
PDE4 Gene Family Variants Are Associated with Response to Apremilast Treatment in Psoriasis
 , ,
, ,  , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Samples, Genotyping, Quality Control and Association Analysis
2.3. ncRNA Analysis
2.4. Protein-Network and Pathway Analysis
2.5. Expression Quantitative Trait Loci (eQTL) Analysis
3. Results
3.1. Clinical Characteristics of the Study Population
3.2. PDE4 and CYP3A4 Variants Are Associated with Response to Treatment
3.3. SNPs within ncRNAs That Target PDE4 mRNAs Are Associated with Apremilast Response
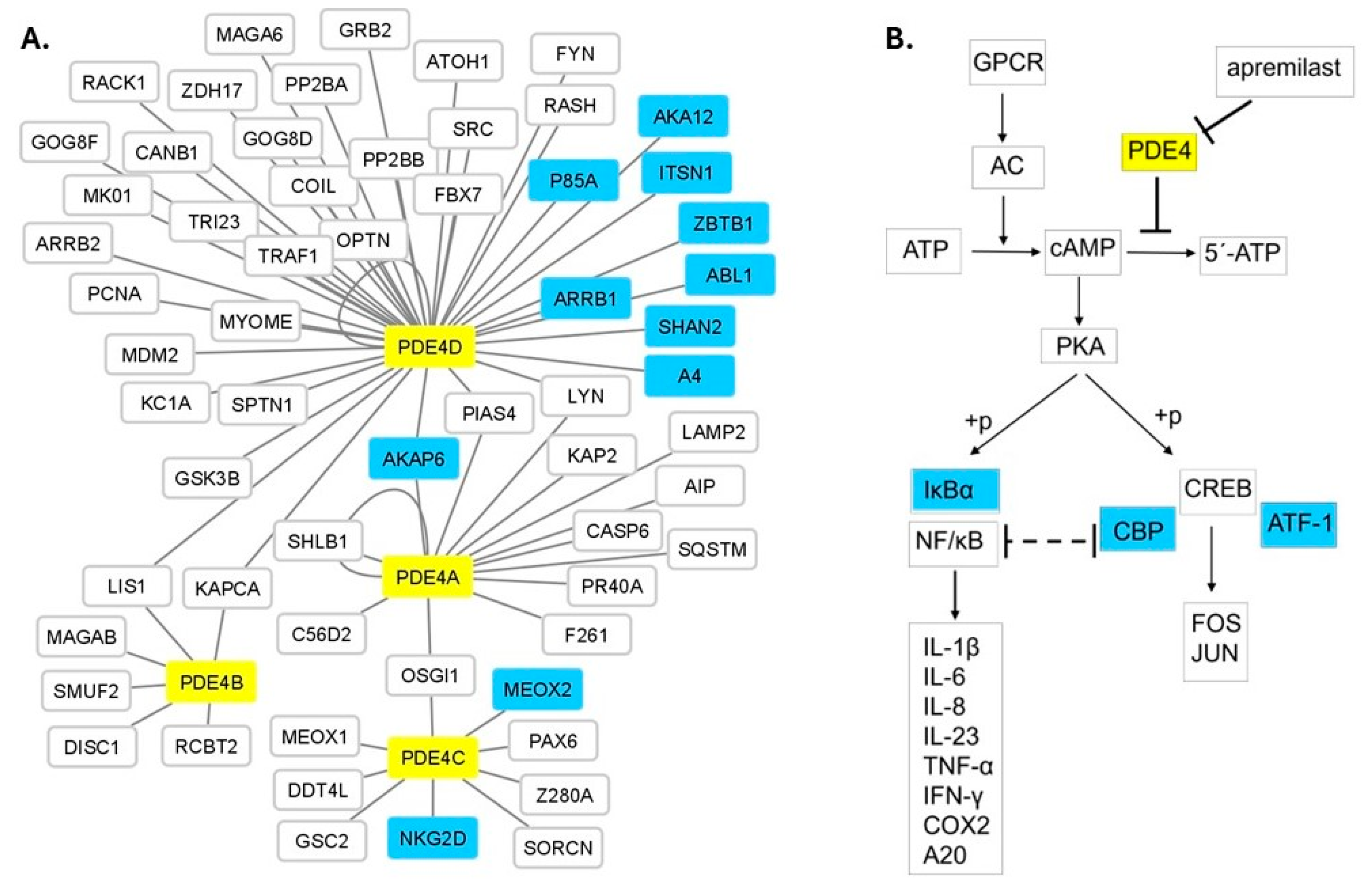
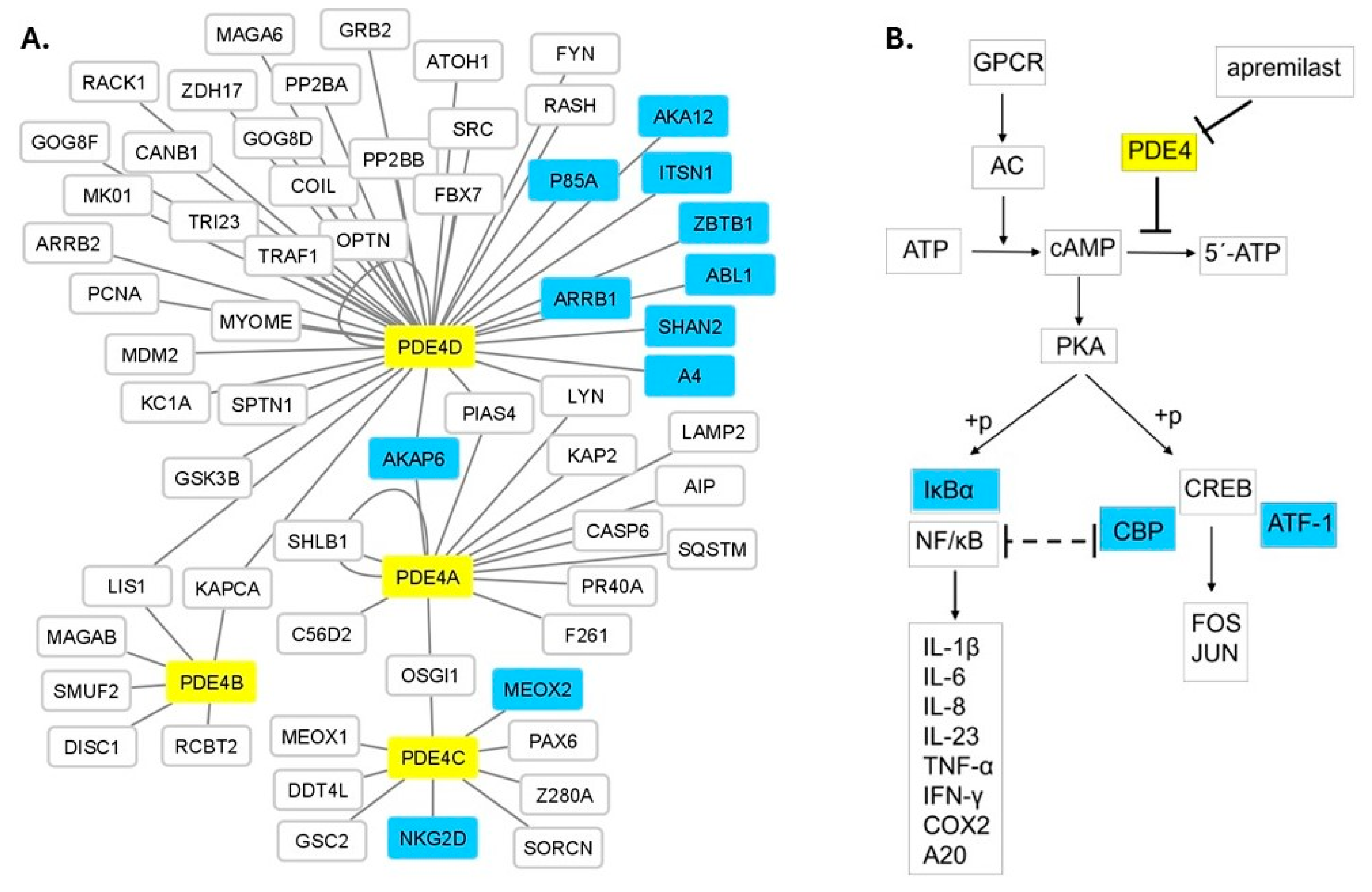
3.4. Protein Network and Pathway Analysis
3.5. eQTL Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boehncke, W.-H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Lee, H.-J.; Kim, M. Challenges and Future Trends in the Treatment of Psoriasis. Int. J. Mol. Sci. 2023, 24, 13313. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Wang, C.-W.; Chen, C.-B.; Chen, W.-T.; Chang, Y.-C.; Hui, R.C.-Y.; Chung, W.-H. Pharmacogenomics on the Treatment Response in Patients with Psoriasis: An Updated Review. Int. J. Mol. Sci. 2023, 24, 7329. [Google Scholar] [CrossRef] [PubMed]
- Ovejero-Benito, M.C.; Muñoz -Aceituno, E.; Reolid, A.; Saiz- Rodríguez, M.; Abad-Santos, F.; Daudén, E. Pharmacogenetics and Pharmacogenomics in Moderate-to-Severe Psoriasis. Am. J. Clin. Dermatol. 2018, 19, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Owczarczyk-Saczonek, A.; Owczarek, W.; Osmola-Mańkowska, A.; Adamski, Z.; Placek, W.; Rakowska, A. Secondary failure of TNF-α inhibitors in clinical practice. Dermatol. Ther. 2019, 32, e12760. [Google Scholar] [CrossRef] [PubMed]
- Elberdín, L.; Fernández-Torres, R.M.; Paradela, S.; Mateos, M.; Blanco, E.; Balboa-Barreiro, V.; Gómez-Besteiro, M.I.; Outeda, M.; Martín-Herranz, I.; Fonseca, E. Biologic Therapy for Moderate to Severe Psoriasis. Real-World Follow-up of Patients Who Initiated Biologic Therapy at Least 10 Years Ago. Dermatol. Ther. 2022, 12, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Nassim, D.; Alajmi, A.; Jfri, A.; Pehr, K. Apremilast in dermatology: A review of literature. Dermatol. Ther. 2020, 33, e14261. [Google Scholar] [CrossRef] [PubMed]
- Chiricozzi, A.; Caposiena, D.; Garofalo, V.; Cannizzaro, M.V.; Chimenti, S.; Saraceno, R. A new therapeutic for the treatment of moderate-to-severe plaque psoriasis: Apremilast. Expert Rev. Clin. Immunol. 2016, 12, 237–249. [Google Scholar] [CrossRef]
- Johnson, K.R.; Nicodemus-Johnson, J.; Danziger, R.S. An evolutionary analysis of cAMP-specific Phosphodiesterase 4 alternative splicing. BMC Evol. Biol. 2010, 10, 247. [Google Scholar] [CrossRef]
- Schafer, P.H.; Parton, A.; Capone, L.; Cedzik, D.; Brady, H.; Evans, J.F.; Man, H.-W.; Muller, G.W.; Stirling, D.I.; Chopra, R. Apremilast is a selective PDE4 inhibitor with regulatory effects on innate immunity. Cell Signal. 2014, 26, 2016–2029. [Google Scholar] [CrossRef]
- Keating, G.M. Apremilast: A Review in Psoriasis and Psoriatic Arthritis. Drugs 2017, 77, 459–472. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Papp, K.; Reich, K.; Leonardi, C.L.; Kircik, L.; Chimenti, S.; Langley, R.G.; Hu, C.; Stevens, R.M.; Day, R.M.; Gordon, K.B.; et al. Apremilast, an oral phosphodiesterase 4 (PDE4) inhibitor, in patients with moderate to severe plaque psoriasis: Results of a phase III, randomized, controlled trial (Efficacy and Safety Trial Evaluating the Effects of Apremilast in Psoriasis [ESTEEM] 1). J. Am. Acad. Dermatol. 2015, 73, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Cather, J.; Gooderham, M.; Poulin, Y.; Mrowietz, U.; Ferrandiz, C.; Crowley, J.; Hu, C.; Stevens, R.M.; Shah, K.; et al. Efficacy and safety of apremilast, an oral phosphodiesterase 4 inhibitor, in patients with moderate-to-severe plaque psoriasis over 52 weeks: A phase III, randomized controlled trial (ESTEEM 2). Br. J. Dermatol. 2015, 173, 1387–1399. [Google Scholar] [CrossRef] [PubMed]
- Reich, K.; Gooderham, M.; Bewley, A.; Green, L.; Soung, J.; Petric, R.; Marcsisin, J.; Cirulli, J.; Chen, R.; Piguet, V. Safety and efficacy of apremilast through 104 weeks in patients with moderate to severe psoriasis who continued on apremilast or switched from etanercept treatment: Findings from the LIBERATE study. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 397–402. [Google Scholar] [CrossRef]
- Wong, T.H.; Sinclair, S.; Smith, B.; Fraser, C.; Morton, C.A. Real-world, single-centre experience of apremilast for the treatment of moderate to severe psoriasis. Clin. Exp. Dermatol. 2017, 42, 674–685. [Google Scholar] [CrossRef]
- Papadavid, E.; Rompoti, N.; Theodoropoulos, K.; Kokkalis, G.; Rigopoulos, D. Real-world data on the effectiveness and safety of apremilast in patients with moderate-to-severe plaque psoriasis. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1173–1179. [Google Scholar] [CrossRef]
- Ighani, A.; Georgakopoulos, J.R.; Zhou, L.L.; Walsh, S.; Shear, N.; Yeung, J. Efficacy and safety of apremilast monotherapy for moderate to severe psoriasis: Retrospective study. J. Cutan. Med. Surg. 2018, 22, 290–296. [Google Scholar] [CrossRef]
- Verbenko, D.A.; Karamova, A.E.; Artamonova, O.G.; Deryabin, D.G.; Rakitko, A.; Chernitsov, A.; Krasnenko, A.; Elmuratov, A.; Solomka, V.S.; Kubanov, A.A. Apremilast Pharmacogenomics in Russian Patients with Moderate-to-Severe and Severe Psoriasis. J. Pers. Med. 2020, 11, 20. [Google Scholar] [CrossRef]
- Martin, F.J.; Amode, M.R.; Aneja, A.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Becker, A.; Bennett, R.; Berry, A.; Bhai, J.; et al. Ensembl 2023. Nucleic Acids Res. 2023, 51, D933–D941. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, P.; Tian, R.; Wang, S.; Guo, Q.; Luo, M.; Zhou, W.; Liu, G.; Jiang, H.; Jiang, Q. LncRNA2Target v2.0: A comprehensive database for target genes of lncRNAs in human and mouse. Nucleic Acids Res. 2019, 47, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Lin, Y.-C.-D.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. miRTarBase update 2022: An informative resource for experimentally validated miRNA–target interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef]
- Dimitrakopoulos, G.N.; Klapa, M.I.; Moschonas, N.K. PICKLE 3.0: Enriching the human meta-database with the mouse protein interactome extended via mouse–human orthology. Bioinformatics 2020, 37, 145–146. [Google Scholar] [CrossRef] [PubMed]
- Dimitrakopoulos, G.N.; Klapa, M.I.; Moschonas, N.K. How Far Are We from the Completion of the Human Protein Interactome Reconstruction? Biomolecules 2022, 12, 140. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Berna-Rico, E.; Perez-Bootello, J.; Abbad-Jaime de Aragon, C.; Gonzalez-Cantero, A. Genetic Influence on Treatment Response in Psoriasis: New Insights into Personalized Medicine. Int. J. Mol. Sci. 2023, 24, 9850. [Google Scholar] [CrossRef] [PubMed]
- Morelli, M.; Galluzzo, M.; Madonna, S.; Scarponi, C.; Scaglione, G.L.; Galluccio, T.; Andreani, M.; Pallotta, S.; Girolomoni, G.; Bianchi, L.; et al. HLA-Cw6 and other HLA-C alleles, as well as MICBDT, DDX58, and TYK2 genetic variants associate with optimal response to anti-IL-17A treatment in patients with psoriasis. Expert Opin. Biol. Ther. 2021, 21, 259–270. [Google Scholar] [CrossRef]
- Stuart, P.E.; Nair, R.P.; Tsoi, L.C.; Tejasvi, T.; Das, S.; Kang, H.M.; Ellinghaus, E.; Chandran, V.; Callis-Duffin, K.; Ike, R.; et al. Genome-wide Association Analysis of Psoriatic Arthritis and Cutaneous Psoriasis Reveals Differences in Their Genetic Architecture. Am. J. Hum. Genet. 2015, 97, 816–836. [Google Scholar] [CrossRef]
- Bowes, J.; Budu-Aggrey, A.; Huffmeier, U.; Uebe, S.; Steel, K.; Hebert, H.L.; Wallace, C.; Massey, J.; Bruce, I.N.; Bluett, J.; et al. Dense genotyping of immune-related susceptibility loci reveals new insights into the genetics of psoriatic arthritis. Nat. Commun. 2015, 6, 6046. [Google Scholar] [CrossRef]
- Yin, X.; Low, H.Q.; Wang, L.; Li, Y.; Ellinghaus, E.; Han, J.; Estivill, X.; Sun, L.; Zuo, X.; Shen, C.; et al. Genome-wide meta-analysis identifies multiple novel associations and ethnic heterogeneity of psoriasis susceptibility. Nat. Commun. 2015, 6, 6916. [Google Scholar] [CrossRef]
- Allanore, Y.; Saad, M.; Dieudé, P.; Avouac, J.; Distler, J.H.; Amouyel, P.; Matucci-Cerinic, M.; Riemekasten, G.; Airo, P.; Melchers, I.; et al. Genome-wide scan identifies TNIP1, PSORS1C1, and RHOB as novel risk loci for systemic sclerosis. PLoS Genet. 2011, 7, e1002091. [Google Scholar] [CrossRef]
- Liu, J.Z.; van Sommeren, S.; Huang, H.; Ng, S.C.; Alberts, R.; Takahashi, A.; Ripke, S.; Lee, J.C.; Jostins, L.; Shah, T.; et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat. Genet. 2015, 47, 979–986. [Google Scholar] [CrossRef]
- Miyagawa, T.; Tokunaga, K. Genetics of narcolepsy. Hum. Genome Var. 2019, 6, 4. [Google Scholar] [CrossRef]
- Wang, H.; Yang, J.; Schneider, J.A.; De Jager, P.L.; Bennett, D.A.; Zhang, H.-Y. Genome-wide interaction analysis of pathological hallmarks in Alzheimer’s disease. Neurobiol. Aging 2020, 93, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Chavez, R.A.; Munro, J.E.; Chiaroni-Clarke, R.C.; Akikusa, J.D.; Allen, R.C.; Craig, J.M.; Ponsonby, A.-L.; Saffery, R.; Ellis, J.A. DNA methylation at IL32 in juvenile idiopathic arthritis. Sci. Rep. 2015, 5, 11063. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Nagappa, M.; Sreekumaran Nair, B.V.; Das, S.K.; Wahatule, R.; Sinha, S.; Vasanthapuram, R.; Taly, A.B.; Debnath, M. Variations within Toll-like receptor (TLR) and TLR signaling pathway-related genes and their synergistic effects on the risk of Guillain-Barré syndrome. J. Peripher. Nerv. Syst. 2022, 27, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Yenmis, G.; Oner, T.; Cam, C.; Koc, A.; Kucuk, O.S.; Yakicier, M.C.; Dizman, D.; Kanigur Sultuybek, G. Association of NFKB1 and NFKBIA polymorphisms in relation to susceptibility of Behçet’s disease. Scand. J. Immunol. 2015, 81, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Bank, S.; Julsgaard, M.; Abed, O.K.; Burisch, J.; Broder Brodersen, J.; Pedersen, N.K.; Gouliaev, A.; Ajan, R.; Nytoft Rasmussen, D.; Honore Grauslund, C.; et al. Polymorphisms in the NFkB, TNF-alpha, IL-1beta, and IL-18 pathways are associated with response to anti-TNF therapy in Danish patients with inflammatory bowel disease. Aliment. Pharmacol. Ther. 2019, 49, 890–903. [Google Scholar] [CrossRef] [PubMed]
- Rezazadeh, M.; Gharesouran, J.; Moradi, M.; Noroozi, R.; Omrani, M.D.; Taheri, M.; Ghafouri-Fard, S. Association study of ANRIL genetic variants and multiple sclerosis. J. Mol. Neurosci. 2018, 65, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Rakhshan, A.; Zarrinpour, N.; Moradi, A.; Ahadi, M.; Omrani, M.D.; Ghafouri-Fard, S.; Taheri, M. Genetic variants within ANRIL (antisense non-coding RNA in the INK4 locus) are associated with risk of psoriasis. Int. Immunopharmacol. 2020, 78, 106053. [Google Scholar] [CrossRef]
- Hao, S.; Zhou, K.; Yu, P.; Tian, J.; Zhang, Y.; Dang, L.; Tong, N.; Li, Y. ANRIL polymorphisms in psoriasis vulgaris patients in northern China. Eur. J. Dermatol. 2022, 32, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Bochenek, G.; Häsler, R.; El Mokhtari, N.E.; König, I.R.; Loos, B.G.; Jepsen, S.; Rosenstiel, P.; Schreiber, S.; Schaefer, A.S. The large non-coding RNA ANRIL, which is associated with atherosclerosis, periodontitis and several forms of cancer, regulates ADIPOR1, VAMP3 and C11ORF10. Hum. Mol. Genet. 2013, 22, 4516–4527. [Google Scholar] [CrossRef]
- Ziegler, C.; Graf, J.; Faderl, S.; Schedlbauer, J.; Strieder, N.; Förstl, B.; Spang, R.; Bruckmann, A.; Merkl, R.; Hombach, S.; et al. The long non-coding RNA LINC00941 and SPRR5 are novel regulators of human epidermal homeostasis. EMBO Rep. 2019, 20, e46612. [Google Scholar] [CrossRef]
- Adylova, A.; Mukhanbetzhanovna, A.A.; Attar, R.; Yulaevna, I.M.; Farooqi, A.A. Regulation of TGFβ/SMAD signaling by long non-coding RNAs in different cancers: Dark Knight in the Castle of molecular oncology. Noncoding RNA Res. 2021, 6, 23–28. [Google Scholar] [CrossRef]
- Dodge-Kafka, K.L.; Soughayer, J.; Pare, G.C.; Carlisle Michel, J.J.; Langeberg, L.K.; Kapiloff, M.S.; Scott, J.D. The protein kinase A anchoring protein mAKAP coordinates two integrated cAMP effector pathways. Nature 2005, 437, 574–578. [Google Scholar] [CrossRef]
- Mauban, J.R.H.; O’ Donnell, M.; Warrier, S.; Manni, S.; Bond, M. AKAP-scaffolding proteins and regulation of cardiac physiology. Physiology 2009, 24, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Qasim, H.; McConnell, B.K. AKAP12 Signaling Complex: Impacts of Compartmentalizing cAMP-Dependent Signaling Pathways in the Heart and Various Signaling Systems. J. Am. Heart Assoc. 2020, 9, e016615. [Google Scholar] [CrossRef] [PubMed]
- Baillie, G.S.; Scott, J.D.; Houslay, M.D. Compartmentalisation of phosphodiesterases and protein kinase A: Opposites attract. FEBS Lett. 2005, 579, 3264–3270. [Google Scholar] [CrossRef]
- Liu, Q.; Yao, F.; Wang, M.; Zhou, B.; Cheng, H.; Wang, W.; Jin, L.; Lin, Q.; Wang, J.-C. Novel human BTB/POZ domain-containing zinc finger protein ZBTB1 inhibits transcriptional activities of CRE. Mol. Cell Biochem. 2011, 357, 405–414. [Google Scholar] [CrossRef]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The role of the transcription factor CREB in immune function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [PubMed]
- Servillo, G.; Della Fazia, M.A.; Sassone-Corsi, P. Coupling cAMP signaling to transcription in the liver: Pivotal role of CREB and CREM. Exp. Cell Res. 2002, 275, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, D.U.; Komives, E.A. Molecular mechanisms of system control of NF-kappaB signaling by IkappaBalpha. Biochemistry 2010, 49, 1560–1567. [Google Scholar] [CrossRef]
- Wang, D.; Sadee, W. CYP3A4 intronic SNP rs35599367 (CYP3A4*22) alters RNA splicing. Pharmacogenet. Genom. 2016, 26, 40–43. [Google Scholar] [CrossRef]
- Wang, D.; Guo, Y.; Wrighton, S.A.; Cooke, G.E.; Sadee, W. Intronic polymorphism in CYP3A4 affects hepatic expression and response to statin drugs. Pharmacogenom. J. 2011, 11, 274–286. [Google Scholar] [CrossRef]
- Reich, K.; Mrowietz, U.; Menter, A.; Griffiths, C.E.M.; Bagel, J.; Strober, B.; Nunez Gomez, N.; Shi, R.; Guerette, B.; Lebwohl, M. Effect of baseline disease severity on achievement of treatment target with apremilast: Results from a pooled analysis. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 2409–2414. [Google Scholar] [CrossRef]
- Wolf, P. Systemic antipsoriatic treatment: Do women respond better than men and if so, why? Br. J. Dermatol. 2021, 185, 1088–1089. [Google Scholar] [CrossRef] [PubMed]
- Guillet, C.; Seeli, C.; Nina, M.; Maul, L.V.; Maul, J.-T. The impact of gender and sex in psoriasis: What to be aware of when treating women with psoriasis. Int. J. Womens Dermatol. 2022, 8, e010. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| All Patients (n = 49) | Responders (n = 22) | Non-Responders (n = 27) | p Value | |
|---|---|---|---|---|
| Male/Female (M/F) | 32/17 | 13/9 | 19/8 | 0.601 |
| Mean baseline age (mean ± s.d.) | 55.4 (±15) | 54.8 (±13.2) | 55.9 (±16.4) | 0.954 |
| Disease duration (years) | 18.4 (±13.1) | 17.1 (±10.5) | 19.2 (±14.6) | 0.634 |
| Baseline weight (kg), M/F | 92.8 (±19.4), 98/83.2 | 90.5 (±20.3), 95.9/82.4 | 94.7 (±18.9), 93.3/84.1 | 0.469 |
| Baseline BMI | 31.6 (±5.7) | 32.3 (±7.3) | 31.1 (±4.3) | 0.513 |
| Baseline PASI | 10.3 (±6.6) | 8.5 (±3.2) | 11.8 (±8.3) | 0.036 |
| PASI at 6 months | 6.1 (±6.4) | 1.7 (±1.4) | 9.8 (±6.6) | 0.003 |
| PASI at 9 months | 4.7 (±4.5) | 1.3 (±0.9) | 7.6 (±4.2) | 0.007 |
| Locus | SNP | Location, HGVS Name | F_R | F_NR | MAF R + NR | MAF (EUR) | p Value | OR |
|---|---|---|---|---|---|---|---|---|
| u. PDE4A | rs12979813 (A/G) | Intergenic, chr19, NC_000019.10:g.11232027 | 0.4545 | 0.2593 | 0.35 | 0.14 | 4.3 × 10−2 | 0.42 |
| rs2305795 * (A/G) | Intergenic, chr19, NC_000019.10:g.10115376 | 0.5909 | 0.3889 | 0.48 | 0.40 | 4.6 × 10−2 | 0.44 | |
| d. PDE4A | rs76966440 (C/T) | Intergenic, chr19, NC_000019.10:g.10687087 | 0.1136 | 0 | 0.05 | 0.02 | 1.1 × 10−2 | 0 |
| rs892085 * (A/G) | Intergenic, chr19, NC_000019.10:g.10707416 | 0.6136 | 0.3889 | 0.49 | 0.48 | 2.7 × 10−2 | 0.40 | |
| rs74942924 (G/A) | Intergenic, chr19, NC_000019.10:g.10952989 | 0 | 0.0926 | 0.05 | 0.03 | 3.8 × 10−2 | NA | |
| rs11085752 * (A/C) | Intergenic, chr19, NC_000019.10:g.10954733 | 0.6136 | 0.3889 | 0.49 | 0.42 | 2.7 × 10−2 | 0.40 | |
| rs7935 (T/C) | Intergenic, chr19, NC_000019.10:g.10994932 | 0.5227 | 0.3148 | 0.41 | 0.35 | 3.7 × 10−2 | 0.42 | |
| rs12980863 (C/T) | Intergenic, chr19, NC_000019.10:g.11199195 | 0.2955 | 0.5000 | 0.41 | 0.39 | 4 × 10−2 | 2.39 | |
| rs12979813 (A/G) | Intergenic, chr19, NC_000019.10:g.11232027 | 0.0682 | 0.2222 | 0.15 | 0.14 | 3.5 × 10−2 | 3.91 | |
| rs420703 (C/A) | Intergenic, chr19, NC_000019.10:g.11301192 | 0.3409 | 0.5741 | 0.47 | 0.30 | 2.1 × 10−2 | 2.61 | |
| rs322151 * (C/T) | Intergenic, chr19, NC_000019.10:g.11306524 | 0.2727 | 0.4815 | 0.39 | 0.23 | 3.5 × 10−2 | 2.48 | |
| rs322144 (G/C) | Intergenic, chr19, NC_000019.10:g.11313027 | 0.3409 | 0.6296 | 0.50 | 0.56 | 4 × 10−3 | 3.29 | |
| rs10424035 (G/A) | Intergenic, chr19, NC_000019.10:g.11400322 | 0 | 0.0926 | 0.05 | 0.01 | 3.8 × 10−2 | NA | |
| u. PDE4B | rs883824 (G/A) | Intergenic, chr1, NC_000001.11:g.64830174 | 0.0455 | 0.1852 | 0.12 | 0.18 | 3.6 × 10−2 | 4.77 |
| rs1045895 (G/A) | Intergenic, chr1, NC_000001.11:g.65432298 | 0.3409 | 0.5556 | 0.46 | 0.40 | 3.4 × 10−2 | 2.42 | |
| rs1045895 (G/A) dom † | 10 GA + AA/12 GG | 21 GA + AA/6 GG | 2 × 10−2 | |||||
| PDE4B | rs61799396 (C/T) | Intron, chr1, NC_000001.11:g.65956761 | 0.3409 | 0.1667 | 0.24 | 0.25 | 4.6 × 10−2 | 0.39 |
| rs1937457 (C/T) | Intron, chr1, NC_000001.11:g.65981104 | 0.4318 | 0.2222 | 0.32 | 0.29 | 2.6 × 10−2 | 0.38 | |
| rs12757542 (A/G) | Intron, chr1, NC_000001.11:g.66030425 | 0.3864 | 0.1667 | 0.27 | 0.28 | 1.4 × 10−2 | 0.32 | |
| rs12406476 (T/C) | Intron, chr1, NC_000001.11:g.66090109 | 0.3409 | 0.5556 | 0.46 | 0.52 | 3.4 × 10−2 | 2.42 | |
| rs12745871 (C/T) | Intron, chr1, NC_000001.11:g.66121693 | 0.6136 | 0.3148 | 0.45 | 0.34 | 3 × 10−3 | 0.29 | |
| rs2503174 (A/G) | Intron, chr1, NC_000001.11:g.66143471 | 0.0909 | 0.2593 | 0.18 | 0.32 | 3.2 × 10−2 | 3.50 | |
| rs2485381 (A/G) | Intron, chr1, NC_000001.11:g.66162049 | 0.0227 | 0.1481 | 0.09 | 0.17 | 3.2 × 10−2 | 7.48 | |
| rs1890196 (T/C) | Intron, chr 1, NC_000001.11:g.66337397 | 0.3182 | 0.5741 | 0.46 | 0.48 | 1.1 × 10−2 | 2.89 | |
| d. PDE4B | rs11208847 (T/C) | Intergenic, chr1, NC_000001.11:g.66400358 | 0.0909 | 0.2593 | 0.18 | 0.22 | 3.2 × 10−2 | 3.50 |
| rs12118088 (A/G) | Intergenic, chr1, NC_000001.11:g.66479505 | 0 | 0.2037 | 0.11 | 0.09 | 1 × 10−3 | NA | |
| rs765685 (G/A) | Intergenic, chr1, NC_000001.11:g.66499792 | 0.5455 | 0.3333 | 0.43 | 0.48 | 3.5 × 10−2 | 0.42 | |
| rs11809759 (A/G) | Intergenic, chr1, NC_000001.11:g.66804222 | 0.4773 | 0.2778 | 0.37 | 0.41 | 4.2 × 10−2 | 0.42 | |
| rs2815378 (C/T) | Intergenic, chr1, NC_000001.11:g.67044015 | 0.0682 | 0.3148 | 0.20 | 0.30 | 3 × 10−3 | 6.28 | |
| rs77040148 (A/G) | Intergenic, chr1, NC_000001.11:g.67138150 | 0 | 0.1296 | 0.07 | 0.05 | 1.3 × 10−2 | NA | |
| d. PDE4C | rs62128111 (C/T) | Intergenic, chr19, NC_000019.10:g.17352405 | 0.1591 | 0.4074 | 0.30 | 0.27 | 7 × 10−3 | 3.63 |
| rs12972417 (C/T) | Intergenic, chr19, NC_000019.10:g.17414505 | 0.0909 | 0.2778 | 0.19 | 0.22 | 2 × 10−2 | 3.85 | |
| rs10410487 (C/T) | Intergenic, chr19, NC_000019.10:g.17718799 | 0.3636 | 0.5741 | 0.48 | 0.43 | 3.8 × 10−2 | 2.36 | |
| rs12151113 (T/C) | Intergenic, chr19, NC_000019.10:g.17748561 | 0.5909 | 0.3333 | 0.45 | 0.51 | 1.1 × 10−2 | 0.35 | |
| rs2161489 (C/T) | Intergenic, chr19, NC_000019.10:g.17753643 | 0 | 0.1296 | 0.07 | 0.07 | 1.3 × 10−2 | NA | |
| rs8108865 (C/T) | Intergenic, chr19, NC_000019.10:g.17761260 | 0.2500 | 0.0556 | 0.14 | 0.19 | 6 × 10−3 | 0.18 | |
| rs4808096 (A/G) | Intergenic, chr19, NC_000019.10:g.17771371 | 0 | 0.0926 | 0.05 | 0.05 | 3.8 × 10−2 | NA | |
| rs897753 (A/C) | Intergenic, chr19, NC_000019.10:g.18043766 | 0.3409 | 0.1481 | 0.23 | 0.20 | 2.5 × 10−2 | 0.34 | |
| u. PDE4C | rs4808781 (G/A) | Intergenic, chr 19, NC_000019.10:g.18306162 | 0.0682 | 0.3333 | 0.21 | 0.23 | 1 × 10−3 | 6.83 |
| rs76580597 (C/T) | Intergenic, chr 19, NC_000019.10:g.18317822 | 0 | 0.0926 | 0.05 | 0.05 | 3.8 × 10−2 | NA | |
| rs10423674 (C/A) | Intergenic, chr 19, NC_000019.10:g.18707093 | 0.4773 | 0.2778 | 0.37 | 0.32 | 4.2 × 10−2 | 0.42 | |
| rs2023878 (C/T) | Intergenic, chr 19, NC_000019.10:g.18723314 | 0.3182 | 0.1481 | 0.22 | 0.21 | 4.5 × 10−2 | 0.37 | |
| rs4808907 (T/C) | Intergenic, chr 19, NC_000019.10:g.19025732 | 0.1818 | 0.4074 | 0.31 | 0.23 | 1.6 × 10−2 | 3.09 | |
| d. PDE4D | rs11960699 (A/C) | Intergenic, chr 5, NC_000005.10:g.58013923 | 0.2955 | 0.5185 | 0.42 | 0.40 | 2.6 × 10−2 | 2.57 |
| rs11741819 (T/C) | Intergenic, chr 5, NC_000005.10:g.58014172 | 0.1364 | 0.3333 | 0.24 | 0.23 | 2.4 × 10−2 | 3.17 | |
| rs72753331 (G/A) | Intergenic, chr 5, NC_000005.10:g.58044825 | 0.0682 | 0.2222 | 0.15 | 0.07 | 3.5 × 10−2 | 3.91 | |
| rs16887824 (C/T) | Intergenic, chr 5, NC_000005.10:g.58109683 | 0.3409 | 0.1111 | 0.21 | 0.18 | 6 × 10−3 | 0.24 | |
| rs9283718 (G/A) | Intergenic, chr 5, NC_000005.10:g.58115743 | 0.3182 | 0.5185 | 0.43 | 0.51 | 4.6 × 10−2 | 2.31 | |
| rs11742503 (A/G) | Intergenic, chr 5, NC_000005.10:g.58234354 | 0.3182 | 0.1481 | 0.22 | 0.18 | 4.5 × 10−2 | 0.37 | |
| rs115791568 (A/G) | Intergenic, chr 5, NC_000005.10:g.58742066 | 0.1136 | 0 | 0.05 | 0.04 | 1.1 × 10−2 | 0 | |
| rs963443 (G/A) | Intergenic, chr 5, NC_000005.10:g.58775972 | 0.3409 | 0.5926 | 0.48 | 0.41 | 1.3 × 10−2 | 2.81 | |
| PDE4D | rs78103527 (T/C) | Intron, chr 5, NC_000005.10:g.59381162 | 0 | 0.0926 | 0.05 | 0.02 | 3.8 × 10−2 | NA |
| rs697076 (C/T) | Intron, chr 5, NC_000005.10:g.59555985 | 0.1364 | 0.3519 | 0.26 | 0.23 | 1.5 × 10−2 | 3.44 | |
| rs295943 (C/T) | Intron, chr 5, NC_000005.10:g.59591047 | 0 | 0.1481 | 0.08 | 0.11 | 8 × 10−3 | NA | |
| rs177077 (C/T) | Intron, chr 5, NC_000005.10:g.59643033 | 0.0909 | 0.2778 | 0.19 | 0.24 | 2 × 10−2 | 3.85 | |
| rs16890078 (T/C) | Intron, chr 5, NC_000005.10:g.59661496 | 0.1364 | 0.0185 | 0.07 | 0.08 | 2.4 × 10−2 | 0.12 | |
| rs2963821 (A/C) | Intron, chr 5, NC_000005.10:g.59813075 | 0.3182 | 0.5185 | 0.43 | 0.49 | 4.6 × 10−2 | 2.31 | |
| d. CYP3A4 | rs73159483 (G/A) | Intergenic, chr 7, NC_000007.14:g.98806461 | 0 | 0.0926 | 0.05 | 0.03 | 3.8 × 10−2 | NA |
| rs1203844 (T/C) | Intergenic, chr 7, NC_000007.14:g.98850871 | 0.5000 | 0.2407 | 0.36 | 0.24 | 8 × 10−3 | 0.32 | |
| rs2395022 * (C/A) | Intergenic, chr 7, NC_000007.14:g.99152756 | 0.1136 | 0 | 0.05 | 0.06 | 1.1 × 10−2 | 0 | |
| CYP3A4 | rs35599367 (C/T) | Intron, chr 7, NC_000007.14:g.99768693 | 0.1136 | 0.0185 | 0.06 | 0.05 | 5 × 10−2 | 0.15 |
| u. CYP3A4 | rs472660 (G/A) | Intergenic, chr 7, NC_000007.14:g.99862484 | 0.1591 | 0.0370 | 0.09 | 0.12 | 3.7 × 10−2 | 0.20 |
| rs7786877 (A/G) | Intergenic, chr 7, NC_000007.14:g.100616392 | 0.2273 | 0.4259 | 0.34 | 0.26 | 3.8 × 10−2 | 2.52 | |
| rs4729597 (T/C) | Intergenic, chr 7, NC_000007.14:g.100624226 | 0.2727 | 0.5000 | 0.40 | 0.37 | 2.2 × 10−2 | 2.67 | |
| rs117406702 (G/A) | Intergenic, chr 7, NC_000007.14:g.100748471 | 0 | 0.0926 | 0.05 | 0.03 | 3.8 × 10−2 | NA |
| Locus | SNP | Location, HGVS Name | F_R | F_NR | MAF R + NR | MAF EUR | p Value | OR |
|---|---|---|---|---|---|---|---|---|
| ANRIL | rs1063192 (A/G) | 3′ UTR, chr9, NM_004936.4:c.*2619 | 0.568 | 0.315 | 0.43 | 0.43 | 1.2 × 10−2 | 0.35 |
| ANRIL | rs10120688 (A/G) | Intron, chr9, NC_000009.12:g.22056500 | 0.636 | 0.407 | 0.51 | 0.50 | 2.4 × 10−2 | 0.39 |
| LINC00941 | rs12297445 (G/A) | Intron, chr12, NC_000012.12:g.30789344 | 0 | 0.093 | 0.05 | 0.09 | 3.8 × 10−2 | NA |
| miR4706 | rs2296316 (T/C) | Intron, chr14, NC_000014.9:g.65053528 | 0.591 | 0.333 | 0.45 | 0.46 | 1.1 × 10−2 | 0.35 |
| Protein (Gene) Name | SNP | Location, HGVS Name | F_R | F_NR | MAF R + NR | MAF EUR | p Value | OR |
|---|---|---|---|---|---|---|---|---|
| AKAP6 (AKAP6) | rs2031106 (G/A) | Intron, chr 14, NC_000014.9:g.32392503 | 0.182 | 0.056 | 0.11 | 0.12 | 4.9 × 10−2 | 0.26 |
| rs7142538 (A/G) | Intron, chr 14, NC_000014.9:g.32404235 | 0.114 | 0 | 0.05 | 0.08 | 1.1 × 10−2 | 0 | |
| rs3742926 (C/T) | Missense, chr 14, NM_004274.5:c.1010 | 0.068 | 0.222 | 0.15 | 0.10 | 3.5 × 10−2 | 3.91 | |
| rs7150894 (A/G) | Synonymous, chr 14, NM_004274.5:c.1155 | 0.523 | 0.296 | 0.40 | 0.34 | 2.3 × 10−2 | 0.38 | |
| rs1956204 (C/T) | Intron, chr 14, NC_000014.9:g.32549256 | 0.136 | 0.315 | 0.23 | 0.20 | 3.8 × 10−2 | 2.91 | |
| rs11624518 (G/A) | Intron, chr 14, NC_000014.9:g.32711145 | 0.205 | 0.389 | 0.31 | 0.29 | 4.9 × 10−2 | 2.48 | |
| rs11622943 (A/G) | Intron, chr 14, NC_000014.9:g.32776891 | 0.159 | 0.352 | 0.27 | 0.39 | 3.2 × 10−2 | 2.87 | |
| rs34628134 (A/G) | Intron, chr 14, NC_000014.9:g.32778457 | 0.455 | 0.241 | 0.34 | 0.23 | 2.6 × 10−2 | 0.38 | |
| A4 (APP) | rs380417 (C/T) | Intron, chr 21, NC_000021.9:g.25899847 | 0.023 | 0.185 | 0.11 | 0.22 | 1.1 × 10−2 | 9.77 |
| rs2829981 (A/G) | Intron, chr 21, NC_000021.9:g.25908384 | 0 | 0.093 | 0.05 | 0.09 | 3.8 × 10−2 | NA | |
| rs1783024 (C/T) | Intron, chr 21, NC_000021.9:g.25966822 | 0.614 | 0.407 | 0.50 | 0.42 | 4.2 × 10−2 | 0.43 | |
| rs128648 (C/T) | Intron, chr 21, NC_000021.9:g.25970679 | 0.136 | 0.315 | 0.23 | 0.28 | 3.8 × 10−2 | 2.91 | |
| rs58908134 (G/T) | Intron, chr 21, NC_000021.9:g.25989938 | 0.068 | 0.222 | 0.15 | 0.18 | 3.5 × 10−2 | 3.91 | |
| SHAN2 (SHANK2) | rs10899236 (A/G) | Intron, chr 11, NC_000011.10:g.70606029 | 0.614 | 0.407 | 0.50 | 0.45 | 4.2 × 10−2 | 0.43 |
| rs12274337 (C/T) | Intron, chr 11, NC_000011.10:g.70643910 | 0 | 0.093 | 0.05 | 0.03 | 3.8 × 10−2 | NA | |
| rs35961474 (C/T) | Intron, chr 11, NC_000011.10:g.70901955 | 0.136 | 0 | 0.06 | 0.05 | 5.1 × 10−3 | 0 | |
| ITSN1 (ITSN1) | rs2251854 (A/G) | Intron, chr 21, NC_000021.9:g.33683942 | 0.205 | 0.426 | 0.33 | 0.32 | 2 × 10−2 | 2.89 |
| rs2256797 (T/C) | Intron, chr 21, NC_000021.9:g.33785207 | 0.114 | 0.333 | 0.23 | 0.23 | 1.1 × 10−2 | 3.90 | |
| rs2834269 (T/C) | Intron, chr 21, NC_000021.9:g.33828070 | 0.455 | 0.241 | 0.34 | 0.25 | 2.6 × 10−2 | 0.38 | |
| rs2834287 (A/G) | 3′ UTR, chr 21, NM_003024.3:c.*357 | 0.091 | 0.315 | 0.21 | 0.17 | 7.2 × 10−3 | 4.60 | |
| P85A (PIK3R1) | rs831122 (G/A) | Intron, chr 5, NC_000005.10:g.68267102 | 0.023 | 0.185 | 0.11 | 0.15 | 1.1 × 10−2 | 9.77 |
| AKA12 (AKAP12) | rs10499266 (G/A) | Intron, chr 6, NC_000006.12:g.151251388 | 0.136 | 0.019 | 0.07 | 0.08 | 2.4 × 10−2 | 0.12 |
| rs73613423 (T/G) | Intron, chr 6, NC_000006.12:g.151263516 | 0.114 | 0 | 0.05 | 0.05 | 1.1 × 10−2 | 0 | |
| rs9397389 (C/T) | Intron, chr 6, NC_000006.12:g.151308730 | 0.205 | 0.389 | 0.31 | 0.26 | 4.9 × 10−2 | 2.48 | |
| MEOX2 (MEOX2) | rs10270030 (T/C) | Intron, chr 7, NC_000007.14:g.15648375 | 0.318 | 0.148 | 0.22 | 0.35 | 4.5 × 10−2 | 0.37 |
| ABL1 (ABL1) | rs75764711 (G/A) | Intron, chr 9, NC_000009.12:g.130794660 | 0 | 0.111 | 0.06 | 0.02 | 2.2 × 10−2 | NA |
| ARRB1 (ARRB1) | rs512797 (G/A) | Intron, chr 11, NC_000011.10:g.75285037 | 0 | 0.167 | 0.09 | 0.16 | 4.5 × 10−3 | NA |
| rs616714 (G/T) | Intron, chr 11, NC_000011.10:g.75333596G | 0.068 | 0.259 | 0.17 | 0.18 | 1.3 × 10−2 | 4.78 | |
| NKG2D (KLRK1) | rs2617151 (G/A) | Intron, chr 12, NC_000012.12:g.10379835 | 0.136 | 0.315 | 0.23 | 0.18 | 3.8 × 10−2 | 2.91 |
| ZBTB1 (ZBTB1) | rs74056445 (A/G) | Intron, chr 14, NC_000014.9:g.64511474 | 0.023 | 0.148 | 0.09 | 0.09 | 3.2 × 10−2 | 7.48 |
| CREBBP (CBP) | rs2239316 * (A/G) | Intron, chr16, NC_000016.10:g.3863994 | 0.091 | 0.259 | 0.18 | 0.25 | 3.2 × 10−2 | 3.5 |
| ATF1 (ATF1) | rs1129406 (C/T) | Synonymous, chr 12, NM_005171.5:c.327 | 0.409 | 0.222 | 0.31 | 0.39 | 4.6 × 10−2 | 0.41 |
| IκBα (NFKBIA) | rs696 * (G/A) | 3′ UTR, chr 14, NM_020529.3:c.*126 | 0.568 | 0.352 | 0.45 | 0.39 | 3.2 × 10−2 | 0.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liadaki, K.; Zafiriou, E.; Giannoulis, T.; Alexouda, S.; Chaidaki, K.; Gidarokosta, P.; Roussaki-Schulze, A.-V.; Tsiogkas, S.G.; Daponte, A.; Mamuris, Z.; et al. PDE4 Gene Family Variants Are Associated with Response to Apremilast Treatment in Psoriasis. Genes 2024, 15, 369. https://doi.org/10.3390/genes15030369
Liadaki K, Zafiriou E, Giannoulis T, Alexouda S, Chaidaki K, Gidarokosta P, Roussaki-Schulze A-V, Tsiogkas SG, Daponte A, Mamuris Z, et al. PDE4 Gene Family Variants Are Associated with Response to Apremilast Treatment in Psoriasis. Genes. 2024; 15(3):369. https://doi.org/10.3390/genes15030369
Chicago/Turabian StyleLiadaki, Kalliopi, Efterpi Zafiriou, Themistoklis Giannoulis, Sofia Alexouda, Kleoniki Chaidaki, Polyxeni Gidarokosta, Angeliki-Viktoria Roussaki-Schulze, Sotirios G. Tsiogkas, Athina Daponte, Zissis Mamuris, and et al. 2024. "PDE4 Gene Family Variants Are Associated with Response to Apremilast Treatment in Psoriasis" Genes 15, no. 3: 369. https://doi.org/10.3390/genes15030369
APA StyleLiadaki, K., Zafiriou, E., Giannoulis, T., Alexouda, S., Chaidaki, K., Gidarokosta, P., Roussaki-Schulze, A.-V., Tsiogkas, S. G., Daponte, A., Mamuris, Z., Bogdanos, D. P., Moschonas, N. K., & Sarafidou, T. (2024). PDE4 Gene Family Variants Are Associated with Response to Apremilast Treatment in Psoriasis. Genes, 15(3), 369. https://doi.org/10.3390/genes15030369