piRNA prediction. PeerJ 2018, 6, e5429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, L.; Li, R.; Liu, N.; Huang, X.; Wong, G. PIWI-interacting RNAs in human diseases: Databases and computational models. Brief. Bioinform. 2022, 23, bbac217. [Google Scholar] [CrossRef]
- Wang, H.; Shi, B.; Zhang, X.; Shen, P.; He, Q.; Yin, M.; Pan, Y.; Ma, J. Exosomal hsa-piR1089 promotes proliferation and migration in neuroblastoma via targeting KEAP1. Pathol. Res. Pract. 2023, 241, 154240. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.P.; Yao, M.J.; Chang, S.Y.; Gou, L.T.; Liu, M.F.; Qiu, Z.L.; Yuan, X.B. Novel function of PIWIL1 in neuronal polarization and migration via regulation of microtubule-associated proteins. Mol. Brain 2015, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Gasperini, C.; Tuntevski, K.; Beatini, S.; Pelizzoli, R.; Lo Van, A.; Mangoni, D.; Cossu, R.M.; Pascarella, G.; Bianchini, P.; Bielefeld, P.; et al. Piwil2 (Mili) sustains neurogenesis and prevents cellular senescence in the postnatal hippocampus. EMBO Rep. 2023, 24, e53801. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Chen, M.; Pang, T.; Li, X.; Long, L.; Liang, D.; Peng, L.; Sun, W.; Xu, E. Attenuation of Piwil2 induced by hypoxic postconditioning prevents cerebral ischemic injury by inhibiting CREB2 promoter methylation. Brain Pathol. 2023, 33, e13109. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, F.; Liu, X.; Liu, Q.; Fu, Y.; Li, R.; Hou, L.; Zhang, J.; He, J.; Kang, L. Piwi/piRNAs control food intake by promoting neuropeptide F expression in locusts. EMBO Rep. 2022, 23, e50851. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.; Madhwani, K.; Chaurasia, B. PiWi RNA in Neurodevelopment and Neurodegenerative Disorders. Curr. Mol. Pharmacol. 2022, 15, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Penning, A.; Tosoni, G.; Abiega, O.; Bielefeld, P.; Gasperini, C.; De Pietri Tonelli, D.; Fitzsimons, C.P.; Salta, E. Adult Neural Stem Cell Regulation by Small Non-coding RNAs: Physiological Significance and Pathological Implications. Front. Cell Neurosci. 2021, 15, 781434. [Google Scholar] [CrossRef] [PubMed]
- Perera, B.P.U.; Tsai, Z.T.; Colwell, M.L.; Jones, T.R.; Goodrich, J.M.; Wang, K.; Sartor, M.A.; Faulk, C.; Dolinoy, D.C. Somatic expression of piRNA and associated machinery in the mouse identifies short, tissue-specific piRNA. Epigenetics 2019, 14, 504–521. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, H.; Perriot, S.; Gastfriend, B.D.; Steinfort, M.; Cibien, C.; Soldati, S.; Matsuo, K.; Guimbal, S.; Mathias, A.; Palecek, S.P.; et al. Intrinsic blood-brain barrier dysfunction contributes to multiple sclerosis pathogenesis. Brain 2022, 145, 4334–4348. [Google Scholar] [CrossRef] [PubMed]
- Hang, Z.; Zhou, L.; Xing, C.; Wen, Y.; Du, H. The blood-brain barrier, a key bridge to treat neurodegenerative diseases. Ageing Res. Rev. 2023, 91, 102070. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Pattnaik, S.; Sivagurunathan, S.; Chidambaram, S. Small ncRNA binding protein, PIWI: A potential molecular bridge between blood brain barrier and neuropathological conditions. Med. Hypotheses 2020, 138, 109609. [Google Scholar] [CrossRef] [PubMed]
- Sivagurunathan, S.; Palanisamy, K.; Arunachalam, J.P.; Chidambaram, S. Possible role of HIWI2 in modulating tight junction proteins in retinal pigment epithelial cells through Akt signaling pathway. Mol. Cell. Biochem. 2016, 427, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, D.; Ghirardi, M.; Skehel, P.A.; Karl, K.A.; Herder, S.P.; Chen, M.; Bailey, C.H.; Kandel, E.R. Aplysia CREB2 represses long-term facilitation: Relief of repression converts transient facilitation into long-term functional and structural change. Cell 1995, 83, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Yang, Z. Dynamical Mechanisms for Gene Regulation Mediated by Two Noncoding RNAs in Long-Term Memory Formation. Neural Plast. 2021, 2021, 6668389. [Google Scholar] [CrossRef]
- Craske, M.G.; Rauch, S.L.; Ursano, R.; Prenoveau, J.; Pine, D.S.; Zinbarg, R.E. What is an anxiety disorder? Depress. Anxiety 2009, 26, 1066–1085. [Google Scholar] [CrossRef] [PubMed]
- Nandi, S.; Chandramohan, D.; Fioriti, L.; Melnick, A.M.; Hebert, J.M.; Mason, C.E.; Rajasethupathy, P.; Kandel, E.R. Roles for small noncoding RNAs in silencing of retrotransposons in the mammalian brain. Proc. Natl. Acad. Sci. USA 2016, 113, 12697–12702. [Google Scholar] [CrossRef] [PubMed]
- Donkin, I.; Versteyhe, S.; Ingerslev, L.R.; Qian, K.; Mechta, M.; Nordkap, L.; Mortensen, B.; Appel, E.V.; Jorgensen, N.; Kristiansen, V.B.; et al. Obesity and Bariatric Surgery Drive Epigenetic Variation of Spermatozoa in Humans. Cell Metab. 2016, 23, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sinnott-Armstrong, N.; Wagschal, A.; Wark, A.R.; Camporez, J.P.; Perry, R.J.; Ji, F.; Sohn, Y.; Oh, J.; Wu, S.; et al. A MicroRNA Linking Human Positive Selection and Metabolic Disorders. Cell 2020, 183, 684–701.e14. [Google Scholar] [CrossRef] [PubMed]
- Vinnikov, I.A.; Hajdukiewicz, K.; Reymann, J.; Beneke, J.; Czajkowski, R.; Roth, L.C.; Novak, M.; Roller, A.; Dörner, N.; Starkuviene, V.; et al. Hypothalamic miR-103 protects from hyperphagic obesity in mice. J. Neurosci. 2014, 34, 10659–10674. [Google Scholar] [CrossRef] [PubMed]
- Falibene, A.; Rossler, W.; Josens, R. Serotonin depresses feeding behaviour in ants. J. Insect Physiol. 2012, 58, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Curcio, M.; Bradke, F. Axon Regeneration in the Central Nervous System: Facing the Challenges from the Inside. Annu. Rev. Cell Dev. Biol. 2018, 34, 495–521. [Google Scholar] [CrossRef] [PubMed]
- Phay, M.; Kim, H.H.; Yoo, S. Analysis of piRNA-Like Small Non-coding RNAs Present in Axons of Adult Sensory Neurons. Mol. Neurobiol. 2018, 55, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Parenti, I.; Rabaneda, L.G.; Schoen, H.; Novarino, G. Neurodevelopmental Disorders: From Genetics to Functional Pathways. Trends Neurosci. 2020, 43, 608–621. [Google Scholar] [CrossRef]
- Kuo, M.C.; Liu, S.C.; Hsu, Y.F.; Wu, R.M. The role of noncoding RNAs in Parkinson’s disease: Biomarkers and associations with pathogenic pathways. J. Biomed. Sci. 2021, 28, 78. [Google Scholar] [CrossRef]
- Kim, Y.S.; Choi, J.; Yoon, B.E. Neuron-Glia Interactions in Neurodevelopmental Disorders. Cells 2020, 9, 2176. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.C.; Bae, Y.; Lee, S.V. The Role of mRNA Quality Control in the Aging of Caenorhabditis elegans. Mol. Cells 2023, 46, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wong, G. An old weapon with a new function: PIWI-interacting RNAs in neurodegenerative diseases. Transl. Neurodegener. 2021, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Dharap, A.; Nakka, V.P.; Vemuganti, R. Altered expression of PIWI RNA in the rat brain after transient focal ischemia. Stroke 2011, 42, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Pierce, L.M.; Kurata, W.E.; Matsumoto, K.W.; Clark, M.E.; Farmer, D.M. Long-term epigenetic alterations in a rat model of Gulf War Illness. Neurotoxicology 2016, 55, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Wang, C.; Chen, L.; Wong, G. Dysregulation of MicroRNAs and PIWI-Interacting RNAs in a Caenorhabditis elegans Parkinson’s Disease Model Overexpressing Human alpha-Synuclein and Influence of tdp-1. Front. Neurosci. 2021, 15, 600462. [Google Scholar] [CrossRef] [PubMed]
- Frost, B.; Hemberg, M.; Lewis, J.; Feany, M.B. Tau promotes neurodegeneration through global chromatin relaxation. Nat. Neurosci. 2014, 17, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Wakisaka, K.T.; Tanaka, R.; Hirashima, T.; Muraoka, Y.; Azuma, Y.; Yoshida, H.; Tokuda, T.; Asada, S.; Suda, K.; Ichiyanagi, K.; et al. Novel roles of Drosophila FUS and Aub responsible for piRNA biogenesis in neuronal disorders. Brain Res. 2019, 1708, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Sarmah, D.; Saraf, J.; Vats, K.; Kalia, K.; Borah, A.; Yavagal, D.R.; Dave, K.R.; Ghosh, Z.; Bhattacharya, P. Noncoding RNAs in ischemic stroke: Time to translate. Ann. N.Y Acad. Sci. 2018, 1421, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Saugstad, J.A. Non-Coding RNAs in Stroke and Neuroprotection. Front. Neurol. 2015, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Potemkin, N.; Clarkson, A.N. Non-coding RNAs in stroke pathology, diagnostics, and therapeutics. Neurochem. Int. 2023, 162, 105467. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Hussen, B.M.; Taheri, M.; Arefian, N. Emerging Impact of Non-coding RNAs in the Pathology of Stroke. Front. Aging Neurosci. 2021, 13, 780489. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.; Madhwani, K. Coding and non-coding nucleotides’: The future of stroke gene therapeutics. Genomics 2021, 113, 1291–1307. [Google Scholar] [CrossRef] [PubMed]
- Carroll, D. RNA in Therapeutics: CRISPR in the Clinic. Mol. Cells 2023, 46, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, S.F.; Shore, M.F. Prevention of psychiatric disorders. Harv. Rev. Psychiatry 1995, 3, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Dwivedi, Y. Non-Coding RNAs in Psychiatric Disorders and Suicidal Behavior. Front. Psychiatry 2020, 11, 543893. [Google Scholar] [CrossRef] [PubMed]
- Ragan, C.; Patel, K.; Edson, J.; Zhang, Z.H.; Gratten, J.; Mowry, B. Small non-coding RNA expression from anterior cingulate cortex in schizophrenia shows sex specific regulation. Schizophr. Res. 2017, 183, 82–87. [Google Scholar] [CrossRef] [PubMed]
- McDaniel Peters, B.C.; Wood, W. Autism and Equine-Assisted Interventions: A Systematic Mapping Review. J. Autism Dev. Disord. 2017, 47, 3220–3242. [Google Scholar] [CrossRef] [PubMed]
- de Giambattista, C.; Ventura, P.; Trerotoli, P.; Margari, M.; Palumbi, R.; Margari, L. Subtyping the Autism Spectrum Disorder: Comparison of Children with High Functioning Autism and Asperger Syndrome. J. Autism Dev. Disord. 2019, 49, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Chiappori, F.; Cupaioli, F.A.; Consiglio, A.; Di Nanni, N.; Mosca, E.; Licciulli, V.F.; Mezzelani, A. Analysis of Faecal Microbiota and Small ncRNAs in Autism: Detection of miRNAs and piRNAs with Possible Implications in Host-Gut Microbiota Cross-Talk. Nutrients 2022, 14, 1340. [Google Scholar] [CrossRef] [PubMed]
- Salloum-Asfar, S.; Elsayed, A.K.; Elhag, S.F.; Abdulla, S.A. Circulating Non-Coding RNAs as a Signature of Autism Spectrum Disorder Symptomatology. Int. J. Mol. Sci. 2021, 22, 6549. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Sohl, K.; Levitskiy, D.; Tennant, P.; Goin-Kochel, R.P.; Shaffer, R.C.; Confair, A.; Middleton, F.A.; Hicks, S.D. Saliva RNA Biomarkers of Gastrointestinal Dysfunction in Children with Autism and Neurodevelopmental Disorders: Potential Implications for Precision Medicine. Front. Psychiatry 2021, 12, 824933. [Google Scholar] [CrossRef]
- Sato, K.; Takayama, K.I.; Inoue, S. Role of piRNA biogenesis and its neuronal function in the development of neurodegenerative diseases. Front. Aging Neurosci. 2023, 15, 1157818. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Kundu, S.; Singh, A.; Singh, S. Understanding the Role of Histone Deacetylase and their Inhibitors in Neurodegenerative Disorders: Current Targets and Future Perspective. Curr. Neuropharmacol. 2022, 20, 158–178. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Vondrakova, D.; Lawson, M.; Valko, M. Metals, oxidative stress and neurodegenerative disorders. Mol. Cell Biochem. 2010, 345, 91–104. [Google Scholar] [CrossRef]
- Cooke, P.; Janowitz, H.; Dougherty, S.E. Neuronal Redevelopment and the Regeneration of Neuromodulatory Axons in the Adult Mammalian Central Nervous System. Front. Cell Neurosci. 2022, 16, 872501. [Google Scholar] [CrossRef] [PubMed]
- Lauretti, E.; Dabrowski, K.; Pratico, D. The neurobiology of non-coding RNAs and Alzheimer’s disease pathogenesis: Pathways, mechanisms and translational opportunities. Ageing Res. Rev. 2021, 71, 101425. [Google Scholar] [CrossRef]
- Tan, L.; Yu, J.T.; Hu, N.; Tan, L. Non-coding RNAs in Alzheimer’s disease. Mol. Neurobiol. 2013, 47, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chetelat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Wang, J.Z.; Grundke-Iqbal, I.; Iqbal, K. Kinases and phosphatases and tau sites involved in Alzheimer neurofibrillary degeneration. Eur. J. Neurosci. 2007, 25, 59–68. [Google Scholar] [CrossRef]
- Saba, R.; Goodman, C.D.; Huzarewich, R.L.; Robertson, C.; Booth, S.A. A miRNA signature of prion induced neurodegeneration. PLoS ONE 2008, 3, e3652. [Google Scholar] [CrossRef] [PubMed]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Papadopoulou, A.S.; Mandemakers, W.; Silahtaroglu, A.N.; Kauppinen, S.; Delacourte, A.; De Strooper, B. Loss of microRNA cluster miR-29a/b-1 in sporadic Alzheimer’s disease correlates with increased BACE1/beta-secretase expression. Proc. Natl. Acad. Sci. USA 2008, 105, 6415–6420. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K.; Ojala, J. ER stress in Alzheimer’s disease: A novel neuronal trigger for inflammation and Alzheimer’s pathology. J. Neuroinflammation 2009, 6, 41. [Google Scholar] [CrossRef]
- Qiu, W.; Guo, X.; Lin, X.; Yang, Q.; Zhang, W.; Zhang, Y.; Zuo, L.; Zhu, Y.; Li, C.R.; Ma, C.; et al. Transcriptome-wide piRNA profiling in human brains of Alzheimer’s disease. Neurobiol. Aging 2017, 57, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Sarkar, A.; Parida, S.; Ghosh, Z.; Mallick, B. Small RNA sequencing revealed dysregulated piRNAs in Alzheimer’s disease and their probable role in pathogenesis. Mol. Biosyst. 2017, 13, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Qiu, W.; Garcia-Milian, R.; Lin, X.; Zhang, Y.; Cao, Y.; Tan, Y.; Wang, Z.; Shi, J.; Wang, J.; et al. Genome-wide significant, replicated and functional risk variants for Alzheimer’s disease. J. Neural Transm. 2017, 124, 1455–1471. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Fan, L.; Wang, X.; Lin, X.; Cao, Y.; Zheng, C.; Zhang, Y.; Zhang, H.; Garcia-Milian, R.; Kang, L.; et al. Transcriptome-wide piRNA profiling in human brains for aging genetic factors. Jacobs J. Genet. 2019, 4, 14. [Google Scholar]
- Jain, G.; Stuendl, A.; Rao, P.; Berulava, T.; Pena Centeno, T.; Kaurani, L.; Burkhardt, S.; Delalle, I.; Kornhuber, J.; Hull, M.; et al. A combined miRNA-piRNA signature to detect Alzheimer’s disease. Transl. Psychiatry 2019, 9, 250. [Google Scholar] [CrossRef]
- Fitz, N.F.; Wang, J.; Kamboh, M.I.; Koldamova, R.; Lefterov, I. Small nucleolar RNAs in plasma extracellular vesicles and their discriminatory power as diagnostic biomarkers of Alzheimer’s disease. Neurobiol. Dis. 2021, 159, 105481. [Google Scholar] [CrossRef]
- Olufunmilayo, E.O.; Holsinger, R.M.D. Roles of Non-Coding RNA in Alzheimer’s Disease Pathophysiology. Int. J. Mol. Sci. 2023, 24, 12498. [Google Scholar] [CrossRef]
- Costa, H.N.; Esteves, A.R.; Empadinhas, N.; Cardoso, S.M. Parkinson’s Disease: A Multisystem Disorder. Neurosci. Bull. 2023, 39, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Paccosi, E.; Proietti-De-Santis, L. Parkinson’s Disease: From Genetics and Epigenetics to Treatment, a miRNA-Based Strategy. Int. J. Mol. Sci. 2023, 24, 9547. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, S.; Darvish, E.; Narimani, F.; Moradi, S.Z.; Abbaszadeh, F.; Khan, H. The regulatory role of non-coding RNAs and their interactions with phytochemicals in neurodegenerative diseases: A systematic review. Brief. Funct. Genom. 2023, 22, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Galvagnion, C.; Marlet, F.R.; Cerri, S.; Schapira, A.H.V.; Blandini, F.; Di Monte, D.A. Sphingolipid changes in Parkinson L444P GBA mutation fibroblasts promote α-synuclein aggregation. Brain 2022, 145, 1038–1051. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, O.; Gana-Weisz, M.; Banfi, S.; Nigro, V.; Bar-Shira, A.; Thaler, A.; Gurevich, T.; Mirelman, A.; Giladi, N.; Alcalay, R.N.; et al. Novel variants in genes related to vesicle-mediated-transport modify Parkinson’s disease risk. Mol. Genet. Metab. 2023, 139, 107608. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Pan, H.; Zhao, Y.; Wang, Y.; Xu, Q.; Tan, J.; Yan, X.; Li, J.; Tang, B.; Guo, J. Association Study of TAF1 Variants in Parkinson’s Disease. Front. Neurosci. 2022, 16, 846095. [Google Scholar] [CrossRef] [PubMed]
- Ohira, K.; Yokota, H.; Hirano, S.; Nishimura, M.; Mukai, H.; Horikoshi, T.; Sawai, S.; Yamanaka, Y.; Yamamoto, T.; Kakeda, S.; et al. DRD2 Taq1A Polymorphism-Related Brain Volume Changes in Parkinson’s Disease: Voxel-Based Morphometry. Park. Dis. 2022, 2022, 8649195. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, M.; Annesi, G.; Procopio, R.; Morelli, M.; Iannello, G.; Bonapace, G.; Mancini, M.; Nicoletti, G.; Quattrone, A. DNAJC13 mutation screening in patients with Parkinson’s disease from South Italy. Park. Relat. Disord. 2018, 55, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.; Sommer, A.; Plotz, S.; Farrell, M.; Winner, B.; Grosch, J.; Winkler, J.; Riemenschneider, M.J. Sporadic Parkinson’s disease derived neuronal cells show disease-specific mRNA and small RNA signatures with abundant deregulation of piRNAs. Acta Neuropathol. Commun. 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Nataf, S.; Guillen, M.; Pays, L. Common Neurodegeneration-Associated Proteins Are Physiologically Expressed by Human B Lymphocytes and Are Interconnected via the Inflammation/Autophagy-Related Proteins TRAF6 and SQSTM1. Front. Immunol. 2019, 10, 2704. [Google Scholar] [CrossRef] [PubMed]
- Aisha, Z.; Lei, J.; Zhang, Y.; Ma, J. EEF1A1 is Involved the Regulating Neuroinflammatory Processes in Parkinson’s Disease. J. Integr. Neurosci. 2023, 22, 122. [Google Scholar] [CrossRef] [PubMed]
- Simoes, F.A.; Joilin, G.; Peters, O.; Schneider, L.S.; Priller, J.; Spruth, E.J.; Vogt, I.; Kimmich, O.; Spottke, A.; Hoffmann, D.C.; et al. Potential of Non-Coding RNA as Biomarkers for Progressive Supranuclear Palsy. Int. J. Mol. Sci. 2022, 23, 14554. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, N.J.; Zhang, Y.J.; Baker, M.; Gass, J.M.; Finch, N.A.; Xu, Y.F.; Stewart, H.; Kelley, B.J.; Kuntz, K.; Crook, R.J.; et al. Novel mutations in TARDBP (TDP-43) in patients with familial amyotrophic lateral sclerosis. PLoS Genet. 2008, 4, e1000193. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, J.; Blair, I.P.; Tripathi, V.B.; Hu, X.; Vance, C.; Rogelj, B.; Ackerley, S.; Durnall, J.C.; Williams, K.L.; Buratti, E.; et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science 2008, 319, 1668–1672. [Google Scholar] [CrossRef] [PubMed]
- Couthouis, J.; Hart, M.P.; Erion, R.; King, O.D.; Diaz, Z.; Nakaya, T.; Ibrahim, F.; Kim, H.J.; Mojsilovic-Petrovic, J.; Panossian, S.; et al. Evaluating the role of the FUS/TLS-related gene EWSR1 in amyotrophic lateral sclerosis. Hum. Mol. Genet. 2012, 21, 2899–2911. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.; Rogelj, B.; Hortobagyi, T.; De Vos, K.J.; Nishimura, A.L.; Sreedharan, J.; Hu, X.; Smith, B.; Ruddy, D.; Wright, P.; et al. Mutations in FUS, an RNA processing protein, cause familial amyotrophic lateral sclerosis type 6. Science 2009, 323, 1208–1211. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, R.F.; Ogawa, K.; Beck, G.; Ikenaka, K.; Takeuchi, E.; Yasumizu, Y.; Jinno, J.; Kimura, Y.; Baba, K.; Nagai, Y.; et al. piRNA/PIWI Protein Complex as a Potential Biomarker in Sporadic Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2022, 59, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Joilin, G.; Gray, E.; Thompson, A.G.; Talbot, K.; Leigh, P.N.; Newbury, S.F.; Turner, M.R.; Hafezparast, M. Profiling non-coding RNA expression in cerebrospinal fluid of amyotrophic lateral sclerosis patients. Ann. Med. 2022, 54, 3069–3078. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Database | Species | Function | Website | Reference |
|---|---|---|---|---|
| Public databases | ||||
| NONCODE | Covers 17 species (Human, Mouse, Cow, Rat, Pig, Drosophila, and Nematode) | annotation, potential, function, and mapping | http://www.noncode.org/, accessed on 9 May 2024 | [72,73] |
| deepBase | Insect, Nematode, Insect, and Mammal | expression profile and interaction function | http://rna.sysu.edu.cn/deepbase3/index.html, accessed on 9 May 2024 | [74,75] |
| Specific databases | ||||
| PiRbase | Covers 44 species (Human, Mouse, Rat, Zebrafish, Chicken, Silkworm and Rabbit) | new sequence, gene target, gold standard set, cluster, function, and network | http://bigdata.ibp.ac.cn/piRBase, accessed on 9 May 2024 | [76] |
| piRNA bank | Human, Mouse, and Rat | piRNAs cluster, map, and homologous and chromosomal locations | http://pirnabank.ibab.ac.in/, accessed on 9 May 2024 | [77] |
| PiRNAclustersDB | Cover 51 species (Mollusks, Arthropods, Fish, Amphibians, Reptiles, and Mammals) | piRNAs cluster database | https://www.smallrnagroup.uni-mainz.de/piRNAclusterDB, accessed on 9 May 2024 | [78] |
| piRTarBase | C. elegans | target prediction | http://cosbi6.ee.ncku.edu.tw/piRTarBase, accessed on 9 May 2024 | [79] |
| piRNA-IPdb | Mouse | length distribution, nucleotide composition, and expression level | https://ipdb2.shinyapps.io/ipdb2/, accessed on 9 May 2024 | [80] |
| Disease | Species | Models | Results | Reference |
|---|---|---|---|---|
| stroke | Rat | ischemia | 54 up- and 51 down-regulated piRNAs were detected | [114] |
| ischemia tolerance | piR-000618, -017990, and -014971 are up-regulated by targeting CREB2 | [89] | ||
| Gulf War illness | Rat | GWI-relevant exposures | piR-007899 and piR-019162 | [115] |
| nerve injury | Mice | MIWI/PIWIL1 KO | up-regulated piR-14384, -69959, -52953, and -52274 | [19] |
| AD | C. elegans | Overexpressing α-syn pan neuron | 21ur-10488 and 21ur-2781 were related to the AD pathology | [116] |
| Drosophila | tau transgenic | down-regulated Ago3 | [117] | |
| Ago3/PIWIL1 KD in tau transgenic | improvement of locomotor defect | [27] | ||
| PD | C. elegans | tau transgenic | 79 up- and 33 down-regulated piRNAs | [117] |
| Overexpressing A53T pan neuron and co-expressing Aβ and α-syn pan neuron | up-regulated 21ur-10824, 21ur-11898, and 21ur-13215 | [28] | ||
| ALS | Drosophila | Caz KD | Aub was overexpressed, while pre-piRNAs were increased | [118] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, X.; Dai, W.; Wang, Z.; Li, S.; Sun, T.; Miao, N. PIWI-Interacting RNAs: A Pivotal Regulator in Neurological Development and Disease. Genes 2024, 15, 653. https://doi.org/10.3390/genes15060653
Pan X, Dai W, Wang Z, Li S, Sun T, Miao N. PIWI-Interacting RNAs: A Pivotal Regulator in Neurological Development and Disease. Genes. 2024; 15(6):653. https://doi.org/10.3390/genes15060653
Chicago/Turabian StylePan, Xian, Wang Dai, Zhenzhen Wang, Siqi Li, Tao Sun, and Nan Miao. 2024. "PIWI-Interacting RNAs: A Pivotal Regulator in Neurological Development and Disease" Genes 15, no. 6: 653. https://doi.org/10.3390/genes15060653
APA StylePan, X., Dai, W., Wang, Z., Li, S., Sun, T., & Miao, N. (2024). PIWI-Interacting RNAs: A Pivotal Regulator in Neurological Development and Disease. Genes, 15(6), 653. https://doi.org/10.3390/genes15060653