

Isolation, Characterization, and Expression Analysis of NAC Transcription Factor from Andrographis paniculata (Burm. f.) Nees and Their Role in Andrographolide Production
 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Retrieval and Identification of Core Fragments of NAC Genes
2.2. Plant Material
2.3. Total RNA Isolation and cDNA Synthesis
2.4. PCR Amplification and Cloning of Core Fragments
2.5. 5′ and 3′ RACE and Sequencing of Products
2.6. Sequence Analysis
2.7. Protein Structure and Their Function Prediction
2.8. Assay of Andrographolide Content
2.9. Expression Analysis Using Real-Time Quantitative PCR (qPCR)
2.10. Quantification of Andrographolide
2.11. Transient Expression of Isolated ApNAC Genes in N. benthamiana
3. Results
3.1. 5′ and 3′ RACE and Sequence Analysis
3.2. Structure-Based Function Prediction
3.3. Diterpenoid Profiling
3.4. Expression Profiling
3.5. Correlation Analysis
3.6. Effects of ApNAC Genes on Terpene Biosynthesis Genes in N. benthamiana
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chandrasekaran, C.; Thiyagarajan, P.; Sundarajan, K.; Goudar, K.S.; Deepak, M.; Murali, B.; Joshua Allan, J.; Agarwal, A. Evaluation of the genotoxic potential and acute oral toxicity of standardized extract of Andrographis paniculata (KalmColdTM). Food Chem. Toxicol. 2009, 47, 1892–1902. [Google Scholar] [CrossRef] [PubMed]
- Akbar, S. Andrographis paniculata: A review of pharmacological activities and clinical effects. Altern. Med. Rev. A J. Clin. Ther. 2011, 16, 66–77. [Google Scholar]
- Mishra, S.K.; Sangwan, N.S.; Sangwan, R.S. Phcog Rev.: Plant Review Andrographis paniculata (Kalmegh): A Review. Rev. Lit. Arts Am. 2007, 1, 283–298. [Google Scholar]
- Nyeem, M.A.; Mannan, M.A.; Nuruzzaman, M.; Kamrujjaman, K.; Das, S. Indigenous king of bitter (Andrographis paniculata): A review. J. Med. Plants Stud. 2017, 5, 318–324. [Google Scholar]
- Garg, A.; Agrawal, L.; Misra, R.C.; Sharma, S.; Ghosh, S. Andrographis paniculata transcriptome provides molecular insights into tissue-specific accumulation of medicinal diterpenes. BMC Genom. 2015, 16, 659. [Google Scholar] [CrossRef]
- Abass, S.; Zahiruddin, S.; Ali, A.; Irfan, M.; Jan, B.; Haq, Q.M.R.; Husain, S.A.; Ahmad, S. Development of Synergy-Based Combination of Methanolic Extract of Andrographis paniculata and Berberis aristata against E. coli and S. aureus. Curr. Microbiol. 2022, 79, 223. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.A.; Sridevi, K.; Kumar, N.V.; Nanduri, S.; Rajagopal, S. Anticancer and immunostimulatory compounds from Andrographis paniculata. J. Ethnopharmacol. 2004, 92, 291–295. [Google Scholar] [CrossRef]
- Bastiana; Widyawaruyanti, A.; Ilmi, H.; Tumewu, L.; Prasetyo, B.; Hafid, A.F.; Aryati. A tablet derived from Andrographis paniculata complements dihydroartemisinin-piperaquine treatment of malaria in pregnant mice. J. Basic Clin. Physiol. Pharmacol. 2021, 33, 175–183. [Google Scholar] [CrossRef]
- Elasoru, S.E.; Rhana, P.; de Oliveira Barreto, T.; Naves de Souza, D.L.; Menezes-Filho, J.E.R.; Souza, D.S.; Moreira, M.V.L.; Campos, M.T.G.; Adedosu, O.T.; Roman-Campos, D.; et al. Andrographolide protects against isoproterenol-induced myocardial infarction in rats through inhibition of L-type Ca2+ and increase of cardiac transient outward K+ currents. Eur. J. Pharmacol. 2021, 906, 174194. [Google Scholar] [CrossRef]
- Gupta, S.; Choudhry, M.A.; Yadava, J.N.S.; Srivastava, V.; Tandon, J.S. Antidiarrhoeal activity of diterpenes of Andrographis paniculata (kal-megh) against escherichia coli enterotoxin in in vivo models. Pharm. Biol. 1990, 28, 273–283. [Google Scholar] [CrossRef]
- Kaur, R.; Sharma, P.; Gupta, G.K.; Ntie-Kang, F.; Kumar, D. Structure-activity-relationship and mechanistic insights for anti-HIV natural products. Molecules 2020, 25, 2070. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Baek, C.Y.; Hwang, J.H.; Kim, M.Y. Andrographis paniculata Extract Relieves Pain and Inflammation in Monosodium Iodoacetate-Induced Osteoarthritis and Acetic Acid-Induced Writhing in Animal Models. Process 2020, 8, 873. [Google Scholar] [CrossRef]
- Loh, S.; Tsai, Y.; Huang, S.; Yu, T.; Kuo, P.; Chao, S.C.; Chou, M.F.; Tsai, C.S.; Lee, S.P. Effects of andrographolide on intracellular ph regulation, cellular migration, and apoptosis in human cervical cancer cells. Cancers 2020, 12, 387. [Google Scholar] [CrossRef] [PubMed]
- Mussard, E.; Cesaro, A.; Lespessailles, E.; Legrain, B.; Berteina-Raboin, S.; Toumi, H. Andrographolide, A Natural Antioxidant: An Update. Antioxidants 2019, 8, 571. [Google Scholar] [CrossRef] [PubMed]
- Ogundola, A.F.; Akhigbe, R.E.; Saka, W.A.; Adeniyi, A.O.; Adeshina, O.S.; Babalola, D.O.; Akhigbe, T.M. Contraceptive potential of Andrographis paniculata is via androgen suppression and not induction of oxidative stress in male Wistar rats. Tissue Cell 2021, 73, 101632. [Google Scholar] [CrossRef] [PubMed]
- Rajanna, M.P.; Bharathi, B.K.; Shivakumar, B.R.; Deepak, M.; Prashanth, D.; Prabakaran, D.; Vijayabhaskar, T.; Arun, B. Immunomodulatory effects of Andrographis paniculata extract in healthy adults—An open-label study. J. Ayurveda Integr. Med. 2021, 12, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singh, R.T.; Sehgal, V.; Handa, S.S. Antihepatotoxic activity of some plants used in herbal formulations. Fitoterapia 1991, 62, 131–138. [Google Scholar]
- Sharma, V.; Qayum, A.; Kapoor, K.K.; Mukherjee, D.; Singh, S.K.; Dhar, M.K.; Kaul, S. Synthesis of 14-deoxy-benzylidene-8,17-epoxy-diene-andrographolide derivatives and evaluation of their anticancer activities. J. Indian Chem. Soc. 2022, 99, 100490. [Google Scholar] [CrossRef]
- Tohkayomatee, R.; Reabroi, S.; Tungmunnithum, D.; Parichatikanond, W.; Pinthong, D. Andrographolide Exhibits Anticancer Activity against Breast Cancer Cells (MCF-7 and MDA-MB-231 Cells) through Suppressing Cell Proliferation and Inducing Cell apoptosis via inactivation of ER-α receptor and PI3K/AKT/mTOR signaling. Molecules 2022, 27, 3544. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, N.P.; Rawal, U.M. Hepatoprotective and antioxidant property of Andrographis paniculata (Nees) in BHC induced liver damage in mice. Indian J. Exp. Biol. 2001, 39, 41–46. [Google Scholar]
- Visen, P.K.; Saraswat, B.; Vuksan, V.; Dhawan, B.N. Effect of andrographolide on monkey hepatocytes against galactosamine induced cell toxicity: An in-vitro study. J. Complement. Integr. Med. 2007, 4, 1. [Google Scholar] [CrossRef]
- Verma, H.; Negi, M.S.; Mahapatra, B.S.; Shukla, A.; Paul, J. Evaluation of an emerging medicinal crop Kalmegh [Andrographis paniculata (Burm. F.) Wall. Ex. Nees] for commercial cultivation and pharmaceutical & industrial uses: A review. Pharmacogn. Phytochem. 2019, 8, 835–848. [Google Scholar]
- Kanjanasirirat, P.; Suksatu, A.; Manopwisedjaroen, S.; Munyoo, B.; Tuchinda, P.; Jearawuttanakul, K.; Seemakhan, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Rangkasenee, N.; et al. High-content screening of Thai medicinal plants reveals Boesenbergia rotunda extract and its component Panduratin A as anti-SARS-CoV-2 agents. Sci. Rep. 2020, 10, 19963. [Google Scholar] [CrossRef] [PubMed]
- Sa-Ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 activity of Andrographis paniculata extract and its major component Andrographolide in human lung epithelial cells and cytotoxicity evaluation in major organ cell representatives. Nat. Prod. 2021, 84, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.H.; Huang, Y.L.; Chen, C.C.; Pi, W.C.; Hsu, Y.L.; Lo, L.C.; Chen, W.Y.; Fu, S.L.; Lin, C.H. Andrographolide and its fluorescent derivative inhibit the main proteases of 2019-nCoV and SARS-CoV through covalent linkage. Biochem. Biophys. Res. Commun. 2020, 533, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Duval, M.; Hsieh, T.F.; Kim, S.Y.; Thomas, T.L. Molecular characterization of AtNAM: A member of the Arabidopsis NAC domain super family. Plant Mol. Biol. 2002, 50, 237–248. [Google Scholar] [CrossRef]
- Ernst, H.A.; Nina Olsen, A.; Skriver, K.; Larsen, S.; Lo Leggio, L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. DNA-binding specificity and molecular functions of NAC transcription factors. Plant Sci. 2005, 169, 785–797. [Google Scholar] [CrossRef]
- Liu, X.; Wang, T.; Bartholomew, E.; Black, K.; Dong, M.; Zhang, Y.; Ren, H. Comprehensive analysis of NAC transcription factors and their expression during fruit spine development in cucumber (Cucumis sativus L.). Horti. Res. 2018, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248. [Google Scholar] [CrossRef]
- Guo, S.; Dai, S.; Singh, P.K.; Wang, H.; Wang, Y.; Tan, J.L.; Wee, W.; Ito, T.A. A membrane-bound NAC-like transcription factor OsNTL5 represses the flowering in Oryza sativa. Front. Plant Sci. 2018, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K.; et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef] [PubMed]
- He, X.J.; Mu, R.L.; Cao, W.H.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.M.; Zhang, H.F.; Liu, S.Y.; Wang, X.K.; Zhang, Y.M.; Meng, Y.C.; Luo, D.; Chen, R.G. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef]
- Jian, W.; Zheng, Y.; Yu, T.; Cao, H.; Chen, Y.; Cui, Q.; Xu, C.; Li, Z. SlNAC6, A NAC transcription factor, is involved in drought stress response and reproductive process in tomato. J. Plant Physiol. 2021, 264, 153483. [Google Scholar] [CrossRef] [PubMed]
- Sablowski, R.W.; Meyerowitz, E.M. A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3PISTILLATA. Cell 1998, 92, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Zhang, J.; Ren, Y.; Li, M.; Tian, S.; Yu, Y.; Zuo, Y.; Gong, G.; Zhang, H.; et al. The NAC transcription factor ClNAC68 positively regulates sugar content and seed development in watermelon by repressing ClINV and ClGH3.6. Hortic. Res. 2021, 8, 214. [Google Scholar] [CrossRef]
- Wang, S.; Huang, J.; Wang, X.; Dang, H.; Jiang, T.; Han, Y. Expression analysis of the NAC transcription factor family of populus in response to salt stress. Forests 2019, 10, 688. [Google Scholar] [CrossRef]
- Zhong, R.; Demura, T.; Ye, Z.H. SND1, a NAC domain transcription factor, is a key regulator of secondary wall synthesis in fibers of Arabidopsis. Plant Cell 2006, 18, 3158–3170. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Okada, K.; Abe, H.; Arimura, G.I. Jasmonates Induce Both Defense Responses and Communication in Monocotyledonous and Dicotyledonous Plants. Plant Cell Physiol. 2015, 56, 16–27. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van Der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Popko, J.; Hänsch, R.; Mendel, R.R.; Polle, A.; Teichmann, T. The role of abscisic acid and auxin in the response of poplar to abiotic stress. Plant Biol. 2010, 12, 242–258. [Google Scholar] [CrossRef]
- Sirichandra, C.; Davanture, M.; Turk, B.E.; Zivy, M.; Valot, B.; Leung, J.; Merlot, S. The arabidopsis ABA-activated kinase OST1 phosphorylates the bZIP transcription factor ABF3 and creates a 14-3-3 binding site involved in its turnover. PLoS ONE 2010, 5, e13935. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wang, S.; Zhang, F.; Chen, L.; Hao, X.; Pan, Q.; Fu, X.; Li, L.; Sun, X.; Tang, K. Overexpression of a Novel NAC Domain-Containing Transcription Factor Gene (AaNAC1) Enhances the Content of Artemisinin and Increases Tolerance to Drought and Botrytis cinerea in Artemisia annua. Plant Cell Physiol. 2016, 57, 1961–1971. [Google Scholar] [CrossRef]
- Ma, W.; Kang, X.; Liu, P.; She, K.; Zhang, Y.; Lin, X.; Li, B.; Chen, Z. The NAC-like transcription factor CsNAC7 positively regulates the caffeine biosynthesis-related gene yhNMT1 in Camellia sinensis. Hortic. Res. 2022, 9, uhab046. [Google Scholar] [CrossRef]
- Kumar, R.; Das, S.; Mishra, M.; Choudhury, D.R.; Sharma, K.; Kumari, A.; Singh, R. Emerging roles of NAC transcription factor in medicinal plants: Progress and prospects. 3 Biotech 2021, 11, 425. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuizen, N.J.; Chen, X.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural variation in monoterpene synthesis in kiwifruit: Transcriptional regulation of terpene synthases by NAC and ETHYLENE-INSENSITIVE3-like transcription factors. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, G.; Zhou, S.; Tu, Y.; Wang, Y.; Dong, T.; Hu, Z. A new tomato NAC (NAM ATAF1/2/CUC2) transcription factor, SlNAC4, functions as a positive regulator of fruit ripening and carotenoid accumulation. Plant Cell Physiol. 2014, 55, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Leng, L.; Yin, Q.; Xu, M.M.; Huang, M.; Xu, Z.; Zhang, Y.; Yao, H.; Wang, C.; Xiong, C.; et al. The genome of the medicinal plant Andrographis paniculata provides insight into the biosynthesis of the bioactive diterpenoid neoandrographolide. Plant J. 2019, 97, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kumar, C.; Jain, R.; Maurya, A.; Kumar, A.; Kumari, A.; Singh, R. Molecular cloning and In-silico characterization of NAC86 of Kalmegh (Andrographis paniculata). Indian J. Hortic. 2022, 79, 9–14. [Google Scholar] [CrossRef]
- Wang, J.; Qi, M.D.; Guo, J.; Shen, Y.; Lin, H.X.; Huang, L.Q. Cloning, subcellular localization, and heterologous expression of ApNAC1 gene from Andrographis paniculata. Zhongguo Zhongyao Zazhi China J. Chin. Mat. Med. 2017, 42, 890–895. [Google Scholar] [CrossRef]
- Singh, R.; Singh, A.; Mahato, A.K.; Paliwal, R.; Tiwari, G.; Kumar, A. De Novo Transcriptome Profiling for the Generation and Validation of Microsatellite Markers, Transcription Factors, and Database Development for Andrographis paniculata. Int. J. Mol. Sci. 2023, 24, 9212. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van De Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER suite: Protein structure and function prediction. Nat. Methods 2014, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Yang, J.; Zhang, Y. COFACTOR: An accurate comparative algorithm for structure-based protein function annotation. Nucleic Acids Res. 2012, 40, W471–W477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Freddolino, P.L.; Zhang, Y. COFACTOR: Improved protein function prediction by combining structure, sequence and protein-protein interaction information. Nucleic Acids Res. 2017, 45, W291–W299. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; An, T.; Xu, Z.; Zhang, L.; Gao, H.; Sun, W.; Liao, B.; Jiang, C.; Liu, Z.; Duan, L.; et al. Genomic-wide identification and expression analysis of AP2/ERF transcription factors related to andrographolide biosynthesis in Andrographis paniculata. Ind. Crops Prod. 2020, 157, 112878. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, Z.; Zhang, L.; Yao, W.; Xu, Z.; Liao, B.; Duan, L.; Ji, A. Genomic Characterization of WRKY Transcription Factors Related to Andrographolide Biosynthesis in Andrographis paniculata. Front. Genet. 2021, 11, 1823. [Google Scholar] [CrossRef]
- Zhong, Y.; Li, L.; Hao, X.; Fu, X.; Ma, Y.; Xie, L.; Shen, Q.; Kayani, S.; Pan, Q.; Sun, X.; et al. AaABF3, an abscisic acid–Responsive transcription factor, positively regulates artemisinin biosynthesis in artemisia annua. Front. Plant Sci. 2018, 9, 1777. [Google Scholar] [CrossRef]
- Manzoor, M.M.; Goyal, P.; Pandotra, P.; Dar, M.S.; Dar, M.J.; Misra, P.; Gupta, A.P.; Vishwakarma, R.A.; Ahuja, A.; Dhar, M.K.; et al. Transcriptome-wide identification of squalene epoxidase genes from Glycyrrhiza glabra L.: Expression analysis and heterologous expression of GgSQE1 suggest important role in terpenoid biosynthesis. Protoplasma 2021, 258, 991–1007. [Google Scholar] [CrossRef]
- Awasthi, P.; Gupta, A.P.; Bedi, Y.S.; Vishwakarma, R.A.; Gandhi, S.G. Mannitol stress directs flavonoid metabolism toward synthesis of flavones via differential regulation of two cytochrome P450 monooxygenases in coleus forskohlii. Front. Plant Sci. 2016, 7, 208138. [Google Scholar] [CrossRef]
- Manzoor, M.M.; Goyal, P.; Gupta, A.P.; Khan, S.; Jaswal, P.; Misra, P.; Pandotra, P.; Ahuja, A.; Vishwakarma, R.A.; Gupta, S. Chemical and real-time based analysis revealed active gene machinery of glycyrrhizin biosynthesis and its accumulation in the aerial tissues of in-vitro regenerated Glycyrrhiza glabra L. Plant Growth Regul. 2020, 92, 263–271. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jha, Z.; Sharam, S.N.; Sharma, D.K. Differential expression of 3-hydroxy-3-methylglutaryl-coenzyme A reductase of Andrographis paniculata in andrographolide accumulation. J. Chem. Pharm. Res. 2011, 3, 499–504. [Google Scholar]
- Patel, A.A.; Shukla, Y.M.; Kumar, S.; Sakure, A.A.; Parekh, M.J.; Zala, H.N. Transcriptome analysis for molecular landscaping of genes controlling diterpene andrographolide biosynthesis in Andrographis paniculata (Burm. f.) Nees. 3 Biotech 2020, 10, 512. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.N.; Jha, Z.; Sinha, R.K.; Geda, A.K. Jasmonate-induced biosynthesis of andrographolide in Andrographis paniculata. Physiol. Plant 2015, 153, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Srinath, M.; Shailaja, A.; Bindu, B.B.V.; Giri, C.C. Comparative analysis of biomass, ethrel elicitation, light induced differential MVA/MEP pathway gene expression and andrographolide production in adventitious root cultures of Andrographis paniculata (Burm. F.) Nees. Plant Cell Tissue Organ Cult. 2022, 149, 335–349. [Google Scholar] [CrossRef]
- Millar, N. Millar Biology statistics made simple using Excel Biology statistics made simple using Excel Spreadsheet programs such as Microsoft Excel can transform the use of statistics in A-level science. Sch. Sci. Rev. 2001, 83, 23–34. [Google Scholar]
- Weigel, D.; Glazebrook, J. Transformation of agrobacterium using the freeze-thaw method. CSH Protoc. 2006, 7, pdb-prot4666. [Google Scholar] [CrossRef]
- Ratner, B. The correlation coefficient: Its values range between 1/1, or do they. J. Targeting, Meas. Anal. Mark 2009, 17, 139–142. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, J.; Chen, H.; Jin, W.; Liang, Z. Characterization of NAC family genes in Salvia miltiorrhiza and NAC2 potentially involved in the biosynthesis of tanshinones. Phytochemistry 2021, 191, 112932. [Google Scholar] [CrossRef]
- Xu, J.; Xu, H.; Zhao, H.; Liu, H.; Xu, L.; Liang, Z. Genome-wide investigation of bHLH genes and expression analysis under salt and hormonal treatments in Andrographis paniculata. Ind. Crops Prod. 2022, 183, 114928. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Anuradha, V.E.; Jaleel, C.A.; Salem, M.A.; Gomathinayagam, M.; Panneerselvam, R. Plant growth regulators induced changes in antioxidant potential and andrographolide content in Andrographis paniculata Wall. ex Nees. Pestic. Biochem. Physiol. 2010, 98, 312–316. [Google Scholar] [CrossRef]
- Murthy, H.N.; Dalawai, D. Biotechnological production of diterpenoid lactones from cell and organ cultures of Andrographis paniculata. Appl. Microbiol. Biotechnol. 2021, 105, 7683–7694. [Google Scholar] [CrossRef] [PubMed]
- Patil, M.; Ramu, S.V.; Jathish, P.; Sreevathsa, R.; Reddy, P.C.; Prasad, T.G.; Udayakumar, M. Overexpression of AtNAC2 (ANAC092) in groundnut (Arachis hypogaea L.) improves abiotic stress tolerance. Plant Biotechnol. Rep. 2014, 8, 161–169. [Google Scholar] [CrossRef]
- Rathnayake, K.; Garcia, T.M.; Cushman, J.C.; Wone, B.W. A Novel NAC83 Transcription Factor from Kalanchoe fedtschenkoi enhances Drought and Salt Tolerance in Arabidopsis. In Proceedings of the Plant and Animal Genome XXVII 789 Conference, San Diego, CA, USA, 12–15 January 2019; PAG: Hongkong, China, 2019. [Google Scholar]
- Shen, J.; Lv, B.; Luo, L.; He, J.; Mao, C.; Xi, D.; Ming, F. The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef] [PubMed]
- Shinde, H.; Dudhate, A.; Tsugama, D.; Gupta, S.K.; Liu, S.; Takano, T. Pearl millet stress-responsive NAC transcription factor PgNAC21 enhances salinity stress tolerance in Arabidopsis. Plant Physiol. Biochem. 2019, 135, 546–553. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, T.; Guo, S.; Hu, J.; Zhan, Y. Ectopic Expression of AeNAC83, a NAC Transcription Factor from Abelmoschus esculentus, Inhibits Growth and Confers Tolerance to Salt Stress in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 10182. [Google Scholar] [CrossRef]


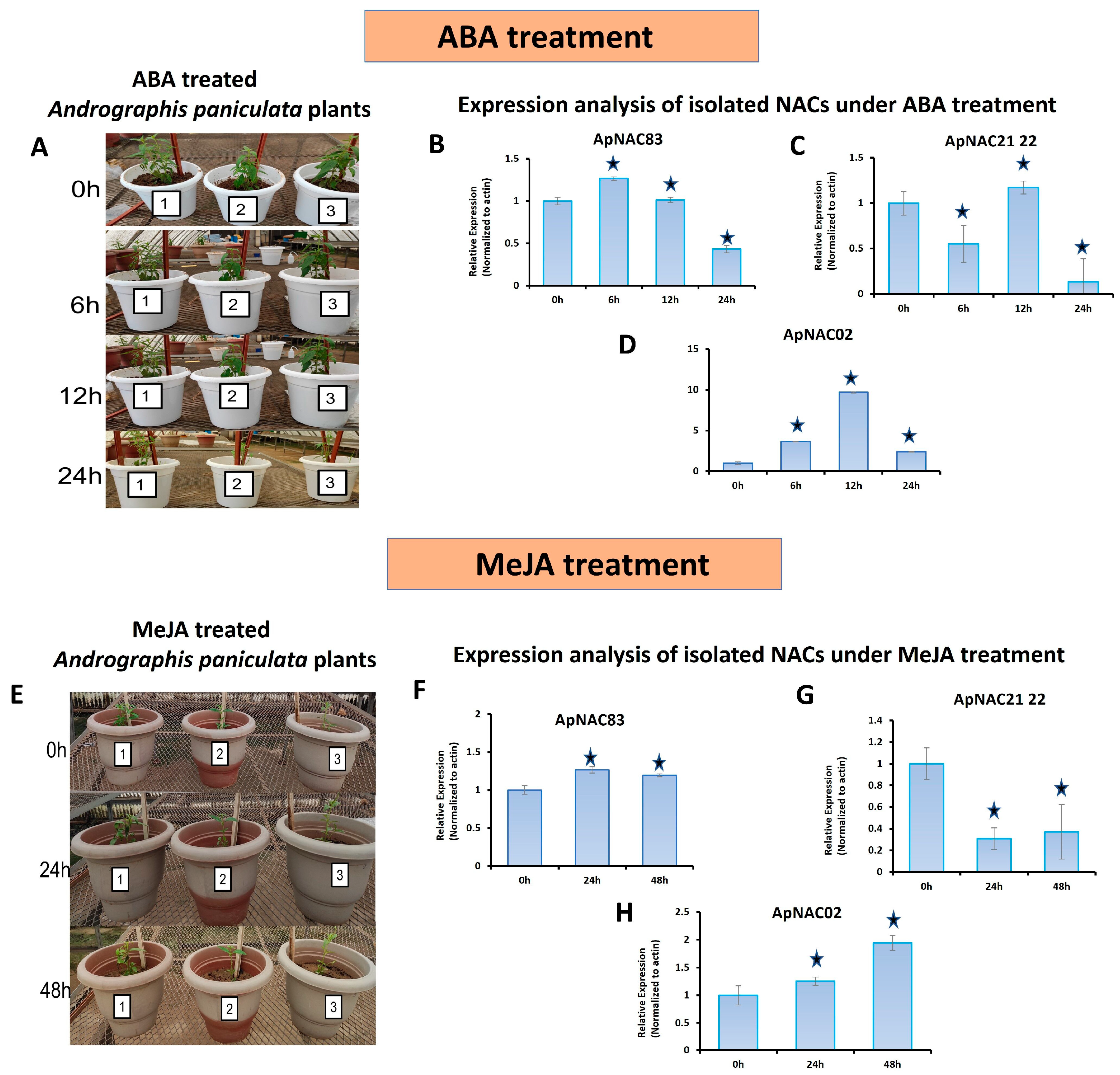
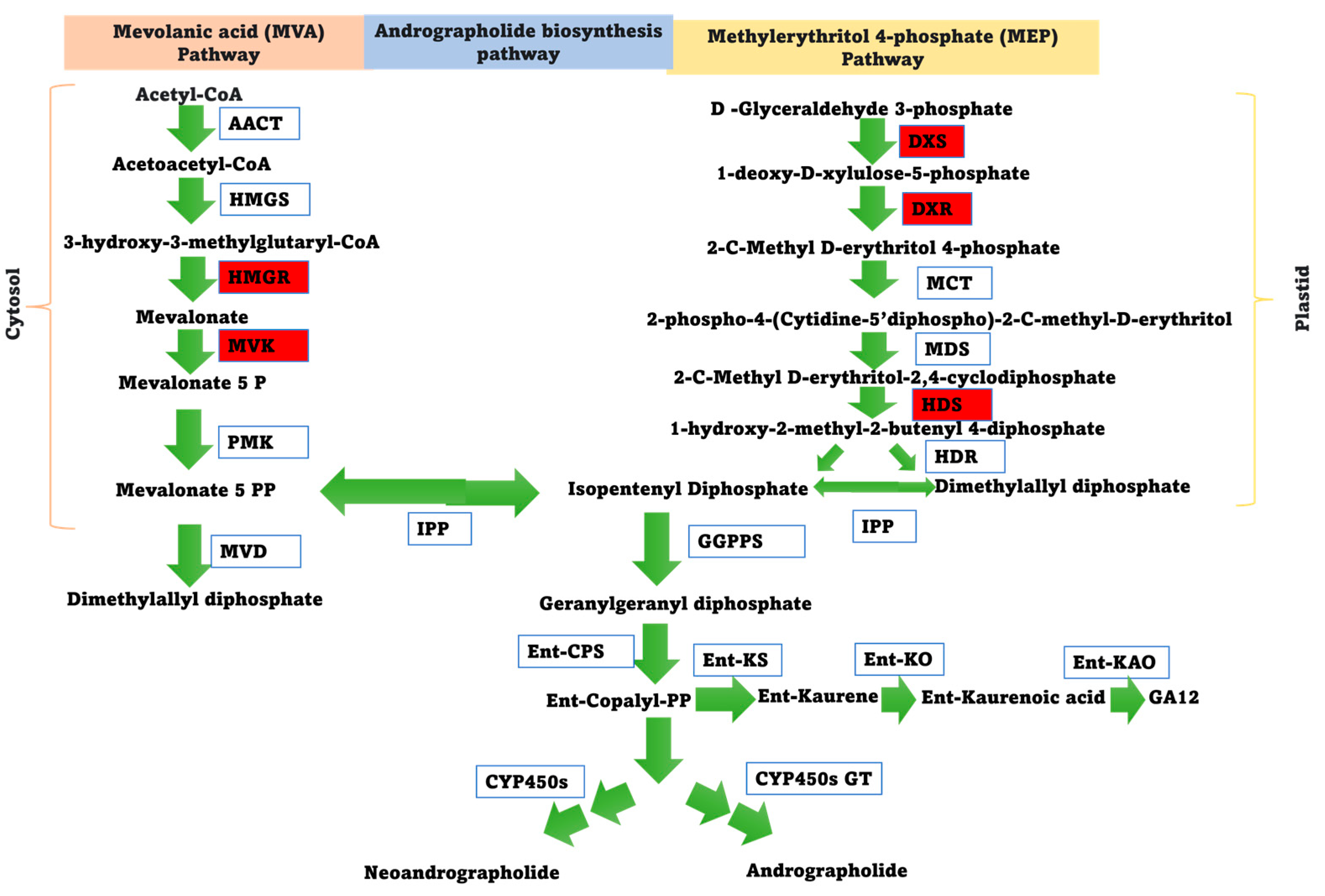
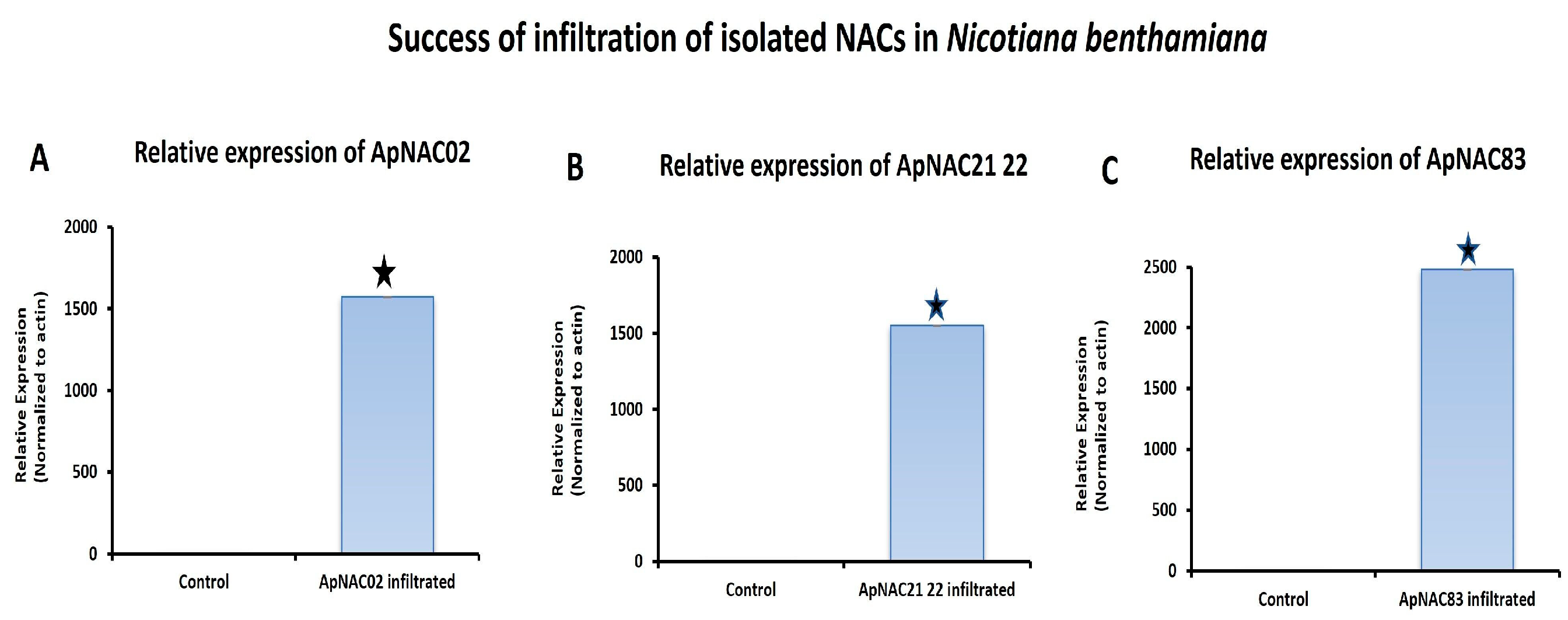
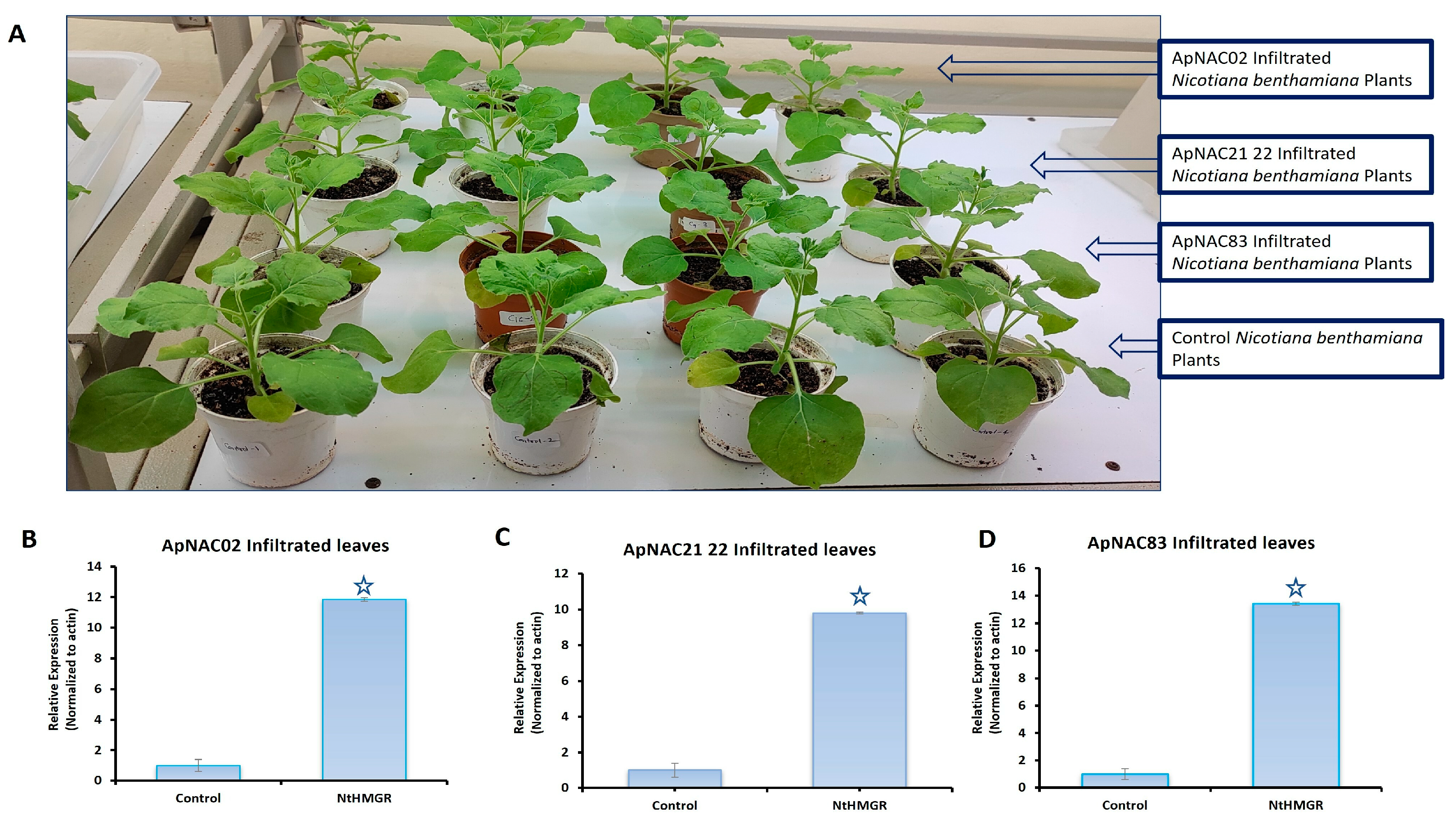

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Bioactive Compounds (ng g−1) | 0 h | 6 h | 12 h | 24 h |
|---|---|---|---|---|---|
| 1 | Andrographolide x | 185.67 | 343.12 | 326.95 | 332.14 |
| 2 | neo andrographolide y | 40.31 | 20.07 | 23.16 | 25.02 |
| 3 | 14-deoxyandrographolide x | 11.39 | 14.41 | 6.41 | 41.02 |
| 4 | andrographolide glycoside y | 1.2 | 1.31 | 1.05 | 1.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, R.; Kumar, C.; Roy Choudhury, D.; Ranjan, A.; Raipuria, R.K.; Dubey, K.K.D.; Mishra, A.; Kumar, C.; Manzoor, M.M.; Kumar, A.; et al. Isolation, Characterization, and Expression Analysis of NAC Transcription Factor from Andrographis paniculata (Burm. f.) Nees and Their Role in Andrographolide Production. Genes 2024, 15, 422. https://doi.org/10.3390/genes15040422
Kumar R, Kumar C, Roy Choudhury D, Ranjan A, Raipuria RK, Dubey KKD, Mishra A, Kumar C, Manzoor MM, Kumar A, et al. Isolation, Characterization, and Expression Analysis of NAC Transcription Factor from Andrographis paniculata (Burm. f.) Nees and Their Role in Andrographolide Production. Genes. 2024; 15(4):422. https://doi.org/10.3390/genes15040422
Chicago/Turabian StyleKumar, Ramesh, Chavlesh Kumar, Debjani Roy Choudhury, Aashish Ranjan, Ritesh Kumar Raipuria, Kaushik Kumar Dhar Dubey, Ayushi Mishra, Chetan Kumar, Malik Muzafar Manzoor, Ashok Kumar, and et al. 2024. "Isolation, Characterization, and Expression Analysis of NAC Transcription Factor from Andrographis paniculata (Burm. f.) Nees and Their Role in Andrographolide Production" Genes 15, no. 4: 422. https://doi.org/10.3390/genes15040422
APA StyleKumar, R., Kumar, C., Roy Choudhury, D., Ranjan, A., Raipuria, R. K., Dubey, K. K. D., Mishra, A., Kumar, C., Manzoor, M. M., Kumar, A., Kumari, A., Singh, K., Singh, G. P., & Singh, R. (2024). Isolation, Characterization, and Expression Analysis of NAC Transcription Factor from Andrographis paniculata (Burm. f.) Nees and Their Role in Andrographolide Production. Genes, 15(4), 422. https://doi.org/10.3390/genes15040422